Introduction
JC virus (JCV), a member of the polyomavirus genus,
which also includes SV40 and BK viruses, is a non-enveloped,
5,130-bp double-stranded DNA virus that forms minichromosomes with
cellular histones (1). The JCV
early region encodes 2 oncoproteins, large T-antigen (T-Ag) and
small t antigen (t-Ag). The late region encodes 3 capsid proteins,
VP1, VP2 and VP3, and a small regulatory protein, agnoprotein
(2). The early viral proteins have
high transformation and oncogenic potential in experimental systems
(3). There are 2 possible outcomes
of JCV infection. One possible outcome is the support of viral DNA
replication by permissive cells, such as oligodendrocytes,
resulting in a lytic infection. Another possibility is silent or
abortive infection that may be linked to cell transformation in
nonpermissive cells, such as those of the pulmonary and digestive
epithelium (4).
As JCV has been frequently detected in tumors of the
human central nervous system (CNS) (5), it has been suggested that this virus
may have a possible association with a variety of human CNS tumors.
However, the mechanisms involved in JCV-associated oncogenesis must
be clarified since studies provide contradictory evidence as to
whether or not JCV plays a causative role in human CNS tumors
(6). Laghi et al(7), followed by Enam et al(8), were the first to report the presence
of JCV DNA in colorectal cancers. We evaluated the presence of JCV
DNA in Japanese patients with colorectal (9), gastric (10), oral (11) and lung cancer (12). While JCV DNA was detected in cancers
of these organs, we could not demonstrate an association between
the presence of JCV DNA and its oncogenic role.
Approximately 4 decades have passed since the first
report that JCV is found in brain tumors following inoculation of
this virus into the brain of newborn hamsters (13). Subsequently, several studies have
shown that brain tumors arise in JCV transgenic mouse models
(14,15). It is well-known that the JCV
receptor is the 5-HT2A serotonin receptor, which is primarily
restricted to glial cells (16).
Therefore, it may be reasonable to conclude that JCV has a strong
tendency to form neurotropic tumors (6). However, since JCV DNA has been shown
to be present in pulmonary and digestive organs, we speculate that
JCV may infect these organs through an unknown receptor. We created
transgenic (TG) mice with a transgene including the K-19 promoter,
which is specific to bronchial and digestive epithelium (17) and the JCV T-antigen. We found
pulmonary tumors in 2/15 mice (13.3%) with no tumors in any other
organs.
Materials and methods
Establishment and generation of K-19/JCV
T-antigen transgenic mice
TG mice were generated using a 3.2-kb
BalI/NciI restriction fragment from the plasmid pBJC,
containing the K-19 promoter (a kind gift from Professor H. Oshima,
Kanazawa University Cancer Research Institute) and the coding
region for the viral early genes, large T- and small t-antigens.
The DNA was microinjected into the pronucleus of fertilized mouse
oocytes generated by the mating of C57BL/6J mice (Chrysalis DNX,
Princeton, NJ, USA). Founder animals were mated with C57BL/6J mice,
and all mice were screened for the presence of the transgene by DNA
extraction from the tail tissue and real-time PCR analysis with
primers specific to the transgene. Real-time quantitative PCR was
performed using the SYBR® Premix DimerEraser (Takara
Bio, Inc., Ohtsu, Japan) kit and LightCycler® (Roche
Applied Science, Indianapolis, IN, USA). The primer set
(5′-TGCCACTGTCTATTGGCCCCT-3′ and 5′-TTGGGGCACATGGCAATGCTGT-3′) was
designed to detect JCV T-antigen DNA (amplicon size 168 bp), and
reaction mixtures were prepared according to the manufacturer’s
protocol. After amplification, using hemizygous and wild-type mice
(C57BL/6J), we generated standard curves. The homozygous,
hemizygous and non-TG states in the target DNA samples were clearly
distinguished by the difference in these Ct values.
Potentially homozygous animals were validated by test breeding with
wild-type partners. Pure JCV T-antigen+/+ mice were
achieved after 3 generations.
In the present study, 15 TG mice (8 males and 7
females) and 10 control mice (5 males and 5 females) were
euthanized by CO2 inhalation at 16 months of age and
were autopsied systemically.
Histologic and immunohistochemical (IHC)
analyses of TG mice
Tissues from the lung, heart, liver, pancreas,
spleen, kidney, esophagus, stomach, small intestine, large
intestine and brain were obtained from TG and C57BL/6J control mice
and fixed in 10% formalin. Histological examination was carried out
by 3 pathologists (A.N., Y.M. and Y.T.).
The IHC procedure was similar to that described
previously (12). Primary
antibodies were directed against JCV T-antigen (1:100 dilution,
PAb416, mouse mAb; EMD Millipore, Darmstadt, Germany), p53 (1:50,
PAb122, mouse mAb; Enzo Life Science, Farmingdale, NY, USA),
β-catenin (1:1,000, rabbit pAb; Cell Signaling Technology, Inc.,
Beverly, MA, USA), IRS-1 (1:50, sc-8038, mouse mAb; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA), cytokeratin CK-19
(1:100, rabbit mAb; Epitomics, Burlingame, CA, USA), CK-7 (1:100,
KRT7, rabbit pAb; Proteintech Group, Chicago, IL, USA), CK-20
(1:50, mouse mAb; GeneTex, San Antonio, TX, USA), CD3 (1:100, goat
pAb, Santa Cruz Biotechnology, Inc.), CD4 (1:50, mouse mAb; EMD
Millipore), CD5 (1:50, mouse mAb; Abnova, Taipei, Taiwan), CD8
(1:50, rabbit mAb; GeneTex), CD20 (1:50, rabbit mAb; EMD
Millipore), CD21 (1:50, mouse mAb; Santa Cruz Biotechnology, Inc.)
and CD79a (1:100, rabbit pAb; LifeSpan Biosciences, Seattle, WA,
USA).
Mutation analysis of K-ras and epidermal
growth factor receptor
DNA samples from tumor lesions and bronchial
epithelium were selectively captured from 10 μm dewaxed sections
using a laser capture microdissection (LCM) system (LM200, Olympus,
Tokyo). Isolation of DNA from LCM specimens was performed with a
PicoPure® DNA Extraction kit (Arcturus Engineering,
Mountain View, CA, USA) according to the manufacturer’s protocol.
The primers used for detection of codon 12 mutations were described
elsewhere (18). The primers used
for detection of epidermal growth factor receptor (EGFR) exon 18–21
mutations were 18S:2349–2369, 5′-TCGTGGAACCTCTCACACCCA-3′ and
18A:2467–2446, 5′-ATACACTGTGCCAAATGCTCCC-3′; 19S:2473–2483,
5′-TCTCTGGATCCCAGAAGGTGA-3′ and 19A:2569–2549,
5′-GTCAAGGATTTCTTTGTTGGC; 20S:2571–2591, 5′-AGCCTATGTGATGGCTAGTG-3′
and 20A:2750–2731, 5′-CAATCTGCACACACCAGTTG-3′; 21S:2757–2778,
5′-GCATGAACTACCTGGAAGATCG-3′ and 21A:2904–2882,
5′-CCCTCGGCATGATATTCTTTCTC-3′. These primers are equipped with the
M13-primer sequence to facilitate sequencing. PCR was performed
according to the manufacturer’s protocol. The PCR products were
purified using the BigDye XTerminator® Purification kit
(Applied Biosystems) to remove unconsumed dNTPs and primers, and
1.5 μl aliquots were then directly sequenced using the
M13-sequencing primers and BigDye Terminator® v3.1 cycle
sequencing kit in an ABI 3130 genetic analyzer (Applied
Biosystems).
Results
Observational findings
The observational finding between TG mice and
control mice included epilation. All of the 15 TG mice sporadically
lost ~50% of their body hair, while 8 control mice lost no hair and
2 control mice had minimally sporadic hair loss. There was no
statistical difference in body weight between the TG (26.7±1.10 g)
and control mice (27.0±1.05 g). Confirmation of K19/JCV
T-antigen+/+ TG mice was accomplished using real-time
PCR targeting for the JCV T-antigen just before the mice were
euthanized.
Histologic and IHC findings
Two pulmonary tumors were identified in the TG mice
(13.3% of TG mice), while no tumors were found in any other organs
examined including the esophagus, stomach, small intestine, large
intestine, heart, liver, pancreas, kidney, spleen and brain. One
pulmonary tumor occupied over half of the left lung without
pulmonary metastasis in a male TG mouse. Histologically, there was
a clear boundary between the tumor and normal lung tissue and a
small tubular, solid arrangement consisting of small irregular and
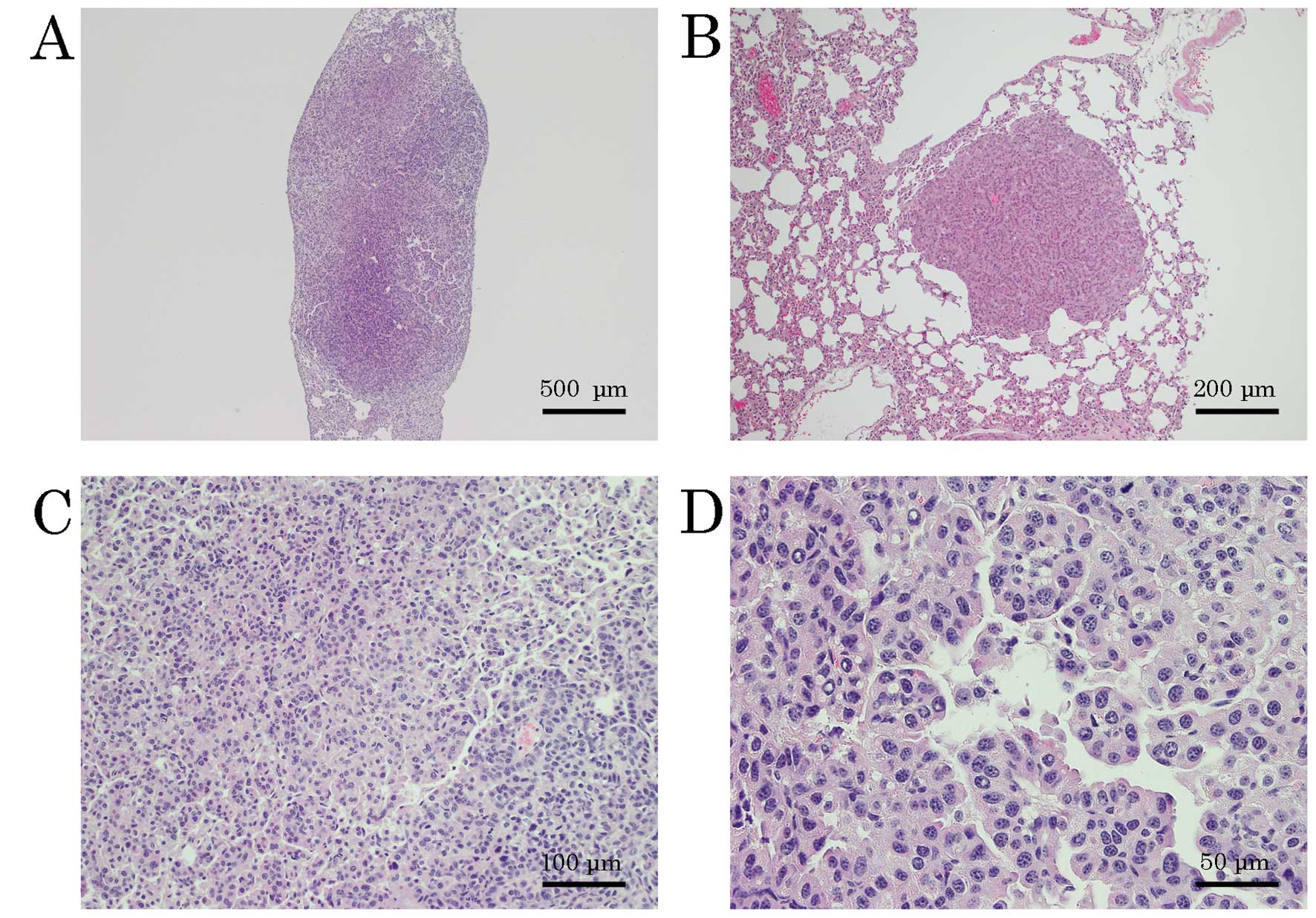
atypical nuclear cells (Fig. 1A and
C). Some of the tumor cells morphologically resembled
hyperplastic bronchial cells (Fig.
1D). Another pulmonary small tumor without pulmonary metastasis
was found in the left lung of a female TG mouse. The histologic
examination of this tumor showed a clear boundary between the tumor
and normal lung tissue and a small, tubular arrangement consisting
of round- to oval-shaped and mildly atypical nuclear cells
(Fig. 1B). Bronchial hyperplasia
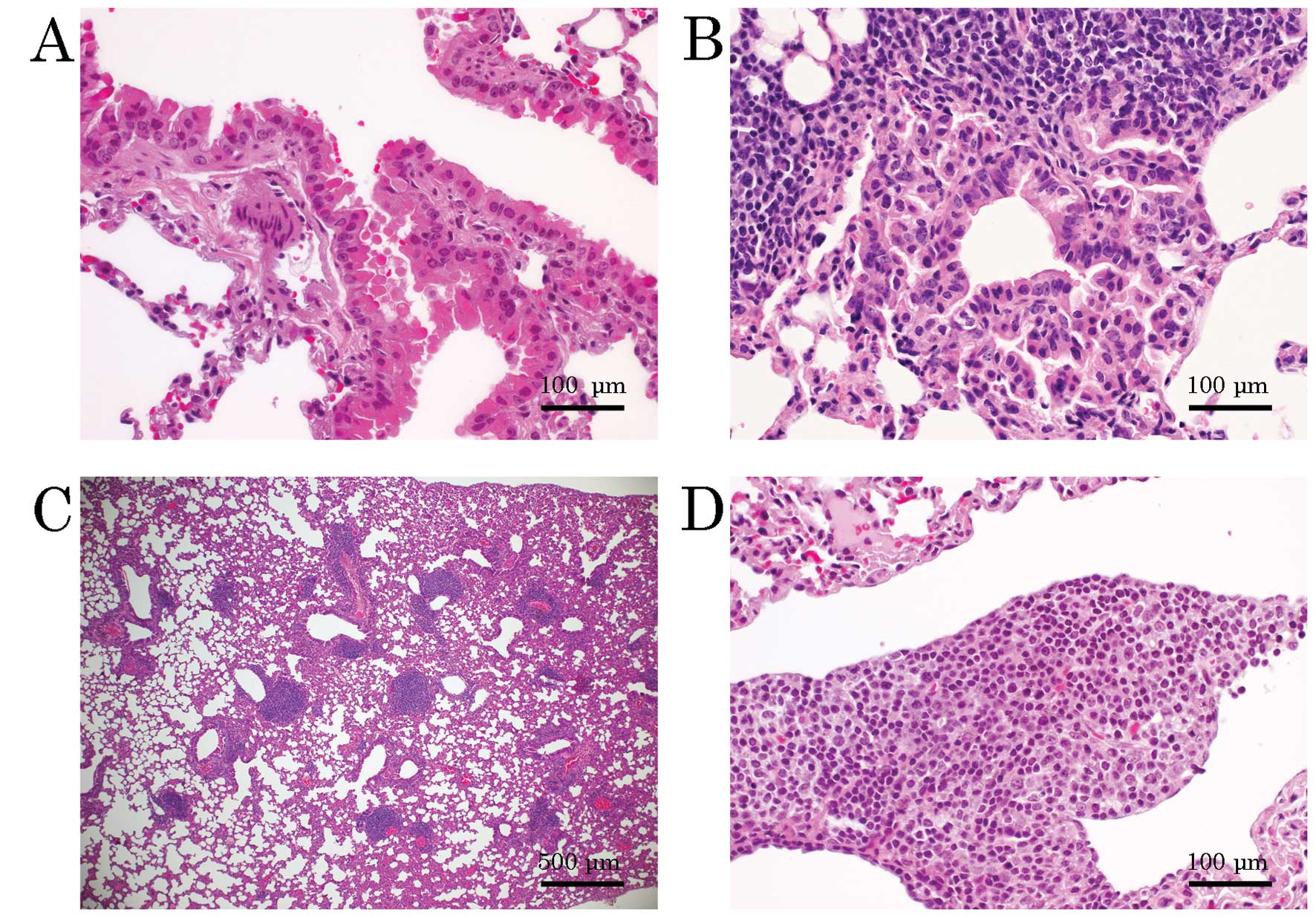
was more prominent in TG mice than in control mice (Fig. 2A). Some bronchial hyperplasia
morphologically resembled a pulmonary tumor, but was characterized
as bronchial hyperplasia due to the small size and mild atypia
(Fig. 2B).
There was a clear tendency toward sporadic
aggregation of lymphocytes in the TG mice (Fig. 2C), some of which showed monotone
features and mild atypia (Fig. 2D).
This finding was not observed in the control mice.
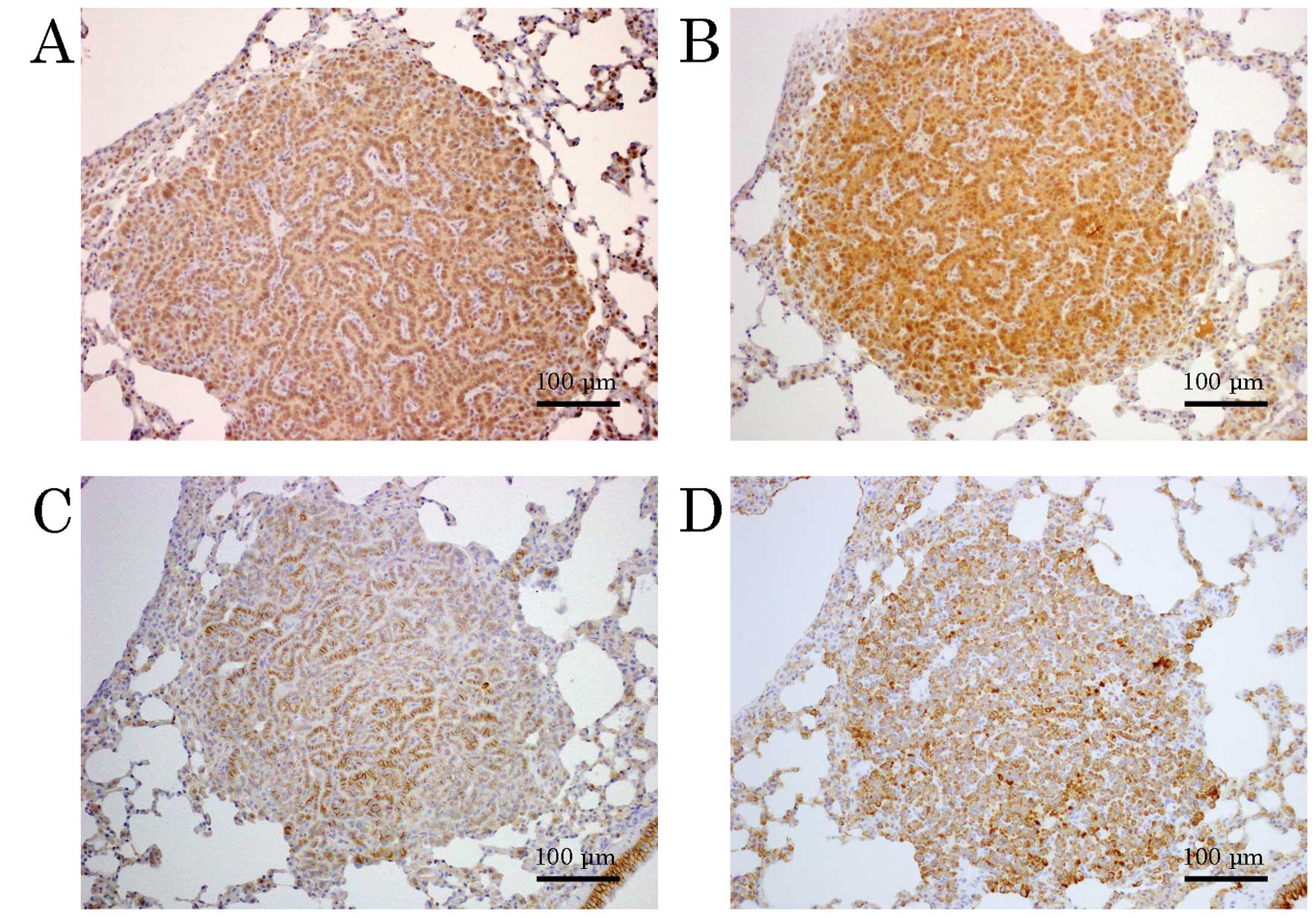
IHC revealed that the pulmonary tumors were positive
for JCV T-antigen primarily in the nuclei (Fig. 3A). p53 was also noted primarily in
the nuclei (Fig. 3B), with
membranous β-catenin (Fig. 3C) and
cytoplasmic CK-19 (Fig. 3D).
Insulin receptor substrate-1 (IRS-1), used as an alternate for
insulin-like growth factor receptor (IGFR), was negative as
determined by IHC. Tumor cells were CK7(+) and CK20(−). Similar IHC
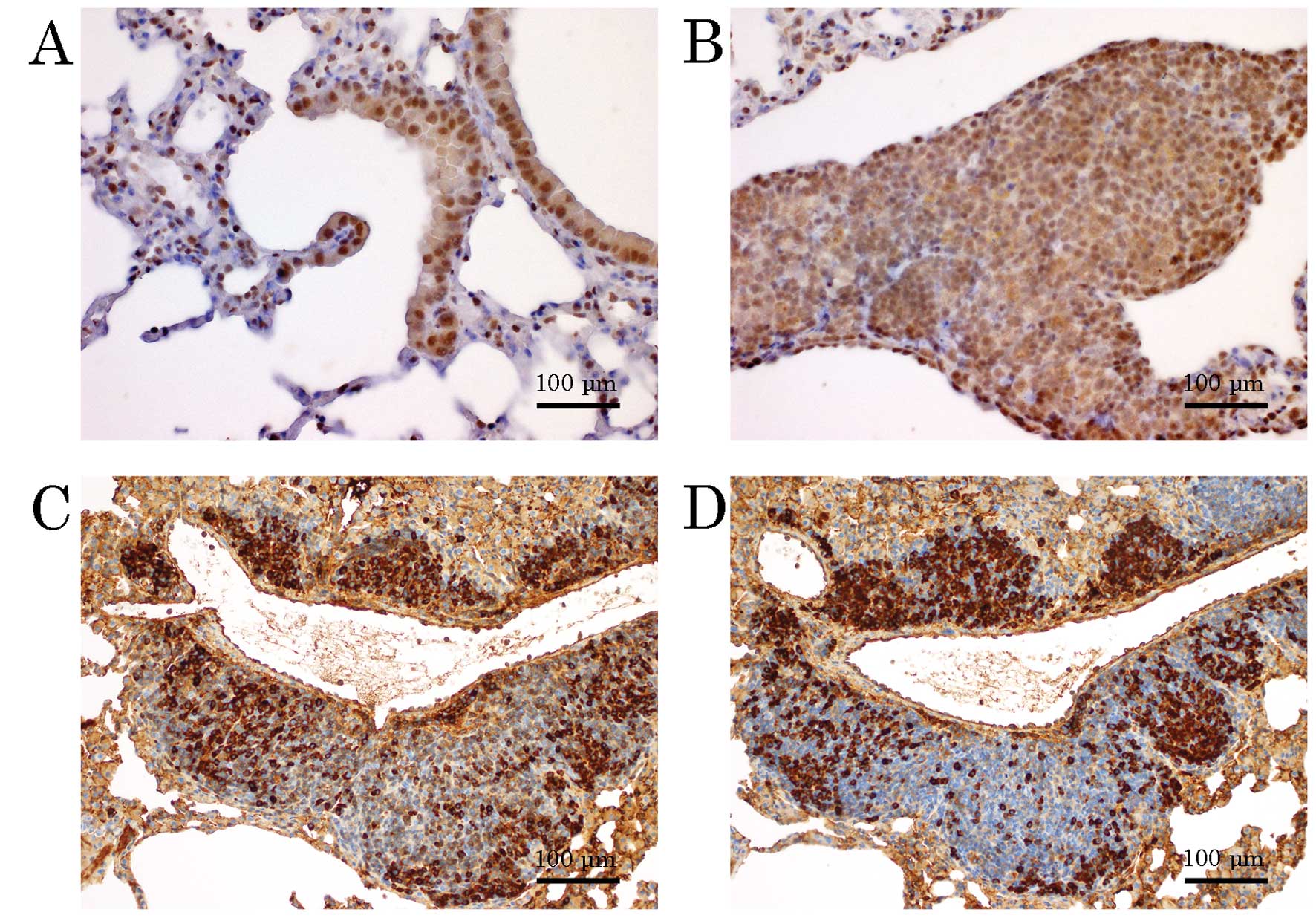
findings were noted in the bronchial epithelium. As shown in
Fig. 4A, positivity for JCV
T-antigen was noted in the bronchial epithelium.
Scattered aggregations of lymphocytes as shown in
Fig. 2C were usually positive for
the JCV T-antigen (Fig. 4B) and
p53. However, lymphocytes demonstrated polyclonal
immunohistochemical reactivity. Some lymphocytes represented T
lymphocyte clones, and were positive for CD3 and CD5, as well as
balanced CD4 and CD8 positivity. Other lymphocytes represented B
lymphocyte clones, and were positive for CD20 and CD79a and, to a
lesser degree, CD10. Fig. 4C
illustrates CD3(+) lymphocytes and Fig.
4D illustrates CD79a(+) lymphocytes.
Mutation analyses of K-ras and EGFR
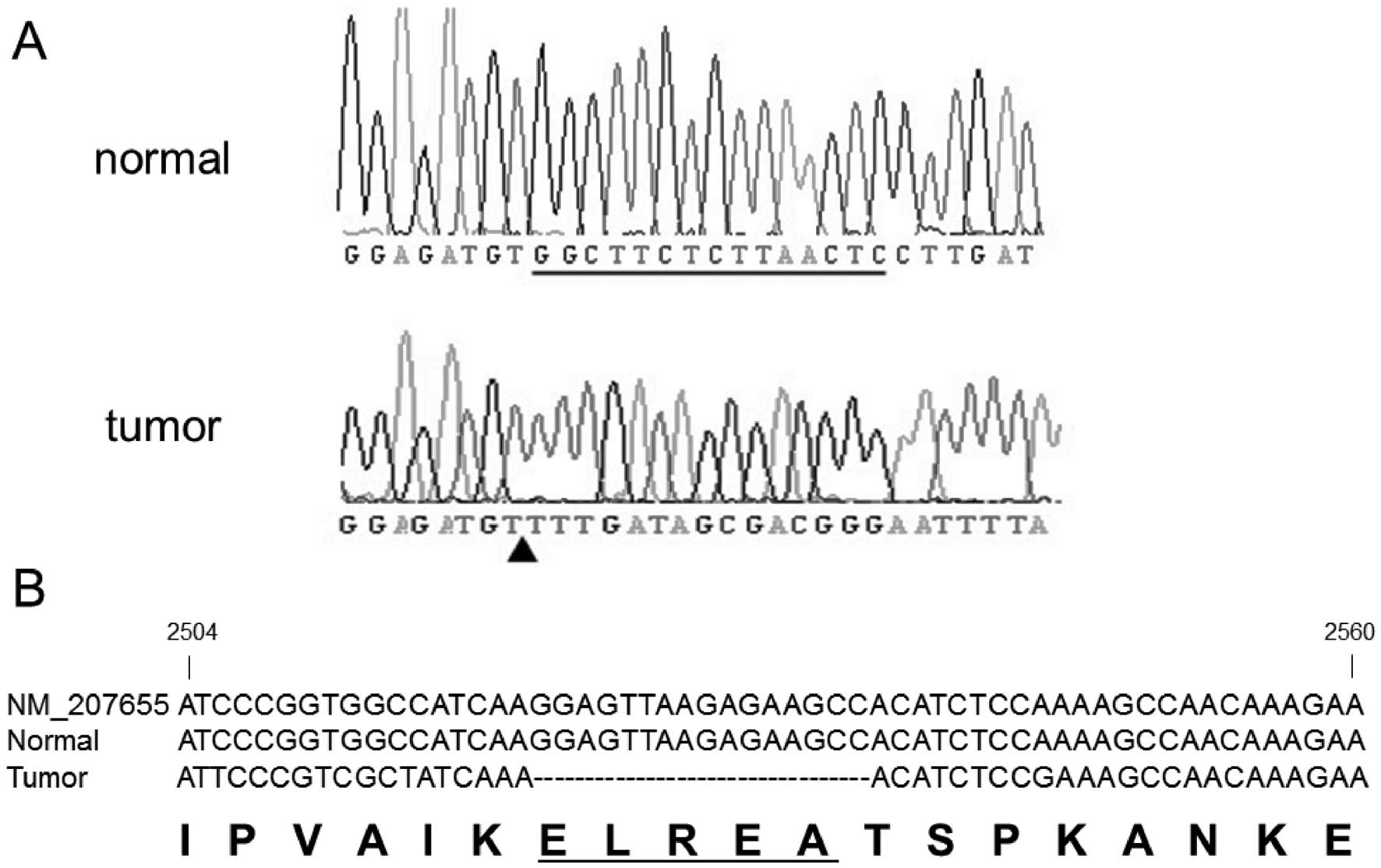
No mutations in K-ras were observed. An
EGFR mutation was detected in the in-frame exon 19 deletion
that eliminates the 5 amino acids
glucine-leucine-arginine-glucine-alanine ELREA (del E841-A845). No
abnormalities were detected in the corresponding normal benign lung
tissue (Fig. 5).
Discussion
This is the first report showing that JCV may be
associated with epithelial tumorigenesis in experimental animals.
The possibility that spontaneous pulmonary tumors should arise in
TG mice is very low in this case, as the back strain of this TG
mouse, C57BL/6J, is almost completely resistant to spontaneous
pulmonary tumors (19). We did not
detect any tumors in other organs; most notably no tumors were
detected in the brain. We also did not detect any intra-pulmonary
or distant metastasis.
Conventional mouse models for pulmonary cancer
almost exclusively give rise to solid, papillary or
bronchiolo-alveolar adenocarcinomas. These single or mixed tumor
types retain marker expression profiles characteristic for both
type II alveolar as well as Clara cells, and therefore both cell
types can serve as hypothetical cells of origin for mouse non-small
cell lung cancer (19). Similar
experiments demonstrated that mice expressing SV40 large T-antigen
from the Clara cell-specific CC10 promoter or alveolar type II
cell-specific promoter developed early multifocal bronchial
alveolar hyperplasia followed by mixed solid and papillary
adenocarcinomas to which the mice succumbed by 4–5 months of age
(20–22). Ectopic expression of the HPV-16
E6/E7 transgene under the control of the keratine-5 promoter
resulted in lung adenocarcinoma after ~6 months of age (23).
The JCV T-antigen can trigger progression of the
cell cycle into the S phase in host cells (3,24) and
interacts with p53, resulting in interference with p53 (25) and allowing additional genetic
damage, representing a step forward in tumorigenesis (26,27).
In the present study, p53 was expressed exclusively in pulmonary
tumors and bronchial epithelium. Although we could not determine
whether this p53 was wild-type or the mutated type, we hypothesized
that p53 may be linked to pulmonary tumorigenesis induced by the
JCV T-antigen. The JCV T-antigen can also interact with IRS-1, a
major protein of the insulin-like growth factor I receptor (IGF-IR)
signaling pathway, which is activated and translocated to the
nucleus in the presence of the JCV T antigen (28). Activated IRS-1 is an adaptor in the
cell response to insulin, activating phosphatidylinositol 3-kinase
(PI3K), which is implicated in cell survival (29) and proliferation signals (30). The JCV T-antigen can also inhibit
homologous recombination-directed DNA repair (HRR) causing DNA
damage, mechanistically by its interaction with IRS-1 (31), which also interacts with Rad51 at
locations of damaged DNA (32), and
thus may contribute to the generation of genetic instability in
cells containing JCV (33). In this
study, expression of IRS-1 was not found in any type of cells, and
therefore we could not find evidence of this mechanism.
The JCV T-antigen also contributes to the
stabilization of β-catenin by a novel mechanism mediated by the
small GTPase Rac1 (34). β-catenin
is an integral component of the Wnt signaling pathway, and
stabilization of β-catenin is associated with increased
transcription of genes that regulate cellular proliferation, e.g.,
c-myc and cyclin D1 (8). In our
study, expression of β-catenin was restricted to the membrane of
tumors and bronchial epithelial cells, but was not expressed in the
cytoplasm or nucleus, which led us to hypothesize that expression
of β-catenin merely played the role of an adhesion molecule in
these cells.
Another mechanism by which JCV may contribute to
pulmonary tumorigenesis is similar in some aspects to the role of
human papilloma virus (HPV) in cervical carcinoma (35). Viral DNA integration has been only
partially addressed in cervical cancer associated with HPV
(36). The chromosomal locations of
HPV viral integration sites coincide with those of fragile
chromosome sites (37) and
translocation break-points which have been already detected in
other types of carcinomas (38). If
the JCV infection is persistent, then there is a high probability
that JCV DNA may integrate into the host cell genome, while some of
the viral DNA remains similarly to what occurs with HPV in cervical
carcinoma (37).
In mouse models of chemical pulmonary tumorigenesis,
in which the type of tumor is adenoma or adenocarcinoma,
K-ras mutations are frequently observed, whereas genetic
alteration of EGFR is generally rare (38). In the present study, only one case
of bronchial epithelial hyperplasia was examined. In this case, an
EGFR mutation was detected, while a K-ras mutation
was not detected. While it is unclear how mutations of EGFR
and other molecular alterations are related to the mechanisms
involved in pulmonary tumorigenesis in mice, both genetic and
epigenetic factors are known to be important for tumorigenesis
(39). In this case, it is
impossible to resolve the mechanism of the phenomenon based on only
one tumor.
In this study, we found that some lymphocytes were
positive for both the JCV T-antigen and p53 in the nuclei. However,
these lymphocytes were not found to be monoclonal by IHC. The
hypothesis suggesting that lymphocytes mediate JCV infection was
reported in a previous study (40).
Lymphocytes may have tolerance for JCV activation, leading to
positivity for both JCV T-antigen and p53.
Acknowledgements
This study was partially supported by a Grant-in-Aid
for Scientific Research (nos. 221S0001 and 23659958) from the
Ministry of Education, Science, Sports and Culture, Japan.
References
|
1
|
Coelho TR, Almeida L and Lazo PA: JC virus
in the pathogenesis of colorectal cancer, an etiological agent or
another component in a multistep process. Virol J. 7:422010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jiang M, Abend JR, Johnson SF and
Imperiale MJ: The role of polyomaviruses in human disease. Virol.
384:266–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Khalili K, Sariyer IK and Safak M: Small
tumor antigen of polyomaviruses: role in viral life cycle and cell
transformation. J Cell Physiol. 215:309–319. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Haggerty S, Walker DL and Frisque RJ: JC
virus-simian virus 40 genomes containing heterologous regulatory
signals and chimeric early regions: identification of regions
restricting transformation by JC virus. J Virol. 63:2180–2190.
1989.
|
|
5
|
Khalili K, Del Valle L, Otte J, et al:
Human neurotropic polyomavirus, JCV, and its role in
carcinogenesis. Oncogene. 22:5181–5191. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Del Valle L, White MK and Khalili K:
Potential mechanisms of the human polyomavirus JC in neural
oncogenesis. J Neuropathol Exp Neurol. 67:729–740. 2008.PubMed/NCBI
|
|
7
|
Laghi L, Randolph AE, Chauhan DP, et al:
JC virus DNA is present in the mucosa of the human colon and in
colorectal cancers. Proc Natl Acad Sci USA. 96:7484–7489. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Enam S, Del Valle L, Lara C, et al:
Association of human polyomavirus JCV with colon cancer: evidence
for interaction of viral T-antigen and β-catenin. Cancer Res.
62:7093–7101. 2002.
|
|
9
|
Hori R, Murai Y, Tsuneyama K, et al:
Detection of JC virus DNA sequences in colorectal cancers in Japan.
Virchows Arch. 447:723–730. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Murai Y, Zheng HC, Abdel Aziz HO, et al:
High JC virus load in gastric cancer and adjacent non-cancerous
mucosa. Cancer Sci. 98:25–31. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kutsuna T, Zheng H, Abdel-Aziz HO, et al:
High JC virus load in tongue carcinomas may be a risk factor for
tongue tumorigenesis. Virchows Arch. 452:405–410. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zheng H, Abdel Aziz HO, Nakanishi Y, et
al: Oncogenic role of JC virus in lung cancer. J Pathol.
212:306–315. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Walker DL, Padgett BL, ZuRhein GM, et al:
Human papovavirus (JC): induction of brain tumors in hamsters.
Science. 181:674–676. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Krynska B, Otte J, Franks R, et al: Human
ubiquitous JCV(CY) T-antigen gene induces brain tumors in
experimental animals. Oncogene. 18:39–46. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shollar D, Del Valle L, Khalili K, et al:
JCV T-antigen interacts with the neurofibromatosis type 2 gene
product in a transgenic mouse model of malignant peripheral nerve
sheath tumors. Oncogene. 23:5459–5467. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Elphick GF, Querbes W, Jordan JA, et al:
The human polyomavirus, JCV, uses serotonin receptors to infect
cells. Science. 306:1380–1383. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Oshima H, Matsunaga A, Fujimura T, et al:
Carcinogenesis in mouse stomach by simultaneous activation of the
Wnt signaling and prostaglandin E2 pathway. Gastroenterology.
131:1086–1095. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Karasaki H, Obata M, Ogawa K and Lee GH:
Roles of the Pas1 and Par2 genes in determination of
the unique, intermediate susceptibility of BALB/cByJ mice to
urethane-induction of lung carcinogenesis: differential effects on
tumor multiplicity, size and Kras2 mutations. Oncogene.
15:1833–1840. 1997.
|
|
19
|
Meuwissen R and Berns A: Mouse models for
human lung cancer. Genes Dev. 19:643–664. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
DeMayo FJ, Finegold MJ, Hansen TN, et al:
Expression of SV40 T antigen under control of rabbit uteroglobin
promoter in transgenic mice. Am J Physiol. 261:L70–L76.
1991.PubMed/NCBI
|
|
21
|
Wikenheiser KA, Clark JC, Linnoila RI, et
al: Simian virus 40 large T antigen directed by transcriptional
elements of the human surfactant protein C gene produces pulmonary
adenocarcinomas in transgenic mice. Cancer Res. 52:5342–5352.
1992.
|
|
22
|
Sandmöller A, Halter R, Gómez-La-Hoz E, et
al: The uteroglobin promoter targets expression of the SV40 T
antigen to a variety of secretory epithelial cells in transgenic
mice. Oncogene. 9:2805–2815. 1994.PubMed/NCBI
|
|
23
|
Carraresi L, Tripodi SA, Mulder LC, et al:
Thymic hyperplasia and lung carcinomas in a line of mice transgenic
for keratin 5-driven HPV16 E6/E7 oncogenes. Oncogene. 20:8148–8153.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Krynska B, Gordon J, Otte J, et al: Role
of cell cycle regulators in tumor formation in transgenic mice
expressing the human neurotropic virus, JCV, early protein. J Cell
Biochem. 67:223–230. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Staib C, Pesch J, Gerwig R, et al: p53
inhibits JC virus DNA replication in vivo and interacts with JC
virus large T-antigen. Virology. 219:237–246. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Niv Y, Goel A and Boland CR: JC virus and
colorectal cancer: a possible trigger in the chromosomal
instability pathways. Curr Opin Gastroenterol. 21:85–89.
2005.PubMed/NCBI
|
|
27
|
Nosho K, Shima K, Kure S, et al: JC virus
T-antigen in colorectal cancer is associated with p53 expression
and chromosomal instability, independent of CpG island methylator
phenotype. Neoplasia. 11:87–95. 2009.PubMed/NCBI
|
|
28
|
Khalili K, Del Valle L, Wang JY, et al:
T-antigen of human polyomavirus JC cooperates with IGF-IR signaling
system in cerebellar tumors of the childhood-medulloblastomas.
Anticancer Res. 23:2035–2041. 2003.PubMed/NCBI
|
|
29
|
del Rincón SV, Rousseau C, Samanta R and
Miller WH Jr: Retinoic acid-induced growth arrest of MCF-7 cells
involves the selective regulation of the IRS-1/PI 3-kinase/AKT
pathway. Oncogene. 22:3353–3360. 2003.PubMed/NCBI
|
|
30
|
Dearth RK, Cui X, Kim HJ, et al: Oncogenic
transformation by the signaling adaptor proteins insulin receptor
substrate (IRS)-1 and IRS-2. Cell Cycle. 6:705–713. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yu Y and Alwine JC: Interaction between
simian virus 40 large T antigen and insulin receptor substrate 1 is
disrupted by the K1 mutation, resulting in the loss of large T
antigen-mediated phosphorylation of Akt. J Virol. 82:4521–4526.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Trojanek J, Croul S, Ho T, et al:
T-antigen of the human polyomavirus JC attenuates faithful DNA
repair by forcing nuclear interaction between IRS-1 and Rad51. J
Cell Physiol. 206:35–46. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Reiss K, Khalili K, Giordano A and
Trojanek J: JC virus large T-antigen and IGF-I signaling system
merge to affect DNA repair and genomic integrity. J Cell Physiol.
206:295–300. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bhattacharyya R, Noch EK and Khalili K: A
novel role of Rac1 GTPase in JCV T-antigen-mediated β-catenin
stabilization. Oncogene. 26:7628–7636. 2007.PubMed/NCBI
|
|
35
|
Lazo PA: The molecular genetics of
cervical carcinoma. Br J Cancer. 80:2008–2018. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
zur Hausen H: Papillomaviruses causing
cancer: evasion from host-cell control in early events in
carcinogenesis. J Natl Cancer Inst. 92:690–698. 2000.PubMed/NCBI
|
|
37
|
Popescu NC and DiPaolo JA: Preferential
sites for viral integration on mammalian genome. Cancer Genet
Cytogenet. 42:157–171. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lazo PA, Gallego MI, Ballester S and
Feduchi E: Genetic alterations by human papillomaviruses in
oncogenesis. FEBS Lett. 300:109–113. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yamakawa K, Kuno T, Hashimoto N, et al:
Molecular analysis of carcinogen-induced rodent lung tumors:
Involvement of microRNA expression and Krαs or Egfr mutations. Mol
Med Rep. 3:141–147. 2010.PubMed/NCBI
|
|
40
|
Monaco MC, Jensen PN, Hou J, et al:
Detection of JC virus DNA in human tonsil tissue: evidence for site
of initial viral infection. J Virol. 72:9918–9923. 1998.PubMed/NCBI
|