Introduction
Endometrial carcinoma is the fourth leading cause of
cancer-related deaths in women worldwide, with an incidence of
approximately 2.26% of the total newly occurring cases among
females in 2008 (1). Surgery,
chemotherapy, radiation and hormonal therapy are common types of
treatment for patients with endometrial cancer (2). Since each patient may have a different
genetic background and pathogenesis, identifying mechanistic
differences at the molecular level will benefit the development of
patient-specific therapies. Extensive research focusing on
endometrial carcinoma has identified several molecules (e.g. PTEN,
CA125 and CA199) that are associated with its initiation (3–5);
however, its molecular pathogenesis has not been fully
elucidated.
OCT4 is a member of the POU domain-containing family
of transcription factors, that is primarily expressed in embryonic
stem (ES) and primordial germ cells, playing an important role in
maintaining pluripotency and self-renewal (6–10). It
is also known that OCT4, together with three other reprogramming
factors, is essential for generating induced pluripotent stem cells
(11,12). Notably, OCT4 expression is closely
related to the occurrence, development and metastasis of multiple
tumors (13–15), suggesting that OCT4 plays a critical
role in carcinogenesis. Previously, we reported that aberrant
expression of OCT4 in ovarian cancer was associated with cancer
cell anti-apoptotic activity (16).
It is known that OCT4 is overexpressed in endometrial carcinoma
(17), although little is known
concerning the function of OCT4 during endometrial
carcinogenesis.
MicroRNAs (miRNAs) are small single-stranded RNAs
which can suppress gene expression transcriptionally and
post-transcriptionally. The effects of miRNAs are mediated by
sequence-specific binding within the 3′-untranslated regions (UTRs)
of cognate mRNA targets, typically resulting in the inhibition of
translation or mRNA degradation (18). A recent study showed that the
tumor-suppressive miRNA, miR-145, directly inhibited the
pluripotency factors OCT4, SOX2 and KLF4 in ES cells, and that
miR-145 inhibited the transcription of OCT4 by binding to its
promoter (19), suggesting that it
may play a critical role in OCT4 dysregulation in carcinogenesis
(20).
In 2011, Salmena et al proposed the competing
endogenous RNA (ceRNA) hypothesis, positing that certain RNAs (i.e.
pseudogenes and long non-coding RNAs) regulate biological function
by competing for miRNA binding (21). Prior to this hypothesis, pseudogenes
were considered to be non-functional genomic sequences (22); however, it has subsequently been
shown that many pseudogenes demonstrate biological activity by
regulating their homologous genes in cells (23,24).
More recent studies indicate that pseudogene transcripts can both
regulate tumor oncogenes and suppressor genes by forming small
interfering RNAs (siRNA) or reducing the concentration of miRNAs in
cells (17,25,26).
It has also been directly shown that pseudogenes can function as
ceRNAs to regulate other RNA transcripts by competing for shared
miRNAs (27). The OCT4 pseudogene 5
(OCT4-pg5) is truncated at the 5′-end and expresses only a partial
open reading frame region followed by a 3′UTR. Furthermore, it is
the target of miR-145 (27,28). Notably, Suo et al found that
OCT4-pg5 was exclusively expressed in cancer tissues but not in
normal tissues (15).
The goal of the present study was to determine the
expression level of OCT4-pg5 in endometrial cancer, and explore the
possibility that it acts as a ceRNA to regulate OCT4 expression,
and activate the OCT4-mediated PI3K/AKT-cyclin D1 signaling pathway
by competitively binding with miR-145.
Materials and methods
Reagents
The primary antibodies raised against OCT4
(ab19857), AKT (ab126811), PCNA (ab29), GAPDH (ab8245)and cyclin D1
(ab134175) were all purchased from Abcam (Cambridge Science Park,
Cambridge, UK), and the anti-p-AKT (244F9) antibody was purchased
from Cell Signaling Technology (Danvers, MA, USA). siRNA
specifically targeting OCT4 (1027416) and negative control siRNA
were obtained from Qiagen (Dusseldorf, Germany). The 3′UTR of
OCT4-pg5 and OCT4 plasmids were constructed by GeneChem (Shanghai,
China). The miRNA inhibitor negative control, miR-145 inhibitor
(4464084), miRNA mimic negative control, and miR-145 mimic
(4464066) were obtained from Ambion (Austin, TX, USA).
Lipofectamine 2000, TRIzol reagent, antibiotics and antimycotics
(Invitrogen, Grand Island, NY, USA); M-MLV reverse transcriptase
(Promega, Madison, WI, USA), Dulbecco’s modified Eagle’s medium
(DMEM), RPMI-1640, fetal bovine serum (FBS; Thermo Scientific,
South Logan, UT, USA), SYBR-Green (Takara, Dalian, China) and PCR
primers (Shanghai Sangon Biotech, Shanghai, China) were all
commercially purchased.
Cell lines and tissue samples
The endometrial cancer cell lines Ishikawa, AN3CA,
SPEC-2, HEC-1b, KLE and RL95–2 were maintained by our laboratory.
Ishikawa, AN3CA, SPEC-2, HEC-1b and KLE cells were cultured in DMEM
with 10% FBS. RL95-2 cells were cultured in RPMI-1640 medium with
10% FBS. All cell lines were maintained with 100 IU/ml penicillin
and 100 μg/ml streptomycin in humidified 5% CO2 at 37°C.
Fourteen samples of fresh human endometrium and 29 cases of
cryopreserved endometrial carcinoma were obtained with informed
consent from patients in the Shanghai First People’s Hospital of
Jiaotong University (Shanghai, China). Half of the fresh
endometrium samples were in the proliferative phase and the other
half were in the secretory phase. All endometrial carcinoma tissues
were diagnosed as endometrioid adenocarcinoma. The use of clinical
specimens in the present study was approved by the Ethics Committee
of Shanghai First People’s Hospital of Jiaotong University.
Transient transfection
Transfections were performed using Lipofectamine
2000 following the manufacturer’s instructions (Invitrogen).
Briefly, the cells were seeded in 60-mm dishes at a density of
1×104 cells/well and cultured to 50–60% confluency.
After serum starvation for 24 h, the double-stranded miR-145 mimic,
inhibitor, OCT4 siRNA or plasmids were transfected into the cells
with Lipofectamine 2000. Twenty four or 48 h later, the cells were
collected for RNA and protein analysis. The sequences of the
miR-145 mimic, inhibitor and OCT4 siRNA are listed in Table I.
 | Table IMicroRNA and OCT4 siRNA
sequences. |
Table I
MicroRNA and OCT4 siRNA
sequences.
| Name | Sequence |
|---|
| hsa-miR-145
mimic |
GUCCAGUUUUCCCAGGAAUCCCUGGAUUCCUGGGAAAACUGGACUU |
| hsa-miR-145
inhibitor |
AGGGAUUCCUGGGAAAACUGGACCAGUACUUUUGUGUAGUACAA |
| OCT4 siRNA |
CAUGUGUAAGCUGCGGCCCTTGGGCCGCAGCUUACACAUGTT |
RNA isolation and real-time quantitative
PCR
Total RNA was isolated using TRIzol reagent
(Invitrogen) following the manufacturer’s recommendations.
Complementary DNA was synthesized from 2.5 μg of total RNA with
random primers and M-MLV reverse transcriptase in a 25-μl reaction
system. The cDNA template (2 μl) was used in a 20-μl reaction
volume with SYBR-Green for real-time PCR. GAPDH served as an
internal normalization control. The PCR primers for OCT4, OCT4-pg5
and GAPDH are listed in Table II.
Amplification was carried out for 38 cycles consisting of 15 sec at
95°C and 30 sec at 62.4°C for OCT4-pg5, and 64°C for OCT4, then 30
sec at 72°C. The 2−ΔΔCT method was used to determine
relative gene expression levels, with the mRNA levels normalized to
GAPDH expression. Each experiment was repeated in triplicate.
| Table IITarget gene PCR primer sequences. |
Table II
Target gene PCR primer sequences.
| Gene | Sequence | Annealing
temperature (°C) |
|---|
| OCT4 |
| Forward
primer |
5′-TCCCTTCGCAAGCCCTCAT-3′ | 64 |
| Reverse
primer |
5′-TGACGGTGCAGGGCTCCGGGGAGGCCCCATC-3′ | |
| Oct4-pg5 |
| Forward
primer |
5′-TTGCTGCAGAAGTGGGTGGAGGAAG-3′ | 62.4 |
| Reverse
primer |
5′-GTACCAAAATGGGAGCCTGGGGC-3′ | |
| GAPDH |
| Forward
primer |
5′-GCACCGTCAAGGCTGAGAAC-3′ | 60 |
| Reverse
primer |
5′-TGGTGAAGACGCCAGTGGA-3′ | |
Western blot analysis
Protein was isolated using cell lysis buffer
supplemented with a protease and phosphatase inhibitor. Isolated
protein (30 μg/lane) was separated on 10% sodium dodecyl
sulfate-polyacrylamide gels, transferred to polyvinylidene fluoride
membranes, and then blocked with 5% skimmed milk for 1 h. Membranes
were incubated overnight with the primary antibodies (OCT4,
1:1,000; AKT, 1:1,000; PCNA, 1:1,000; GAPDH, 1:10,000 and cyclin D1
1:10,000). Antibody binding was revealed after incubation with an
appropriate secondary antibody coupled to peroxidase by using
enhanced chemiluminescence.
Cell proliferation assay
To assess cell proliferation, the cells were stained
with sulforhodamine B (SRB). Briefly, the cells were trypsinized
and seeded in 96-well plates for experimental assessment 24 h after
transfection. The following day (d 0), 1 plate per day was washed
twice with PBS and fixed in 10% trichloroacetic acid solution at
4°C. On day 5, all the plates were stained with 0.4% SRB for 30 min
at room temperature, and rinsed with 1% acetic acid 4 times. After
lysis with 10 mM Tris, the optical density (OD) of the cell lysates
was measured at 490 nm using a microplate reader (Bio-Rad,
Hercules, CA, USA). This experiment was performed in 96-well plates
with 6 replicate wells at least 3 times.
Colony formation assay
After the cells were transfected with the plasmids
or the miR-145 mimic, they were seeded at ~600 cells/well in a
6-well plate and cultured for 10 days. Following paraformaldehyde
fixation and crystal violet staining, the stained cell colonies
were counted. The experiment was carried out in triplicate wells
and repeated 3 times.
Statistical analyses
Statistical analysis was performed using SPSS 21.0
(IBM, Armonk, NY, USA). All data are presented as mean ± standard
deviation (SD). Statistical significance of the real-time PCR and
cell growth assay was analyzed by the Student’s t-test. A P-value
<0.05 was considered to indicate a statistically significant
result.
Results
OCT4-pg5 is overexpressed in endometrial
cancer and is directly targeted by miR-145
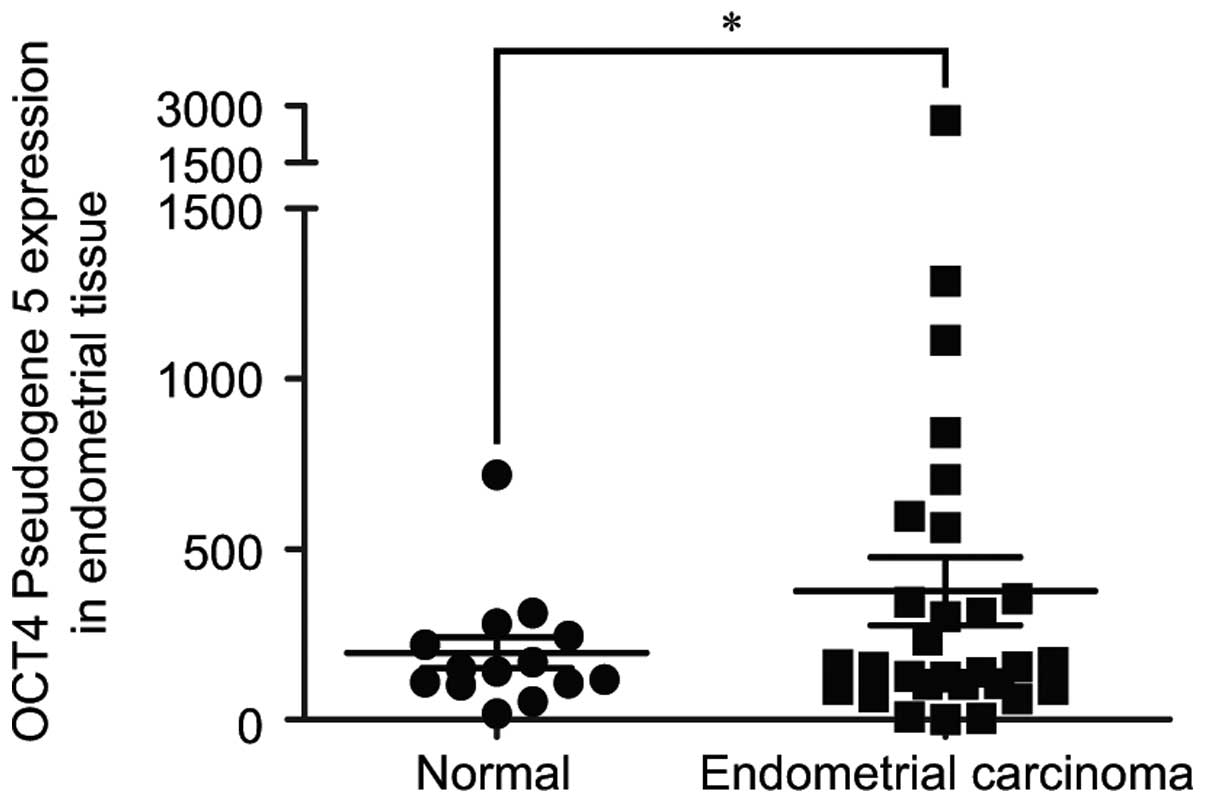
To investigate the relative expression level of
OCT4-pg5 in endometrial cancer, real-time PCR was performed to
assess its mRNA expression level (Fig.
1). OCT4-pg5 mRNA levels were significantly higher in the
endometrial cancer samples compared to the level in the normal
endometrial tissues (P<0.05). Further studies revealed that
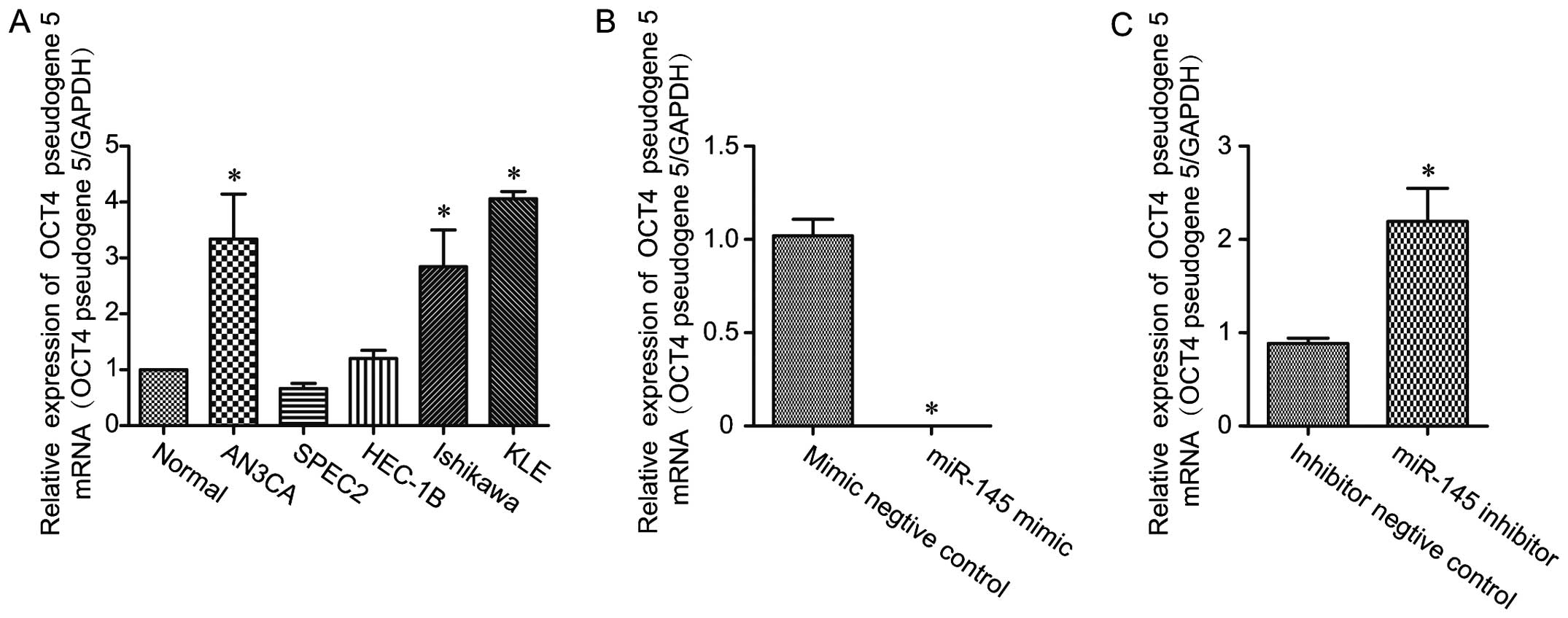
OCT4-pg5 expression was also significantly elevated in the
endometrial cancer cell lines AN3CA, Ishikawa and KLE, with an
upward trend in HEC-1B cells and no change in SPEC-2 cells, as
compared to the expression level in the primary cultured
endometrial cells (Fig. 2A). The
elevated level of OCT4-pg5 in endometrial cancers suggests that it
may play a functional role during carcinogenesis.
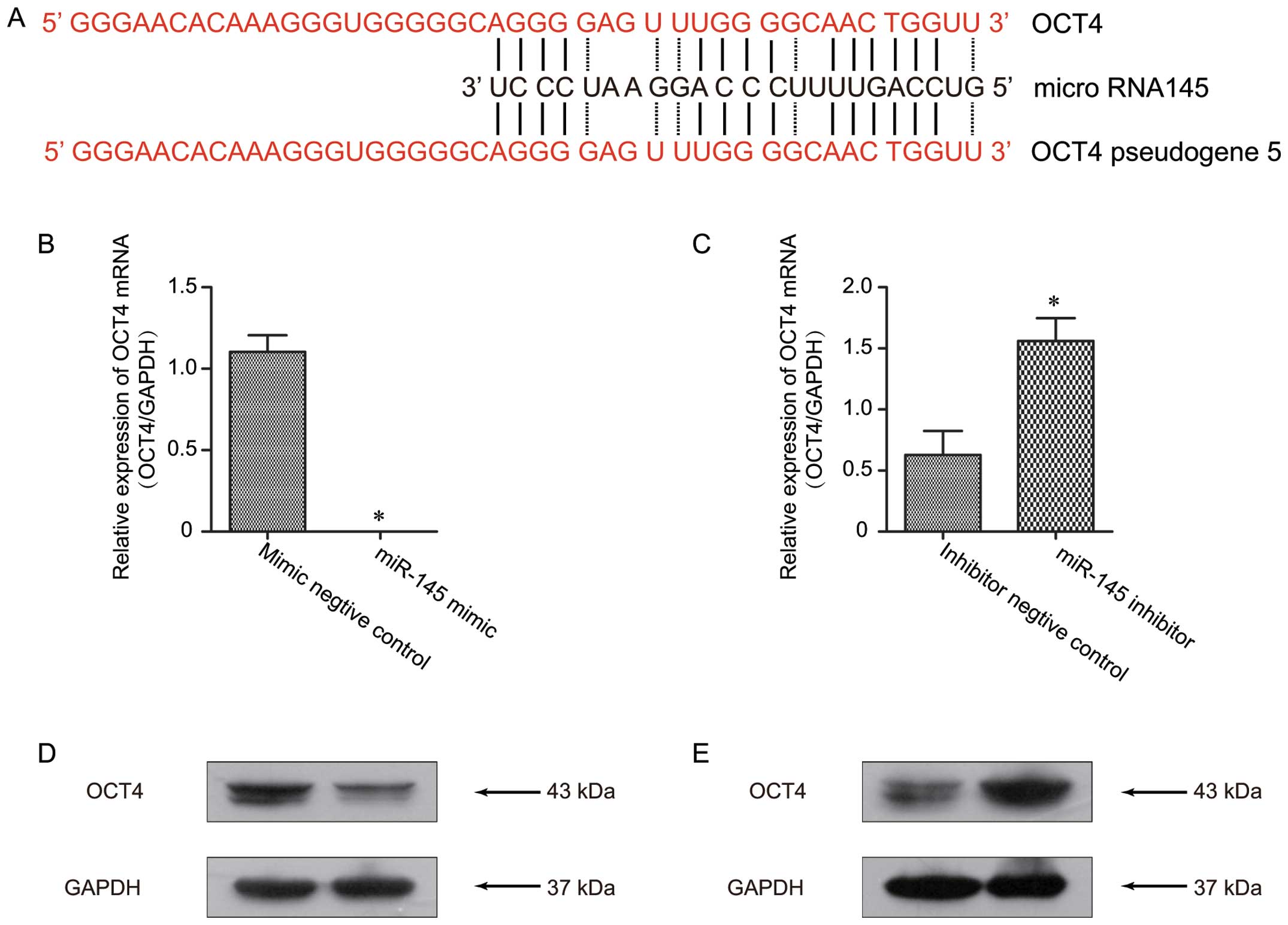
It has been previously reported that miR-145 can
regulate OCT4 expression by targeting its 3′UTR in various cancer
types (17,29). Given that OCT4-pg5 and miR-145 share
a binding site within the OCT4 3′UTR (Fig. 3A), it was hypothesized that miR-145
may also target OCT4-pg5. In agreement with this hypothesis, it was
observed that treatment with 30 nM with miR-145 mimic in Ishikawa
cells resulted in a significant reduction in OCT4-pg5 expression,
compared with the negative control group (Fig. 2B). On the other hand, treatment with
a miR-145 inhibitor resulted in a potent increase in OCT4-pg5
expression, compared to the negative control (Fig. 2C).
OCT4 is directly targeted by miR-145
In order to confirm previous studies that miR-145
regulates OCT4 expression (17,29),
OCT4 expression was assessed in the Ishikawa cells treated with 30
nM of the miR-145 mimic. The results showed a marked suppression in
the OCT4 mRNA (Fig. 3B) and protein
levels (Fig. 3D) in the cells
treated with the miR-145 mimic, as compared to the levels in the
negative control group. Conversely, treatment with the miR-145
inhibitor was associated with increased OCT4 mRNA (Fig. 3C) and protein expression levels
(Fig. 3E), when compared with these
levels in the inhibitor negative control. These data suggest that
miR-145 regulates OCT4 expression via transcriptional inhibition in
Ishikawa cells.
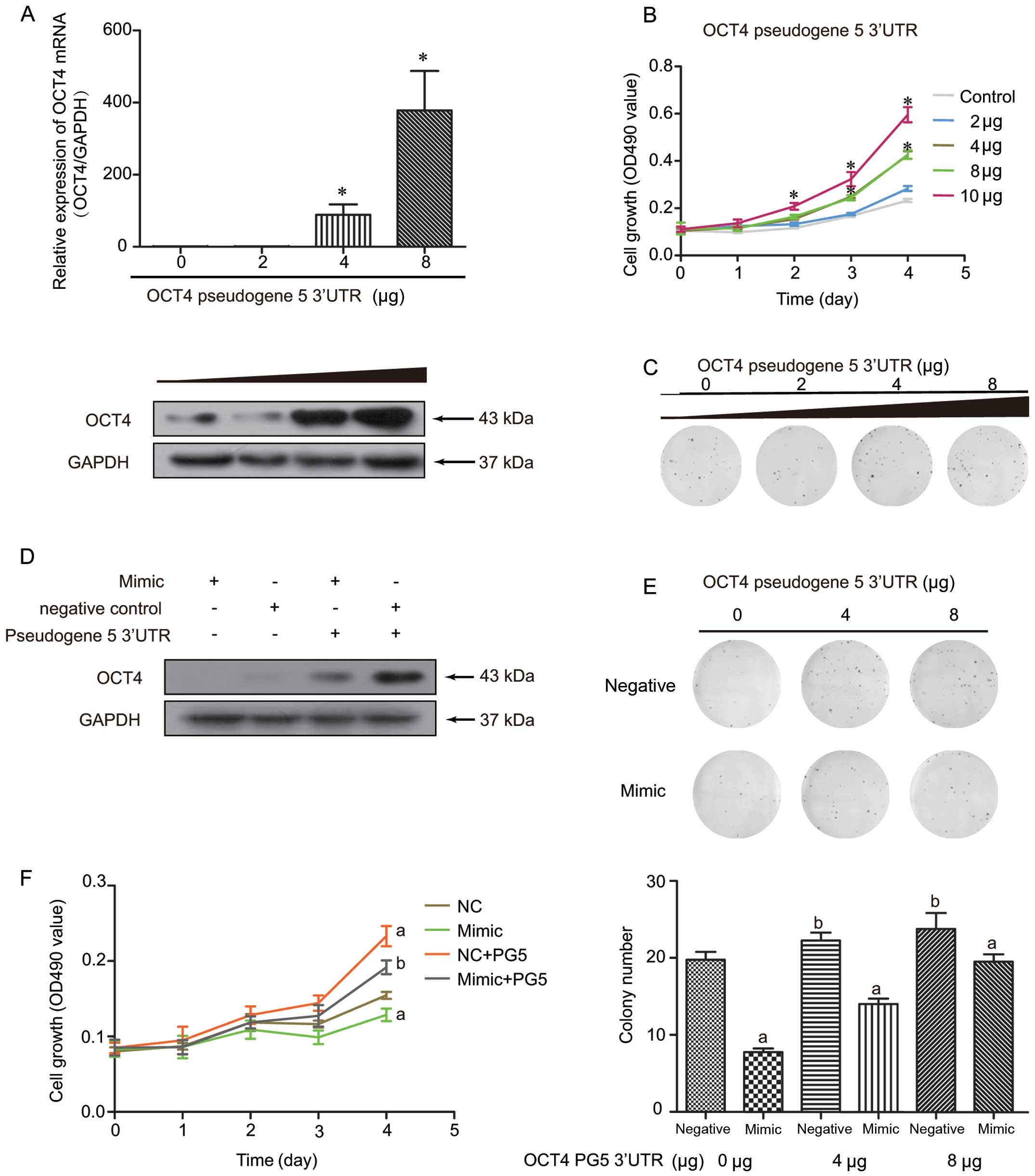
OCT4-pg5 functions as an antagonist
against miR-145 binding to OCT4 to promote proliferation
The data presented above indicated that OCT4-pg5 may
function as an antagonist against miR-145 binding with the OCT4
promoter in Ishikawa cells. To investigate this interaction,
overexpression of OCT4-pg5 3′UTR was performed in Ishikawa cells
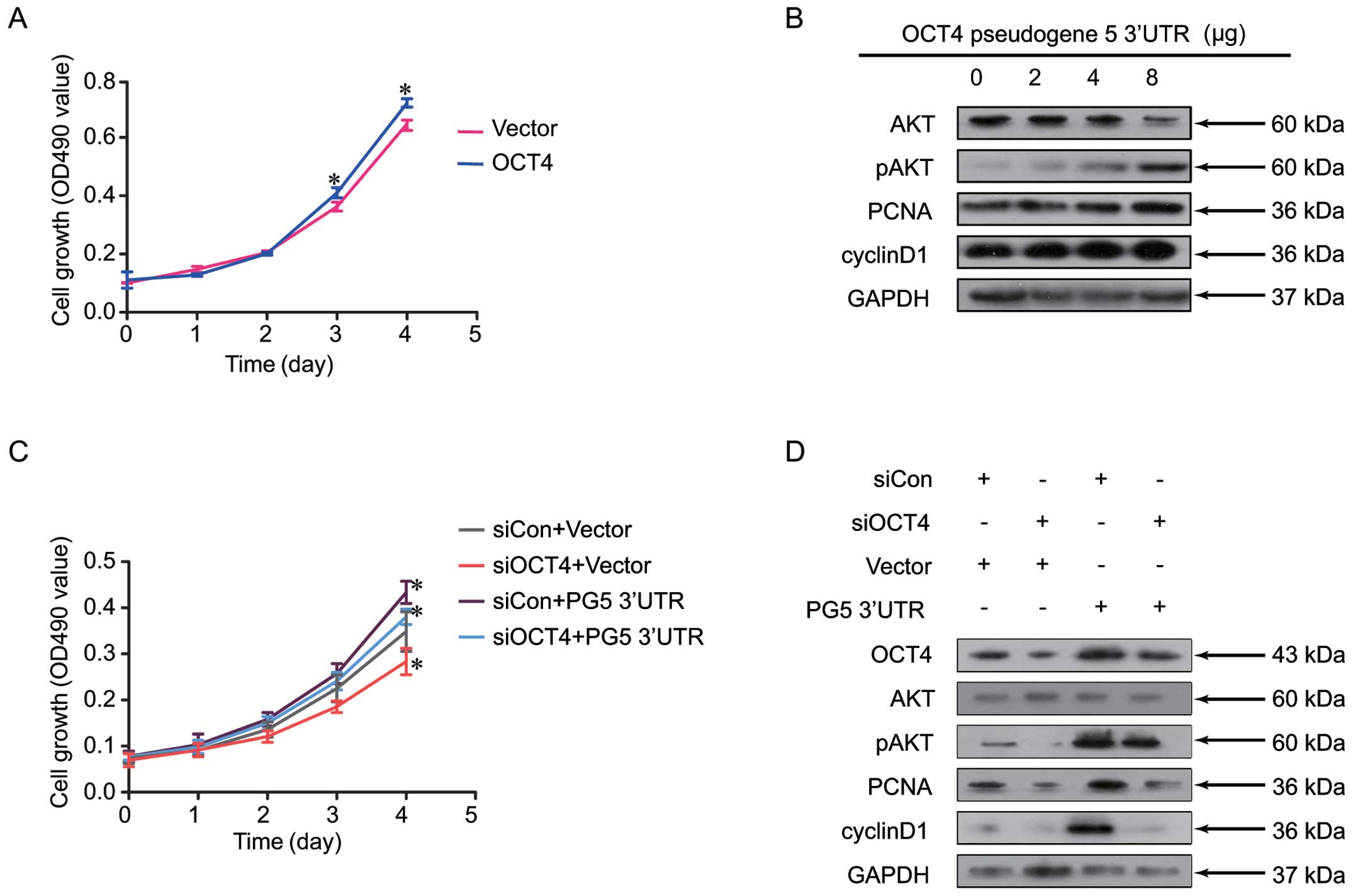
(Fig. 4). It was observed that
increasing the expression level of OCT4-pg5 3′UTR markedly elevated
the OCT4 mRNA and protein levels in a dose-dependent manner
(Fig. 4A). In terms of its
biological function, our SRB assay showed that overexpression of
the OCT4-pg5 3′UTR significantly accelerated cell proliferation
(Fig. 4B) and increased the colony
formation rate (Fig. 4C).
Given that miR-145 targets both OCT4 and OCT4-pg5,
we investigated whether OCT4-pg5 regulates OCT4 expression by
competitive binding with miR-145. As expected, OCT4-pg5-induced
OCT4 expression was attenuated by treatment with the miR-145 mimic
(Fig. 4D), which we attributed to
reduced miR-145 mimic binding to the OCT4 3′UTR as a result of
increasing miR-145 mimic absorption by elevated OCT4-pg5 3′UTR. We
subsequently observed that the exogenous miR-145 mimic also
inhibited OCT4-pg5-induced clone formation (Fig. 4E), although a similar proliferative
profile was observed (Fig. 4F),
suggesting that the miR-145 mimic only partly blocked the role of
OCT4-pg5 3′UTR on proliferation. All things considered, these
results imply that the increase in OCT4 expression and cell
proliferation mediated by OCT4-pg5 overexpression was largely
dependent on endogenous miR-145 expression in endometrial cancer.
It also suggests that OCT4-pg5 functions as a competitive
antagonist of miR-145 to protect OCT4 from being repressed by
miR-145 in endometrial cancer.
OCT4-pg5-mediated Ishikawa cell
proliferation is dependent on OCT4-PI3K/AKT-cyclin D1
signaling
To investigate whether OCT4-pg5-induced
proliferation is dependent on the OCT4-PI3K/AKT-cyclin D1 signaling
pathway in Ishikawa cells, the role of OCT4 on cellular growth and
the effect of OCT4-pg5 3′UTR on this signaling pathway were
studied. Similar to the effect of OCT4-pg5 overexpression described
above (Fig. 4B), increased
expression of OCT4 was associated with increased cell proliferation
(Fig. 5A). Dissection of the
OCT4-PI3K/AKT-cyclin D1 signaling pathway in these cells revealed
that OCT4-pg5 3′UTR transfection resulted in increased protein
expression levels of p-AKT, PCNA and cyclin D1 (Fig. 5B). To confirm that activation of the
PI3K/AKT-cyclin D1 signaling pathway was mediated by OCT4,
knockdown of OCT4 or overexpression of OCT4-pg5 3′UTR was performed
(Fig. 5C and D). The results from
these studies demonstrated that downregulation of p-AKT, PCNA and
cyclin D1 by the miR-145 mimic was rescued by overexpression of
OCT4-pg5, and their upregulation by OCT4-pg5 was blocked by OCT4
siRNA (Fig. 5D). Similarly, the
increased cell proliferation induced by OCT4-pg5 3′UTR was also
attenuated by OCT4 siRNA (Fig.
5C).
Discussion
The molecular mechanisms underlying the development
of endometrial carcinoma have not been fully elucidated. Recently,
OCT4 was observed to be aberrantly expressed in endometrial
carcinoma, with a resultant association with malignancy (17,30,31).
Aberrant OCT4 expression has also been detected in a broad spectrum
of other cancers, including non-small cell lung cancer, hepatoma,
breast and bladder cancer (32–35).
Indeed, it has been suggested that elevated expression of OCT4
contributes to cancer chemotherapy resistance, anti-apoptotic
properties, proliferation and invasion (36–39).
Previously, we observed that OCT4 is over-expressed in ovarian
carcinomas and inhibited FSH-induced apoptosis through enhancing
stem cell-like cell expansion and signaling through the
OCT4-AKT-survivin pathway (16).
All things considered, these facts imply that OCT4 may be an
important trigger in carcinogenesis. It has been reported that OCT4
expression is increased in endometrial cancer, that OCT4-induced
cell growth was blocked by miR-145 through targeting its 3′UTR, and
that OCT4 and miR-145 expression are inversely correlated (17). The goal of the present study was to
investigate the factors affecting miR-145 and OCT4 in endometrial
carcinogenesis.
OCT4 pseudogenes are reportedly overexpressed in
various cancer types (15). In
agreement with this notion, we observed markedly elevated OCT4-pg5
expression levels in primary endometrial carcinoma samples and
several endometrial cancer cell lines. Indeed, consistent with Wu
et al, we observed that upregulation of OCT4-pg5 was
correlated with OCT4 expression (17). Furthermore, exogenous transfection
of OCT4-pg5 3′UTR resulted in a dose-dependent increase in OCT4 at
the mRNA and protein levels, suggesting that OCT4 is a target of
OCT4-pg5, and that this pseudogene is a critical factor affecting
OCT4-mediated carcinogenesis.
Our subsequent experiments were designed to gain a
mechanistic insight into the role of OCT4-pg5, with special
emphasis on miR-145. It is known that OCT4 can be transcriptionally
activated by itself (40), and
negatively regulated by miR-145 (19). Indeed, miR-145 is a well-known
tumor-suppressor miRNA that is downregulated in numerous types of
malignancies, including endometrial cancer (17,41).
Poliseno et al previously suggested that OCT4 and its
pseudogenes share a similar sequence and have conserved microRNA
binding sites (27). In this
regard, we confirmed by sequence alignment that there is a
conserved miR-145 binding site in the OCT4 and OCT4-pg5 3′UTR.
Furthermore, treatment with the miR-145 mimic potently inhibited
both OCT4 and OCT4-pg5 expression, whereas its inhibitor sharply
increased their expression levels. These data indicate that miR-145
is an important modulator of OCT4-pg5′s regulation of OCT4
expression.
All transcribed pseudogenes are theoretically
capable of competing with cognate mRNAs for miRNA binding; however,
a small percentage of pseudogenes could be processed into
endogenous siRNAs to inhibit the expression of cognate genes
through RNA interference (42).
Ebert et al introduced the concept of ‘miRNA sponges’ as a
means to create loss-of-function phenotypes for miRNAs in mammalian
cells (43). Similarly, Poliseno
et al found that the mammalian pseudogene PTENP1 could
function as a ceRNA to regulate PTEN expression by competing for
shared miRNAs with PTENP1 3′UTR, with a similar effect found
between KRAS and its pseudogene KRAS1P (27). In the context of the present study,
Wang et al reported that OCT4-pg4 functions as a natural
microRNA sponge to regulate OCT4 expression by competing for
miR-145 in hepatocellular carcinoma (44). Here, we observed that the inhibition
of OCT4 expression by the miR-145 mimic could be reversed by
overexpression of OCT4-pg5 3′UTR, indicating that exogenous
OCT4-pg5 3′UTR ‘absorbed’ more miR-145, thereby preventing it from
binding to the OCT4 3′UTR. These data are consistent with the
hypothesis that OCT4-pg5 upregulates OCT4 expression by competing
against miR-145 in endometrial cancer.
Our subsequent functional studies demonstrated that
OCT4-pg5 promoted the cell proliferation and colony-forming ability
in endometrial cancer, and that the negative effects of the miR-145
mimic on these parameters could be restored by OCT4-pg5. It was
also observed that knockdown of OCT4 in Ishikawa cells resulted in
a lower rate of cell proliferation, suggesting an oncogenic role
for OCT4 in endometrial carcinogenesis. In our previous studies,
the OCT4-PI3K/AKT-survivin signaling pathway was found to be
involved in the development of ovarian cancer (16). Similarly, Hu et al showed
that OCT4 may maintain the survival of cancer stem cell-like cells
in murine 3LL and human MCF7 tumor cell lines partly through
Oct4/Tcl1/Akt1 signaling, by inhibiting apoptosis (45). Wang et al also observed that
OCT4 can mediate chemoresistance through a potential Oct4-AKT-ABCG2
pathway (36). Here, we
demonstrated that overexpression of OCT4-pg5 increased the protein
levels of cyclin D1 and PCNA and activated the PI3K/AKT signaling
pathway. Nevertheless, OCT4-pg5-induced phosphorylation of AKT and
the increase in cyclin D1 and PCNA were blocked by depletion of
OCT4, as well as transfection with the miR-145 mimic, suggesting
that activating OCT4-PI3K/AKT-cyclin D1 signaling is required for
OCT4-pg5 to promote endometrial cancer proliferation by acting as a
miR-145 antagonist.
This is the first study to investigate the
relationship among OCT4-pg5, miR-145 and OCT4 expression in
endometrial carcinogenesis. We observed that the OCT4-pg5
expression level was significantly higher in endometrial carcinoma
tissues than in benign endometrial tissues. Our results demonstrate
that OCT4-pg5 plays an essential role in miR-145-mediated
endometrial carcinoma cell proliferation through regulation of OCT4
expression and the PI3K/AKT-cyclin D1 signaling pathway. These
findings provide novel insight into the molecular mechanism of cell
proliferation in endometrial carcinoma cells, and may help to
reveal novel targets for the treatment of endometrial cancer.
Acknowledgements
The present study was supported by grants from The
National Natural Science Foundation of China (NSFC nos. 81272883,
81202044, 81370074 and 81172478), The Science and Technology
Commission of Shanghai Municipality (STCSM no. 12ZR1447600), The
Shanghai Municipal Public Health Bureau (no. XYQ2013119), The
Shanghai Jiaotong University (no. BXJ201339) and the grant of
‘Chenxin plan’ from Shanghai Jiaotong University for Z.Z.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
NCCN Practice Guidelines in Oncology.
Uterine Neoplasms Version 1.2013. 2013, http://www.nccn.org/professionals/physician_gls/f_guidelines.asp#uterine.
|
|
3
|
Cheng H, Liu P, Zhang F, et al: A genetic
mouse model of invasive endometrial cancer driven by concurrent
loss of Pten and Lkb1 is highly responsive to mTOR inhibition.
Cancer Res. 74:15–23. 2014. View Article : Google Scholar :
|
|
4
|
Daniilidou K, Frangou-Plemenou M,
Grammatikakis J, Grigoriou O, Vitoratos N and Kondi-Pafiti A:
Prognostic significance and diagnostic value of PTEN and p53
expression in endometrial carcinoma. A retrospective
clinicopathological and immunohistochemical study. J BUON.
18:195–201. 2013.PubMed/NCBI
|
|
5
|
Patsner B and Yim GW: Predictive value of
preoperative serum CA-125 levels in patients with uterine cancer:
the Asian experience 2000 to 2012. Obstet Gynecol Sci. 56:281–288.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Niwa H: Molecular mechanism to maintain
stem cell renewal of ES cells. Cell Struct Funct. 26:137–148. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Niwa H, Miyazaki J and Smith AG:
Quantitative expression of Oct-3/4 defines differentiation,
dedifferentiation or self-renewal of ES cells. Nat Genet.
24:372–376. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pesce M and Scholer HR: Oct-4: control of
totipotency and germline determination. Mol Reprod Dev. 55:452–457.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pesce M and Scholer HR: Oct-4: gatekeeper
in the beginnings of mammalian development. Stem Cells. 19:271–278.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pan GJ, Chang ZY, Schöler HR and Pei D:
Stem cell pluripotency and transcription factor Oct4. Cell Res.
12:321–329. 2002. View Article : Google Scholar
|
|
11
|
Yu J, Vodyanik MA, Smuga-Otto K, et al:
Induced pluripotent stem cell lines derived from human somatic
cells. Science. 318:1917–1920. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Takahashi K and Yamanaka S: Induction of
pluripotent stem cells from mouse embryonic and adult fibroblast
cultures by defined factors. Cell. 126:663–676. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang YD, Cai N, Wu XL, Cao HZ, Xie LL and
Zheng PS: OCT4 promotes tumorigenesis and inhibits apoptosis of
cervical cancer cells by miR-125b/BAK1 pathway. Cell Death Dis.
4:e7602013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Monk M and Holding C: Human embryonic
genes re-expressed in cancer cells. Oncogene. 20:8085–8091. 2001.
View Article : Google Scholar
|
|
15
|
Suo G, Han J, Wang X, et al: Oct4
pseudogenes are transcribed in cancers. Biochem Biophys Res Commun.
337:1047–1051. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang Y, Jin H, Liu Y, et al: FSH inhibits
ovarian cancer cell apoptosis by up-regulating survivin and
down-regulating PDCD6 and DR5. Endocr Relat Cancer. 18:13–26. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu Y, Liu S, Xin H, et al: Up-regulation
of microRNA-145 promotes differentiation by repressing OCT4 in
human endometrial adenocarcinoma cells. Cancer. 117:3989–3998.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bartel DP: MicroRNAs: target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu N, Papagiannakopoulos T, Pan G, Thomson
JA and Kosik KS: MicroRNA-145 regulates OCT4, SOX2, and KLF4 and
represses pluripotency in human embryonic stem cells. Cell.
137:647–658. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hu J, Guo H, Li H, et al: MiR-145
regulates epithelial to mesenchymal transition of breast cancer
cells by targeting Oct4. PLoS One. 7:e459652012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: the Rosetta Stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Petrov DA and Hartl DL: Pseudogene
evolution and natural selection for a compact genome. J Hered.
91:221–227. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hirotsune S, Yoshida N, Chen A, et al: An
expressed pseudogene regulates the messenger-RNA stability of its
homologous coding gene. Nature. 423:91–96. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Korneev SA, Park JH and O’Shea M: Neuronal
expression of neural nitric oxide synthase (nNOS) protein is
suppressed by an antisense RNA transcribed from an NOS pseudogene.
J Neurosci. 19:7711–7720. 1999.PubMed/NCBI
|
|
25
|
Harrison PM, Zheng D, Zhang Z, Carriero N
and Gerstein M: Transcribed processed pseudogenes in the human
genome: an intermediate form of expressed retrosequence lacking
protein-coding ability. Nucleic Acids Res. 33:2374–2383. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Han YJ, Ma SF, Yourek G, Park YD and
Garcia JG: A transcribed pseudogene of MYLK promotes cell
proliferation. FASEB J. 25:2305–2312. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Poliseno L, Salmena L, Zhang J, Carver B,
Haveman WJ and Pandolfi PP: A coding-independent function of gene
and pseudogene mRNAs regulates tumour biology. Nature.
465:1033–1038. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pain D, Chirn GW, Strassel C and Kemp DM:
Multiple retrop-seudogenes from pluripotent cell-specific gene
expression indicates a potential signature for novel gene
identification. J Biol Chem. 280:6265–6268. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang S, Wu Y, Feng D, et al: miR-145
inhibits lung adenocarcinoma stem cells proliferation by targeting
OCT4 gene. Zhongguo Fei Ai Za Zhi. 14:317–322. 2011.(In Chinese).
PubMed/NCBI
|
|
30
|
Mak VC, Siu MK, Wong OG, Chan KK, Ngan HY
and Cheung AN: Dysregulated stemness-related genes in gynecological
malignancies. Histol Histopathol. 27:1121–1130. 2012.PubMed/NCBI
|
|
31
|
Zhou X, Zhou YP, Huang GR, et al:
Expression of the stem cell marker, Nanog, in human endometrial
adenocarcinoma. Int J Gynecol Pathol. 30:262–270. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Karoubi G, Gugger M, Schmid R and Dutly A:
OCT4 expression in human non-small cell lung cancer: implications
for therapeutic intervention. Interact Cardiovasc Thorac Surg.
8:393–397. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Huang PZ, Lu CL, Li BK, et al: OCT4
expression in hepatocellular carcinoma and its clinical
significance. Chin J Cancer. 29:111–116. 2010.(In Chinese).
View Article : Google Scholar
|
|
34
|
Ezeh UI, Turek PJ, Reijo RA and Clark AT:
Human embryonic stem cell genes OCT4, NANOG, STELLAR, and GDF3 are
expressed in both seminoma and breast carcinoma. Cancer.
104:2255–2265. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu K, Zhu Z and Zeng F: Expression and
significance of Oct4 in bladder cancer. J Huazhong Univ Sci
Technolog Med Sci. 27:675–677. 2007. View Article : Google Scholar
|
|
36
|
Wang XQ, Ongkeko WM, Chen L, et al:
Octamer 4 (Oct4) mediates chemotherapeutic drug resistance in liver
cancer cells through a potential Oct4-AKT-ATP-binding cassette G2
pathway. Hepatology. 52:528–539. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang Z, Zhu Y, Lai Y, et al:
Follicle-stimulating hormone inhibits apoptosis in ovarian cancer
cells by regulating the OCT4 stem cell signaling pathway. Int J
Oncol. 43:1194–1204. 2013.PubMed/NCBI
|
|
38
|
Li C, Yan Y, Ji W, et al: OCT4 positively
regulates survivin expression to promote cancer cell proliferation
and leads to poor prognosis in esophageal squamous cell carcinoma.
PLoS One. 7:e496932012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dai X, Ge J, Wang X, Qian X, Zhang C and
Li X: OCT4 regulates epithelial-mesenchymal transition and its
knockdown inhibits colorectal cancer cell migration and invasion.
Oncol Rep. 29:155–160. 2013.
|
|
40
|
Pei D: Regulation of pluripotency and
reprogramming by transcription factors. J Biol Chem. 284:3365–3369.
2009. View Article : Google Scholar
|
|
41
|
Slaby O, Svoboda M, Fabian P, et al:
Altered expression of miR-21, miR-31, miR-143 and miR-145 is
related to clinicopathologic features of colorectal cancer.
Oncology. 72:397–402. 2007. View Article : Google Scholar
|
|
42
|
Tam OH, Aravin AA, Stein P, et al:
Pseudogene-derived small interfering RNAs regulate gene expression
in mouse oocytes. Nature. 453:534–538. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ebert MS, Neilson JR and Sharp PA:
MicroRNA sponges: competitive inhibitors of small RNAs in mammalian
cells. Nat Methods. 4:721–726. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang L, Guo ZY, Zhang R, Xin B, Chen R,
Zhao J, Wang T, Wen WH, Jia LT, Yao LB and Yang AG: Pseudogene
OCT4-pg4 functions as a natural micro RNA sponge to regulate OCT4
expression by competing for miR-145 in hepatocellular carcinoma.
Carcinogenesis. 34:1773–1781. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hu T, Liu S, Breiter DR, Wang F, Tang Y
and Sun S: Octamer 4 small interfering RNA results in cancer stem
cell-like cell apoptosis. Cancer Res. 68:6533–6540. 2008.
View Article : Google Scholar : PubMed/NCBI
|