Introduction
Renal cell carcinoma (RCC) is the most lethal
urologic tumor and the sixth leading cause of cancer-related
mortality in Western countries. Each year, approximately 200,000
patients are diagnosed with this malignancy resulting in
approximately 100,000 deaths, and its incidence has steadily
increased in the last decades (1–3). At
present, the most effective treatment for localized RCC is surgical
resection; however, 30% of patients develop metastatic disease
after surgery, and the median survival for these patients is only
13 months. Therefore, novel therapeutic strategy as well as
prophylactic regimens are urgently required.
Prevention of numerous types of cancer through a
diet rich in fruits and vegetables has been in part attributed to
plant-derived botanicals, including polyphenolics, which may
protect against various chronic degenerative diseases such as
cancer or cardiovascular disease (4). Quercetin is a dietary flavonoid found
in tea, onion, grapes, wines and apples, and the anticancer
activities of quercetin have been assessed in several cancer cell
lines, as well as in vivo(5,6).
Hyperoside is a stilbene analog and a phytoalexin produced by
plants as a defense mechanism in response to fungal diseases,
stress and UV radiation; the primary dietary sources of hyperoside
are Malus micromalus and abelmosk (7). Hyperoside was found to exhibit
anticancer properties by inhibiting cell proliferation, inducing
apoptosis, decreasing angiogenesis, and causing cell cycle arrest
in several cancer cell lines (8). A
combination of quercetin and hyperoside (QH) previously exhibited
synergistic interactions in their anticancer effects on MOLT-4
leukemia cells (9). In a study with
mouse skin tumors, a combination of hyperoside with tea
polyphenolics demonstrated synergistic anticancer effects (10). In pancreatic cancer stem cells,
sulforaphane was found to synergize with quercetin in the
inhibition of the self-renewal of cells (11). Quercetin and epigallocatechin
gallate (EGCG) synergistically inhibited invasion, migration and
epithelial-mesenchymal transition in prostate cancer stem cells
(12).
Increasing evidence suggests that specificity
protein (Sp) transcription factors Sp1, Sp3 and Sp4 are
overexpressed in tumors and regulate genes important for cancer
cell death and survival (13,14).
More recently, the involvement of microRNA-27a (miR-27a) in the
regulation of Sp1 transcription factor through zinc finger protein
ZBTB10, an inhibitor of Sp1, has been demonstrated. Additionally,
botanical compounds were also found to inhibit Sp transcription
factors through the miR-27a-ZBTB10-Sp1 axis (15–17).
Therefore, the aim of this study was to investigate the effects of
a mixture of quercetin and hyperoside on Sp1, Sp3 and Sp4
expression and the potential involvement of miR-27a in the
downregulation of Sp transcription factors in 786-O renal cell
carcinoma cells.
Materials and methods
Botanical extract
Polyphenolics were extracted from a standardized QH
dehydrate supplement (ratio 1:1) in capsule form which was provided
by Suzhong Pharmaceutical Co., Ltd. (Taizhou, Jiangsu, China).
Polyphenolics were extracted with methanol (50 mg/50 ml) and then
centrifuged at room temperature for 10 min at 3,000 rpm to remove
inactive and insoluble components. Methanol was evaporated in a
rotavapor (BÜCHI Labortechnik AG, Flawil, Switzerland) at 40°C.
Residual moisture was evaporated in a speedvac concentrator (Thermo
Scientific, Waltham, MA, USA) at 43°C. The final QH mixture was
stored at −80°C and dissolved in dimethyl sulfoxide (DMSO) prior to
use.
HPLC-PDA analysis
The polyphenolic mixture was analyzed and quantified
by retention time and photodiode array (PDA) spectra by HPLC-PDA.
The chromatographic separation was performed in an Alliance 2695
system (Waters Corporation, Milford, MA, USA) and carried out in a
Discovery C18 column (Supelco Inc., Bellefonte, PA, USA) (250×4.6
mm, 5 μm) at room temperature. The chromatographic
conditions used were mobile phase A: water/acetic acid (98:2),
mobile phase B: acetonitrile/water/acetic acid (68:30:2). A
gradient program with 1 ml/min was used as follows: 0 min 100%, A;
20 min 60%, A; 30 min 30%, A; 32 min 0%, A; 35 min 100%, A. The
detection wavelengths were set at 360 and 306 nm for quercetin and
hyperoside, respectively. Standard compounds for the identification
and quantitative analysis of quercetin and hyperoside were obtained
from Acros Organics (Morris Plains, NJ, USA).
Oxygen radical absorbance capacity
The antioxidant capacity was determined using the
oxygen radical absorbance capacity assay (ORAC) (22) with fluorescein as the fluorescent
probe in a FLUOstar fluorescence microplate reader (485 nm
excitation and 538 nm emission; BMG Labtech Inc., Durham, NC, USA).
Results were reported in μmol of Trolox equivalents/ml.
Cell culture
Human clear cell renal carcinoma 786-O cells
[American Type Culture Collection (ATCC), Manassas, VA, USA] were
cultured in Mc-Coy’s medium supplemented with 10% fetal bovine
serum (FBS) and 1% antibiotics (100,000 U/l penicillin and 100 mg/l
streptomycin). Cells were maintained in a humidified atmosphere of
95% air and 5% CO2 at 37°C. For experiments, cells were
seeded in DMEM/F-12 medium with 2.5% FBS, and 1% antibiotics
(100,000 U/l penicillin and 100 mg/l streptomycin). Generation of
reactive oxygen species (ROS) was determined using the
2′,7′-dichlorofluorescein diacetate (DCFH-DA) assay. In summary,
1×104 cells/well were seeded in a clear bottom 96-well
plate and incubated for 24 h, and then treated with different
concentrations of QH (0–60 μg/ml). After 24 h, cells were
washed twice with phosphate-buffered saline (PBS) and incubated
with 200 μM hydrogen peroxide for 2 h at 37°C. Hydrogen
peroxide was removed with PBS washes, and 10 μM DCFH-DA
diluted in PBS was added to the cells. Next, cells were incubated
for 15 min at 37°C, DCFH-DA was removed, and the fluorescence
intensity was measured after 15 min at 37°C using a FLUOstar
fluorescent microplate reader (485 nm excitation and 538 nm
emission).
Cell viability
Cells were seeded (3×103 cells/well) in a
96-well plate for 24 h, and the growth medium was then replaced
with the experimental medium containing various extract
concentrations of QH (0–60 μg/ml). Cell viability was
assessed at 48, 72 and 96 h with the CellTiter 96®
AQueous One Solution Cell Proliferation assay (Promega, Madison,
WI, USA) according to the manufacturer’s protocol using a FLUOstar
microplate reader at 490 nm. The concentration at which cell
viability was inhibited by 50% (IC50) was calculated by
sigmoidal nonlinear regression analyses of the percentage of cell
inhibition, expressed as a ratio to the control, using GraphPad
Prism 5.01 software (GraphPad Software Inc., La Jolla, CA,
USA).
Cell proliferation
786-O cells (2×104 cells/well) were grown
in a 24-well plate for 24 h. The growth medium was then replaced
with experimental medium containing QH extract concentrations
ranging from 0 to 60 μg/ml. After 72 h cell proliferation
was determined using a cell counter (Beckman Coulter Inc.,
Fullerton, CA, USA). Cell counts were expressed as a percentage of
the control cells.
Cleaved caspase-3 activation
Cells were grown (6×105 cells/well) for
24 h and then incubated with different QH concentrations (0–30
μg/ml) for 24 h. Cleaved caspase-3 activation was determined
using an ELISA kit (Cell Signaling Technology Inc., Danvers, MA,
USA) according to the manufacturer’s protocol using a FLUOstar
microplate reader at 450 nm.
Real-time PCR analysis of miRNA and
mRNA
786-O cells were grown (2×105 cells/well)
in a 6-well plate for 24 h before incubation with different
concentrations of QH (0–30 μg/ml). Total RNA, which contains
both mRNA and miRNA, was isolated using the mirVana™ miRNA
Isolation kit (Applied Biosystems Inc., Foster City, CA, USA)
following the manufacturer’s recommended protocol. Extracted
nucleic acid was evaluated for quality and quantity using the
NanoDrop® ND-1000 Spectrophotometer (NanoDrop
Technologies, Wilmington, DE, USA) at 260 and 280 nm.
SuperScript™ III First-Strand (Invitrogen, Carlsbad,
CA, USA) was used to reverse-transcribe mRNA. TATA-binding protein
(TPB) was used as an mRNA endogenous control. For real time PCR
(RT-PCR), proprietary primers for Sp3 and Sp4 (Qiagen Inc.,
Valencia, CA, USA) were used. The following primers were purchased
from Integrated DNA Technologies, Inc. (Coralville, IA, USA) and
used for amplification as follows: TBP (sense, 5′-TGC ACA GGA GCC
AAG AGT GAA-3′ and antisense, 5′-CAC ATC ACA GCT CCC CAC CA-3′),
caspase-3 (sense, 5′-CTG GAC TGT GGC ATT GAG ACA-3′ and antisense,
5′-CGG CCT CCA CTG GTA TTT TAT G-3′), ZBTB10 (sense, 5′-GCT GGA TAG
TAG TTA TGT TGC-3′ and antisense, 5′-CTG AGT GGT TTG ATG GAC AGA
G-3′), Sp1 (sense, 5′-TCA CCA ATG CCA ATA GCT ACT CA-3′ and
antisense, 5′-GAG TTG GTC CCT GAT GAT CCA-3′), survivin (sense,
5′-CCA TGC AAA GGA AAC CAA CAA T-3′ and antisense, 5′-ATG GCA CGG
CGC ACT T-3′). RT-PCR for mRNA was performed using the SYBR GreenER
qPCR SuperMix (Invitrogen) on a 7900HT Fast Real-Time PCR System
(Applied Biosystems Inc.).
The TaqMan® MicroRNA Assay for miR-27a
and RNU6B (used as the control; Applied Biosystems Inc.) was used
to reverse-transcribe mature miRNA following the manufacturer’s
protocol in a MasterCycler (Eppendorf North America Inc., Westbury,
NY, USA). RT-PCR for miRNA was carried out with the
TaqMan® assay, which contained the forward and reverse
primers as well as the TaqMan® probe and
TaqMan® Universal PCR Master mix, No
AmpErase® UNG (Applied Biosystems Inc.). After
completion of RT-PCR, relative quantification for both mRNA and
miRNA gene expression was evaluated by using the comparative
critical threshold (CT) method. Transfections with 50 and 100 nM
miR-27a mimic (Dharmacon, Lafayette, CO, USA) were performed using
Lipofectamine 2000 (Invitrogen) for 6 h. After transfection, cells
were incubated with 20 μg/ml of QH for 24 h.
Cell cycle kinetics
Cells were seeded (5×105 cells/plate)
with medium containing 2.5% FBS for 24 h and were subsequently
treated with QH (0–20 μg/ml) for 24 h. Cells were fixed with
90% ethanol and stored at −20°C. DNA was stained with propidium
iodide containing RNAse (0.2 mg/ml) solution and analysis was
carried out at 488 nm excitation and 620 nm emission wavelengths on
a FACScan flow cytometer (BD Immonunocytometry Systems, San José,
CA, USA). The percentage of cells in each cell cycle phase was
analyzed using the ModFit LT version 3.2 for Macintosh by Verity
Software House (Topsham, ME, USA).
Western blotting
Cells were grown (2×106 cells/plate) for
24 h and treated with QH (0–30 μg/ml) for 24 h. Cells were
harvested and cell lysates were obtained using a high-salt buffer
(1.5 mmol/l MgCl2, 500 mmol/l NaCl, 1 mmol/l EGTA, 50
mmol/l HEPES, 10% glycerol, 1% Triton X-100; adjusted to pH 7.5)
supplemented with protease inhibitors (Sigma-Aldrich, St. Louis,
MO, USA). Samples were incubated at 100°C for 5 min in 1X Laemmli
buffer (0.1% bromophenol blue, 175 mM β-mercaptoethanol, 50 mM
Tris-HCl, 2% SDS). Proteins were separated on a 10% (Sp proteins)
or 12% (survivin) SDS-PAGE at 100 V and transferred to PVDF
membranes (Bio-Rad, Hercules, CA, USA). Membranes were blocked in
nonfat milk in PBS-Tween and incubated with primary antibodies
overnight at 4°C. After multiple washing steps, membranes were
incubated with secondary antibodies. Membranes were washed and
incubated with chemiluminescence substrate (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA), and proteins were
visualized with a Kodak Molecular Imaging System (Carestream Health
Inc., Rochester, NY, USA).
Statistical analyses
Data from the in vitro experiments were
analyzed by one-way analysis of variance with JMP 8.0 (SAS
Institute, Cary, NC, USA). Differences were deemed significant at
p≤0.05 using a Tukey-Cramer HSD comparison for all pairs. For
transfections with the miR-27a mimic, differences were deemed
significant at p≤0.05 using a Student’s t-test comparison for all
pairs. The analysis of linear (pairwise) correlations was performed
where correlations with a p-value <0.05 were deemed significant.
Nonlinear modeling of sigmoidal curves for cell viability was
performed using GraphPad Prism 5.01 software (GraphPad Software
Inc.).
Results
Chemical composition
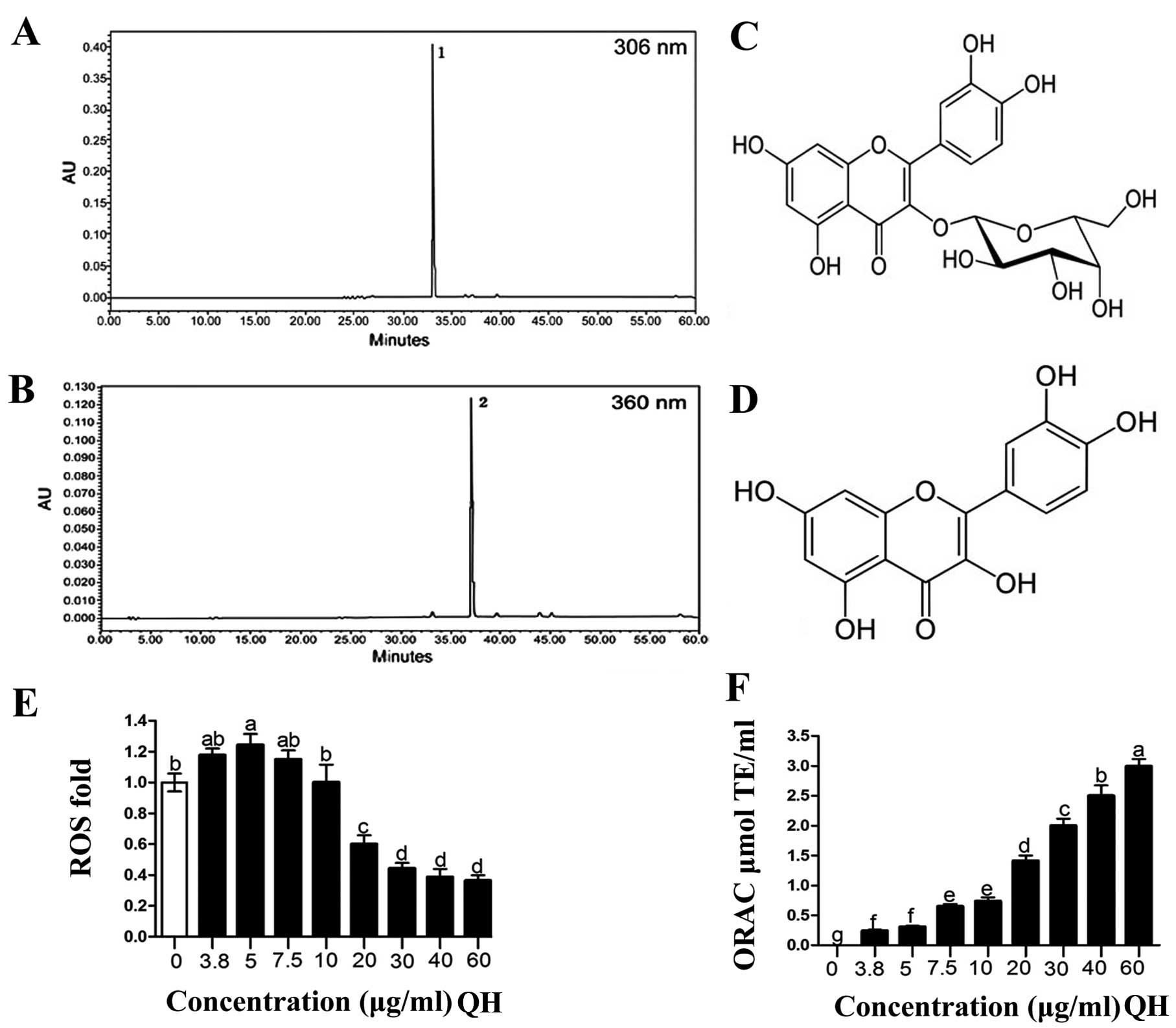
The chromatographic profiles (Fig. 1A and B) of QH demonstrated the
presence of 2 major polyphenols, hyperoside (peak 1) and quercetin
(peak 2), respectively, as major ingredients of this botanical
supplement. The chemical structures of quercetin and hyperoside are
listed in Fig. 1C and D. In fact,
both quercetin and hyperoside are among the most abundant
flavonoids in the routine human diet (4,18).
Generation of intracellular ROS and
ORAC
Intracellular generation of ROS was investigated
after 786-O cells were challenged with hydrogen peroxide. QH
slightly induced the generation of ROS at low concentrations (0–10
μg/ml) whereas at concentrations >20 μg/ml,
generation of ROS was significantly reduced (Fig. 1E) by up to 66% compared to that of
solvent-treated control cells. The antioxidant capacity values
(ORAC) (Fig. 1F), were
significantly increased by all concentrations of QH in a
dose-dependent manner.
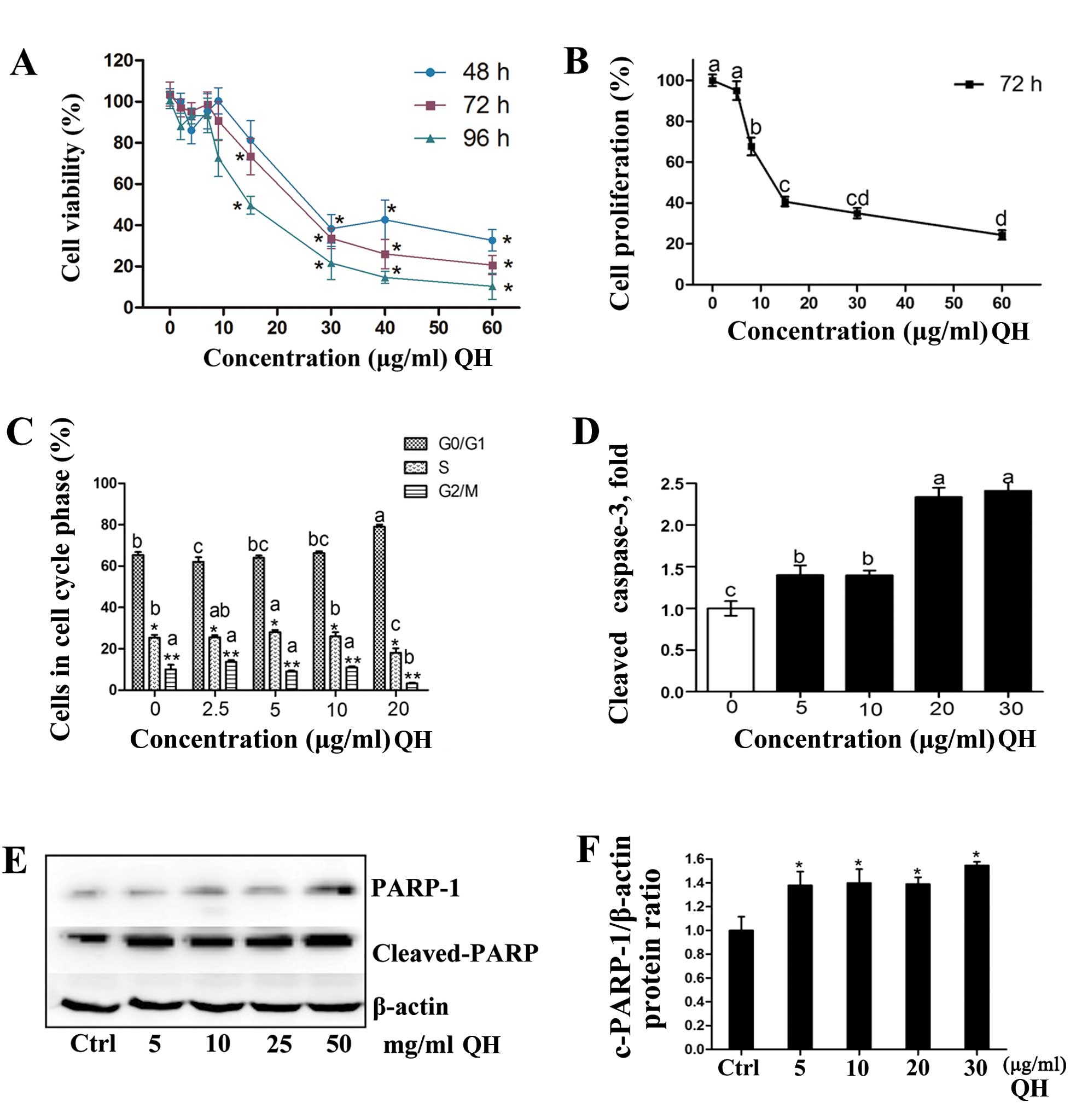
Cell death and cell cycle kinetics
786-O cell viability (Fig. 2A) was significantly decreased by QH
in a dose- and time-dependent manner. The IC50 values
were 18.2, 18.7 and 11.8 μg/ml at 48, 72 and 96 h,
respectively. QH also inhibited cell proliferation after treatment
for 72 h (Fig. 2B) illustrated by a
decrease in cell counts in a dose-dependent manner. The net number
of cells remaining was the combined results of cancer cell
proliferation as well as the cytotoxicity of QH. Cell numbers were
significantly decreased at all concentrations (3.75–60
μg/ml), and at 60 μg/ml QH cell proliferation was
reduced by up to 73%.
The effects of QH on cell-cycle progression were
determined by fluorescence-activated cell sorting (FACS) analysis
(Fig. 2C). There were minor but
significant changes in the percentage of cells in different phases
of the cell cycle using 2.5, 5, or 10 μg/ml of extract.
There was a significant G0/G1 to S-phase
arrest when compared to the control cells when cells were treated
with 20 μg/ml of the mixture (Fig. 2C).
Cleaved caspase-3, the activated form, is involved
in the execution of apoptosis and was induced up to 1.5-fold even
at low concentrations of QH (5 and 10 μg/ml) (Fig. 2D). At higher concentrations (20 and
30 μg/ml), the induction increased up to 2.3-fold.
Poly(ADP-ribose) polymerase 1 (PARP-1) is a substrate for caspase-3
cleavage and produces cleaved PARP-1 (19). There was an increase in PARP
cleavage when cells were treated with QH, as also shown in the
densitometry analysis for PARP-1 cleavage/β-actin (Fig. 2E).
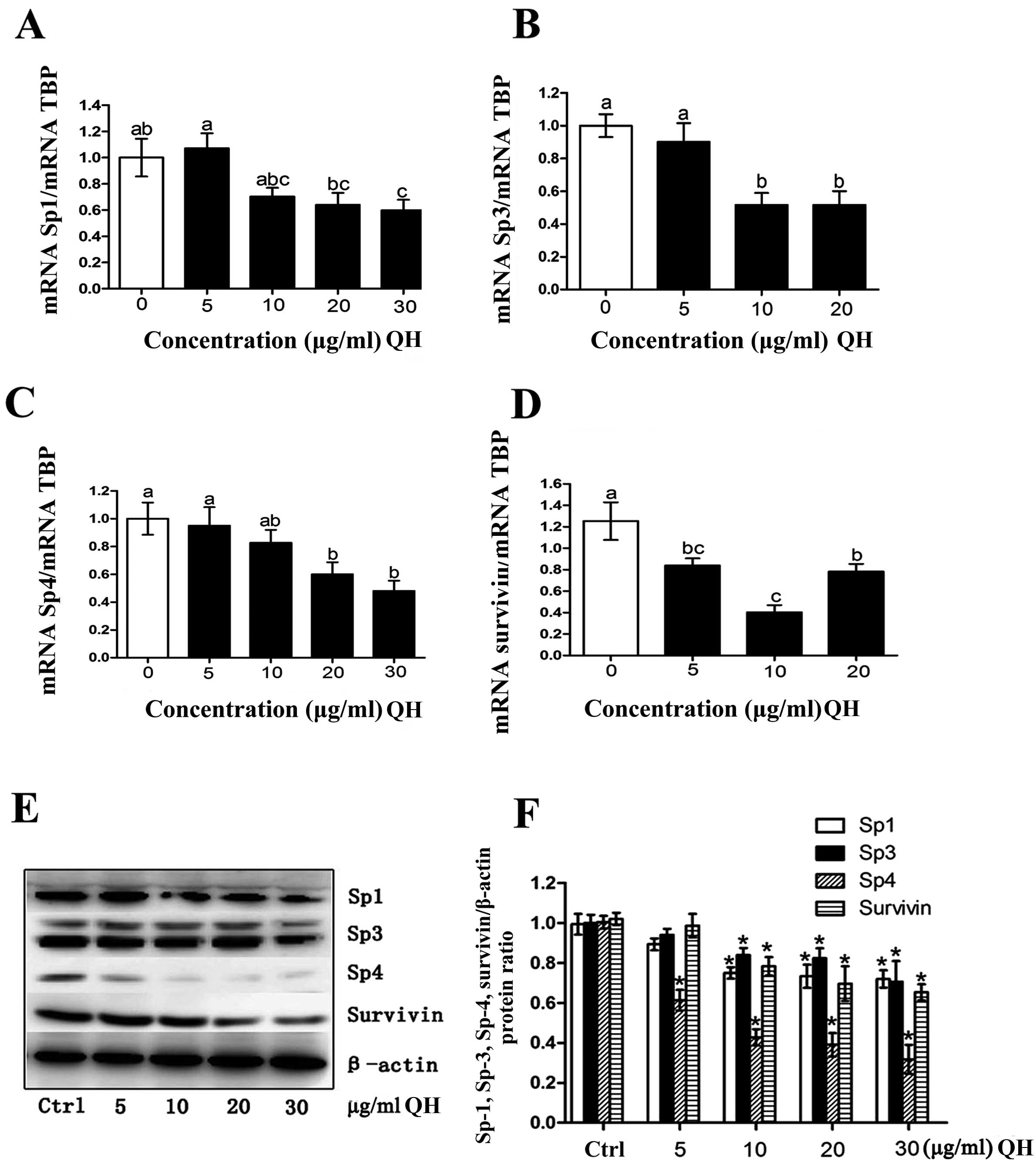
Modulation of Sp1 transcription factors
and dependent genes
Previous studies with several botanical anticancer
agents and their synthetic derivatives have demonstrated that these
compounds downregulate Sp transcription factors and Sp-regulated
genes (20). The present results
showed that QH (5–30 μg/ml) decreased Sp1, Sp3 and Sp4 mRNA
levels, and there was also a parallel decrease in Sp1, Sp3 and Sp4
proteins (Fig. 3A–C and E). In
addition, QH also decreased expression of survivin protein and
mRNA, and these results are consistent with QH-mediated suppression
of Sp transcription factors since survivin is an Sp-regulated gene.
We also carried out densitometry for the Sp1, Sp3, Sp4, and
Sp-regulated genes, expressed as a ratio to β-actin (Fig. 3F).
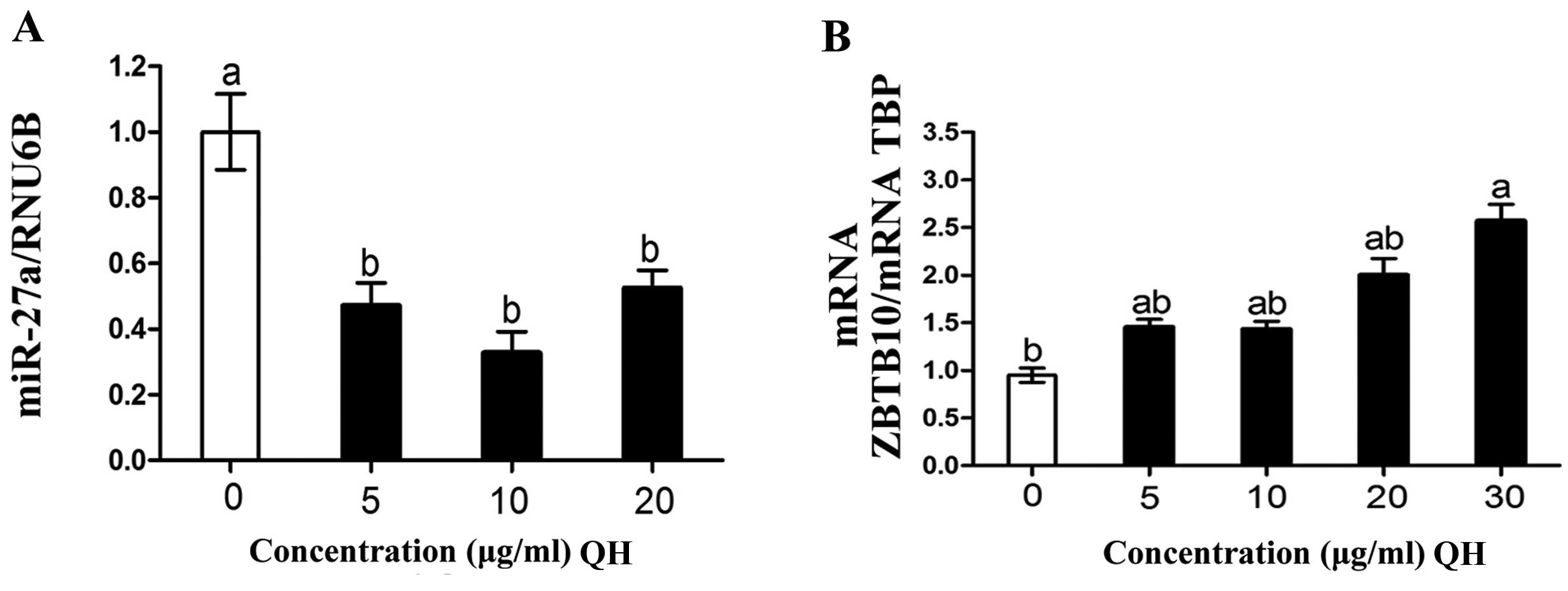
Previous studies have demonstrated that the
inhibition of ZBTB10 expression by miR-27a is at least in part
responsible for the increased expression of Sp transcription
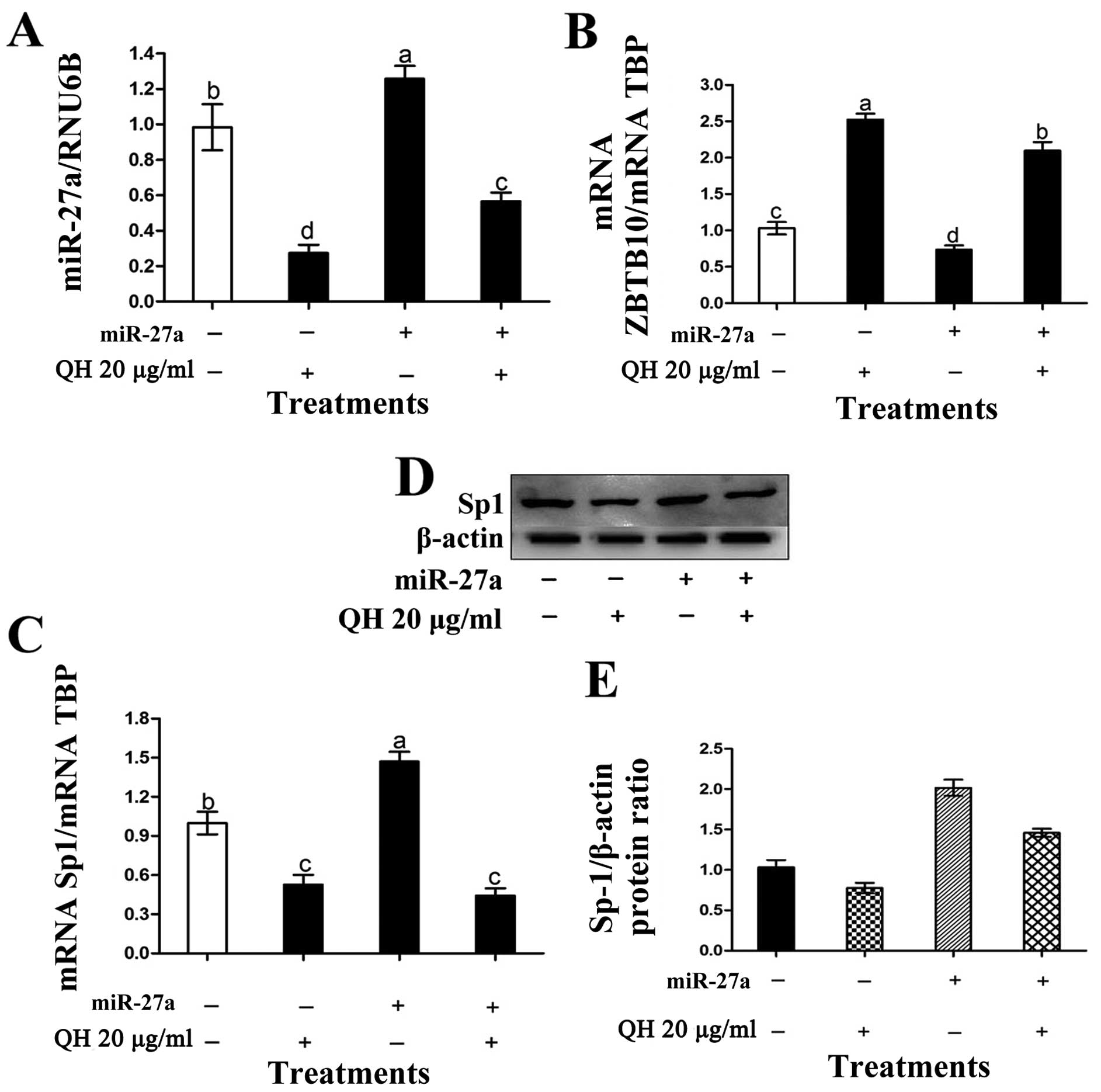
factors in cancer cells and tumors (21). In the present study, QH
significantly decreased miR-27a (Fig.
4A) and upregulated ZBTB10 mRNA (Fig. 4B), and this was accompanied by
decreased Sp proteins and the Sp-regulated gene survivin (Fig. 3D and E). These results are
consistent with previous studies regarding the effects of other
botanicals and their derivatives on the miR-27a-ZBTB10-Sp1 axis in
multiple cancer cell lines (21).
In this study, the overexpression of the miR-27a mimic in 786–0
cells increased miR-27a, decreased ZBTB10 and increased Sp1 mRNA
levels and this was consistent with the inactivation of endogenous
ZBTB10 expressed in these cells (Fig.
5A–D). Moreover, the miR-27a mimic also partially reversed
QH-induced downregulation of miR-27a, induction of ZBTB10, and
downregulation of Sp1 (Fig. 5).
Densitometry was performed for Sp-1/β-actin (Fig. 5E).
Discussion
Generation of intracellular ROS and
ORAC
The generation of ROS has been associated with
oxidative cellular damage that may be involved in the development
of various pathological conditions (22). The role of ROS in carcinogenesis is
complex. Cancer cells tend to have higher constitutive levels of
ROS than normal cells, due in part to mutations in nuclear and
mitochondrial genes responsible for the electron transport chain
and also due to increased metabolism and mitochondrial activity
(23). Elevated levels of ROS may
enhance cell proliferation and other events relevant to cancer
progression (23). ROS can also
cause DNA damage and oxidation of fatty acids in cellular membrane
structures, thereby facilitating mutagenesis and cancer development
(23). However, many anticancer
drugs are mitochondro-toxic and induce ROS, which in turn can lead
to cancer cell death. Polyphenolics, such as quercetin and
hyperoside, have the ability to scavenge free radicals and also
induce activation of antioxidant and detoxifying enzymes, thus
protecting cells against oxidative damage from carcinogenic
compounds (24). In this study, QH
induced a concentration-dependent increase in intracellular
antioxidant capacity (Fig. 1E) and
the intracellular concentration of ROS increased at a low QH
concentration (5 μg/ml) but decreased at high concentrations
(20–60 μg/ml) (Fig. 1E). One
possible explanation for this biphasic effect may be the
mitochondrotoxic properties of polyphenolics, which also induce
ROS, whereas at higher concentrations their antioxidant properties
are predominant. Studies with polyphenolics in cancer cells
demonstrate their protective effects against oxidative damage in
some conditions (25). In this
study, QH induced ROS at lower concentrations, potentially because
of the additional formation of radicals, where QH concentrations
were too low to have a protective effect. At higher concentrations,
QH inhibited the generation of ROS, potentially through scavenging
ROS.
Cell death and cell cycle kinetics
The anti-proliferative effects of quercetin and
hyperoside were previously demonstrated in MOLT-4 leukemia cells,
and the combination of resveratrol and quercetin exhibited
synergistic effects (9). Based on
the previous study, a combination of quercetin and hyperoside at a
ratio of 1:1 was investigated in the present study, resulting in a
significant decrease in cell viability and cell proliferation
following treatment with QH (Fig. 2A
and B). However, it must be considered that for the chosen cell
model, another ratio may have been more effective.
It was previously demonstrated that cell cycle
arrest is induced by polyphenolics, including resveratrol and
quercetin in several cancer cell lines within different phases
(26). Tan et al(27) studied the effect of quercetin on
HepG2 cells and found that following treatment with quercetin for
48 h, cells were arrested in the G0/G1 phase.
In MOLT-4 leukemia cells, polyphenolic-mediated cell cycle arrest
was influenced by the duration of treatment and the type of
polyphenolic. In the present study, QH (20 μg/ml) decreased
the percentage of cells in the S-phase and increased the percentage
of cells in the G0/G1 phase, which is
consistent with the inhibition of the progression from
G0/G1 to S-phase (Fig. 2C).
Caspase-3 is a major executive enzyme in apoptosis
and a commonly used indicator for the induction of apoptosis
(28). PARP-1, an abundant
chromatin-associated protein, plays an important role in
maintaining genome integrity and is cleaved during apoptosis by
caspase-3 (29). Previous studies
have demonstrated the effects of quercetin, as well as of a number
of other polyphenolics, on caspase-3 and PARP-1 activity (30). In general, the findings of this
study are in concordance with previous studies that found that
polyphenolics induce apoptosis through the activation of caspase-3
accompanied by cleavage of the DNA repair enzyme poly(ADP-ribose)
polymerase (PARP) (28). Previous
studies have shown that resveratrol causes induction of caspase-3
and cleavage of PARP in human articular chondrocytes and myeloid
leukemia cells (28).
Modulation of Sp1 transcription factors
and dependent genes
Numerous botanical compounds have previously been
demonstrated to downregulate Sp transcription factors and
Sp-regulated genes (21). For
example, curcumin decreased Sp1, Sp3, and Sp4 levels in bladder and
pancreatic cancer cells. The terpenoid betulinic acid also
decreased expression of these transcription factors in prostate
cancer cells, and synthetic analogs of the triterpenoids oleanolic
and glycyrrhetinic acid also exhibited comparable effects (22). The importance of Sp1 downregulation
in terms of the anticancer activity of these compounds is that
Sp-regulated genes play an important role in cancer cell and tumor
growth (cyclin D1, c-MET and EGFR), survival (NF-κB-p65, survivin
and Bcl-2), and angiogenesis (VEGF and its receptors) (31).
Sp1 overexpression in gastric and pancreatic cancer
patients is a negative prognostic factor, as there is evidence that
Sp1 exhibits oncogenic properties and plays a role in cell
transformation and maintenance of the cancer phenotype (32). The pathways associated with the
induction of high levels of Sp1, Sp3 and Sp4 during transformation
are not known. However, our study indicated that at least one
mechanism for their elevated expression in cancer cells and tumors
was due to inhibition of ZBTB10 expression by miR-27a. ZBTB10 is a
translational repressor and binds GC-rich sites to decrease
Sp-dependent trans-activation (33). In the present study, the effects of
QH were not dose-dependent for miR-27a itself, but for ZBTB10
(Fig. 4A and B). Many miRNAs play
important roles in carcinogenesis, with either oncogenic or
tumor-suppressing activities (34).
The overexpression of oncogenic miRNAs has been demonstrated in
several cancer cell lines (35). In
previous studies, miR-27a expression was increased in 6 breast
cancer cell lines and also in colon cancer cells (35,36).
In the present study, a mimic for miR-27a partially reversed the
effects of the extracts on Sp1 protein; however these effects were
not significant for Sp1 mRNA.
Results from this study for the first time
demonstrated that apoptosis induced by QH and inhibition of HT-29
cell growth was associated with decreased expression of Sp1, Sp3,
Sp4, and Sp-regulated survivin. These latter responses were
paralleled by perturbation of the miR-27a-ZBTB10 axis, resulting in
the induction of ZBTB10, a potent Sp-repressor gene (36). These effects of QH on miR-27a-ZBTB10
were observed at concentrations of QH that decreased ROS in the
HT-29 cells (Fig. 1E), whereas
previous studies with curcumin and the synthetic triterpenoid
methyl-2-cyano-3, 12-dioxooleana-1, 9-dien-28-oate (CDDOMe) showed
that their effects on miR-27a-ZBTB10 were dependent on the
generation of ROS (36). Ongoing
studies in our laboratory are focused on mechanisms of
ROS-independent downregulation of miR-27a by QH and other
anticancer agents as this pathway plays an important role in the
anticancer activity of botanicals and their derivates.
In conclusion, the present results revealed that a
combination of quercetin and hyperoside had cytotoxic effects on
colon cancer cells, resulting in apoptosis. Interactions of QH and
the miR-27a-ZBTB10-Sp1 axis were identified as one possible
underlying mechanism. Further studies are needed to assess the role
of miR-27a and its clinical relevance in the anticancer effects
exhibited by botanicals.
Acknowledgements
This study was partially supported by grants from
the National Natural Science Foundation of China (nos. 81000311 and
81270831).
References
|
1
|
Miyamoto H, Miller JS, Fajardo DA, et al:
Non-invasive papillary urothelial neoplasms: the 2004 WHO/ISUP
classification system. Pathol Int. 60:1–8. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zisman A, Pantuck AJ, Wieder J, et al:
Risk group assessment and clinical outcome algorithm to predict the
natural history of patients with surgically resected renal cell
carcinoma. J Clin Oncol. 20:4559–4566. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Eble JN, Sauter G, Epstein JI and
Sesterhenn I: World Health Organization Classification of Tumors.
Pathology and Genetics of Tumors of the Urinary System and Male
Genital Organs. IARC Press; Lyon: 2005
|
|
4
|
Theodoratou E, Kyle J, Cetnarskyj R, et
al: Dietary flavonoids and the risk of colorectal cancer. Cancer
Epidemiol Biomarkers Prev. 16:684–693. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim MJ, Kim YJ, Park HJ, et al: Apoptotic
effect of red wine polyphenols on human colon cancer SNU-C4 cells.
Food Chem Toxicol. 44:898–902. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mertens-Talcott SU and Percival SS:
Ellagic acid and quercetin interact synergistically with
resveratrol in the induction of apoptosis and cause transient cell
cycle arrest in human leukemia cells. Cancer Lett. 218:141–151.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chu CY, Lee HJ, Chu CY, et al: Protective
effects of leaf extract of Zanthoxylum ailanthoides on
oxidation of low-density lipoprotein and accumulation of lipid in
differentiated THP-1 cells. Food Chem Toxicol. 47:1265–1271.
2009.PubMed/NCBI
|
|
8
|
Gao LL, Feng L, Yao ST, et al: Molecular
mechanisms of celery seed extract induced apoptosis via S phase
cell cycle arrest in the BGC-823 human stomach cancer cell line.
Asian Pac J Cancer Prev. 12:2601–2606. 2011.PubMed/NCBI
|
|
9
|
Mertens-Talcott SU, Talcott ST and
Percival SS: Low concentrations of quercetin and ellagic acid
synergistically influence proliferation, cytotoxicity and apoptosis
in MOLT-4 human leukemia cells. J Nutr. 133:2669–2674. 2003.
|
|
10
|
George J, Singh M, Srivastava AK, et al:
Resveratrol and black tea polyphenol combination synergistically
suppress mouse skin tumors growth by inhibition of activated MAPKs
and p53. PLoS One. 6:e233952011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Srivastava RK, Tang SN, Zhu W, et al:
Sulforaphane synergizes with quercetin to inhibit self-renewal
capacity of pancreatic cancer stem cells. Front Biosci (Elite Ed).
3:515–528. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tang SN, Singh C, Nall D, et al: The
dietary bioflavonoid quercetin synergizes with epigallocatechin
gallate (EGCG) to inhibit prostate cancer stem cell
characteristics, invasion, migration and epithelial-mesenchymal
transition. J Mol Signal. 5:142010. View Article : Google Scholar
|
|
13
|
Chadalapaka G, Jutooru I, Burghardt R and
Safe S: Drugs that target specificity proteins downregulate
epidermal growth factor receptor in bladder cancer cells. Mol
Cancer Res. 8:739–750. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chadalapaka G, Jutooru I, Chintharlapalli
S, et al: Curcumin decreases specificity protein expression in
bladder cancer cells. Cancer Res. 68:5345–5354. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chintharlapalli S, Papineni S, Abdelrahim
M, et al: Oncogenic microRNA-27a is a target for anticancer agent
methyl 2-cyano-3,11-dioxo-18β-olean-1,12-dien-30-oate in colon
cancer cells. Int J Cancer. 125:1965–1974. 2009.PubMed/NCBI
|
|
16
|
Jutooru I, Chadalapaka G, Abdelrahim M, et
al: Methyl 2-cyano-3,12-dioxooleana-1,9-dien-28-oate decreases
specificity protein transcription factors and inhibits pancreatic
tumor growth: role of microRNA-27a. Mol Pharmacol. 78:226–236.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jutooru I, Chadalapaka G, Lei P and Safe
S: Inhibition of NF-κB and pancreatic cancer cell and tumor growth
by curcumin is dependent on specificity protein down-regulation. J
Biol Chem. 285:25332–25344. 2011.
|
|
18
|
Frémont L: Biological effects of
resveratrol (Review). Life Sci. 66:663–673. 2000.
|
|
19
|
Brauns SC, Dealtry G, Milne P, et al:
Caspase-3 activation and induction of PARP cleavage by cyclic
dipeptide Cyclo(Phe-Pro) in HT-29 cells. Anticancer Res.
25:4197–4202. 2005.PubMed/NCBI
|
|
20
|
Meng Q, Velalar CN and Ruan R: Effects of
epigallocatechin-3-gallate on mitochondrial integrity and
antioxidative enzyme activity in the aging process of human
fibroblast. Free Rad Biol Med. 44:1032–1041. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mertens-Talcott SU, Chintharlapalli S, Li
MR and Safe S: The oncogenic microRNA-27a targets genes that
regulate specificity protein transcription factors and the
G2-M checkpoint in MDA-MB-231 breast cancer cells.
Cancer Res. 67:11001–11011. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fresco P, Borges F, Diniz C and Marques
MP: New insights on the anticancer properties of dietary
polyphenols. Med Res Rev. 26:747–766. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schumacker PT: Reactive oxygen species in
cancer cells: live by the sword, die by the sword. Cancer Cell.
10:175–176. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fiander H and Schneider H: Dietary ortho
phenols that induce glutathione S-transferase and increase the
resistance of cells to hydrogen peroxide are potential cancer
chemopreventives that act by two mechanisms: the alleviation of
oxidative stress and the detoxification of mutagenic xenobiotics.
Cancer Lett. 156:117–124. 2000.
|
|
25
|
Hsu SC, Lu JH, Kuo CL, et al: Crude
extracts of Solanum lyratum induced cytotoxicity and
apoptosis in a human colon adenocarcinoma cell line (colo 205).
Anticancer Res. 28:1045–1054. 2008.
|
|
26
|
Schlachterman A, Valle F, Wall KM, et al:
Combined resveratrol, quercetin, and catechin treatment reduces
breast tumor growth in a nude mouse model. Transl Oncol. 1:19–27.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tan J, Wang B and Zhu L: Regulation of
survivin and Bcl-2 in HepG2 cell apoptosis induced by quercetin.
Chem Biodivers. 6:1101–1110. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shakibaei M, John T, Seifarth C and
Mobasheri A: Resveratrol inhibits IL-1β-induced stimulation of
caspase-3 and cleavage of PARP in human articular chondrocytes
in vitro. Ann NY Acad Sci. 1095:554–563. 2007.
|
|
29
|
Wang ZQ, Stingl L, Morrison C, et al: PARP
is important for genomic stability but dispensable in apoptosis.
Genes Dev. 11:2347–2358. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hsu CP, Lin YH, Chou CC, et al: Mechanisms
of grape seed procyanidin-induced apoptosis in colorectal carcinoma
cells. Anticancer Res. 29:283–289. 2009.PubMed/NCBI
|
|
31
|
Lee HS, Park CK, Oh E, et al: Low SP1
expression differentially affects intestinal-type compared with
diffuse-type gastric adenocarcinoma. PLoS One. 8:e555222013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Deacon K, Onion D, Kumari R, et al:
Elevated SP-1 transcription factor expression and activity drives
basal and hypoxia-induced vascular endothelial growth factor (VEGF)
expression in non-small cell lung cancer. J Biol Chem.
287:39967–39981. 2012. View Article : Google Scholar
|
|
33
|
Kim K, Chadalapaka G, Lee SO, et al:
Identification of oncogenic microRNA-17–92/ZBTB4/specificity
protein axis in breast cancer. Oncogene. 31:1034–1044.
2012.PubMed/NCBI
|
|
34
|
Lynam-Lennon N, Maher SG and Reynolds JV:
The roles of microRNA in cancer and apoptosis (Review). Biol Rev
Camb Philos Soc. 84:55–71. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tang W, Zhu J, Su S, et al: MiR-27 as a
prognostic marker for breast cancer progression and patient
survival. PLoS One. 7:e517022012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Noratto GD, Jutooru I, Safe S, et al: The
drug resistance suppression induced by curcuminoids in colon cancer
SW-480 cells is mediated by reactive oxygen species-induced
disruption of the microRNA-27a-ZBTB10-Sp axis. Mol Nutr Food Res.
57:1638–1648. 2013. View Article : Google Scholar : PubMed/NCBI
|