Introduction
Corticotropin-releasing factor (CRF) was originally
reported as a major hypothalamic mediating factor produced in
response to stress. The function of CRF in the homeostatic response
to stress is well-characterized. The CRF system, consisting of CRF
and CRF receptor (CRFR), has been identified in the cardiovascular,
reproductive and gastrointestinal systems. CRF systems have been
found in the immune system and are shown to possess
immunomodulatory properties. Several studies have reported that CRF
and its receptors have been found in many types of tumors (1–3).
Urocortin, a CRF family member, was detected in various tumors,
including pituitary adenoma, prostatic carcinoma and endometrial
carcinoma (4–6). Notably, many studies report that CRF
has both pro- and anticancer activity. Graziani et al
revealed that CRF inhibited the proliferation of endometrial
adenocarcinoma cells and human mammary cancer cells (7). CRF also inhibits proliferation of
melanoma cells through mediation of CRFR1 (8). In contrast, other evidence suggests
that CRF can promote cancer malignancy (9). Yang et al observed that CRF
promoted the migration of melanoma cells via the ERK pathway
(10). CRF can protect cancer cells
from pro-apoptotic signals blocking activation of pro-caspase-3
(11). However, the cause of these
opposing effects in tumors is not well understood.
Natural killer (NK) cells can recognize and lyse
virus-infected cells and tumor cells (12). NK killing activity against tumor
cells is a process that results from counterbalance between
inhibitory and stimulatory signals. Specifically, NKG2D (activating
receptor) mediates NK cell activation by overcoming the inhibitory
signals from self-recognition (13). Moreover, NKG2D ligand is expressed
on tumor cells and is induced by danger signals such as cellular
stress and virus infection, suggesting that NKG2D ligands are
potentially useful therapeutic targets for eliminating tumor- and
virus-infected cells using NK cells (14–17).
However, some studies have reported that tumor cells can produce
soluble forms of NKG2D ligands that lead to downregulated surface
NKG2D expression on NK and CTL cells (18–20).
Salih et al observed that soluble NKG2D ligands secreted by
tumor cells downregulate surface expression of NKG2D on NK cells
and lead to NK cell malfunction. Moreover, we previously reported
that soluble ULBP2 produced by gastric cancer inhibits function of
NK cells through downregulation of NKG2D expression (21–23).
In the present study, we investigated whether
expression of NKG2D ligands is regulated by CRF in human cervical
cancer cell lines, and we also defined the functional consequences
of CRF-regulated NKG2D ligand production in cervical cancer cells
following interaction between tumor and NK cells.
Materials and methods
Cell culture and reagents
Human cervical carcinoma cell line, HeLa, human
alveolar basal epithelial cells, A549, and human leukemia cells,
NK92 MI, were purchased from the American Type Culture Collection
(ATCC; Manassas, VA, USA). HeLa and A549 cells were cultured in
RPMI-1640 medium supplemented with 10% heat-inactivated fetal calf
serum. NK92 MI cells were cultured in α modification of Minimum
Essential Medium Eagle (α-MEM) supplemented with 2 mM L-glutamine,
0.2 mM inositol, 20 mM folic acid, 12.5% FBS and 12.5% horse serum.
Anti-NKG2D antibodies were purchased from R&D Systems
(Minneapolis, MN, USA). 1-[6-(17-3-methoxyestra-1,3,5(10)-trien-17-yl)amino)hexyl]-1H-pyrrole-2,5-dione
(U73122) was purchased from Calbiochem (La Jolla, CA, USA). CRF and
1,10-phenanthroline (pan-matrix metalloprotease inhibitor) were
purchased from Sigma-Aldrich (St. Louis, MO, USA).
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA was extracted from tumor cells using an
RNeasy kit (Qiagen, Valencia, CA, USA). cDNA was prepared and used
for RT-PCR with the following primers: human ULBP2 primer (sense,
5′-CTGCAGGTAAGGATGTCTTGTGAG-3′ and antisense,
5′-TGAGGGTGGTGGCTATGGC-3′, product 350 bp); human CRF (sense,
5′-CAACTTTTTCCGCGTGTTGCT-3′ and antisense,
5′-ATGGCATAAGAGCAGCGCTAT-3′, product 360 bp); human CRFR1 primer
(sense, 5′-CACTACCATGTTGCAGTCATC-3′ and antisense,
5′-CGAACATCCAGAAGAAGTTGG-3′, product 300 bp); CRFR2 primer (sense,
5′-TCGTCAACTACCTGCGCCAC-3′ and antisense,
5′-GTCATTAGGATCCTGACGATGT-3′, product 522 bp). Cycling conditions
for ULBP2, CRF, CRFR1 and CRFR2 were 1 min at 94°C, 1 min at 60°C,
and 1 min at 72°C for 32 cycles. β-actin was amplified as a control
using sense primer, 5′-GTGGGGCGCCCCAGGCACCA-3′ and antisense
primer, 5′-CTCCTTAATGTCACGCACGATTTC-3′, product 200 bp. Cycling
conditions for β-actin were 1 min at 94°C, 1 min at 55°C and 1 min
at 72°C for 25 cycles. The PCR products were analyzed by
EtBr-stained 1.2% agarose gel subjected to electrophoresis.
Flow cytometry analysis
Surface antigen FACS analysis was performed to
detect NKG2D ligands on tumor cell lines and to detect NKG2D in NK
cells. Briefly, cells were washed twice with ice-cold
phosphate-buffered saline (PBS) and then incubated with mouse
anti-human MHC class I-related chain A (MICA) monoclonal antibody,
anti-human ULBP monoclonal antibodies (anti-ULBP1, anti-ULBP2 and
anti-ULBP3) or anti-NKG2D antibody (both from R&D Systems) for
30 min on ice. After two washes, cells were incubated with an
appropriate FITC-conjugated secondary antibody diluted in PBS for
30 min on ice. Intracellular FACS analysis was performed to detect
intracellular levels of NKG2D ligands in cervical cancer cells.
Cells were washed twice with ice-cold PBS containing 0.05% BSA and
0.02% sodium azide (PBS-BSA), and fixed in 2% paraformaldehyde in
PBS for 15 min on ice. The cells were then washed once in cold
PBS-BSA and resuspended in PBS containing 0.1% saponin and 0.05%
sodium azide (permeabilization buffer) for 15 min, followed by
incubation with an anti-NKG2D ligand antibody for 30 min on ice.
After two washes, cells were incubated with an appropriate
FITC-conjugated secondary antibody in permeabilization buffer for
30 min on ice. Samples were analyzed using a FACSCalibur flow
cytometer (BD Biosciences).
Cytotoxicity assays
NK cell cytotoxic activity was determined by
calcein-AM assays (24). K562
target cells were washed twice with PBS and incubated with 5 mM
calcein-AM (Molecular Probes) in serum-free RPMI medium for 10 min
at 37°C. Labeled target cells were distributed into U-bottom
microtiter plates at a concentration of 1×104
cells/well. Effector cells were added at various effector cell to
target cell (E:T) ratios in quadruplicate. Target cells in complete
RPMI medium alone were used to determine spontaneous calcein-AM
retention. Maximal lysis was determined by solubilizing three wells
of target cells in lysis buffer (0.1% Triton X-100). After
incubation for 5 h, the assays were analyzed using a fluorescence
reader. Percent specific cytotoxicity was calculated as follows: %
Cytotoxicity = [(retention of experimental well − retention of
spontaneous well)/(retention of maximal lysis well − retention of
spontaneous well)] × 100.
Results
CRF enhances expression of ULBP2 in
cervical cancer cells
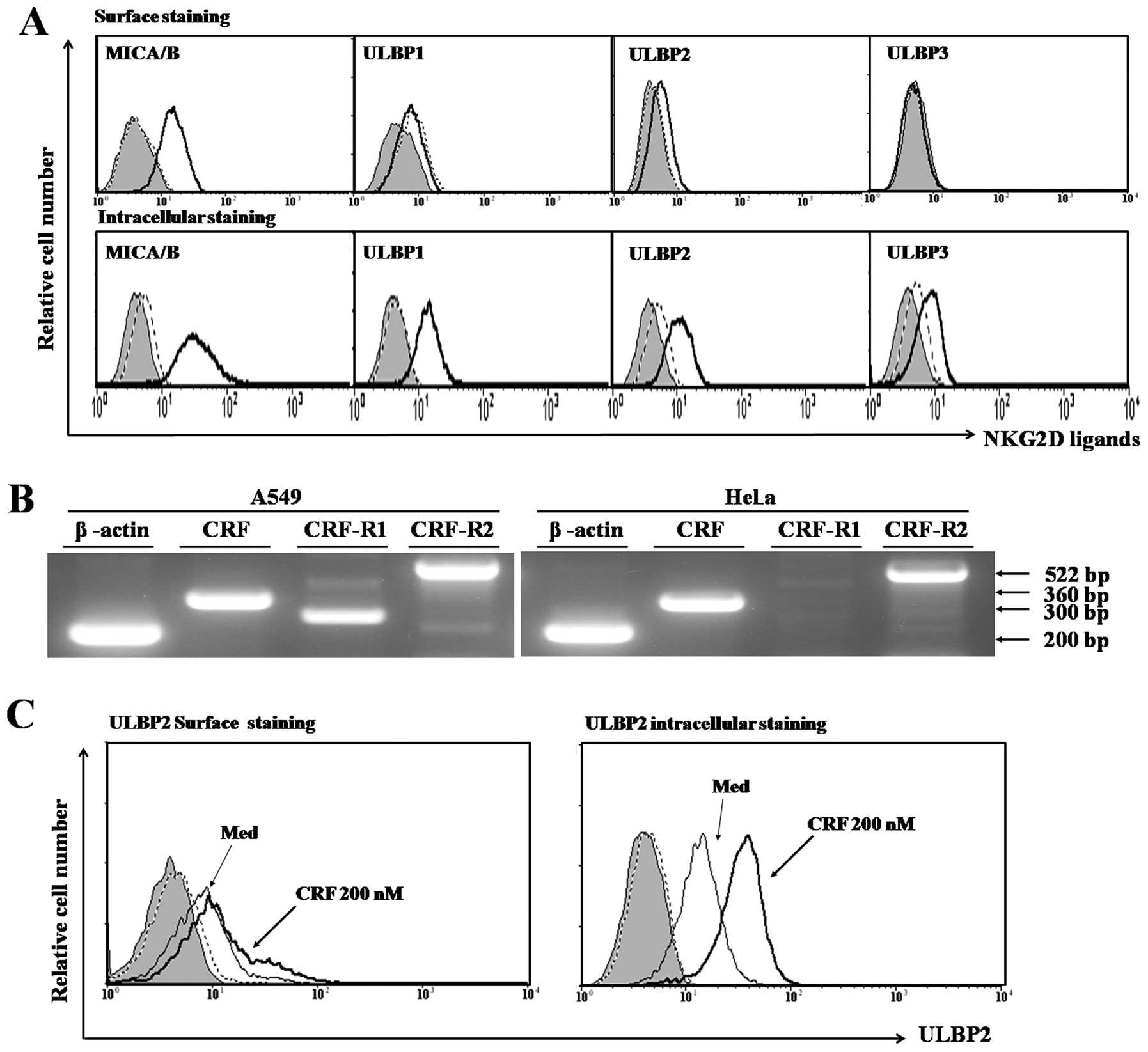
To evaluate expression of NKG2D ligands in cervical
cancer cells, surface and intracellular expression of MICA and ULBP
was analyzed (Fig. 1A). HeLa cells
showed surface expression of MICA and ULBP2, but not ULBP1 and
ULBP3. Intracellular expression of all NKG2D ligands was observed
within HeLa cells by intracellular FACS staining analysis.
We next examined expression of CRF and the CRF
receptor system in HeLa cells. A549 cells, which are known to
express CRF, CRF receptor 1 and 2, were found to be positive for
CRF and CRF receptor system by RT-PCR analysis (Fig. 1B, left panel). CRF and CRF receptor
2 genes were expressed in HeLa cells, but CRF receptor 1 was not
detected by RT-PCR analysis (Fig.
1B, right panel). Several studies have reported that CRF and
CRF receptors are expressed in many human cancers. We hypothesized
that CRF and the CRF receptor system are involved in the response
of stress associated NKG2D ligand expression in cervical cancer
cells. To investigate the ability of CRF to alter NKG2D ligand
expression on cervical cancer cells, we analyzed the intracellular
level of NKG2D ligand in HeLa cells after treatment with CRF. We
found that intracellular level of ULBP2 was strongly induced by CRF
(Fig. 1C). However, other NKG2D
ligands were not induced by CRF treatment (data not shown).
Notably, ULBP2 surface expression was slightly induced in cervical
cancer cells after treatment with CRF (Fig. 1C). These data suggest that
production of CRF in HeLa cells may potentially induce ULBP2
expression.
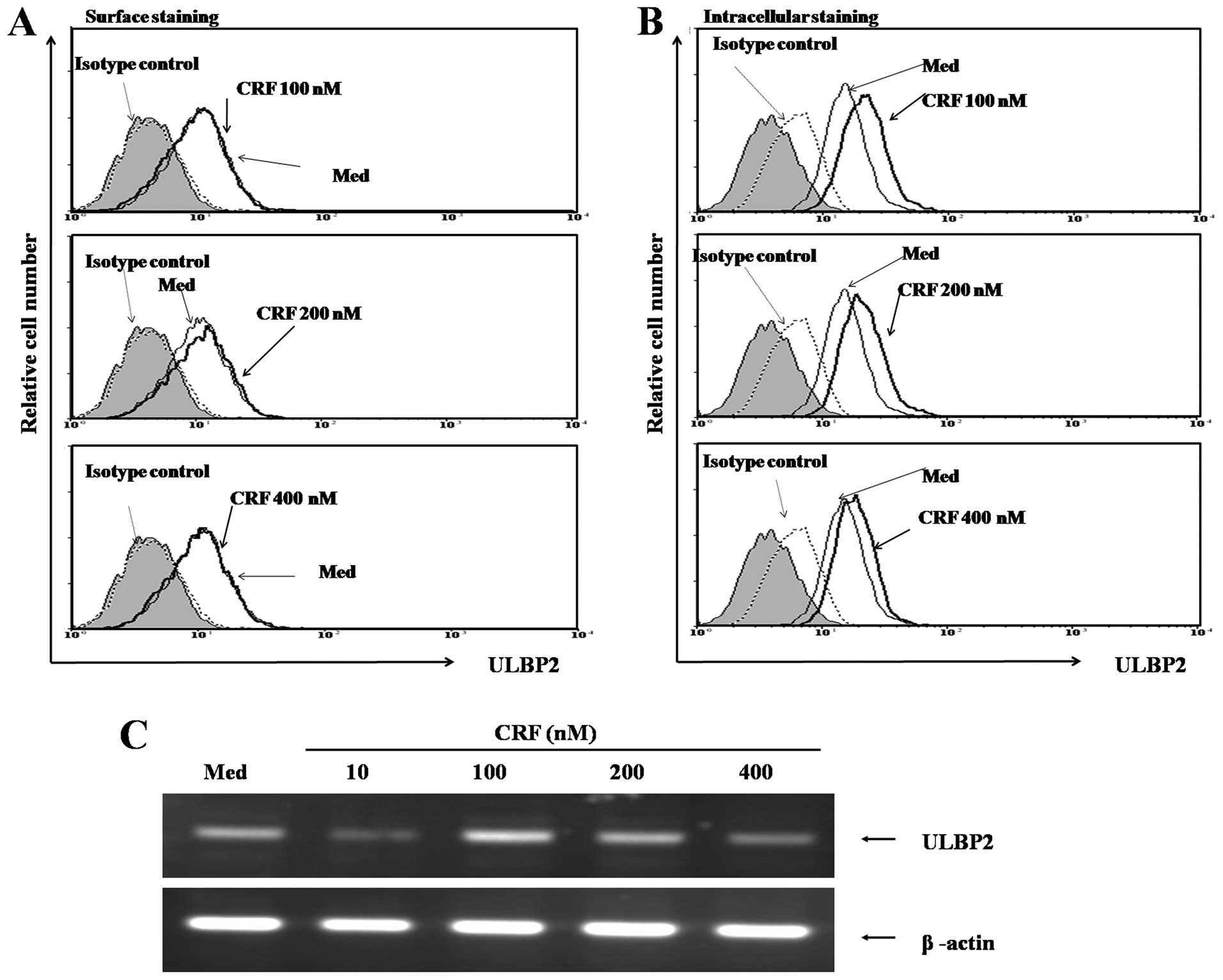
To further confirm our findings, we evaluated the
effects of various concentrations of CRF on ULBP2 expression in
HeLa cells after treatment for 48 h. As shown in Fig. 2A and B, CRF treatment significantly
induced intracellular expression of ULBP2 in a dose-dependent
manner, but surface expression was not induced. As shown in
Fig. 2C, ULBP2 mRNA expression was
markedly induced by CRF in a dose-dependent manner.
Regulation of surface ULBP2 expression in
cervical cancer
We observed that ULBP2 expression was markedly
induced intracellularly following CRF treatment, but was not
induced at the surface level (Fig.
2). We therefore addressed the question of whether CRF-induced
ULBP2 could be expressed at surface level. ULBP 1, 2 and 3 are
known to have a GPI-linked molecule, which can be released by
phosphatidylinositol-specific phosphase C (PI-PLC). Moreover,
Waldhauer and Steinle showed that ULBP2 can be released from tumor
cells by metalloprotease (25).
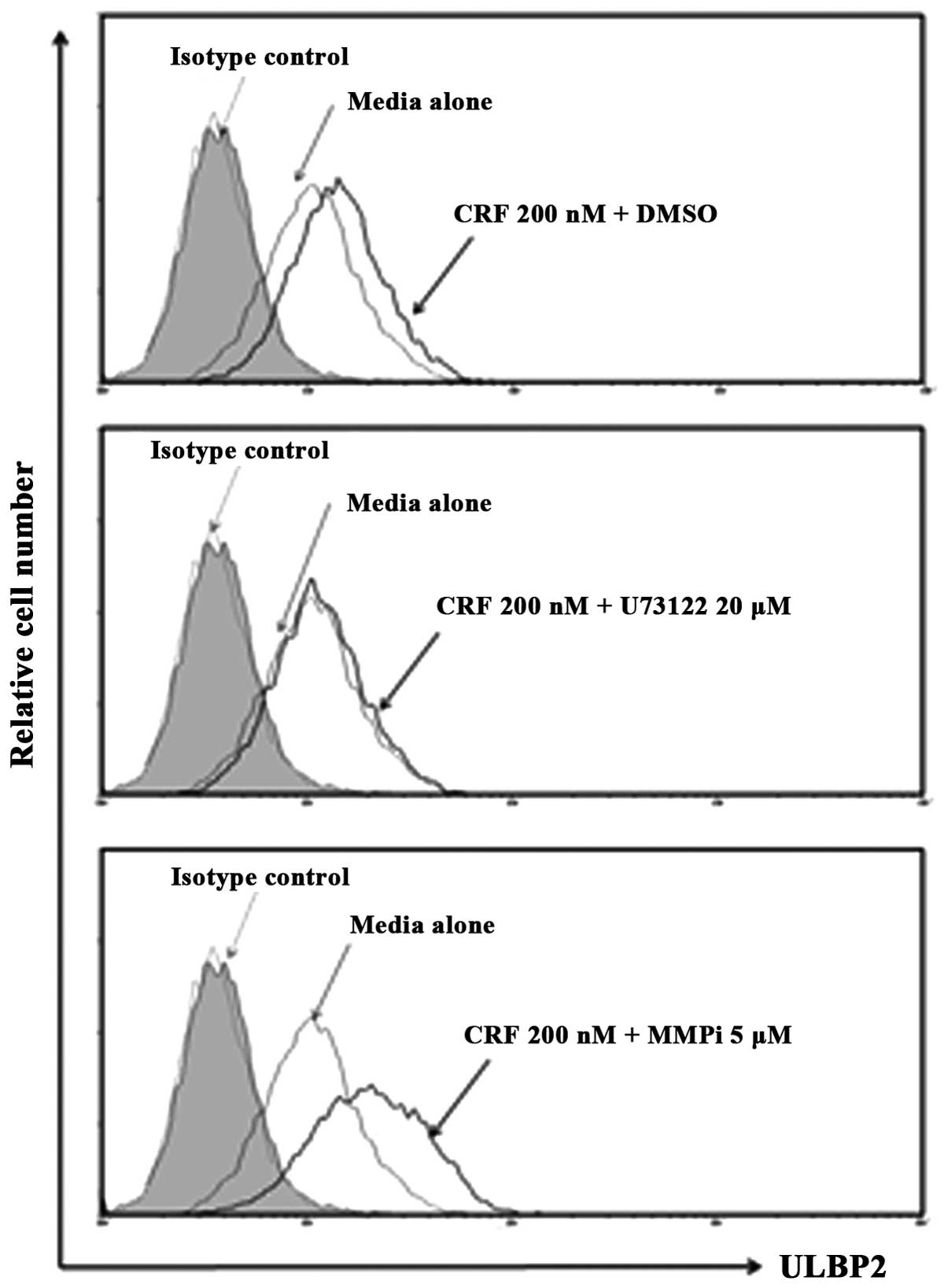
Based on this evidence, we investigated the effect of treatment
with U73122 (PI-PLC inhibitor) and MMPi (pan-metalloprotease
inhibitor) on ULBP2 surface expression following CRF treatment.
After treatment with 5 μM MMPi, CRF-treated HeLa cells showed
strongly enhanced expression of ULBP2 compared with vehicle
control-treated cells (Fig. 3A). In
contrast, HeLa cells treated with PI-PLC inhibitor did not have
enhanced surface expression of ULBP2 following CRF treatment. These
data suggest that PI-PLC is not involved in the release of the
soluble form of ULBP2 on CRF-treated HeLa cells.
Functional ability of soluble ULBP2
secreted from CRF-treated HeLa cells
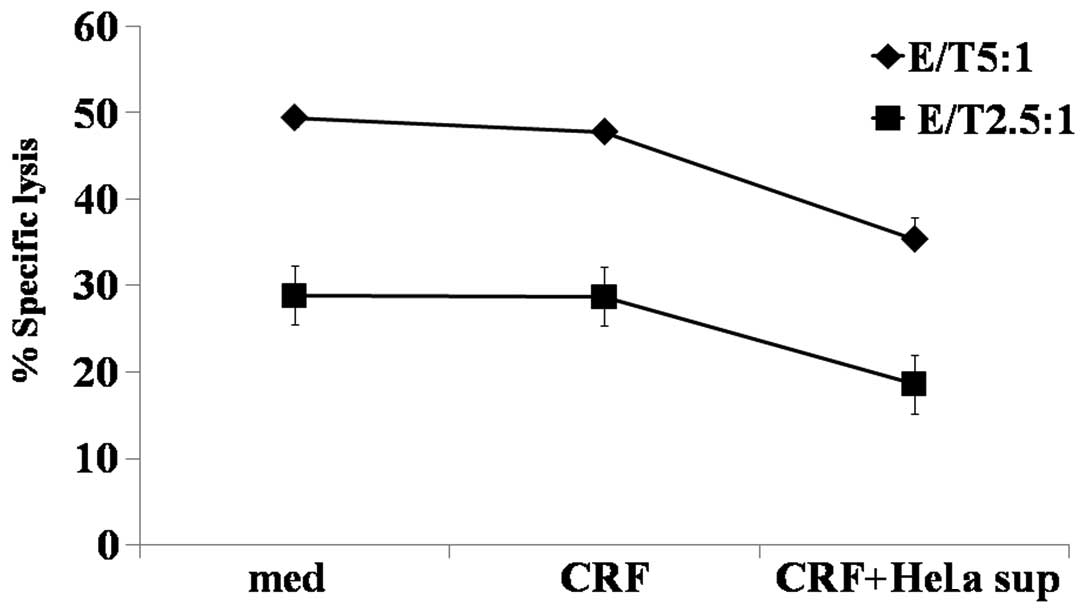
In the present study, we observed that ULBP2 was
expressed on the cell surface of HeLa cells following treatment
with CRF and was released from the cell by metalloprotease. To
determine the functions of soluble ULBP2, we treated NK cells with
culture supernatant from CRF-treated HeLa cells. As shown in
Fig. 4, we found that NK cells
treated with CRF-treated HeLa cell culture supernatant for 48 h
displayed significantly reduced activity in contrast to untreated
NK cell control cells or NK cells treated directly with CRF. These
results suggest that production of CRF from cervical cancer cells
can reduce NK cell function via the NKG2D-mediated pathway.
Discussion
The present study demonstrated the inhibitory
function of CRF in NKG2D-mediated NK killing activity against
cervical cancer cells by inducing release of soluble NKG2D ligand
from cancer cells and suggests the possible role of cancer produced
CRF as an immunomodulator between immune cells and cancer cells.
Corticotrophin-releasing factor (CRF) was originally identified as
a regulator of the stress response in
hypothalamic-pituitary-adrenal axis (26). CRF and the CRF homologues urocortins
(UCN) have previously been detected in many types of cancer, such
as endometrial carcinoma, prostatic carcinoma and pituitary adenoma
(4–6). Reubi et al showed that the CRF
receptors or CRF are expressed in various human cancers (2). Klimaviciute et al reported that
mRNA and proteins of CRF receptors are expressed in human cervix
and CRF is detected in cervical tissue, indicating that the cervix
is a target for the action of CRF (27). Moreover, Minas et al observed
that expression of CRF and CRF receptors detected in ovarian cancer
in situ and corresponding secretion of CRF modulate survival
of ovarian cancer cells (28). This
evidence suggests that CRF and its receptor systems have critical
functions in the regulation of cancer cell progression. In this
study, we observed that CRF and CRF receptor 2 were strongly
expressed in cervical cancer cells (Fig. 1). As shown in Fig. 1, exogenous treatment with CRF can
induce intracellular levels of an NKG2D ligand (ULBP2) in cervical
cancer cells. Moreover, we found that CRF-induced ULBP2 was
released to the surface of cancer cells by metalloprotease
(Fig. 3). Based on these results,
we speculate that CRF produced by cancer cells may be involved in
evasion of cervical cancer through downregulation of NKG2D in NK
cells.
The NKG2D/NKG2D ligand system is particularly
important to immune surveillance of tumors since expression of
NKG2D ligands is induced by carcinogens and stress such as
genotoxic stress (29–31). Human NKG2D ligands consist of two
families: MHC class I-related molecules (MICs) and UL16-binding
proteins (ULBPs) (16). We
originally hypothesized that CRF induces MICA/B in cervical cancer
cells since stress-inducible MICA/B is expressed in certain
epithelial origin tumors following stress signals. In addition,
several studies have revealed that ULBP1 is preferentially induced
over other ULBPs in certain cancer cells (32,33).
In contrast to our expectations, we found that CRF treatment
elevated levels of intracellular ULBP2 in cervical cancer cells.
Moreover, intracellular expression of ULBP2 increased in a
dose-dependent manner following CRF treatment. The ability of CRF
to alter expression of various NKG2D ligands in various cancer
cells may be an interesting topic for further studies.
It is well described that soluble forms of NKG2D
ligands can be used by cancer cells to escape immunosurveillance of
immune effector cells. Specifically, shedding of MICA has been
shown to be an immune escape mechanism of tumors via downregulation
of NKG2D on immune effector cells (34–36).
Some studies have observed soluble forms of ULBP2 in certain cancer
cells that are secreted by phospholipase since the ULBP2 structure
consists of GPI-anchored molecule at the cell surface (37). However, we found that surface
expression of ULBP2 does not increase following treatment with
phospholipase inhibitor (U73122), but treatment with
metalloprotease significantly induced ULBP2 surface expression,
suggesting that ULBP2 cleavage was performed by metalloprotease
(Fig. 3). Some studies have
reported that cleavage of MICA and ULBP2 on cancer cells was
significantly inhibited by metalloprotease inhibitor (20,25,38).
In addition, several studies have also reported that CRF can induce
expression of metalloprotease in cervical tissue (39,40).
These results suggest that proteolytic release of ULBP2 on cancer
cells occurs through CRF-induced metalloprotease during cancer
immune escape. This aspect should be investigated in future
studies.
In conclusion, ULBP2 is expressed and released from
cervical cancer cells by CRF, which regulates NKG2D expression in
NK cells. The stability of cell surface expression and amount of
ULBP2 released seems to be related to NK susceptibility. Mechanisms
to modulate surface expression and soluble ULBP2 may provide
another novel concept for immunotherapy to improve the function of
T and NK cells in killing cervical cancer cells under stress
conditions.
Acknowledgements
This study was supported by a grant from the
National R&D Program for Cancer Control, Ministry for Health,
Welfare and Family Affairs, Republic of Korea (0920060), and by the
Basic Science Research Program through the National Research
Foundation of Korea (NRF) funded by the Ministry of Education,
Science and Technology (2009-0073592).
References
|
1
|
Ciocca DR, Puy LA, Fasoli LC, et al:
Corticotropin-releasing hormone, luteinizing hormone-releasing
hormone, growth hormone-releasing hormone, and somatostatin-like
immunoreactivities in biopsies from breast cancer patients. Breast
Cancer Res Treat. 15:175–184. 1990. View Article : Google Scholar
|
|
2
|
Reubi JC, Waser B, Vale W and Rivier J:
Expression of CRF1 and CRF2 receptors in human cancers. J Clin
Endocrinol Metab. 88:3312–3320. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sato H, Nagashima Y, Chrousos GP,
Ichihashi M and Funasak Y: The expression of
corticotropin-releasing hormone in melanoma. Pigment Cell Res.
15:98–103. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Arcuri F, Cintorino M, Florio P, et al:
Expression of urocortin mRNA and peptide in the human prostate and
in prostatic adenocarcinoma. Prostate. 52:167–172. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Florio P, De Falco G, Leucci E, et al:
Urocortin expression is downregulated in human endometrial
carcinoma. J Endocrinol. 190:99–105. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Iino K, Sasano H, Oki Y, et al: Urocortin
expression in human pituitary gland and pituitary adenoma. J Clin
Endocrinol Metab. 82:3842–3850. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Graziani G, Tentori L, Muzi A, et al:
Evidence that corticotropin-releasing hormone inhibits cell growth
of human breast cancer cells via the activation of CRH-R1 receptor
subtype. Mol Cell Endocrinol. 264:44–49. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Carlson KW, Nawy SS, Wei ET, et al:
Inhibition of mouse melanoma cell proliferation by
corticotropin-releasing hormone and its analogs. Anticancer Res.
21:1173–1179. 2001.PubMed/NCBI
|
|
9
|
Arbiser JL, Karalis K, Viswanathan A, et
al: Corticotropin-releasing hormone stimulates angiogenesis and
epithelial tumor growth in the skin. J Invest Dermatol.
113:838–842. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang Y, Park H, Kim TS, Bang SI and Cho D:
Enhancement of cell migration by corticotropin-releasing hormone
through ERK1/2 pathway in murine melanoma cell line, B16F10. Exp
Dermatol. 16:22–27. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Radulovic M, Hippel C and Spiess J:
Corticotropin-releasing factor (CRF) rapidly suppresses apoptosis
by acting upstream of the activation of caspases. J Neurochem.
84:1074–1085. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Trinchieri G: Biology of natural killer
cells. Adv Immunol. 47:187–376. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Raulet DH: Interplay of natural killer
cells and their receptors with the adaptive immune response. Nat
Immunol. 5:996–1002. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bahram S, Inoko H, Shiina T and
Radosavljevic M: MIC and other NKG2D ligands: from none to too
many. Curr Opin Immunol. 17:505–509. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Groh V, Steinle A, Bauer S and Spies T:
Recognition of stress-induced MHC molecules by intestinal
epithelial γδ T cells. Science. 279:1737–1740. 1998.
|
|
16
|
Raulet DH: Roles of the NKG2D
immunoreceptor and its ligands. Nat Rev Immunol. 3:781–790. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Welte SA, Sinzger C, Lutz SZ, et al:
Selective intracellular retention of virally induced NKG2D ligands
by the human cytomegalovirus UL16 glycoprotein. Eur J Immunol.
33:194–203. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Groh V, Wu J, Yee C and Spies T:
Tumour-derived soluble MIC ligands impair expression of NKG2D and
T-cell activation. Nature. 419:734–738. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pende D, Rivera P, Marcenaro S, et al:
Major histocompatibility complex class I-related chain A and
UL16-binding protein expression on tumor cell lines of different
histotypes: analysis of tumor susceptibility to NKG2D-dependent
natural killer cell cytotoxicity. Cancer Res. 62:6178–6186.
2002.
|
|
20
|
Waldhauer I, Goehlsdorf D, Gieseke F, et
al: Tumor-associated MICA is shed by ADAM proteases. Cancer Res.
68:6368–6376. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Salih HR, Antropius H, Gieseke F, et al:
Functional expression and release of ligands for the activating
immunoreceptor NKG2D in leukemia. Blood. 102:1389–1396. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Salih HR, Holdenrieder S and Steinle A:
Soluble NKG2D ligands: prevalence, release, and functional impact.
Front Biosci. 13:3448–3456. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Song H, Kim J, Cosman D and Choi I:
Soluble ULBP suppresses natural killer cell activity via
down-regulating NKG2D expression. Cell Immunol. 239:22–30. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lichtenfels R, Biddison WE, Schulz H, Vogt
AB and Martin R: CARE-LASS (calcein-release-assay), an improved
fluorescence-based test system to measure cytotoxic T lymphocyte
activity. J Immunol Methods. 172:227–239. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Waldhauer I and Steinle A: Proteolytic
release of soluble UL16-binding protein 2 from tumor cells. Cancer
Res. 66:2520–2526. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bale TL and Vale WW: CRF and CRF
receptors: role in stress responsivity and other behaviors. Annu
Rev Pharmacol Toxicol. 44:525–557. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Klimaviciute A, Calciolari J, Bertucci E,
et al: Corticotropin-releasing hormone, its binding protein and
receptors in human cervical tissue at preterm and term labor in
comparison to non-pregnant state. Reprod Biol Endocrinol. 4:292006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Minas V, Rolaki A, Kalantaridou SN, et al:
Intratumoral CRH modulates immuno-escape of ovarian cancer cells
through FasL regulation. Br J Cancer. 97:637–645. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cerwenka A and Lanier LL: Ligands for
natural killer cell receptors: redundancy or specificity. Immunol
Rev. 181:158–169. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Diefenbach A and Raulet DH: Strategies for
target cell recognition by natural killer cells. Immunol Rev.
181:170–184. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lanier LL: NKG2D. J Biol Regul Homeost
Agents. 17:338–340. 2003.
|
|
32
|
López-Soto A, Quiñones-Lombraña A,
López-Arbesú R, López-Larrea C and González S: Transcriptional
regulation of ULBP1, a human ligand of the NKG2D receptor. J Biol
Chem. 281:30419–30430. 2006.PubMed/NCBI
|
|
33
|
Rohner A, Langenkamp U, Siegler U,
Kalberer CP and Wodnar-Filipowicz A: Differentiation-promoting
drugs up-regulate NKG2D ligand expression and enhance the
susceptibility of acute myeloid leukemia cells to natural killer
cell-mediated lysis. Leuk Res. 31:1393–1402. 2007. View Article : Google Scholar
|
|
34
|
Raffaghello L, Prigione I, Airoldi I, et
al: Downregulation and/or release of NKG2D ligands as immune
evasion strategy of human neuroblastoma. Neoplasia. 6:558–568.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Salih HR, Rammensee HG and Steinle A:
Cutting edge: down-regulation of MICA on human tumors by
proteolytic shedding. J Immunol. 169:4098–4102. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu JD, Higgins LM, Steinle A, Cosman D,
Haugk K and Plymate SR: Prevalent expression of the
immunostimulatory MHC class I chain-related molecule is
counteracted by shedding in prostate cancer. J Clin Invest.
114:560–568. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Onda H, Ohkubo S, Shintani Y, et al: A
novel secreted tumor antigen with a
glycosylphosphatidylinositol-anchored structure ubiquitously
expressed in human cancers. Biochem Biophys Res Commun.
285:235–243. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fernández-Messina L, Ashiru O, Boutet P,
et al: Differential mechanisms of shedding of the
glycosylphosphatidylinositol (GPI)-anchored NKG2D ligands. J Biol
Chem. 285:8543–8551. 2010.PubMed/NCBI
|
|
39
|
Dubicke A, Akerud A, Sennstrom M, et al:
Different secretion patterns of matrix metalloproteinases and IL-8
and effect of corticotropin-releasing hormone in preterm and term
cervical fibroblasts. Mol Hum Reprod. 14:641–647. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li W and Challis JR:
Corticotropin-releasing hormone and urocortin induce secretion of
matrix metalloproteinase-9 (MMP-9) without change in tissue
inhibitors of MMP-1 by cultured cells from human placenta and fetal
membranes. J Clin Endocrinol Metab. 90:6569–6574. 2005. View Article : Google Scholar
|