Introduction
Prostate cancer (PrC) is a highly prevalent
oncologic pathology in most countries throughout the world
(1). During its early stages this
disease is usually asymptomatic and exhibits slow progression,
which carries the risk of having it diagnosed at an advanced stage.
Research indicates a direct correlation between the appearance of
symptoms and the spreading of the cancer or metastasis. This late
diagnosis may decrease the treatment options available for patients
and also the chances of recovery (2).
Due to the above factors and in light of the
aggressive nature of this disease in its late stages, it has become
critical to search for more effective tools with which to detect
PrC while it is still in its early stages, and for better treatment
options. These two aspects would greatly improve the quality of
life and the overall survival expectancy of these patients
(3,4).
Regarding the molecular pathogenesis of PrC, it has
been observed that certain genetic alterations may provoke the
transformation of normal prostatic cells into cancerous cells. In
this context, it has been reported that certain mutations in key
specific genes such as PTEN, TP53, E-cadherin and β-catenin
(5) are important in this
transformation process. In addition, it has been demonstrated that
several growth factors, such as insulin growth factor I (IGF-I),
transforming growth factor α and β (TGFα or β) and members of the
fibroblast growth factor (FGF) family, may be involved in the
proliferation and metastasis of PrC (6).
The molecular factors involved in the development of
PrC are various and as the transformation process in the cell
evolves, it acquires a malignant phenotype with the ability to
invade and generate metastasis in different parts of the body, but
with a preference to form metastatic lesions in the bones (5,6). For
this reason, many compounds have been used to try and control the
progression of prostate cancer. It has been suggested that
marijuana (Cannabis sativa) through some of its active
compounds could be involved in halting tumoral growth and thus
delaying its progression to more advanced and aggressive stages
(7–9).
Marijuana acts in the organism as a psychoactive
agent through the production of its active component, the
cannabinoid (−)-Δ9-tetrahidrocanabinol (or THC), which
is an aromatic terpenoid compound with a very low solubility in
water. Two different cannabinoid receptors have been described,
characterized and cloned: cannabinoid receptor 1 (or CB1),
originally found in the brain, and cannabinoid receptor 2 (or CB2),
which was first described in the spleen. Both receptors are part of
the superfamily of G protein-coupled receptors (10,11).
Other than its exogenous ligands such as THC, the
organism can produce similar compounds that have been termed as
endocannabinoids, which are capable of modulating several
physiological mechanisms through the CB1 and CB2 membrane receptors
(12), although it has also been
reported recently that they may also act in an independent receptor
manner (13).
Endocannabinoids are molecules that are derived from
unsaturated fatty acids acting as endogenous ligands for
cannabinoid receptors CB1 and CB2 and originate in the plasma
membrane from phospholipids in response to a rise in the
intracellular concentrations of calcium. These endogenous ligands
bind to the CB1 and CB2 receptors with great affinity and
participate in many biological processes in the immune,
respiratory, circulatory and reproductive systems (9,14–17).
Endocannabinoids are then eliminated in a two step process: first,
an intracellular accumulation, followed by an enzymatic
metabolization by an enzyme that belongs to the serine hydrolase
family, the fatty acid amide hydrolase (or FAAH) or by the monoacyl
glycerol lipase (a soluble serine hydrolase enzyme). These two
enzymes are the main proteins in charge of degrading
endocannabinoids (18).
On a cellular level, it has been found that
endocannabinoids may modulate cell proliferation, viability and
differentiation. This evidence suggests that endocannabinoids may
also be involved in controlling the growth and transformation of
tumor cells (8,19,20).
Regarding this, evidence shows that endocannabinoids may inhibit
the growth of several types of tumors through the inhibition of
proliferative pathways such as adenylate cyclase (21) and protein kinase A (22), arrest of the cell cycle by induction
of p27 (23), downregulation of the
EGF receptor (EGF-R) and other molecules related to growth pathways
such as the nerve growth factor receptor (NGF-R), the vascular
endothelial growth factor (VEGF) and prolactine (24). Likewise, when tumor cells are
treated with specific antagonists for endocannabinoids, the
invasive ability of tumors increases. The great advantage that the
use of endocannabinoids may bring to the battle against prostate
cancer is that it has been demonstrated that the receptors for
these molecules are substantially overexpressed in cancerous
prostatic cells when compared to normal, healthy prostate tissues
(9,25).
There are two classes of endocannabinoids. Among
those that are derived from fatty acids, anandamide is the most
studied and was the first to be described. Other members of this
family include N-oleoylethanolamide and N-palmitoylethanolamide,
which have shown strong dietetic effects independent of CB1 or CB2
activation. The second class of endocannabinoids are those bound to
glycerol, of which 2-arachidonoyl glycerol (2-AG) is the most
commonly used in research due to its demonstrated effects on cancer
cells. Another member of this family is 2-arachidonyl glyceryl
ether, the endogenous relevance of which is currently being studied
(9).
Due to the previous effects noted in other types of
cancer and increased knowledge of anandamide (and its synthetic
analog methanandamide) and 2-AG, further investigation of the
effects of these endocannabinoids for the treatment of PrC is
warranted. Recently, it was reported that an increment in
intracellular levels of 2-AG inhibited the invasive ability of the
PrC cell lines PC3 and DU-145 by a mechanism involving the
activation of the CB1 receptor and through the inactivation of
protein kinase A (22). In the same
manner, the direct participation of anandamide (Ana) in decreasing
the proliferative action of EGF in cell lines has been proven.
Following treatment with anandamide in DU-145 and LNCaP cells, a
downregulating effect on the expression levels for the receptor of
the EGF growth factor accompanied by a proliferative arrest in the
G1 phase of the cell cycle and a rise in the levels of apoptosis
and necrosis in the cells were found (26–28).
Not with standing these results, information
concerning the signaling pathways mediating these effects in PrC is
still scarce and somewhat contradictory in nature. Some information
has been reported in other types of cancer, for example, the
activation of the AKT pathway in astrocytoma cells after treatment
with THC or Ana (29), activation
of the ERK pathway when using THC in glioma cell lines (30) or the activation of the JNK pathway
when using endocannabinoids in different types of nerve cells that
express the CB1 receptor (31).
To date, all of the studies using endocannabinoids
have been made in cell lines. The objective of the present study
was to analyze the effect of endocannabinoids, not only on cell
lines, but also on primary cultures of PrC and the signaling
pathways involved in order to obtain a better understanding of the
possible effects following treatment with these molecules against
prostate cancer.
Materials and methods
Materials
Endocanabinoids anandamide (Ana) and 2-AG were
purchased from Calbiochem (San Diego, CA, USA) (cat. nos. 172100
and 181251, respectively) and methanandamide (Me) was purchased
from Biomol (Plymouth Meeting, PA, USA) (cat. no. FA-021). CB1
receptor antagonist, SR141716, was purchased from Sanofi
(Montpellier, France).
Primary antibodies for the CB1 and CB2 receptors
were obtained from Cayman Chemical (Ann Arbor, MI, USA) (cat. nos.
10006590 and 101550, respectively), and caspase-3 and Bcl-2 from
Cell Signaling Technology, Inc. (Danvers, MA, USA) (cat. nos. 9661
and 2876, respectively). Phospho-p44/42 MAPK (Erk1/2) was purchased
from Cell Signaling Technology, Inc. (Danvers) (cat. no. 9101) and
p-Akt 1/2/3 (Thr308)-R from Santa Cruz Biotechnology, Inc. (Dallas,
TX, USA) (cat. no. 16646-R). Anti-mouse and anti-rabbit secondary
peroxidase-conjugated antibodies were obtained from Jackson Immuno
Research (West Grove, PA, USA) (cat. nos. 115-035-003 and
111-035-003, respectively). Anti-rabbit FITC-conjugated secondary
antibodies were obtained from Jackson Immuno Research (cat. no.
305-095-003). For immunohistochemistry the subsequent kits were
used: Histostain®-Plus Bulk kit (cat. no. 85-8943) and
the DAB-Plus Substrate kit (cat. no. 00-2020) (both from
Invitrogen, Carlsbad, CA, USA). Annexin V-FITC Apoptosis Detection
kit was obtained from BD Pharmingen (Franklin Lakes, NJ, USA) (cat.
no. 556547).
Methods
Immunohistochemistry
Formalin-fixed and paraffin-embedded prostate
specimens were obtained from the archives of the Pathological
Anatomy Service, Clinic Hospital of the University of Chile, with
the corresponding authorization. All samples were evaluated by an
expert pathologist (I.G.) and grouped as following: benign prostate
hyperplasia (BPH) as a non-malignant control and PrC samples of
high and low Gleason score. Tumor and control samples were cut into
5-μm sections, mounted on silane-treated slides, deparaffinized in
xylene and dehydrated in a series of ethanol solutions with
increasing ethanol content up to 100%. The sections were washed
with phosphate-buffered solution (PBS) (5 min, 3 times). The
sections were incubated in a steam bath for 10 min at 95–100°C in
retrieval buffer (10 mM citrate buffer, pH 6.0). After cooling
down, the samples were incubated with 3% H2O2
for 10 min, in order to inhibit the activity of endogenous
peroxidase. Then, the sections were washed with PBS 2–3 times,
incubated in blocking solution (PBS 2% BSA) at room temperature for
1 h and washed again with PBS 2–3 times. Primary antibodies were
added and the sections were incubated for 1 h at 37°C or overnight
at 4°C. After incubation with the primary antibodies, a secondary
antibody was added to the sections for 30 min at 37°C. Then, the
samples were washed 3 times with PBS for 5 min. Next, the sections
were stained with the strepavidin-biotin system followed by
counterstaining with hematoxylin. Finally, all of the specimens
were sealed with neutral glue.
Cell cultures
PrC primary cell cultures were obtained from fresh
samples of patients with prostate adenocarcinoma. The protocol for
obtaining the sample and its use was approved by the Universidad de
Chile Bioethics Committee including the required informed consent
(DI MULT 05/36-2 project Universidad de Chile). The human prostate
carcinoma cell line (PC3) was obtained from the American Type
Culture Collection (Rockville, MD, USA). PC3 cells were cultured in
Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10%
fetal bovine serum and 1% antibiotic penicillin and streptomycin.
PrC primary cell cultures were grown in DMEM supplemented with 7%
fetal bovine serum and 1% antibiotic penicillin and streptomycin.
All the cells were maintained under standard cell culture
conditions at 37°C in 5% CO2 in a humid environment
(32).
Cell treatments
Anandamide (dissolved in DMSO), 2-arachidonoyl
glycerol (2-AG; dissolved in DMSO) and methanandamide (Me;
dissolved in methanol) were used for the treatment of cells. The
final concentrations of DMSO and the methanol used were proven
harmless for the treated cells. For the dose-dependent studies, the
cells were treated with Ana, 2-AG and Me at final concentrations of
2.5, 5.0 and 10.0 μM for 48 h. For the rest of the experiments 5.0
μM was used since it showed the best concentration/effect ratio. To
establish the role of CB1 and CB2 receptors in the endocannabinoid
effects, cells were treated with 20 μM of CB1 antagonist SR141716
(diluted in DMSO) for 30 min at 37°C in the absence of light.
Afterwards, the antagonist was removed and the cells were washed
with PBS. Finally, the cells were treated with the different
endocannabinoids under the same conditions as explained above.
Cell viability
The effect of endocannabinoids on cell viability was
determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl
tetrazoliumbromide (MTT) assays. The cells were plated at
5×103/well in 200 μl of complete culture medium
containing 2.5, 5.0 and 10.0 μM concentrations of Ana, 2-AG or Me
in 96-well microtiter plates for 48 h at 37°C in a humidified
chamber. Each condition was repeated 5 times. After incubation, MTT
reagent (100 μl 5 mg/ml in PBS) was added to each well, and the
microplates were incubated for 3 h at 37°C in the dark. The MTT
solution was removed from the wells by aspiration and the crystals
were dissolved in DMSO (150 μl). Absorbance was recorded on a
microplate reader (Mod. DNM-9602; Perlong, Beijing, China) at a 550
nm wavelength. The effect of the three endocannabinoids on growth
inhibition was assessed as the percentage of inhibition in regards
to the untreated controls (100%).
Western blot analysis
Following the treatment of cells with
endocannabinoids at a concentration of 5 μM for 48 h, the medium
was aspirated and the cells were washed with PBS and then
trypsinized and centrifuged at 2,500 rpm for 5 min. The resulting
pellet was resuspended in a lysis buffer with a protease inhibitor
cocktail. Later, the cells were scraped and the lysate was
collected in a microfuge tube and passed through a syringe to break
up the cell aggregates. The lysate was cleared by centrifugation at
13,500 × g for 15 min at 4°C, and the supernatant was discarded and
the protein pellet was collected for protein quantification using
the Bradford method at 570 nM in a Rayleigh spectrophotometer
(UV-1600 model). For western blot analysis, 40 μg of protein was
resolved over 10% polyacrylamide gels, with a molecular weight
standard (cat. no. 161-0374; Pierce, Rockford, IL, USA), and
electro-transferred onto a nitrocellulose membrane (cat. no.
162-0115; Bio-Rad, Berkeley, CA, USA). The nonspecific sites on the
membranes were blocked by being incubated with a blocking buffer
for 1 h at room temperature. Then, the membranes were incubated
with the corresponding primary antibody in blocking buffer,
overnight at 4°C, followed by incubation with anti-mouse or
anti-rabbit peroxidase-conjugated secondary antibodies and detected
by chemiluminescence (EZ-ECL kit, cat. no. 20-500-120; Biological
Industries, Beit-Haemek, Israel). The bands were scanned and then
analyzed using the scientific software program Un-Scan-It (Silk
Scientific Corporation, Orem, UT, USA).
Quantification of cell cycle distribution
by flow cytometry
The cells were grown at a 106 density in
culture dishes and were treated with endocannabinoids at a 5.0 μM
concentration for 48 h. Then, the cells were trypsinized, washed
with PBS and fixed in cold ethanol (70% v/v) and stored in
cytometry tubes at −20°C. At the time of the analysis, the cells
were centrifuged and resuspended in a labeling solution (propidium
iodide 0.5 mg/ml and RNase 100 μg/ml). The labeled cells were
incubated for 30 min at 37°C in the dark and then analyzed in a
FACScan cytometer (Becton-Dickinson, Franklin Lakes, NJ, USA). Data
analysis was assessed using WinMD1 version 2.8 software.
Annexin V assay
Cells were grown at a 106 density and
treated with endocannabinoids at a 5.0 μM concentration for 6 h.
After the treatment, the cells were trypsinized, washed with cold
PBS and then centrifuged at 2,500 rpm for 5 min. The supernatant
was discarded and the pellet was resuspended in 1 ml of cold PBS.
The cells were counted with a Neubauer chamber and 105
cells were transferred to a cytometry tube. Then, the cells were
processed and labeled according to the BD Pharmingen Annexin V-FITC
Apoptosis Detection kit that was used for this assay. The labeled
cells were analyzed in a FACScan cytometer (Becton-Dickinson). Data
analysis was assessed using WinMD1 version 2.8 software.
Statistical analysis
Data are expressed as mean ± SD. The significance
between the control and treated cells was calculated using an
unpaired t-test for P-value. P<0.05 was considered to indicate a
statistically significant difference. The GraphPad Quick Calcs
program was used.
Results
Immunohistochemistry of CB1 and CB2
receptors in prostatic tissue
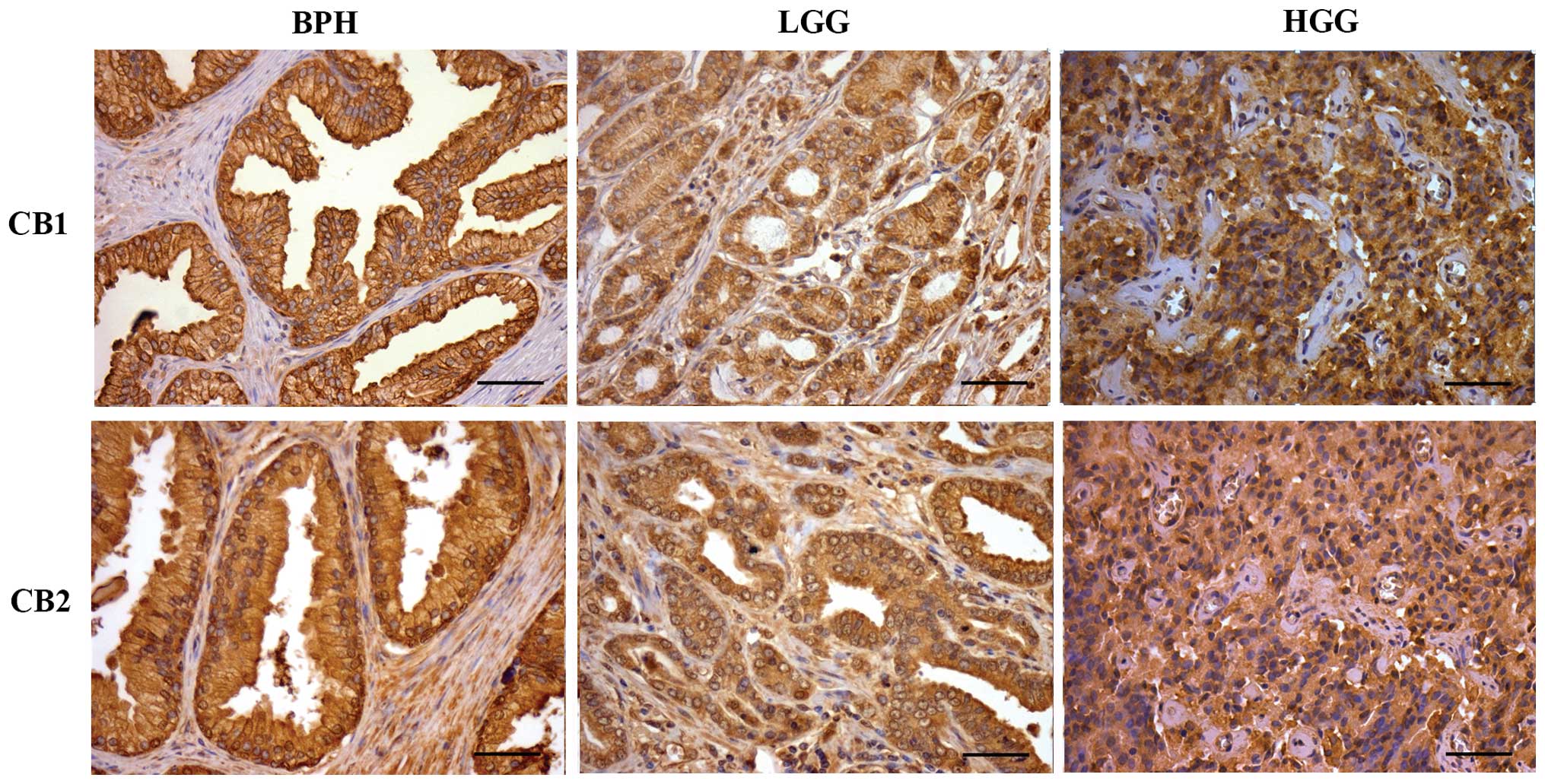
The immunodetection of CB1 and CB2 receptors in PrC
and BPH tissues is shown in Fig. 1.
For both receptors, the staining pattern was similar. In BPH, the
staining was strong in the apical cell membrane and mild in the
cytoplasm. Low Gleason grade (LGG) and high Gleason grade (HGG)
samples showed a similar pattern with homogeneous and diffuse
cytoplasmic staining, but the intensity of the staining was
stronger in the HGG samples. Exceptionally, there were positive
nuclei of isolated lymphocytes.
Additionally, the presence of CB1 and CB2 receptors,
in the PC3 cells and BPH and PrC primary cultures, was evaluated by
immunocytochemistry and western blot analysis. Both types of cells
showed the presence of the receptors. In addition, the receptors
appeared to be homogeneously distributed throughout the cell
surface of PC3, BPH and PrC cells in a similar way (data not
shown).
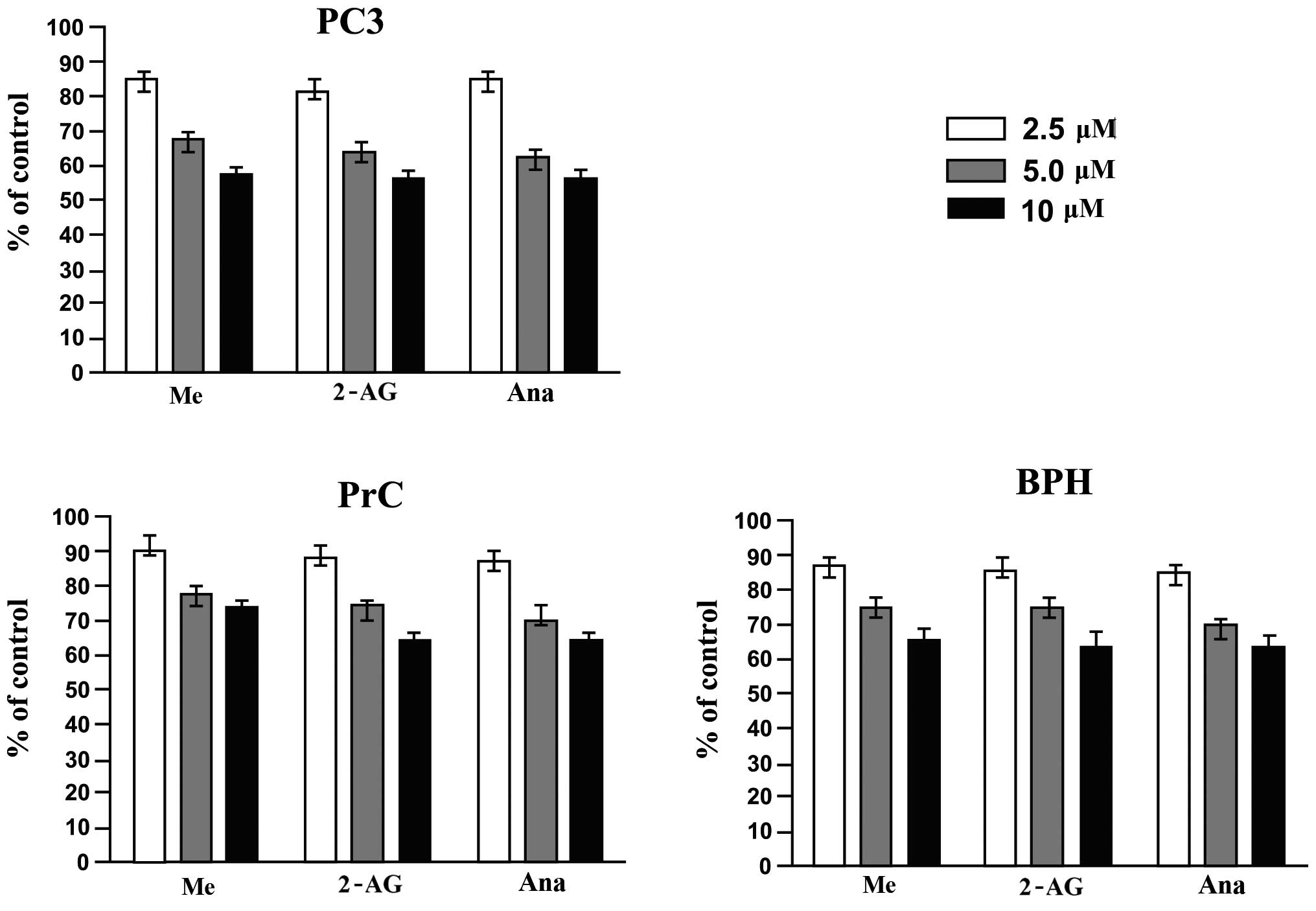
Effect of endocannabinoids on the cell
viability of PC3 cells and primary cultures of BPH and PrC
To assess the cell viability response of the PC3
cell line and the primary cultures of PrC and BPH and the primary
cultures to the different endocannabinoids, MTT assay was employed.
Fig. 2 shows that treatment of PC3
cells and primary cultures of PrC and BPH for 48 h with the
different endocannabinoids (at 2.5, 5.0 and 10 μM concentrations)
significantly decreased the viability of the cells (P<0.05).
Even more, the effect was proven to be dose-dependent and PC3 cells
were more sensitive to the treatments than both primary cultures.
Among the primary cultures, BPH cultures proved to be more
sensitive to the treatments than the PrC cultures.
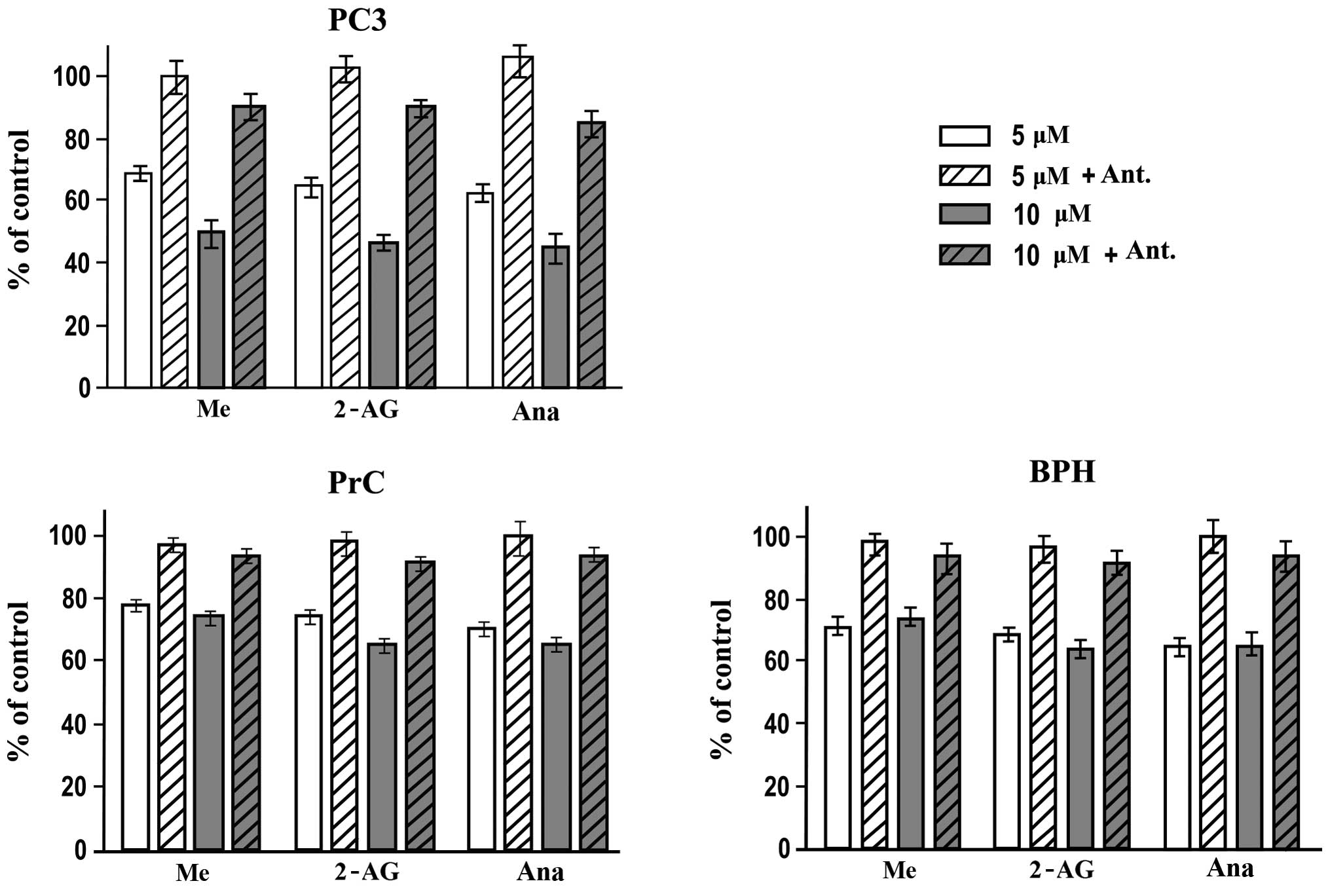
Participation of CB1 and CB2 receptors in
the effect by endocannabinoids on the viability of PC3 cells and
primary cultures of BPH and PrC
To study the role of CB1 and CB2 receptors in the
endocannabinoid-mediated suppression of viability of the different
cell cultures, we evaluated the effect of a CB1 antagonist SR141716
using the MTT assay. Cells were incubated with 20 μM SR141716 for
30 min (which had no effect on the cell viability) and then were
treated normally with the different endocannabinoids (Fig. 3). The pretreatment with the
antagonist showed a significant protective effect on both PC3 cells
and primary cultures of BPH and PrC. Only by using a 10 μM
concentration of the endocannabinoids were we able to observe a
small effect on the cell viability again (P<0.05) but only in
the PC3 cells. The effect of the treatments on both primary
cultures remained the same even at the highest dose used. These
results showed that the activation of the CB1 receptors was
required in the endocannabinoid-mediated growth inhibition process
or cell death noted in PC3 cells and primary cultures of BPH and
PrC when treated with endocannabinoids.
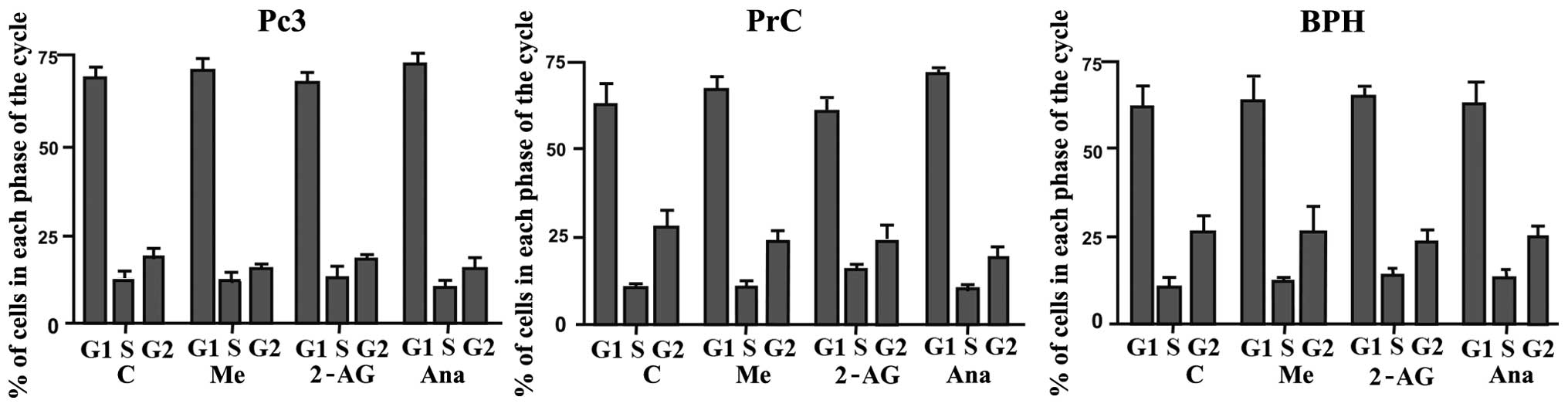
Effect of endocannabinoids on the cell
cycle distribution of PC3 cells and primary cultures of BPH and
PrC
Previous research from our group showed a decrease
in the expression of PCNA (a cell proliferation marker) in response
to endocannabinoid treatment (data not shown). Due to these
results, we hypothesized that the effect on cell viability
demonstrated by the endocannabinoids on the different cell cultures
could be produced through induction of apoptosis by cell cycle
arrest. PC3 cells and primary cultures of BPH and PrC were treated
with the different endocannabinoids for 48 h at a 5 μM
concentration. As shown in Fig. 4
there were no significant variations in the distribution of cells
throughout the cell cycle when comparing control untreated cells
with PC3 cells and PrC and BPH cultures that were treated with the
different endocannabinoids (P>0.05).
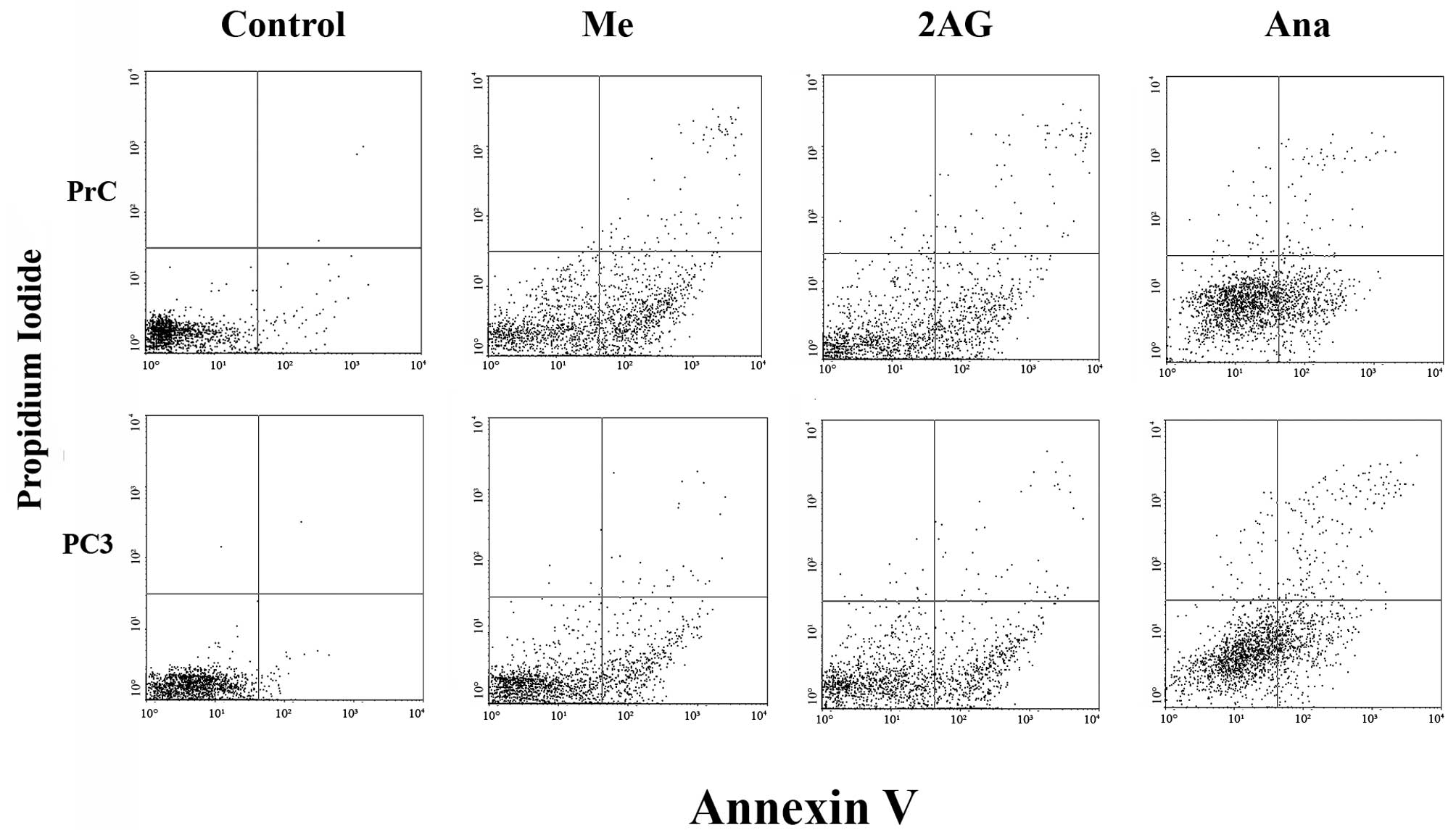
Flow cytometric analysis of FITC Annexin
V staining in the PC3 cells and primary cultures of PrC
To analyze the action of endocannabinoids on the
different cultures of PrC cells and the possibility of a
proapoptotic effect we used a FITC Annexin V assay. As shown in
Fig. 5, the number of cells in the
apoptotic state was higher in all of the treated cell cultures when
compared with the untreated ones, while the number of necrotic
cells remained statistically without variation (P>0.05). This
effect was noted in both types of cells, but it was greater in the
PC3 cells than that noted in the primary cultures of PrC. In
addition, none of the different endocannabinoids showed a higher
effect when compared with the others in the cell lines, yet the
results varied among the different endocannabinoids in the primary
cultures, with Me showing a higher effect than the other two
agents. These results suggest that the effect of endocannabinoids
on PC3 cells and primary cultures of PrC may be caused by the
activation of the apoptotic pathway.
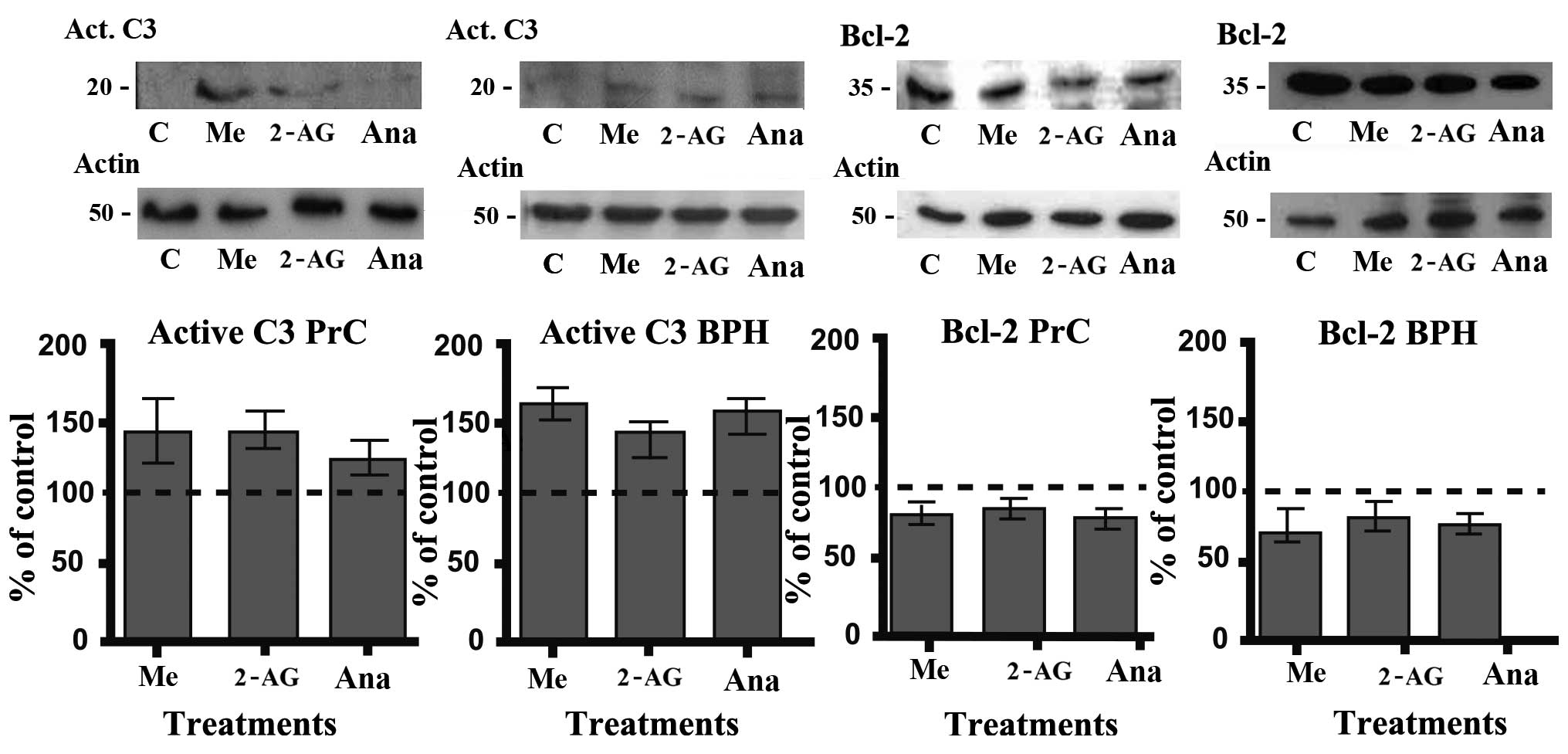
Effect of endocannabinoids on the
expression of active caspase-3 and Bcl-2 in primary cultures of PrC
and BPH
The results of the Annexin V assay indicated that
endocannabinoids may exert their action through the activation of
the apoptotic pathway. To corroborate this we analyzed their effect
on the expression of important proteins of the classical apoptotic
pathway such as the active form of caspase-3 and Bcl-2. Cells were
treated for 48 h with the different endocannabinoids at a 5 μM
concentration. After the treatments (Fig. 6), there was a significant increase
in the level of active caspase-3 and a consequent decrease in the
level of Bcl-2 (both in PrC and BPH cultures), which corresponds
with an activation of the classical apoptotic pathway.
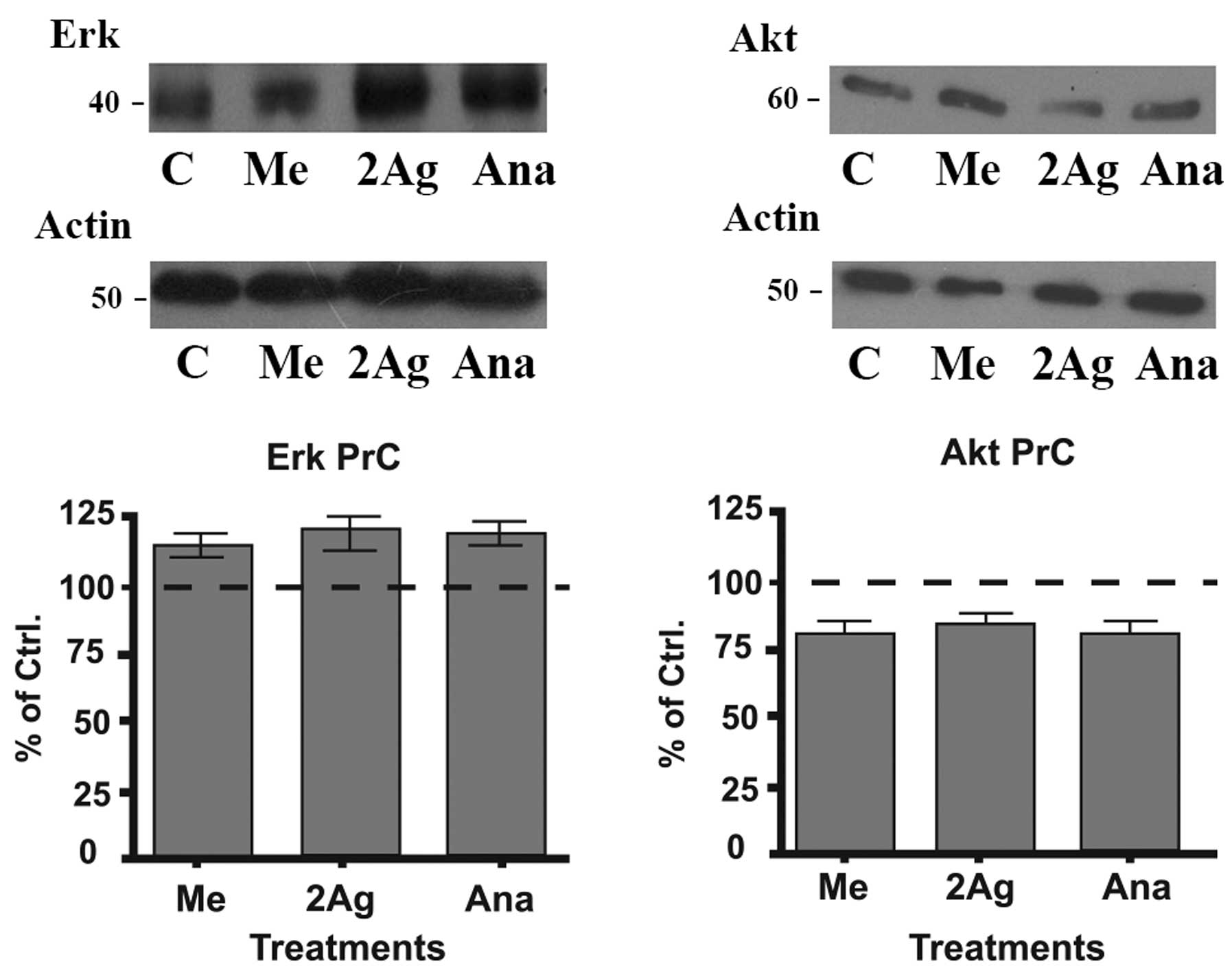
Effect of endocannabinoids on the
expression of Erk and Akt in primary cultures
It has been shown that the Erk and Akt signaling
pathways play a very important role in regulating the cellular
response to proliferative signals and that mutations in these
pathways are often observed in various types of cancer (26). Therefore, we analyzed the effect of
endocannabinoids on the expression of these proteins to test the
possibility of their involvement in the apoptotic process as noted
in the above experiments. Cells were treated with different
endocannabinoids for 48 h at a 5 μM concentration. After the
treatments, we observed that Erk protein expression was augmented
in the treated cells when compared to the control-untreated cells
(Fig. 7). In contrast, the levels
of Akt decreased following all the treatments in contrast to the
increase noted in the Erk expression. These results indicate that
Erk activation/Akt downregulation may be involved in the suppresive
effect that endocannabinoids have on the cell viability of PrC
cells.
Discussion
Prostate cancer has become the most commonly
diagnosed cancer in men and is one of the most threatening diseases
in Western countries. Although there are therapies that have been
proven effective, such as androgen deprivation, these treatment
strategies are not able to eliminate all tumor cells. In addition,
current chemotherapy treatments cause many undesirable side effects
for patients.
It has been reported that endocannabinoids have a
wide range of regulatory effects in a variety of physiological
processes. One of their most promising actions has to do with their
effect on various types of cancers and their potential use in the
treatment of these diseases (18).
In the present study, it was demonstrated by
immunohistochemistry that CB1 and CB2 receptors are highly
expressed in prostate cancer samples with different degrees of
malignancy, as well as in BPH tissue. The presence of receptors
CB1/CB2 in commercial cell lines (PC3), as well as in primary
cultures of PrC, was additionally demonstrated by
immunocytochemistry and western blot analysis (data not shown) as
it was coincidentally reported by other studies (25,30,33).
Furthermore, we observed that the expression of these receptors was
associated with the degree of malignancy in PrC, with higher
expression in the most aggressive samples of PrC. To analyze the
effect that the analogs of endocannabinoids have on the various PrC
cell cultures, the sensitivity to three different concentrations of
endocannabinoids was evaluated, which established that primary
cultures were more resistant to endocannabinoids than PC3 cells.
This differential effect may be explained by a different expression
of metabolizing enzymes, such as FAAH, which metabolize and degrade
endocannabinoids in situ. Several studies have illustrated
differences in the expression of these enzymes in prostate cancer
cell lines and the effect this has on treatment with
endocannabinoids in cancer cells (34,35).
The action of these enzymes, their expression and related
metabolism of endocannabinoids may account for the weaker effect
that the treatments had on the primary cultures. Moreover, it was
also observed that the effect of the endocannabinoids in regards to
the viability of the different cell cultures occurred in a
dose-dependent manner and that there may be a saturation of the
receptors associated with the increasing concentration of each
treatment.
Furthermore, the effect of the different treatments
on both cell lines and primary cultures was almost totally reversed
when they were previously incubated with the CB1R antagonist
SR141716. As this is a selective antagonist for the CB1 receptor,
the results obtained lead us to conclude that the observed effect
of the different treatments was produced mainly by the activation
of CB1 and not CB2; these results also discard a possible
receptor-independent effect of the endocannabinoids.
Olea-Herrero et al (36) recently postulated that the effect of
methanandamide on the survival of PC3 cells may be carried out
specifically by activation of the CB2 receptor. This finding not
only differs from our results but also of the published data by
Agudelo et al (37); who
despite not using methanandamide used its endogenous analog
anandamide. This group reported that the effect of anandamide was
produced by the activation of CB1 receptors. Furthermore, the
affinity of methanandamide for the CB2 receptor was very low (with
a Ki of 815 nM for CB2 and a Ki of 20 nM for CB1). Some researchers
even consider this molecule to be a specific agonist for CB1. In
previous experiments from our laboratory using radioligand binding
studies, we observed that in both cell lines and primary cultures,
endocannabinoids bound more readily to CB1 receptors than to CB2
receptors (data not shown). This may account for the effect
observed when using the specific inhibitor for CB1.
Although previous results depicted a decrease in
PCNA in PrC cell lines following endocannabinoid treatment, the
cell cycle analysis assessed by flow cytometry showed no variations
in the distribution of the different stages of the cell cycle.
Other studies have reported cell cycle arrest following treatment
with different THC analogs but using them at higher concentrations
(39,40), which may be one reason why even
though we previously observed that levels of PCNA were lower after
the treatments, that was not enough to modify the cell cycle
distribution of the different cell cultures. Another reason for
this may be that the decrease in the PCNA levels was not high
enough to have a significant effect on the cell cycle (38).
Once we concluded that the effect of
endocannabinoids on cell viability was not exerted through a
modification of the cell cycle, we analyzed whether this effect may
be due to activation of the apoptotic pathway. Through Annexin V
assays we observed no change in the percentage of necrotic cells
after the treatment with endocannabinoids, but we observed an
increase in the number of cells in early apoptosis after the
treatments. These results were confirmed by evaluating the
expression of caspase-3 and Bcl-2 post-treatment with the
endocannabinoids. The results revealed an increase in the levels of
active caspase-3 and a decrease in the expression levels of Bcl-2.
This simultaneous effect observed in both proteins may be explained
by a change in the activity of the nuclear factor-κB (NF-κB)
induced by the endocannabinoids. NF-κB is involved in cell
proliferation through multiple mechanisms, some of which control
the expression of Bcl-2 and also the activation of proteins
involved in the apoptotic pathway such as caspases. It has been
reported that in prostate cancer NF-κB is overactivated, thus
endocannabinoids may act through this pathway, simultaneously
activating caspase-3, downregulating Bcl-2 expression; and other
proteins that may also be involved in the apoptotic process
(7,41).
All of our results suggest that endocannabinoids may
exert their effect by activating the apoptotic pathway without
modifying the cell cycle stage or inducing necrosis. Moreover, a
decrease in the levels of AKT in the primary cultures was detected
after treatment with endocannabinoids. AKT functions as a critical
positive regulator of metabolism and cell proliferation (39) which is positively correlated with
the effect caused by the different treatments on prostate cancer
cultures. These results are supported by other reports that have
shown that AKT is decreased in other types of cancer following
treatment with THC (41).
Furthermore, increases in the activation of Erk and treatment with
cannabinoids have been correlated with antiproliferative processes
in other types of cancer (42). The
results observed in our study depict a simultaneous activation of
the ERK pathway and a decrease in AKT levels. The combination of
these two events may contribute to the activation of
antiproliferative pathways and a decrease in the proliferative
pathways in primary cultures of PrC. A more in-depth analysis of
the proteins involved in these pathways may be important to better
describe the intracellular mechanisms that may be activated by
endocannabinoids in prostate cancer cells and thus search for other
possible targets of intervention to potentiate the effects depicted
by the treatment of cancer cells with endocannabinoids.
Finally and in accordance with our findings, we
conclude that endocannabinoids are capable of halting the growth of
prostate cancer cells through activation of apoptotic mechanisms.
Furthermore, we suggest that this effect may be through the
modulation of the Erk and Akt signaling pathways by
endocannabinoids. Therefore, endocannabinoids appear to be a
powerful tool for investigation in the development of drugs and
treatments against advanced PrC.
Acknowledgements
We acknowledge Ms. Graciela Caroca for her excellent
technical assistance. The present study was supported by
Vicerrectoria de Investigación y Desarrrollo of Universidad de
Chile (VID) Grant DI MULT 05/36-2. Grants FONDECYT, 1060500 (H.C.),
1110269 (H.C.) and 1140417 (E.C.).
References
|
1
|
Jemal A, Bray F, Center MM, et al: Global
cancer statistics. CA Cancer J Clin. 61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sark W: Epidemiology of prostate cancer
and its precursors. Mod Pathol. 10:10382004.(Epub ahead of print).
View Article : Google Scholar
|
|
3
|
Tefekli A and Tunc M: Future prospects in
the diagnosis and management of localized prostate cancer. Sci
World J. 2013:3472632013. View Article : Google Scholar
|
|
4
|
Hsing A and Chokkalingam A: Prostate
cancer epidemiology. Front Biosci. 11:1388–1413. 2006. View Article : Google Scholar
|
|
5
|
Porkka K and Visakorpi T: Molecular
mechanisms of prostate cancer. Eur Urol. 45:683–691. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee C, Jia Z, Rahmatpanah F, Zhang Q, Zi
X, McClelland M and Mercola D: Role of the adjacent stroma cells in
prostate cancer development and progression: synergy between TGF-β
and IGF signaling. Biomed Res Int. 2014:5020932014.
|
|
7
|
Bifulco M, Malfitano A, Pisanti S and
Laezza C: Endocannabinoids in endocrine and related tumours. Endocr
Relat Cancer. 15:391–408. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hermanson DJ and Marnett LJ: Cannabinoids,
endocannabinoids, and cancer. Cancer Metastasis Rev. 30:599–612.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Díaz-Laviada I: The endocannabinoid system
in prostate cancer. Nat Rev Urol. 8:553–561. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Devane WA, Dysarz FA, Johnson MR, et al:
Determination and characterization of a cannabinoid receptor in rat
brain. Mol Pharmacol. 34:605–613. 1998.
|
|
11
|
Van Dross R, Soliman E, Jha S, et al:
Receptor-dependent and receptor-independent endocannabinoid
signaling: A therapeutic target for regulation of cancer growth.
Life Sci. 92:463–466. 2013. View Article : Google Scholar
|
|
12
|
Guindon J and Hohmann AG: The
endocannabinoid system and cancer: therapeutic implication. Br J
Pharmacol. 16:1447–1463. 2011. View Article : Google Scholar
|
|
13
|
Pisanti S and Bifulco M: Endocannabinoid
system modulation in cancer biology and therapy. Pharmacol Res.
60:107–116. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kirkham TC and Tucci SA: Endocannabinoids
in appetite control and the treatment of obesity. CNS Neurol Disord
Drug Targets. 5:272–292. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bovolin P, Cottone E, Pomatto V, et al:
Endocannabinoids are involved in male vertebrate reproduction:
regulatory mechanisms at central and gonadal level. Front
Endocrinol (Lausanne). 5:542014.
|
|
16
|
Jiang W, Zhang Y, Xiao L, Van Cleemput J,
Ji SP, Bai G and Zhang X: Cannabinoids promote embryonic and adult
hippocampus neurogenesis and produce anxiolytic- and
antidepressant-like effects. J Clin Invest. 115:3104–3116. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cridge BJ and Rosengren RJ: Critical
appraisal of the potential use of cannabinoids in cancer
management. Cancer Manag Res. 30:301–313. 2013.
|
|
18
|
Batista LA, Gobira PH, Viana TG, et al:
Inhibition of endocannabinoid neuronal uptake and hydrolysis as
strategies for developing anxiolytic drugs. Behav Pharmacol.
25:425–433. 2014.PubMed/NCBI
|
|
19
|
Bifulco M, Laezza C, Pisanti S and
Gazzerro P: Cannabinoids and cancer: pros and cons of an antitumor
strategy. Br J Pharmacol. 148:123–135. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ramos JA and Bianco FJ: The role of
cannabinoids in prostate cancer: Basic science perspective and
potential clinical applications. Indian J Urol. 28:9–14. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Melck D, Rueda D, Galve-Roperh I, et al:
Involvement of the cAMP/protein kinase A pathway and of
mitogen-activated protein kinase in the anti-proliferative effects
of anandamide in human breast cancer cells. FEBS Lett. 463:235–240.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nithipatikom K, Endsley M, Isbell M, et
al: 2-Arachidonoyl-glycerol: a novel inhibitor of
androgen-independent prostate cancer cell invasion. Cancer Res.
15:8826–8830. 2004. View Article : Google Scholar
|
|
23
|
Portella G, Laezza C, Laccetti P, et al:
Inhibitory effects of cannabinoid CB1 receptor stimulation on tumor
growth and metastatic spreading: actions on signals involved in
angiogenesis and metastasis. FASEB J. 17:1771–1773. 2003.PubMed/NCBI
|
|
24
|
Melck D, De Petrocellis L, Orlando P, et
al: Suppression of nerve growth factor Trk receptor and prolactin
receptors by endocannabinoids leads to inhibition of human breast
and prostate cancer cell proliferation. Endocrinology. 141:118–126.
2000.
|
|
25
|
Czifra G, Varga A, Nyeste K, et al:
Increased expressions of cannabinoid receptor-1 and transient
receptor potential vanilloid-1 in human prostate carcinoma. J
Cancer Res Clin Oncol. 135:507–514. 2009. View Article : Google Scholar
|
|
26
|
Mimeault M, Pommery N, Wattez N, et al:
Anti-proliferative and apoptotic effects of anandamide in human
prostatic cancer cell lines: implication of epidermal growth factor
receptor down-regulation and ceramide production. Postate. 15:1–12.
2003. View Article : Google Scholar
|
|
27
|
Ruiz L, Miguel A and Díaz-Laviada I:
Delta9-tetrahydro-cannabinol induces apoptosis in human prostate
PC-3 cells via a receptor-independent mechanism. FEBS Lett.
458:400–404. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sarfaraz S, Afaq F, Adhami VM and Mukhtar
H: Cannabinoid receptor as a novel target for the treatment of
prostate cancer. Cancer Res. 65:1635–1641. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gómez del Pulgar T, Velasco G and Guzmán
M: The CB1 cannabinoid receptor is coupled to the activation of
protein kinase B/Akt. Biochem J. 347:369–373. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gómez del Pulgar T, Velasco G, Sánchez C,
et al: De novo-synthesized ceramide is involved in
cannabinoid-induced apoptosis. Biochem J. 363:183–188. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rueda D, Galve-Roperh I, Haro A and Guzmán
M: The CB1 cannabinoid receptor is coupled to the activation of
c-Jun N-terminal kinase. Mol Pharmacol. 58:814–820. 2000.PubMed/NCBI
|
|
32
|
Mendoza P, Sánchez C, Contreras HR, et al:
Evaluation of MENT on primary cell cultures from benign prostatic
hyperplasia and prostate carcinoma. Int J Androl. 32:607–615. 2009.
View Article : Google Scholar
|
|
33
|
Chunga S, Hammarstenb P, Josefssonb A, et
al: A high cannabinoid CB(1) receptor immunoreactivity is
associated with disease severity and outcome in prostate cancer.
Eur J Cancer. 45:174–182. 2009. View Article : Google Scholar
|
|
34
|
Brown L, Cascio M, Wahle K, et al:
Cannabinoid receptor- dependent and -independent anti-proliferative
effects of omega-3 ethanolamides in androgen receptor-positive
and-negative prostate cancer cell lines. Carcinogenesis.
31:1584–1591. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nithipatikom K, Isbell M, Endsleya M, et
al: Anti-proliferative effect of a putative endocannabinoid,
2-arachidonylglyceryl ether in prostate carcinoma cells.
Prostaglandins Other Lipid Mediat. 94:34–43. 2011. View Article : Google Scholar :
|
|
36
|
Olea-Herrero N, Vara D, Malagarie-Cazenave
S and Díaz-Laviada I: Inhibition of human tumour prostate PC-3 cell
growth by cannabinoids R(+)-Methanandamide and JWH-015: involvement
of CB2. Br J Cancer. 101:940–950. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Agudelo M, Newton C, Widen R, et al:
Cannabinoid receptor 2 (CB2) mediates immunoglobulin class
switching from IgM to IgE in cultures of murine-purified B
lymphocytes. J Neuroimmune Pharmacol. 3:35–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kelman Z: PCNA: structure, function and
interactions. Oncogene. 6:629–640. 1997. View Article : Google Scholar
|
|
39
|
Song G, Ouyang G and Bao S: The activation
of Akt/PKB signaling pathway and cell survival. J Cell Mol Med.
1:59–71. 2005. View Article : Google Scholar
|
|
40
|
Greenhough A, Patsos H, Ann C, et al: The
cannabinoid D9-tetrahydrocannabinol inhibits RAS-MAPK and PI3K-AKT
survival signalling and induces BAD-mediated apoptosis in
colorectal cancer cells. Int J Cancer. 121:2172–2180. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
De Petrocellis L, Ligresti A, Moriello A,
et al: Non-THC cannabinoids inhibit prostate carcinoma growth in
vitro and in vivo: pro-apoptotic effects and underlying mechanisms.
Br J Pharmacol. 168:79–102. 2013. View Article : Google Scholar :
|
|
42
|
Hsu SS, Huang CJ, Cheng HH, et al:
Anandamide-induced Ca2+ elevation leading to p38 MAPK
phosphorylation and subsequent cell death via apoptosis in human
osteosarcoma cells. Toxicology. 28:21–29. 2007. View Article : Google Scholar
|