1. Introduction
Cancer is a major cause of morbidity and mortality
worldwide and remains a formidable disease. According to the
National Cancer Institute (NCI), it was estimated that there would
be 1,665,540 new cases of all cancer types and 585,720 people would
succumb to cancer in 2014. Conventional treatment for cancer
patients include surgery, radiotherapy and chemotherapy. Over the
last few decades, adjuvant chemotherapy following surgery has been
proven to decrease recurrence and improve patient survival, yet may
subsequently promote other malignancies such as ovarian (1), bowel (2), gastro-oesophageal (3) and pancreatic (4) cancers. Recently, several advanced
alternative treatments including gene and targeted therapy are
attracting worldwide attention (5,6).
However, these treatments are not effective, and resistance against
chemotherapy, toxicity, side-effects and the unaffordable cost for
most patients require the search for comparatively natural products
or non-toxic drugs (7). Therefore,
it is of urgent need in the biomedical science to develop novel
anticancer agents with fewer and less debilitating side-effects and
reduced drug resistance to satisfy the unfulfilled therapeutic
demand of cancer patients.
Agents derived from natural sources exert a
multitude of effects on chronic inflammation-driven diseases
including cancer (8). Recently,
much attention has been focused on medicinal plants or herbs as
potential sources of new therapeutic anticancer drugs due to their
lack of toxic effects, relatively lower cost and enhanced
effectiveness (9–11). It was reported that these agents
increase chemotherapeutic efficacy, reduce toxicity, and prolong
the survival time, improve the immune functions and the quality of
life of cancer patients (12).
Trametes robiniophila Murr. (Huaier) is a
sandy beige mushroom found on the trunks of trees and was first
recorded by Shi-Zhen Li, a famous Chinese practitioner in the Ming
Dynasty. Huaier has been widely used in traditional Chinese
medicine (TCM) for ~1,600 years (13). However, the anticancer effects of
Huaier which have attracted increasing worldwide interest have only
been researched in recent years (14–16).
Accumulating evidence suggests that the anticancer mechanisms of
Huaier effects may be associated with various biological
activities, such as inhibition of cell proliferation (15), anti-metastasis (17), interference with tumor angiogenesis
(14) and tumor-specific
immunomodulatory effect (16,18).
Huaier was found to be a putative anticancer agent with broad
therapeutic value following elaborate molecular studies conducted
in the recent past. Irrespective of its effectiveness as a
complementary therapy for cancer, to date, there is no systematic
review available concerning the anticancer effects of Huaier and
the underlying mechanisms. Understanding the mechanisms involved in
the anticancer action of Huaier should provide useful information
for its possible application in clinical cancer therapy and also in
cancer prevention. This review briefly summarizes the literature
published to date reporting the anticancer effect of Huaier and its
underlying mechanisms.
2. Anti-proliferative effect of Huaier
Cell proliferation is an indispensible process in
the occurrence and development of cancer. In previous studies, the
MTT, CCK-8 cell viability or sulforhodamine B (SRB) assay has been
used to measure the cell viability after Huaier treatment (16,17,19,20).
The results showed that W-NTRP (a neutral water-soluble
polysaccharide isolated from the fruit bodies of Huaier)
dose-dependently inhibited the proliferation of three human
cholangiocarcinoma cell lines (QBC939, Sk-ChA-1 and MZ-ChA-1) in
vitro (16). The same trend was
also found in Huaier polysaccharide (HP)-treated hepatocarcinoma
cells (MHCC97-H) (21), Huaier
extract-treated melanoma cells (A875) (20), ovarian epithelial cancer cells
(SKOV3, SKOV3. ip1 and HEY) (17)
and breast cancer cells (MCF-7 and MDA-MB-231) (19). The underlying mechanism associated
with the proliferation inhibitory effect of Huaier was further
revealed in which several well-studied proteins and classic
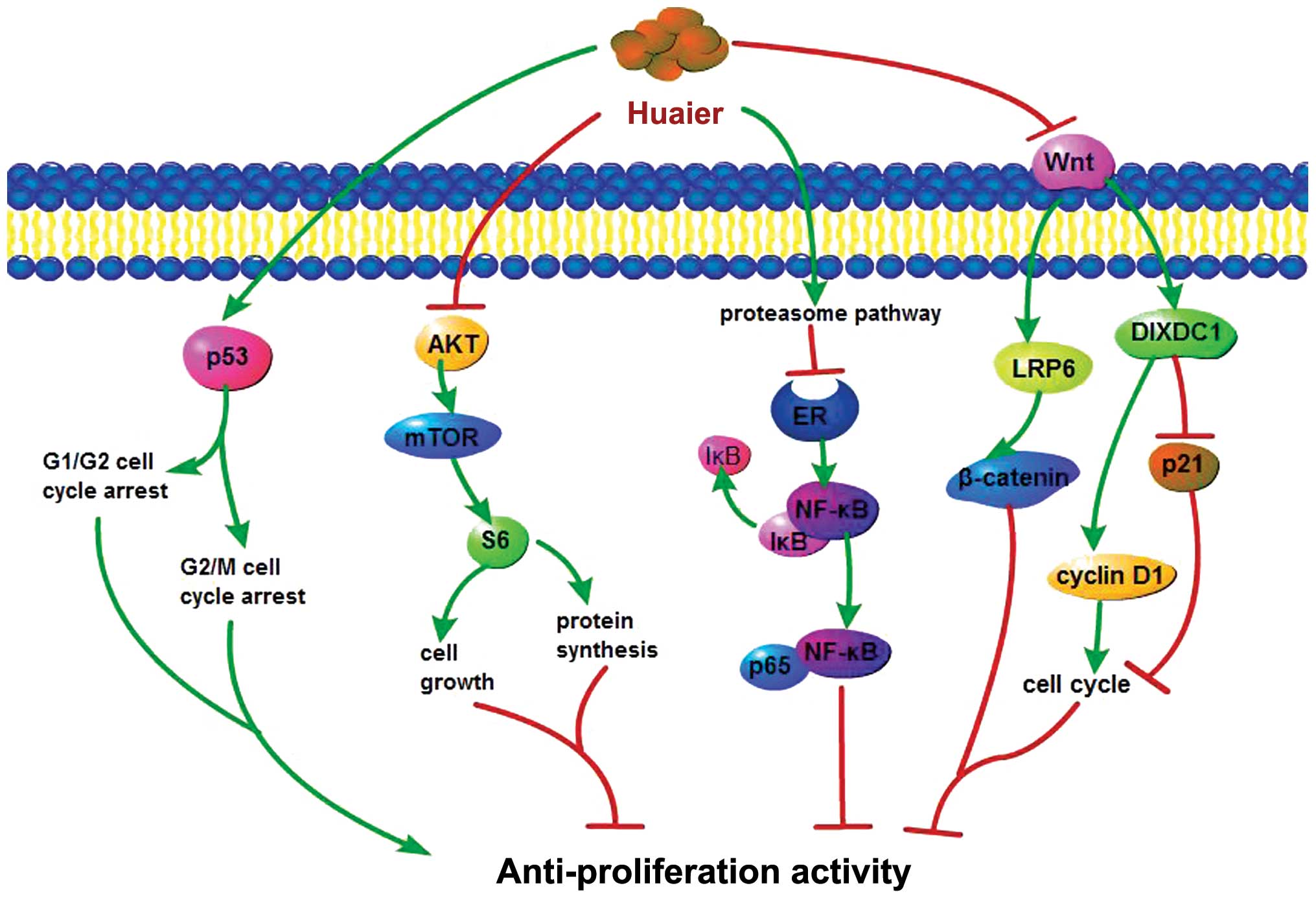
pathways were involved (Fig.
1).
The p53 tumor-suppressor gene can influence cell
cycle progression, DNA damage repair, genomic stability and thus
inhibit the proliferation of cancer cells (22). To determine whether the
proliferation inhibitory effect of Huaier is due to p53-induced
cell-cycle arrest, cell cycle distribution of Huaier-treated MCF-7,
MDA-MB-231 and A875 cells was analyzed by flow cytometry (19,20).
G0/G1 arrest was found in the MCF-7 and MDA-MB-231 cells,
suggesting that Huaier inhibits cell proliferation via cell-cycle
arrest at the G0/G1 phase (19).
The time- and dose-dependent increase in the fraction of cells in
the G2/M phase and the decrease in PCNA protein (a molecular marker
for proliferation) expression in the A875 cells treated with Huaier
extract confirmed that Huaier extract inhibited melanoma A875 cell
proliferation via cell-cycle arrest at the G2/M phase (20). The enhanced p53 expression detected
in the MCF-7, MDA-MB-231 and A875 cells led to the conclusion that
Huaier inhibits cell proliferation via p53-induced cell-cycle
arrest (19,20).
The pAKT/mTOR/S6 pathway
It has been demonstrated that the AKT/mTOR (protein
kinase B/mammalian target of rapamycin) signaling pathway is
important for cell growth (23).
Ribosomal S6 kinase, a key target of mTOR, controls protein
synthesis in cells. Reduced S6 kinase activity leads to a decrease
in protein synthesis and inhibits cell growth (24). To detect whether Huaier inhibits
proliferation though the pAKT/mTOR/S6 pathway, the protein levels
of pAKT, AKT, pS6 (S235-236) and pS6 (S240-244) were evaluated by
western blot analysis. The results showed that phosphorylation of
S6 at S235-236 and S240-244 was significantly downregulated in
three ovarian epithelial cancer cell lines (SKOV3, SKOV3.ip1 and
HEY) after 72 h of Huaier treatment, in accordance with the
decreased pAKT expression in SKOV3 and SKOV3.ip1 cells, indicating
that the pAKT/mTOR/S6 kinase pathway is involved in the
anti-proliferative effects of Huaier (17).
The ER/NF-κB pathway
Estrogen receptors (ER-α and ER-β) are members of
the super family of nuclear steroid hormone receptors (25). They regulate the expression of
target genes through their binding to specific DNA target sequences
or by interacting with other transcription factors (26). A recent study found that Huaier
extract efficiently inhibited estrogen-stimulated proliferation in
three ERα-positive breast cancer cell lines (MCF-7, T47D and
ZR-75-1), confirmed by marked downregulation of ERα mRNA and
protein levels in Huaier-treated cells. This process was associated
with activation of the proteasome (27). The nuclear factor κB (NF-κB)
pathway, an essential pathway for tumorigenesis (28), is involved in this process.
Following Huaier treatment, phosphorylated levels of p65 were
reduced almost to the basal level without estrogen, suggesting that
Huaier extract abolished the effect of estrogen on the activation
of NF-κB and thus suppressed the proliferation of breast cancer
cells induced by estrogen (27).
The Wnt/β-catenin pathway
The classic Wnt/β-catenin signaling pathway is
essential to human beings since aberrant Wnt/β-catenin signaling
leads to a series of human diseases including various types of
cancers (29). It was demonstrated
that Huaier inhibits the proliferation of cancer cells via the
Wnt/β-catenin signaling pathway (17). The detection of decreased Wnt target
gene expression after Huaier treatment provides evidence in support
of Wnt/β-catenin pathway inhibition. A significant reduction in the
expression of DIXDC1 and LRP6 was detected upon Huaier treatment
(17). DIXDC1, the human homolog of
Ccd1, is a positive Wnt signaling pathway protein that functions
downstream of Wnt and upstream of axin and promotes cancer cell
proliferation by targeting p21 and cyclin D1 both of which
facilitate cell cycle progression from G1 to S-phase (30). LRP6 recruits axin and Dishevelled to
the plasma membrane, thereby disrupting the degradation of
β-catenin and facilitating β-catenin nuclear translocation
(31). The downregulation of both
DIXDC1 and LRP6 confirmed the anti-proliferative effect of Huaier
via the Wnt/β-catenin pathway.
3. Anti-metastatic effect of Huaier
As one of the major causes of cancer treatment
failure and ultimately mortality in cancer patients (32), metastasis is an extremely
complicated multi-step process involving the separation of cancer
cells from their primary site by penetrating the stromal tissue,
circulation through the blood vessels or the lymph nodes, adhesion
to the basement membrane and invasion of the target organ for
distinct metastasis (33), thereby
causing poor prognosis and survival of patients. Moreover, over 90%
of cancer-related deaths are due to metastatic disease (34). Recent studies have shown that Huaier
extract suppresses metastasis in ovarian cancer SKOV3, SKOV3.ip1
and HEY cell lines and in the human breast cancer MCF-7 and
MDA-MB-231 cell lines, as well as in hepatocarcinoma MHCC97-H cells
(17,19,21),
indicating that Huaier may serve as a potent anti-metastatic agent
for cancer therapy. Migration and scratch assays in vitro
were performed to determine the cell migratory ability. In
comparison with untreated cells, significant inhibition of
migrational movement was observed in migration assays in MDA-MB-231
and MCF-7 cells treated with 4 mg/ml Huaier (19). In scratch assays, the migration
index (corresponding to wound-healing capacity) was significantly
inhibited in Huaier-treated SKOV3, SKOV3. ip1 and HEY cells
compared with untreated cells, respectively (17). HP exerted the same effect in the
scratch assays. The cell migration of human hepatocellular
carcinoma cell line MHCC97-H was controlled in a time-dependent
manner by HP, being inhibited by up to 28.57, 47.81 and 57.19% at
25, 50 and 100 µg/ml, respectively (21). Invasion assays were conducted to
compare the invasive potential of Huaier extract-treated cells
including MDA-MB-231, SKOV3, SKOV3. ip1, HEY and HP-treated
MHCC97-H cells with untreated cells. The results revealed that
following Huaier treatment the numbers of invading cells through
the Matrigel-coated membrane were significantly decreased compared
with the untreated groups (17,19,21).
In cell adhesion assays performed on MHCC97-H cells, the numbers of
adhesive cells on Matrigel-coated plates were stained with
hematoxylin and eosin (H&E) reagent and counted under an
inverted microscope. HP treatment significantly inhibited cell
adhesion to the Matrigel-coated substrate in a dose-dependent
manner; the reduction was 28.55, 47.48 and 56.47% with HP at 25, 50
and 100 µg/ml, respectively (21). The underlying mechanisms of these
effects were revealed in several studies suggesting that the
GSK3β/β-catenin and AEG-1 pathways were involved (17,19,21)
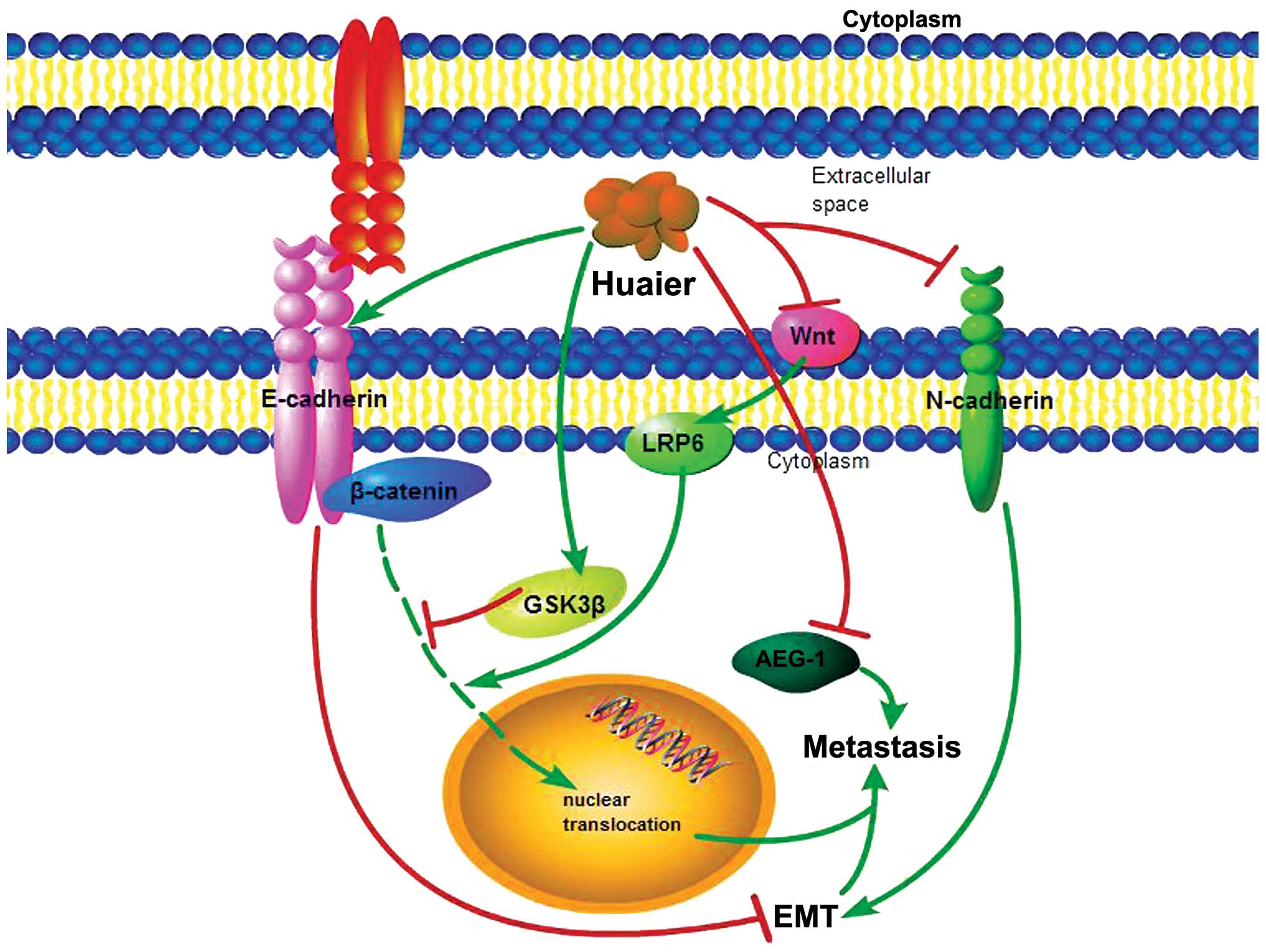
(Fig. 2). These previous results
suggest that Huaier may serve as a potent anti-metastatic agent of
enormous clinical value in cancer therapy.
The GSK3β/β-catenin pathway
In the E-cadherin-mediated cell-cell adhesion
cascade, β-catenin is a key component which can be inhibited by
glycogen synthase kinase 3β (GSK3β) via phosphorylation of
β-catenin leading to its ubiquitination and proteasomal degradation
(35,36). Yan et al found that following
treatment with Huaier aqueous extract for increasing time periods
(24, 48 and 72 h) and concentrations (5.0 and 7.5 mg/ml), the
cytosolic accumulation and nuclear expression of β-catenin were
markedly decreased in a time-dependent manner in SKOV3, SKOV3. ip1
and HEY cells. Huaier treatment also markedly enhanced total GSK3β
expression in a dose-dependent manner in these cell lines while
inhibiting GSK3β S9 phosphorylation. The expression of LRP6, a
promoter of β-catenin nuclear translocation, was significantly
decreased in the Huaier-treated SKOV3 and SKOV3.ip1 cells. These
data indicate that Huaier suppresses not only protein expression,
but also the nuclear translocation of β-catenin due to GSK3β and
LRP6. Increased E-cadherin expression was found in the
Huaier-treated HEY cells, which inhibits the invasive ability of
carcinoma cells. These studies revealed that Huaier inhibited the
cell mobility of ovarian cancer cells via the GSK3β/β-catenin
signaling pathway (17).
Inactivation of epithelial-mesenchymal
transition (EMT) and the AEG-1 pathway
HP was obtained as a water-soluble brown powder from
Huaier extract and was found to exert an anti-metastatic effect on
human hepatocellular carcinoma cell line MHCC97-H (21). Following wound-healing, cell
adhesion and cell invasion assays on MHCC97-H cells treated with
various concentrations of HP (0, 25, 50 or 100 µg/ml), it
was demonstrated that HP inhibited cell adhesion, invasion and
migration in vitro. AEG-1, which has emerged as a vital
oncogene in multiple aspects of the development and progression of
cancers including HCC (37–39), was significantly decreased after HP
treatment. The expression of E-cadherin and N-cadherin as markers
of epithelial-mesenchymal transition (EMT) was also detected by
western blot analysis. Reduced N-cadherin expression and enhanced
E-cadherin expression indicate that the anti-metastatic effect of
HP is through inactivation of EMT and the AEG-1 pathway (21).
4. Anti-angiogenic effect of Huaier
Angiogenesis is a complex process which is defined
as the formation of new blood vessels from pre-existing ones. It
involves a multi-step process which includes degradation of the
extra cellular matrix, migration, proliferation, sprouting,
elongation and tube formation of endothelial cells (40,41).
Angiogenesis plays an essential role in tumor growth, metastasis
and recurrence (42–45). Without vascularization, solid tumors
grow only to 1–2 mm (46). Thus,
preventing angiogenesis is a potential strategy for cancer therapy
and many anti-angiogenic substances have entered the clinic and
have been used as therapeutic options for various types of cancer
(47).
Wang et al (14) reported that exposure to Huaier
aqueous extract led to cell-cycle arrest, decreased mobility and
reduced angiogenesis ability of human umbilical vein endothelial
cells (HUVECs), indicating that Huaier may inhibit tumor-induced
angiogenesis. The study showed that Huaier increased the proportion
of HUVECs in the G0/G1 phase in a dose-dependent manner (from
36.79±2.25% in the control group to 62.41±9.77% in the 8 mg/ml
Huaier group) and promoted the accumulation of p21, a well-studied
cyclin-dependent kinase inhibitor, thus resulting in cell-cycle
arrest. The motility of HUVECs was then examined using modified
scratch and cell migration assays, both of which showed that the
motility of the HUVECs was dose- and time-dependently inhibited
(P<0.01). To further evaluate the effect of Huaier on
angiogenesis, tube formation, chick embryo chorioallantoic membrane
(CAM) and aortic ring assays were subsequently performed in
vitro and ex vivo with direct Huaier treatment. The
assays revealed that Huaier extract caused a marked decrease in the
angiogenesis ability of HUVECs (14). The potential signaling pathways
underlying the potent anti-angiogenic activity of Huaier extract
were also uncovered. Vascular endothelial growth factor (VEGF), one
of the most potent pro-angiogenic factors both physiological and
pathological, is a highly specific mitogen for vascular endothelial
cells and a vascular permeability enhancer which is essential for
endothelial cell proliferation, migration and anti-apoptosis
(48,49). Studies have demonstrated that the
overexpression of VEGF in cancer patients is associated with poor
prognosis and decreased survival (50,51).
Huaier extract inhibited the expression of VEGF and the activation
of ERK in a dose-dependent manner and eventually exhibited
anti-angiogenic activity. Huaier extract also suppressd the
phosphorylation of JNK, STAT3 and p65 (major components in the
NF-κB complex), all of which are important pathways that regulate
cell migration.
The anti-angiogenic activity in vivo was
evaluated in BALB/c mice (14).
Tumor tissues from the mice injected with 4T1 cells were stained
with H&E, CD34 and TUNEL. Significantly reduced blood vessels
and enhanced apoptosis were observed in mice administered a
100-µl solution containing 50 mg Huaier extract by gavage
daily. The same effect was also found in tumor-bearing New Zealand
rabbits (52). Lower expression of
VEGF and P53 and significantly decreased microvessel density (MVD)
were observed in the rabbits following treatment with Huaier. These
results suggest that Huaier extract may serve as a potent
anti-angiogenic and anticancer agent with broad therapeutic
value.
5. Apoptosis induction by Huaier
Apoptosis is a genetically encoded program of cell
suicide, characterized by the morphological features observed upon
cell death which include nuclear condensation, nuclear and cellular
fragmentation, membrane blebbing and phagocytosis of the dying cell
in the absence of inflammation (53). It is a process that plays an
indispensable role in the control of the growth and development of
organisms, and serves as a natural barrier to cancer development
(54). Blockage of apoptosis may
determine the sensitivity of cancer cells to a wide range of
diverse chemotherapy agents and explain the frequently observed
phenomenon of multidrug resistance. Thus, apoptosis induction is
considered to be a crucial strategy for cancer prevention and
treatment since successful eradication of cancer cells through
apoptosis is one of the ultimate aims of chemotherapy (55). Morphological changes have been
observed in cancer cells exposed to Huaier extract. The majority of
Huaier-treated melanoma A875 and breast cancer MCF-7 cells became
enlarged, irregular-shaped and showed vacuolated changes in the
cytoplasm (19,20). These morphological changes
demonstrated cell damage following treatment with Huaier extract.
Experiments in vitro showed that Huaier inhibited the
proliferation of melanoma (A875), lung adenocarcinoma (A549),
breast cancer (MCF-7 and MDA-MB-231), hepatocellular carcinoma
(Hep-G2) and ovarian cancer (SKOV3, SKOV3.ip1 and HEY) cells by
inducing apoptosis (15,17,19,20,52).
These previous studies have confirmed that Huaier induces the
apoptosis of cancer cells, suggesting that Huaier is an effective
anticancer agent for the treatment of cancer patients.
Mitochondria are the targets of several molecular
pro-apoptotic signal transduction pathways (56). Intrinsic control of apoptosis
requires activation of cytosolic caspases by mitochondrial
cytochrome c release and involves regulation of
mitochondrial outer membrane permeabilization by Bcl-2 family
proteins (57).
Mitochondrial-mediated apoptosis has been studied in breast cancer
MCF-7, melanoma A875, HCC Hep-G2 and ovarian epithelial cancer
SKOV3.ip1 cell lines (17,19,20,52).
Following PI/Annexin V double staining performed in MCF-7, A875 and
SKOV3.ip1 cells treated with Huaier extract, apoptosis was induced
as confirmed by increased late apoptosis or cell death rate (UR)
and the early apoptosis rate (LR). Decreased MMP also confirmed
Huaier-induced apoptosis in MCF-7 cells (19). Subsequent western blot analysis
suggested that this effect was at least partly induced via the
mitochondrial pathway. After treatment with Huaier aqueous extract
for increasing time periods (48 and 72 h) and concentrations (4 and
8 mg/ml), the expression of p53, phosphorylated-p53 (p-p53), bcl-2
and BAX were tested with western blot analysis. The expression of
p53 and p-p53 were upregulated, indicating the accumulation and
activation of p53 in response to Huaier treatment in the breast
cancer cell line MCF-7. Meanwhile, treatment with Huaier extract
suppressed Bcl-2 expression and upregulated BAX expression in a
time- and dose-dependent manner, suggesting mitochondrial-mediated
apoptosis (19). The same trend was
also found in melanoma A875 and ovarian cancer SKOV3.ip1 cells
(17,20). In addition, caspase-3 is the most
studied caspase concerning apoptosis induced by natural products
(58). It has been reported in
previous studies that the activation of caspase-3 was significantly
increased in A875, MCF-7 and MDA-MB-231 cells following Huaier
treatment, confirmed by increased cleaved caspase-3 expression and
decreased pro-caspase-3 expression (19,20).
As detected by immunohistochemistry, Huaier-treated tumor-bearing
New Zealand rabbits had lower expression of P53 and Bcl-2 and
higher expression of Bax with the differences being significant
when compared with the control group (52). The results revealed that Huaier
extract promoted cell apoptosis through the mitochondrial pathway,
thus inhibiting tumor occurrence and progression.
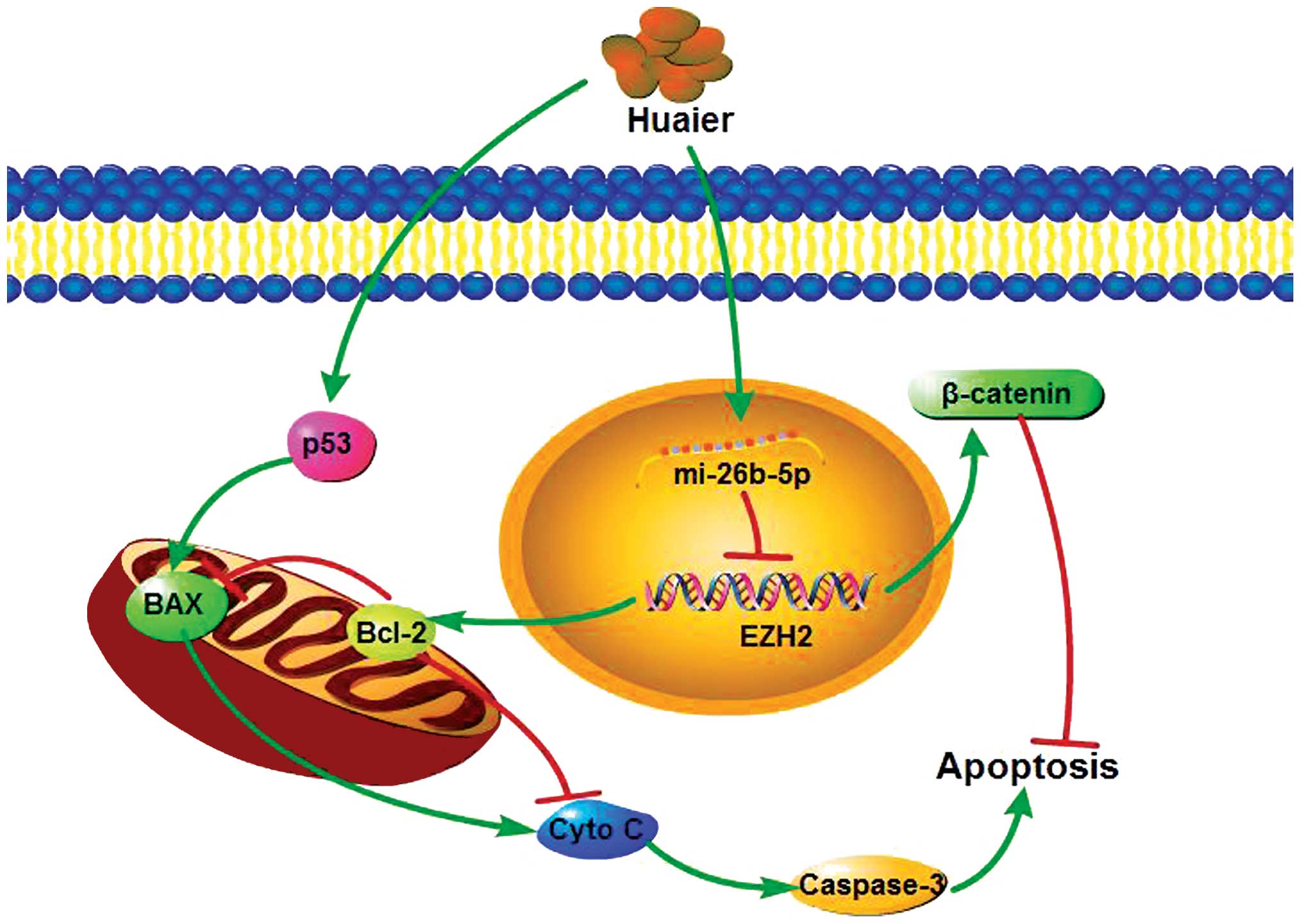
MicroRNAs may act as oncogenes or tumor suppressors
involved in the tumorigenesis and aggression of human cancers
(59). It was observed in a recent
study that Huaier treatment (2–8 mg/ml for 24–72 h) induced cell
apoptosis of pulmonary adenocarcinoma A549 cells via upregulation
of miR-26b-5p (15), an miRNA known
to be significantly downregulated in various types of cancer and to
induce apoptosis in cancer cells (60,61).
Flow cytometric analysis showed that cells treated with 4 mg/ml
Huaier and an miR-26b-5p inhibitor exhibited a 50% decrease in the
rate of apoptosis as compared with the group treated with Huaier
and the inhibitor control, indicating that Huaier induced apoptosis
via miR-26b-5p. EZH2, a target gene of miR-26b-5p, is involved in
the apoptosis of cancer cells and was found to be downregulated in
A549 cells exposed to Huaier. EZH2 is the histone H3 lysine 27
methyltransferase of polycomb-repressive complex 2 and is
overexpressed in multiple cancer types (62). Several EZH2-related proteins were
detected by western blot analysis in both A549 and non-small cell
lung cancer (NSCLC) H1299 cell lines and decreased expression of
both β-catenin and bcl-2 was found in consensus with the previous
finding that EZH2 can ultimately enhance Bcl-2 expression via the
activation of Wnt/β-catenin signaling (63). Thus, Huaier may induce apoptosis in
lung cancer cells via the miR-26b-5p-EZH2-mediated Wnt/β-catenin
pathway (Fig. 3).
6. Inhibition of cancer stem cells by
Huaier
Cancer stem cells (CSCs) represent a small subset of
cancer-initiating cells endowed with self-renewal and multi-lineage
differentiation capacity that promote tumor growth and recurrence
(64–66). Clinically, CSCs resist conventional
cancer therapies including chemotherapy and radiation therapy and
are involved in relapse which is one of the most important features
leading to the poor clinical outcome of cancer patients (67–69).
Therefore, major clinical challenges towards the complete
eradication of cancers are likely to target CSCs (70). In the past few years, the potential
role of naturally occurring agents as potent antitumor agents
functioning by targeting CSCs has been highlighted (71).
Huaier extract inhibited the number and the size of
spheroids formed in both colorectal cancer cells and mammospheres
of breast cancer MCF7 cells at a significantly lower concentration
than those exerting anti-proliferative impact on bulk cancer cells,
indicating that CSCs were preferentially targeted by Huaier extract
(72,73). Considering that CD44, CD24 and ALDH
are cell surface markers for breast cancer MCF7 and colorectal
cancer (CRC) cells, respectively, a decrease in the cell population
of CD446+/CD24− breast cancer cells and
ALDH-positive CRC cells suggested an inhibitory effect of Huaier on
CSCs.
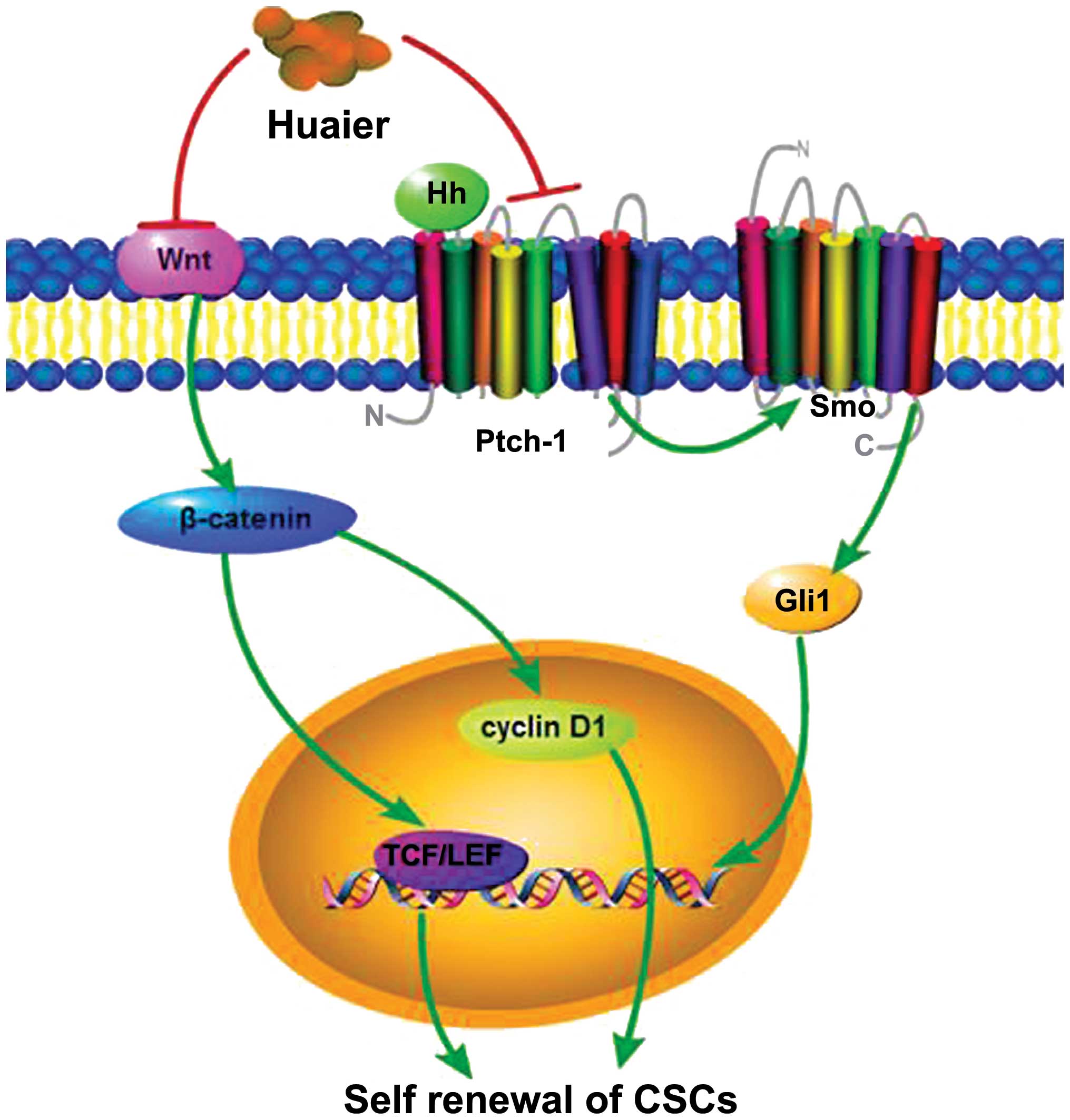
The underlying mechanism of the inhibitory effect of
Huaier on CSCs was subsequently studied, among which downregulation
of the Wnt/β-catenin pathway and inhibition of the hedgehog (Hh)
pathway were highlighted (Fig. 4).
The Wnt/β-catenin pathway is one of the critical pathways
demonstrated to mediate the self-renewal of CSCs (74). The activation of Wnt target genes
depends on mediation by β-catenin, which enters the nucleus to
activate the TCF/LEF transcription factor (74). Treatment of the CRC cells with
Huaier led to dose-dependent downregulation of the levels of total
β-catenin protein, decreased activation of TCF/LEF in the nucleus
and reduced expression of cyclin D1, one of the Wnt/β-catenin
downstream genes (72). Therefore,
the downregulation of the Wnt/β-catenin self-renewal pathway may be
a potential target of the Huaier extract. The inhibitory effect of
Huaier extract on CSCs was also found to be partly dependent on the
inactivation of the Hh pathway which is associated with both normal
mammary gland development and breast cancer progression (73,75).
After treatment with Huaier extract, the expression of Gli1 was
obviously declined, showing that Huaier extract was highly
effective to eradicate breast (CSCs) through regulating Hh
signaling (73). These results
identified Huaier as an effective agent of great clinical value to
eradicate CSCs and to improve current cancer treatment.
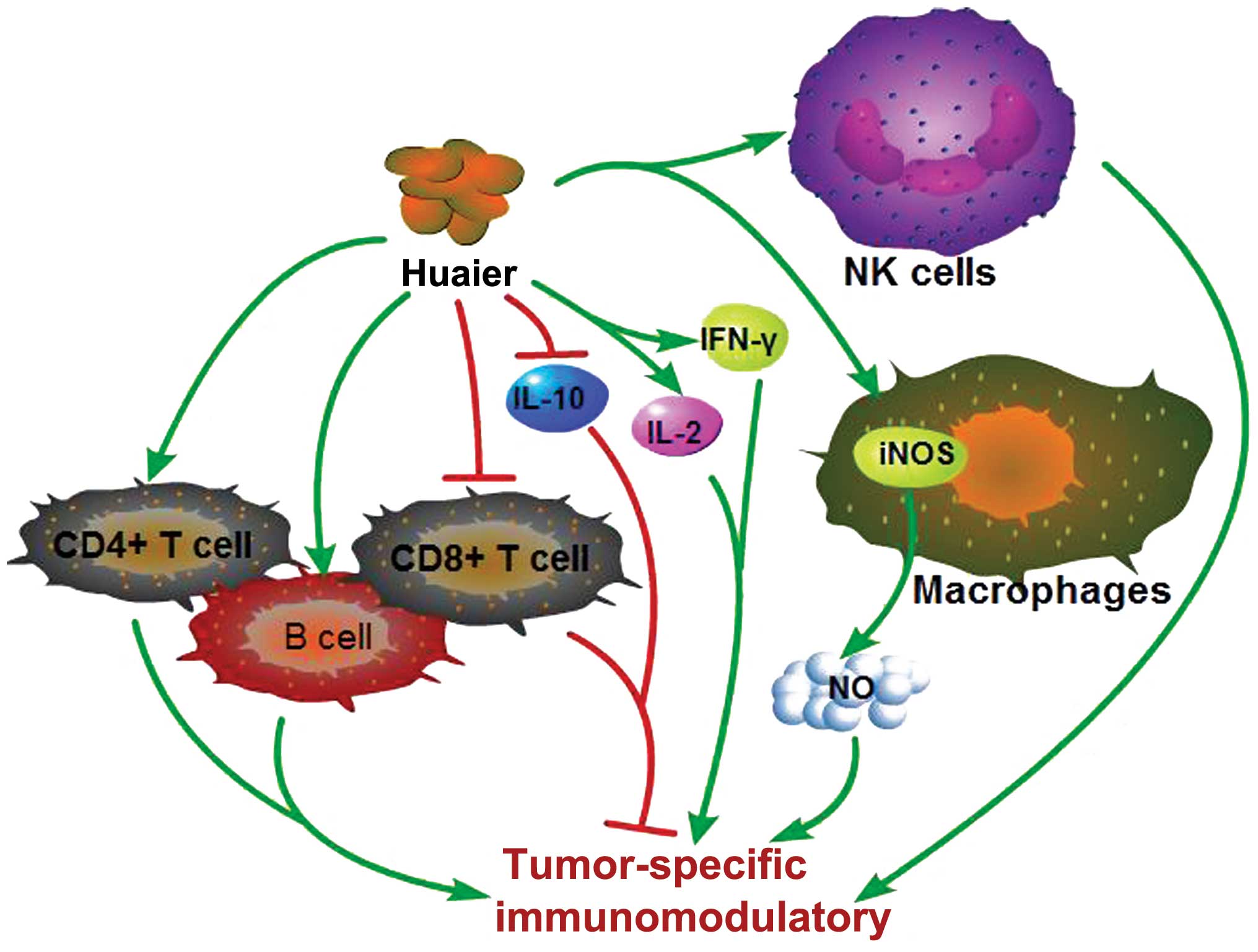
7. Tumor-specific immunomodulatory effect of
Huaier
Tumor growth and the immune system are intertwined
in a complex competition where tilting the subtle balance between
tumor-specific immunity and tolerance can ultimately decide the
fate of the host (76). Recently,
experimental and clinical testing of novel types of
immunotherapeutic agents have been rapidly developing due to the
advances in tumor immunology and a better understanding of the
mechanisms regulating the immune response (77). The water extract from Huaier
consists mainly of polysaccharide protein which has been proved to
be of great clinical value as the main active ingredient in the
anticancer effects and immunity-enhancing actions of Huaier
(Fig. 5) (16). Lymphocytes are the key effector
cells of the mammalian immune system, whose proliferation is
considered an indicator of immunopotentiation. W-NTRP (a neutral
water-soluble polysaccharide isolated from the fruit bodies of
Huaier) activated different subpopulations of lymphocytes and was
suggested to be a potent immunomodulating and immunoenhancing agent
(16). TP-1 (a Huaier
polysaccharide) administered at two doses (50 or 100 mg/kg)
significantly prompted the T- or B-lymphocyte proliferation induced
by ConA or LPS, respectively, when compared to those in H22-bearing
mice (Kunming mice weekly transplanted with murine H22 ascitic
hepatoma cells into the peritoneal cavities). Li et al
(78) also noted increased
CD4+ T cells and decreased CD8+ T cells
induced by TP-1 in tumor-bearing mice (78). Macrophages and NK cells are capable
of inducing the death of tumor cells and play essential roles in
host anticancer immune response (79). Macrophages are an essential
component of the host defense against tumor growth by releasing
various cell factors, such as TNF-α, NO, reactive oxygen
intermediates (ROI) and other substances to kill tumor cells
(80). It was reported that W-NTRP
significantly enhanced macrophage phagocytosis, increasing NO
production and iNOS activity (16).
An increased quantity of NK cells and enhanced NK cell activity
were detected after exposure to Huaier polysaccharide (18,78).
Huaier polysaccharide (TP-1) also modulated the cytokine release,
as confirmed by the increased IFN-γ and IL-2 expression and
inhibition of IL-10 expression. Taken together, these results
suggest that the polysaccharide isolated from Huaier has an
immunoregulatory effect and potent antitumor activities with great
clinical value.
8. Therapeutic perspectives and
conclusions
Huaier, as a type of officinal fungi, has been used
in China for nearly 1,600 years, and has been reported to exert
potent anticancer activities. This review aimed at obtaining a
clear picture regarding the anticancer effects of Huaier and the
underlying mechanisms. The anticancer potential of Huaier,
including proliferation inhibitory, anti-metastasis,
anti-angiogenesis, apoptosis induction, tumor-specific
immunomodulatory and cancer stem cell inhibitory activities, have
been validated in in vitro studies and in various animal
models, indicating its potential therapeutic value against various
types of cancers. Another encouraging finding was that Huaier may
serve as an efficient anticancer agent with little toxicity.
Preliminary toxicological evaluation demonstrated that TP-1 (a
Huaier polysaccharide) had no obvious systemic toxicity on the
kidney and liver of tumor-bearing mice at the therapeutic dose (25
and 50 mg/kg). This was confirmed by the results that the
tumor-caused changes in hepatic function markers (ALT and AST) and
renal function parameters (BUN, UA, and CRE) (78) were ameliorated, or even restored to
the normal level after TP-1 treatment. Xenograft experiments in
mice were also performed. The weight of tumors isolated from the
Huaier-treated groups (at concentration of 2.5 g/kg/day) was
significantly decreased compared to the control group with no
significant difference in body weight, which indicated no obvious
toxicity to mice at the curative dose (17). Several molecular targets and classic
pathways of Huaier which play an important role in the development
of cancer have also been identified. These results highlight the
possible application of Huaier in cancer chemoprevention and lay a
solid foundation for its clinical use in humans, opening a more
effective anticancer treatment option even though most of the
research is still in an experimental stage. We hope and believe
that Huaier may provide a benefit in the treatment of malignant
tumors as a prospective anticancer drug candidate.
Although the findings documented in this review are
quite encouraging for the use of Huaier as a novel anticancer
agent, several limitations exist in the current scenario. Most of
the studies were conducted using Huaier aqueous extract, and the
observed biological effects could be due to the combined effects of
various components. Thus, further research on the specific
effective anticancer components of Huaier is warranted. Although
animal and experimental studies have been successfully carried out
showing Huaier’s efficacy as an anticancer agent, no information on
the absorption, distribution, metabolism, and excretion of Huaier
in humans, or clinical trial was found in the search of the
available literature. Therefore, clinical studies should be carried
out to illustrate the untapped chemopreventive and therapeutic
potential of Huaier either alone or in conjunction with existing
therapies.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (nos. 81272903, 81172529 and
81072150), and the Shandong Science and Technology Development Plan
(nos. 2012GZC22115 and 2013GRC31801) to Q.Y.
References
|
1
|
Winter-Roach B, Hooper L and Kitchener H:
Systematic review of adjuvant therapy for early stage (epithelial)
ovarian cancer. Int J Gynecol Cancer. 13:395–404. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wolpin BM, Meyerhardt JA, Mamon HJ and
Mayer RJ: Adjuvant treatment of colorectal cancer. CA Cancer J
Clin. 57:168–185. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Macdonald JS, Smalley SR, Benedetti J,
Hundahl SA, Estes NC, Stemmermann GN, Haller DG, Ajani JA,
Gunderson LL, Jessup JM and Martenson JA: Chemoradiotherapy after
surgery compared with surgery alone for adenocarcinoma of the
stomach or gastroesophageal junction. New Engl J Med. 345:725–730.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liao WC, Chien KL, Lin YL, Wu MS, Lin JT,
Wang HP and Tu YK: Adjuvant treatments for resected pancreatic
adenocar-cinoma: a systematic review and network meta-analysis.
Lancet Oncol. 14:1095–1103. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wirth T, Parker N and Ylä-Herttuala S:
History of gene therapy. Gene. 525:162–169. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Parums DV: Current status of targeted
therapy in non-small cell lung cancer. Drugs Today (Barc).
50:503–525. 2014. View Article : Google Scholar
|
|
7
|
Pan MH and Ho CT: Chemopreventive effects
of natural dietary compounds on cancer development. Chem Soc Rev.
37:2558–2574. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kannaiyan R, Shanmugam MK and Sethi G:
Molecular targets of celastrol derived from Thunder of God Vine:
potential role in the treatment of inflammatory disorders and
cancer. Cancer Lett. 303:9–20. 2011. View Article : Google Scholar
|
|
9
|
Yu J, Liu H, Lei J, Tan W, Hu X and Zou G:
Antitumor activity of chloroform fraction of Scutellaria barbata
and its active constituents. Phytotherapy Res. 21:817–822. 2007.
View Article : Google Scholar
|
|
10
|
Harhaji Lj, Mijatović S, Maksimović-Ivanić
D, Stojanović I, Momcilović M, Maksimović V, Tufegdzić S,
Marjanović Z, Mostarica-Stojković M, Vucinić Z, et al: Anti-tumor
effect of Coriolus versicolor methanol extract against mouse B16
melanoma cells: in vitro and in vivo study. Food Chem Toxicol.
46:1825–1833. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rashid S, Unyayar A, Mazmanci MA, McKeown
SR, Banat IM and Worthington J: A study of anti-cancer effects of
Funalia trogii in vitro and in vivo. Food Chem Toxicol.
49:1477–1483. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
McCulloch M, See C, Shu XJ, Broffman M,
Kramer A, Fan WY, Gao J, Lieb W, Shieh K and Colford JM Jr:
Astragalus-based Chinese herbs and platinum-based chemotherapy for
advanced non-small-cell lung cancer: meta-analysis of randomized
trials. J Clin Oncol. 24:419–430. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li LX, Ye SL, Wang YH and Tang ZZ:
Progress on experimental research and clinical application of
Trametes robiniophila. Bull Chin Cancer. 16:110–113. 2007.
|
|
14
|
Wang X, Zhang N, Huo Q and Yang Q:
Anti-angiogenic and antitumor activities of Huaier aqueous extract.
Oncol Rep. 28:1167–1175. 2012.PubMed/NCBI
|
|
15
|
Wu T, Chen W, Liu S, Lu H, Wang H, Kong D,
Huang X, Kong Q, Ning Y and Lu Z: Huaier suppresses proliferation
and induces apoptosis in human pulmonary cancer cells via
upregulation of miR-26b-5p. FEBS Lett. 588:2107–2114. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun Y, Sun T, Wang F, Zhang J, Li C, Chen
X, Li Q and Sun S: A polysaccharide from the fungi of Huaier
exhibits anti-tumor potential and immunomodulatory effects.
Carbohydr Polym. 92:577–582. 2013. View Article : Google Scholar
|
|
17
|
Yan X, Lyu T, Jia N, Yu Y, Hua K and Feng
W: Huaier aqueous extract inhibits ovarian cancer cell motility via
the AKT/GSK3beta/beta-catenin pathway. PloS One. 8:e637312013.
View Article : Google Scholar
|
|
18
|
Zheng J, Li C, Wu X, Liu M, Sun X, Yang Y,
Hao M, Sheng S, Sun Y, Zhang H, et al: Astrocyte elevated gene-1
(AEG-1) shRNA sensitizes Huaier polysaccharide (HP)-induced
anti-metastatic potency via inactivating downstream P13K/Akt
pathway as well as augmenting cell-mediated immune response. Tumour
Biol. 35:4219–4224. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang N, Kong X, Yan S, Yuan C and Yang Q:
Huaier aqueous extract inhibits proliferation of breast cancer
cells by inducing apoptosis. Cancer Sci. 101:2375–2383. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang F, Zhang Z and Liu Z: Effects of
Huaier aqueous extract on proliferation and apoptosis in the
melanoma cell line A875. Acta Histochem. 115:705–711. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zheng J, Li C, Wu X, Liu M, Sun X, Yang Y,
Hao M, Sheng S, Sun Y, Zhang H, et al: Huaier polysaccharides
suppresses hepatocarcinoma MHCC97-H cell metastasis via
inactivation of EMT and AEG-1 pathway. Int J Biol Macromol.
64:106–110. 2014. View Article : Google Scholar
|
|
22
|
Pflaum J, Schlosser S and Müller M: p53
Family and cellular stress responses in cancer. Front Oncol.
4:2852014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Advani AS, Mahfouz RZ, Maciejewski J,
Rybicki L, Sekeres M, Tripp B, Kalaycio M, Bates J and
Saunthararajah Y: Ribosomal S6 kinase and AKT phosphorylation as
pharmacodynamic biomarkers in patients with myelodysplastic
syndrome treated with RAD001. Clin Lymphoma Myeloma Leuk.
14:172–177. 2014. View Article : Google Scholar
|
|
24
|
Magnuson B, Ekim B and Fingar DC:
Regulation and function of ribosomal protein S6 kinase (S6K) within
mTOR signalling networks. Biochem J. 441:1–21. 2012. View Article : Google Scholar
|
|
25
|
Hedden A, Müller V and Jensen EV: A new
interpretation of antiestrogen action. Ann NY Acad Sci.
761:109–120. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Klinge CM: Estrogen receptor interaction
with estrogen response elements. Nucleic Acids Res. 29:2905–2919.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang X, Zhang N, Huo Q, Sun M, Lv S and
Yang Q: Huaier aqueous extract suppresses human breast cancer cell
proliferation through inhibition of estrogen receptor α signaling.
Int J Oncol. 43:321–328. 2013.PubMed/NCBI
|
|
28
|
Shen HM and Tergaonkar V: NFkappaB
signaling in carcinogenesis and as a potential molecular target for
cancer therapy. Apoptosis. 14:348–363. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Barker N and Clevers H: Mining the Wnt
pathway for cancer therapeutics. Nat Rev Drug Discov. 5:997–1014.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang L, Cao XX, Chen Q, Zhu TF, Zhu HG and
Zheng L: DIXDC1 targets p21 and cyclin D1 via PI3K pathway
activation to promote colon cancer cell proliferation. Cancer Sci.
100:1801–1808. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
MacDonald BT, Tamai K and He X:
Wnt/beta-catenin signaling: components, mechanisms, and diseases.
Dev Cell. 17:9–26. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jin X, Zhu Z and Shi Y: Metastasis
mechanism and gene/protein expression in gastric cancer with
distant organ metastasis. Bull Cancer. Oct 8–2014.Epub ahead of
print.
|
|
33
|
Song IH: Cancer metastasis and metastasis
suppressors. Korean J Gastroenterol. 43:1–7. 2004.In Korean.
PubMed/NCBI
|
|
34
|
Grinshpun A, Ben-Porath I, Peretz T and
Salmon A: Tumor, metastasis and what’s in between. Harefuah.
152:30–33. 59582013.
|
|
35
|
Bajpai S, Feng Y, Wirtz D and Longmore GD:
β-Catenin serves as a clutch between low and high intercellular
E-cadherin bond strengths. Biophys J. 105:2289–2300. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu H, Xu GH, Wang K, Cao JL, Gu EW, Li YH
and Liu XS: Involvement of GSK3β/β-catenin signaling in the
impairment effect of ketamine on spatial memory consolidation in
rats. Neurobiol Learn Mem. 111:26–34. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yuan L, Shi RR, Rao SM, Song JL and Cui
MC: Reversal of resistance to adriamycin in human breast cancer
cell line MCF-7/ADM by silencing AEG-1 gene and its mechanism.
Sheng Li Xue Bao. 66:625–630. 2014.In Chinese. PubMed/NCBI
|
|
38
|
Ke ZF, Mao X, Zeng C, He S, Li S and Wang
LT: AEG-1 expression characteristics in human non-small cell lung
cancer and its relationship with apoptosis. Med Oncol. 30:3832013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hu G, Wei Y and Kang Y: The multifaceted
role of MTDH/AEG-1 in cancer progression. Clin Cancer Res.
15:5615–5620. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Risau W: Mechanisms of angiogenesis.
Nature. 386:671–674. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Carmeliet P and Jain RK: Angiogenesis in
cancer and other diseases. Nature. 407:249–257. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tassi E and Wellstein A: Tumor
angiogenesis: initiation and targeting – therapeutic targeting of
an FGF-binding protein, an angiogenic switch molecule, and
indicator of early stages of gastrointestinal adenocarcinomas.
Cancer Res Treat. 38:189–197. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hanahan D and Folkman J: Patterns and
emerging mechanisms of the angiogenic switch during tumorigenesis.
Cell. 86:353–364. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bonapace L, Coissieux MM, Wyckoff J, Mertz
KD, Varga Z, Junt T and Bentires-Alj M: Cessation of CCL2
inhibition accelerates breast cancer metastasis by promoting
angiogenesis. Nature. 515:130–133. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Phoophitphong T, Hanprasertpong J,
Dechsukhum C and Geater A: Correlation of angiogenesis and
recurrence-free survival of early stage cervical cancer patients
undergoing radical hysterectomy with pelvic lymph node dissection.
J Obstet Gynaecol Res. 33:840–848. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Folkman J: Tumor angiogenesis: therapeutic
implications. N Engl J Med. 285:1182–1186. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tang HS, Feng YJ and Yao LQ: Angiogenesis,
vasculogenesis, and vasculogenic mimicry in ovarian cancer. Int J
Gynecol Cancer. 19:605–610. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Giuliano S and Pagès G: Mechanisms of
resistance to anti-angiogenesis therapies. Biochimie. 95:1110–1119.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Neufeld G, Cohen T, Gengrinovitch S and
Poltorak Z: Vascular endothelial growth factor (VEGF) and its
receptors. FASEB J. 13:9–22. 1999.PubMed/NCBI
|
|
50
|
Paley PJ, Staskus KA, Gebhard K, Mohanraj
D, Twiggs LB, Carson LF and Ramakrishnan S: Vascular endothelial
growth factor expression in early stage ovarian carcinoma. Cancer.
80:98–106. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Chen P, Zhu J, Liu DY, Li HY, Xu N and Hou
M: Overexpression of survivin and VEGF in small-cell lung cancer
may predict the poorer prognosis. Med Oncol. 31:7752014. View Article : Google Scholar
|
|
52
|
Ren J, Zheng C, Feng G, Liang H, Xia X,
Fang J, Duan X and Zhao H: Inhibitory effect of extract of fungi of
Huaier on hepatocellular carcinoma cells. J Huazhong Univ Sci
Technolog Med Sci. 29:198–201. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wyllie AH, Kerr JF and Currie AR: Cell
death: the significance of apoptosis. Int Rev Cytol. 68:251–306.
1980. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Adams JM and Cory S: The Bcl-2 apoptotic
switch in cancer development and therapy. Oncogene. 26:1324–1337.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lowe SW and Lin AW: Apoptosis in cancer.
Carcinogenesis. 21:485–495. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Green DR, Galluzzi L and Kroemer G: Cell
biology. Metabolic control of cell death. Science. 345:12502562014.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Degli Esposti M: Mitochondria in
apoptosis: past, present and future. Biochem Soc Trans. 32:493–495.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Costa M, Costa-Rodrigues J, Fernandes MH,
Barros P, Vasconcelos V and Martins R: Marine cyanobacteria
compounds with anticancer properties: a review on the implication
of apoptosis. Mar Drugs. 10:2181–2207. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Huang Y, Yang YB, Zhang XH, Yu XL, Wang ZB
and Cheng XC: MicroRNA-21 gene and cancer. Med Oncol. 30:3762013.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Solomides CC, Evans BJ, Navenot JM,
Vadigepalli R, Peiper SC and Wang ZX: MicroRNA profiling in lung
cancer reveals new molecular markers for diagnosis. Acta Cytol.
56:645–654. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Liu XX, Li XJ, Zhang B, Liang YJ, Zhou CX,
Cao DX, He M, Chen GQ, He JR and Zhao Q: MicroRNA-26b is
underexpressed in human breast cancer and induces cell apoptosis by
targeting SLC7A11. FEBS Lett. 585:1363–1367. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Koh CM, Iwata T, Zheng Q, Bethel C,
Yegnasubramanian S and De Marzo AM: Myc enforces overexpression of
EZH2 in early prostatic neoplasia via transcriptional and
post-transcriptional mechanisms. Oncotarget. 2:669–683.
2011.PubMed/NCBI
|
|
63
|
Yuan JB, Yang LY, Tang ZY, Zu XB and Qi L:
Down-regulation of EZH2 by RNA interference inhibits proliferation
and invasion of ACHN cells via the Wnt/β-catenin pathway. Asian Pac
J Cancer Prev. 13:6197–6201. 2012. View Article : Google Scholar
|
|
64
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Dalerba P, Cho RW and Clarke MF: Cancer
stem cells: models and concepts. Annu Rev Med. 58:267–284. 2007.
View Article : Google Scholar
|
|
66
|
Ricci-Vitiani L, Lombardi DG, Pilozzi E,
Biffoni M, Todaro M, Peschle C and De Maria R: Identification and
expansion of human colon-cancer-initiating cells. Nature.
445:111–115. 2007. View Article : Google Scholar
|
|
67
|
Rycaj K and Tang DG: Cancer stem cells and
radioresistance. Int J Radiat Biol. 90:615–621. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Chen K, Huang YH and Chen JL:
Understanding and targeting cancer stem cells: therapeutic
implications and challenges. Acta Pharmacol Sin. 34:732–740. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Beier D, Schulz JB and Beier CP:
Chemoresistance of glioblastoma cancer stem cells--much more
complex than expected. Mol Cancer. 10:1282011. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Tsujii M: Cancer therapy targeting cancer
stem cell. Nihon rinsho. 72:35–41. 2014.In Japanese. PubMed/NCBI
|
|
71
|
Bao B, Li Y, Ahmad A, Azmi AS, Bao G, Ali
S, Banerjee S, Kong D and Sarkar FH: Targeting CSC-related miRNAs
for cancer therapy by natural agents. Curr Drug Targets.
13:1858–1868. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhang T1, Wang K, Zhang J, Wang X, Chen Z,
Ni C, Qiu F and Huang J: Huaier aqueous extract inhibits colorectal
cancer stem cell growth partially via downregulation of the
Wnt/β-catenin pathway. Oncol Lett. 5:1171–1176. 2013.PubMed/NCBI
|
|
73
|
Wang X, Zhang N, Huo Q, Sun M, Dong L,
Zhang Y, Xu G and Yang Q: Huaier aqueous extract inhibits stem-like
characteristics of MCF7 breast cancer cells via inactivation of
hedgehog pathway. Tumour Biol. 35:10805–10813. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Clevers H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Gangopadhyay S, Nandy A, Hor P and
Mukhopadhyay A: Breast cancer stem cells: a novel therapeutic
target. Clin Breast Cancer. 13:7–15. 2013. View Article : Google Scholar
|
|
76
|
Townsend KN, Hughson LR, Schlie K, Poon
VI, Westerback A and Lum JJ: Autophagy inhibition in cancer
therapy: metabolic considerations for antitumor immunity. Immunol
Rev. 249:176–194. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Ladányi A and Balatoni T: Unblocking
antitumor immune response: novel possibilities for the
immunotherapy of melanoma. Magy Onkol. 57:100–107. 2013.In
Hungarian.
|
|
78
|
Li C, Wu X, Zhang H, Yang G, Hao M, Sheng
S, Sun Y, Long J, Hu C, Sun X, et al: A Huaier polysaccharide
inhibits hepatocel-lular carcinoma growth and metastasis. Tumour
Biol. Nov 6–2014.Epub ahead of print.
|
|
79
|
Stuelp-Campelo PM, de Oliveira MB, Leão
AM, Carbonero ER, Gorin PA and Iacomini M: Effect of a soluble
alpha-D-glucan from the lichenized fungus Ramalina celastri on
macrophage activity. Int Immunopharmacol. 2:691–698. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Fidler IJ and Kleinerman ES: Therapy of
cancer metastasis by systemic activation of macrophages: from the
bench to the clinic. Res Immunol. 144:284–287. 1993. View Article : Google Scholar : PubMed/NCBI
|