Introduction
As one of the three most common cancers in the
world, colorectal cancer (CRC) occurs with a global incidence
exceeding 1.2 million new cases and 600,000 deaths per year.
Endoscopic removal of precursor lesions appears to be the only
effective treatment strategy for CRC (1). The most frequently used chemotherapy
regimens for CRC to date are respective combinations of leucovorin,
oxaliplatin, irinotecan and fluorouracil (5-FU), which are not
highly efficient target therapies, and are accompanied by a series
of adverse side effects, such as peripheral nerve toxicity and
severe neutrophil reduction (2,3). It is
essential to identify and verify potential endogenic biomarkers for
the diagnosis, therapy and further prevention of CRC. Early-stage
CRC is usually misdiagnosed as chronic appendicitis or
cholelithiasis. The carcinoembryonic antigen (CEA) level in patient
blood is a common chemical marker for the clinical diagnosis of
CRC, yet a high CEA level often suggests that tumors have spread to
distant sites. Therefore, it is necessary to explore potential
therapeutic biomarkers and targets which may be effective with
fewer side effects. Here, we suggest that endogenetic cannabinoids
and ceramides may be a possible option.
As two major endocannabinoids, anandamide (AEA) was
first isolated, followed by 2-AG (4,5), and
both are found within the gastrointestinal tract (6).
N-acyl-phosphatidylethanolamine-hydrolyzing phospholipase D
(NPLD) is a key enzyme that participates in a series of AEA
synthesis reactions, and synthesizes and releases AEA from the
direct biosynthetic precursor
N-arachidonoylphosphatidylethanolamine (NAPE) (7,8). Fatty
acid amide hydrolase (FAAH) acts as the main hydrolytic enzyme for
AEA, which is widely distributed in tissues containing the CB1 and
CB2 receptors (9), and mainly
catalyzes AEA into arachidonic acid (AA) and ethanolamine.
Diacylglycerol lipase-α (DGL-α) and monoacylglycerol lipase (MGL)
are respectively responsible for 2-AG biosynthesis and hydrolysis
(8). Both AEA and 2-AG have been
verified to bind to CB receptors. The CB1 receptor is not only
distributed in regions of the brain for central nerve system
control, but is also present in the prostate, spleen, lymphocytes
and small intestine (10,11). The CB2 receptor is mainly found in
immune tissues and immunocompetent blood cells, and is highly
expressed in natural killer cells, mast cells and monocytes
(11). AEA seems to have more
affinity for the CB1 receptor than CB2, but 2-AG prefers to bind to
the CB2 receptor (12,13). Recently, a series of studies
investigated the antineoplastic role of endocannabinoids (14). AEA efficaciously inhibited the
proliferation of multiple tumor cell lines with CB
receptor-mediated antitumor effects, including human breast,
prostatic and colorectal cancer cell lines (15–17).
De Petrocellis et al reported that AEA prevented the
proliferation of cancer cells by inhibiting DNA synthesis and
arresting the G1/S transition of the cell cycle, which was mainly
mediated via the interaction with the transmembrane CB1 receptor
rather than the CB2 receptor (16).
Furthermore, the pro-apoptotic effect of AEA was also realized by
acting on the vanilloid receptors (18). Notably, another pro-apoptotic
endogenic lipid, ceramide, was also confirmed to be elevated and to
participate in apoptosis via the endocannabinoid system (16,19).
Ceramides are vital signaling sphingolipids involved
in cell growth arrest, differentiation and apoptosis, and contain
two frame structures, a sphinganine base and a fatty acyl with
varying lengths of carbon chains (20). The most abundant ceramides in human
tissues are those with C16 to C24 fatty acyl bases, which are
mainly synthesized by six ceramide synthases (CerSs) (CerS1–CerS6).
CerS1, CerS2, CerS5 and CerS6 are relatively plentiful in colon
tissues. CerSs show different specificity to produce ceramides with
differing chain lengths. CerS1 is mainly responsible for C18
ceramides, and CerS2 for C20–C24 ceramides, CerS5 for C16 ceramides
and CerS6 for C16/18 ceramides, respectively (21). Ceramidase inhibition or application
of ceramide analogues promoted the apoptosis of human CRC cells
(22). However, pharmacological
inhibition of ceramide biosynthesis protected cancer cells from
apoptosis (23). In addition,
cancer cell lines with low ceramide levels or a defect in ceramide
synthesis can escape from apoptosis induced by radiation (24). One study found that patients with
head and neck cancer resistant to radiotherapy had decreased
ceramide levels in tumor tissues (25). Higher or lower levels of ceramide
appear to be closely linked to cancer cell apoptosis or malignant
progression. Hence, in addition to endocannabinoids, ceramides may
be another group of important lipids involved in CRC
progression.
In order to reveal the expression patterns of
endocannabinoids and ceramides in human CRC, we evaluated the
endogenic levels of AEA, 2-AG, ceramides and their metabolites via
liquid chromatography-tandem mass spectrometry (LC-MS/MS). The
expression levels of two cannabinoid receptors and the relevant
enzymes, such as NPLD, FAAH, DGL-α, MGL and CerSs, were also
analyzed to evaluate the potential effects of these lipids on tumor
treatment.
Materials and methods
Patients and tissue sampling
Colorectal cancer samples were obtained from 47
Chinese patients undergoing surgical resection at the Department of
Colorectal Surgery, Fujian Medical University Union hospital
(Fuzhou, China) during 2011, and were confirmed with TNM stage
according to the World Health Organization TNM staging 7th edition
by pathological analysis. Non-tumor control tissues were resected
from the same patients at the region 5–7 cm above the tumor
position. All tissues were divided into several slices for lipid,
gene and protein analysis, and were freshly frozen and stored at
−80°C immediately after resection. Informed consent was obtained
from each patient before surgery. Study of the resected human
tissues was approved by the Ethics Committee of Fujian Medical
University Union hospital, following clinical registration
guidelines in China.
Lipid extraction and analysis
Lipids were extracted from the tissue samples using
a previously described method with modifications (26,27).
In brief, the samples were homogenized in methanol/H2O
(v/v, 50/50) containing internal standards (C17:1 FAE, C17:0
heptadecanoic acid and C17:0 ceramide) and were then extracted with
chloroform. The organic phase was transferred and dried under
nitrogen flow (N2). After being redissolved with
chloroform, the lipid solution underwent solid-phase extraction,
and was eluted by methanol/chloroform (v/v, 10/90). The elution
containing endocannabinoids and ceramides was dried and
reconstituted with 100 µl of methanol/chloroform (v/v,
75/25), and then 10 µl of this elution was used for
detection using the 3200 Q Trap LC-MS/MS system (Applied
Biosystems, USA) coupling with the 1100-LC system (Agilent, China).
The mobile phase for endocannabinoids and ceramides consisted of
methanol and ultra-pure water (pH 7.4), and the gradient elution
was as follows: 85% methanol was kept for the first 3 min, followed
by a linear gradient from 85 to 100% methanol for 2 min and then
100% methanol was continued for 15 min. The elution condition was
finally returned to 85% methanol at a flow rate of 0.7 ml/min. The
column temperature was maintained at 40°C. Ion detection was
monitored by APCi+-MRM mode. The molecular ions were
monitored at m/z 348.00/62.00 for AEA, m/z
379.10/287.10 for 2-AG, m/z 313.1/62.0 for C17:1 FAE,
m/z 520.4/264.2 for C16:0 ceramide, m/z 534.3/264.2
for C17:0 ceramide, m/z 548.4/264.2 for C18:0 ceramide,
m/z 576.4/264.2 for C20:0 ceramide, m/z 632.4/264.2
for C24:0 ceramide and m/z 630.4/264.2 for C24:1 ceramide.
Elution condition, ion monitor model and the molecular ion
parameters of free fatty acids were previously described in detail
(26).
Assessment of enzyme activity
Analysis of enzyme activity was also carried out
using a previously described method (27). Samples were homogenized in 50 mM
Tris-HCl buffer (pH 7.4) containing 0.32 M sucrose to obtain the
total proteins. For NPLD activity, in 50 mM Tris-HCl buffer (0.1%
Triton X-100 and 1 mM phenylsulphonylfluoride), 100 µg of
sample protein was incubated with 50 µM
1-palmitoyl,2-palmitoyl-sn-glycero-3-phosphoethanolamine-N-heptadecenoyl
(C17:1 NAPE) at 37°C for 30 min, and then 0.2 ml methanol
containing [2H4]-OEA was added to halt the
reaction. DGL-α activity was measured at 37°C for 30 min in 50 mM
Tris-hCl containing 0.1% Triton X-100. The reaction between the
sample proteins and 50 µM diheptadecanoyl-sn-glycerol
as substrates were stopped by 0.2 ml methanol containing C17:1 FAE.
FAAH and MGL activity were measured by incubating the sample
proteins with AEA or 2-oleoylglycerol as respective substrates in
Tris-HCl buffer containing fatty acid-free BSA (0.05%). The
reactions were also terminated by adding 0.2 ml methanol containing
C17:0 heptadecanoic acid. The reaction products of the enzyme
assays above were monitored by LC-MS and the molecular ions were:
m/z 312.1/62.0 for heptadecanoylethanolamide, m/z
330.1/66.0 for [2H4]-OEA, m/z 303 for
arachidonic acid, m/z 281 for oleic acid, m/z 269 for
17:0 heptadecanoic acid, and m/z 367.1 for
1(3)-heptadecanoyl-sn-glycerol (HDG).
Evaluation of protein and mRNA
levels
Protein expression levels of the tissue samples were
evaluated by western blot analysis. First, the concentration of
homogenized tissue protein was measured using the BCA protein assay
kit (Pierce, China). Proteins were separated by sodium dodecyl
sulfate polyacrylamide gel electrophoresis (SDS-PAGE), and then
transferred to PVDF membranes (Amersham Biosciences, China).
Membranes with the proteins were incubated overnight with primary
antibodies against the CB1 receptor (1:500), CB2 receptor (1:500;
Cayman, China) or β-actin at 4°C, followed by incubation with
HRP-conjugated anti-rabbit IgG antibody (1:10,000; Sigma, China)
for 1 h at room temperature. Protein bands were visualized using an
ECL Plus kit (Amersham Biosciences). mRNA expression levels were
measured by RT-qPCR assay. Tissue samples were ground using
ice-cold porcelain mortars with TRIzol to extract total RNA. The
RNA concentration was measured with a spectrophotometer (Beckman
Coulter, USA). Reverse transcription was completed with a ReverTra
Ace qPCR RT kit (Toyobo, Japan) following the manufacturer’s
instructions. Finally quantitative PCR was performed and analyzed
in a 7500 Fast real-time PCR System (Applied Biosystems). The mRNA
expression levels were normalized by housekeeping gene 18S as the
internal standard.
Primers were as follows: NPLD forward primer (F),
5′-TGGCTGGGACACGCG-3′ and reverse primer (R),
5′-GGGATCCGTGAGGAGGATG-3′; FAAH F, 5′-GCCTCAAGGAATGCTTCAGC-3′ and
R, 5′-TGCCCTCATTCAGGCTCAAG-3′; MGL F, 5′-CATGTGGATTCCATGCAGAAAG-3′
and R, 5′-AGGATTGGCAAGAACCAGAGG-3′; DGL-α F,
5′-AGAATGTCACCCTCGGAATGG-3′ and R, 5′-GTGGCTCTCAGCTTGACAAAGG-3′;
CB1 F, 5′-AGCCTCTGGATAACAGCATGG-3′ and R,
5′-AATCTTGACCGTGCTCTTGATG-3′; CB2 F, 5′-CTCAGTGACCAGGTCAAGAAGG-3′
and R, 5′-TTTTGCCTCTGACCCAAGG-3′; CerS1 F,
5′-TTTGGCTCCCGCACAATGT-3′ and R, 5′-AAAAGCGAGATA GAGGTCCTCA-3′;
CerS2 F, 5′-GCTCTTCCTCATCGTTCGATAC-3′ and R,
5′-GTGTAGCCACGTACAGCTCA-3′; CerS5 F, 5′-GCTGCTCTTCGAGCGATTTAT-3′
and R, 5′-CCTCCGATGGCGAAACCAG-3′; CerS6 F,
5′-TTTGGCTCCCGCACAATGT-3′ and R, 5′-AAAAGCGAGATAGAGGTCCTCA-3′; 18S
F, 5′-CAGCCACCCGAGATTGAGCA-3′ and R,
5′-TAGTAGCGACGGGCGGTGTG-3′.
Statistical analysis
Data of the results are expressed as means ± SEM.
Statistical significance was analyzed with one-way variance
analysis (ANOVA) followed by the Bonferroni all pairs test.
Statistical analysis was completed using GraphPad Prism (GraphPad
Software, USA), and the difference was considered statistically
significant at p<0.05.
Results
Characteristics of the patients
Tumor samples were collected from 47 patients with
CRC during surgical removal. Detailed baseline characteristics of
the 47 patients are listed in Table
I. Their median age was 60 years (range, 33 to 88). Sixty four
percent of the patients were male. Approximately 68% of the
patients were at tumor stage T3 with tumor stage T2 and T4,
respectively, consisting of 4 and 28% of the cases. Regarding the
nodal status, the patients were distributed almost equally between
N0 (47%) and N1+ (53%). TNM staging was as follows: stage I, 2%;
stage II, 45%; stage III, 42%; stage IV, 11%. Here, nodal status
was applied to classify all samples because of equalization and
comprehensiveness. The groups were classified as follows: n0 group,
tumors with no lymph node metastasis; N1+ group, tumors with
metastasis of one or more lymph nodes; and N group, adjacent
non-tumor tissues.
 | Table ICharacteristics of the CRC
patients. |
Table I
Characteristics of the CRC
patients.
|
Characteristics | Data |
|---|
| Age (years) |
| Median | 60 |
| Range | 33–88 |
| Gender, n (%) |
| Male | 31 | (66) |
| Female | 16 | (34) |
| Tumor stage, n
(%) |
| T2 | 2 | (4) |
| T3 | 32 | (68) |
| T4 | 13 | (28) |
| Nodal status, n
(%) |
| N0 | 22 | (47) |
| N1+ | 25 | (53) |
| TNM staging, n
(%) |
| I | 1 | (2) |
| II | 21 | (45) |
| III | 20 | (42) |
| IV | 5 | (11) |
Alteration of endogenic lipid levels in
human CRC tissues
After detection by LC-MS/MS assay and normalization
by the wet weight of each sample, the levels of AEA, 2-AG,
ceramides and free fatty acids were analyzed. Regarding the
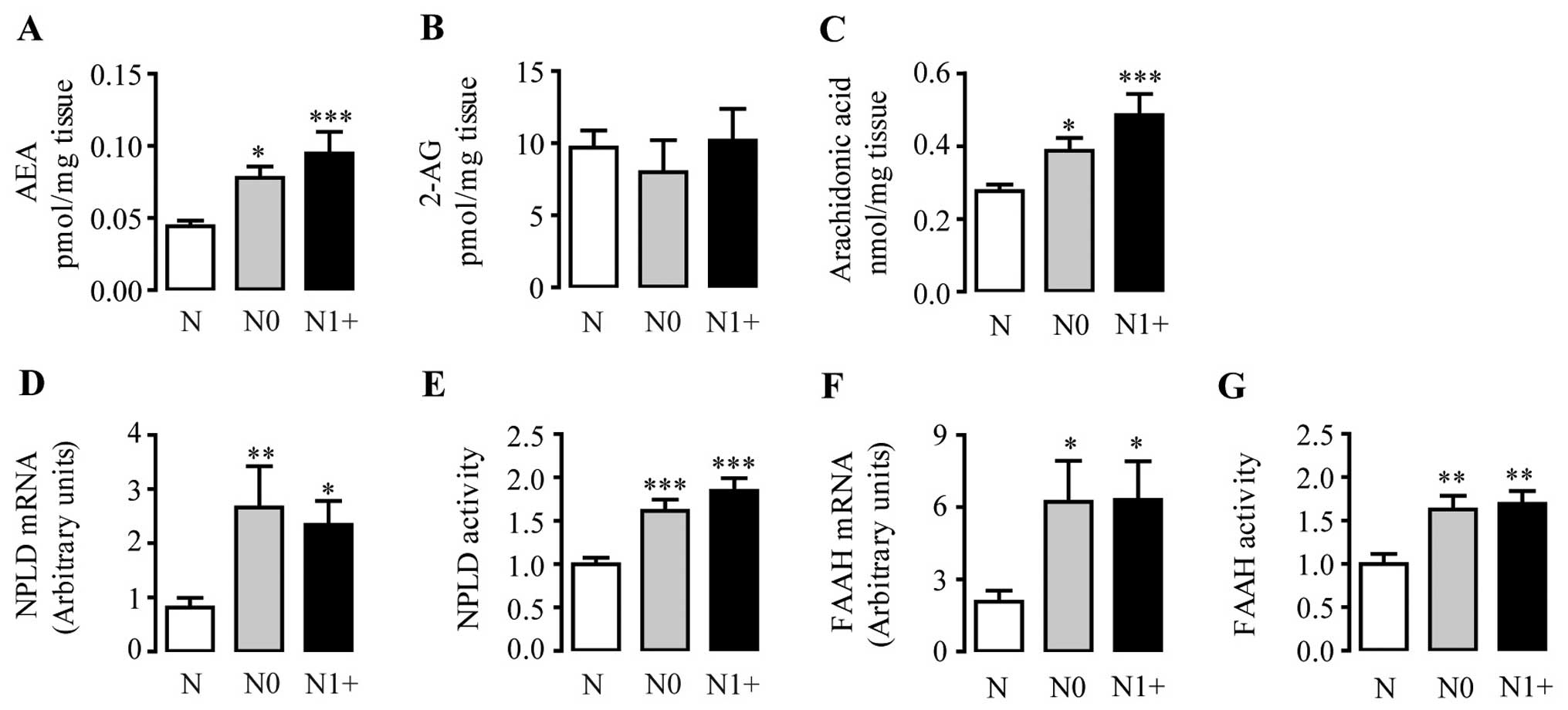
endocanabinoids, the anandamide level was obviously elevated in the
CRC tissues when compared with this level in the non-tumor control
tissues, and was nearly 2-fold higher in the tissues with
metastasis of the lymph nodes (Fig.
1A). However, change in the level of the other endocannabinoid
2-AG was not detected between the tumor tissues and the control
tissues (Fig. 1B). Levels of
long-chain ceramides also showed variation with different trends.
Levels of C16:0 and C24:0 ceramides in human non-tumor colon
tissues were, respectively, 102.6±6.0 and 8.6±0.8 pmol/mg tissue,
and both were obviously increased in the tumor tissues (Fig. 2A-a and -b). A 1.5-fold higher level
of C24:1 ceramide was detected in the tumor tissues with lymph node
metastasis (Fig. 2A-c). Elevation
of C24 ceramide showed a positive correlation with lymph node
invasion. In contrast, reduced levels of both C18 and C20 ceramides
were observed in the tumor tissues as compared with these levels in
the adjacent noncancerous tissues (Fig.
2A-d and -e). Regarding a comprehensive series of multiple free
fatty acids (FFA), levels of saturated fatty acids (SFAs),
monounsaturated fatty acids (MUFAs) and polyunsaturated fatty acids
were also detected using the LC-MS/MS method. Unexpectedly, as the
main metabolic product of AEA, levels of arachidonic acid (C20:4
FFA) were markedly increased in the tumor tissues, and were even
higher in the N1+ group (Fig. 1C).
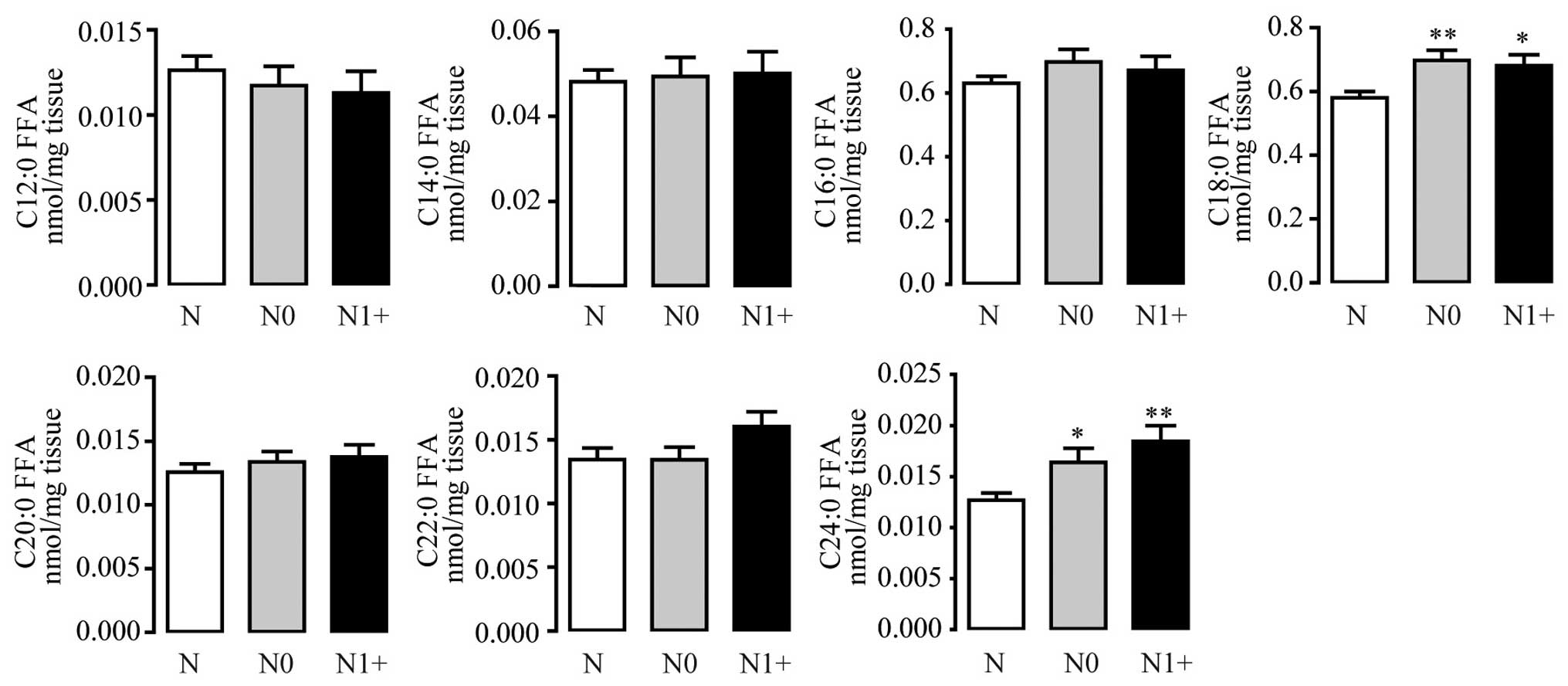
In addition, compared with the non-tumor tissues, most FFAs
presented upregulated trends in the CRC tissues. As for the SFAs,
C18:0 and C24:0 FFAs displayed higher levels in the tumor tissues
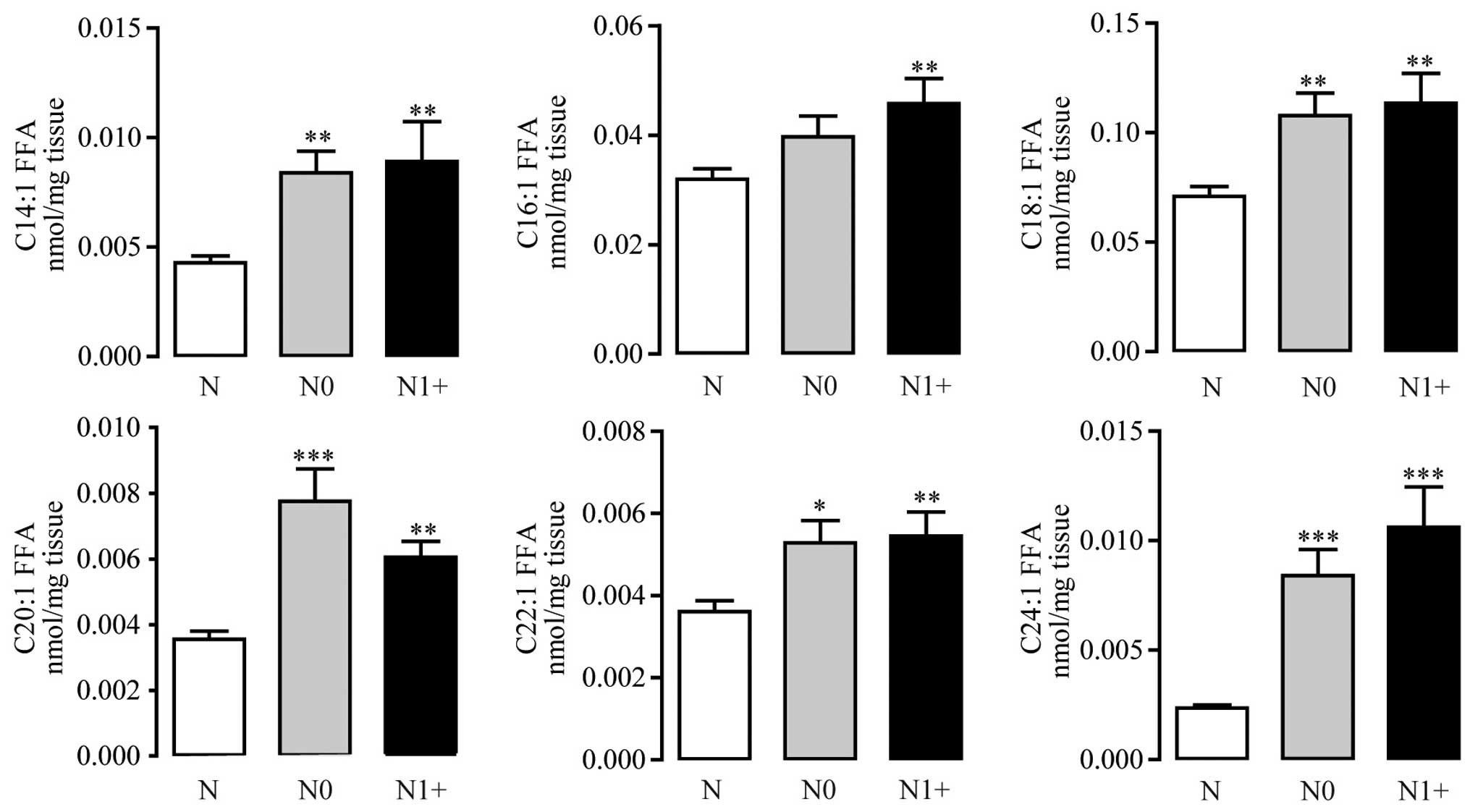
(Fig. 3). MUFAs from C14 to C24
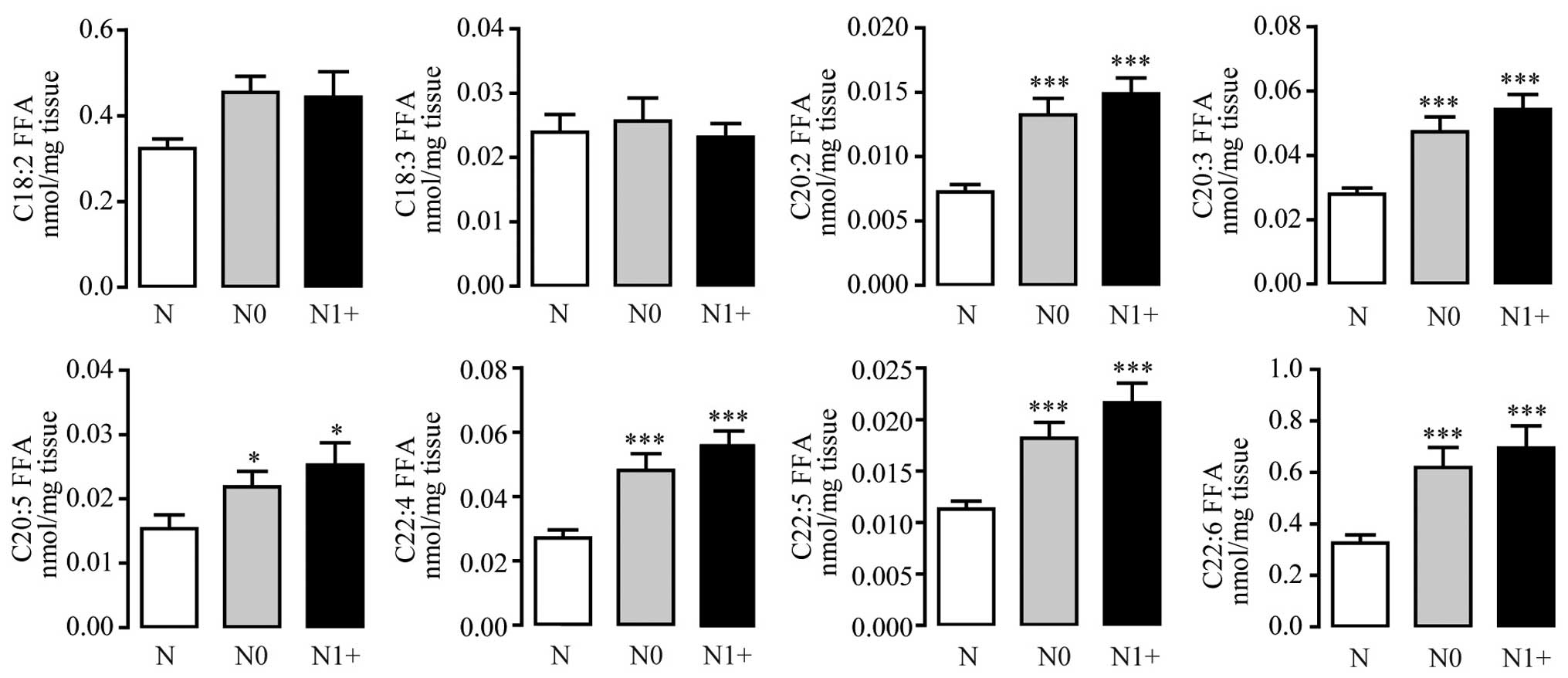
were consistently elevated in the tumor tissues (Fig. 4). Except C18 FFAs, most PUFAs also
revealed highly increasing trends (Fig.
5).
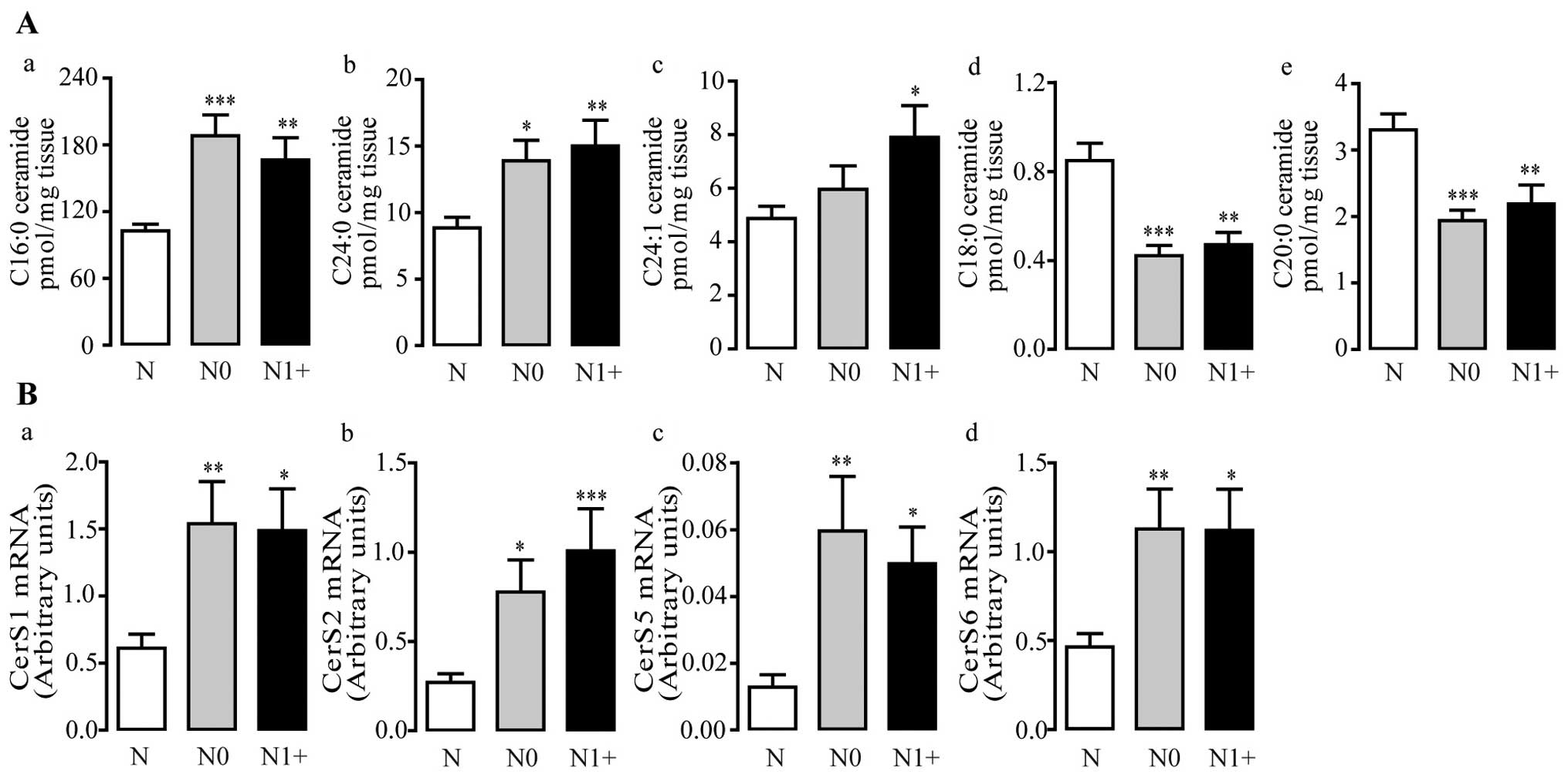 | Figure 2Varied levels of ceramides and
ceramide synthases in the human colorectal cancer tissues. (A)
C16:0, C24:0 and C24:1 ceramides were elevated, and C18:0/C20:0
ceramides were downregulated in the CRC tissues. (B) The mRNA
levels of CerS1, CerS2, CerS5 and CerS6 were upregulated in the CRC
tissues (N0, gray bar, CRC without lymph node metastasis; N1+,
solid bars, CRC with lymph node metastasis) compared with the
non-tumor colon tissue controls (N, open bars).
*p<0.05, **p<0.01,
***p<0.001, n=22–47. |
Changes in metabolic enzymes for
corresponding lipids in human CRC tissues
In order to explain the variation of lipids in human
CRC, mRNA expression levels and activities of metabolic enzymes
were respectively analyzed by quantitative PCR assay and enzyme
activity assay as described above. We first measured the mRnA
expression levels and activities of associative metabolic enzymes
responsible for AEA and 2-AG. As a main synthetase of AEA, NPLD was
2- to 3-fold overexpressed in the tumor tissues; the enzyme
activities of which showed the same upward trend (Fig. 1D and E). Notably, FAAH, a key
hydrolase of AEA, also presented higher mRNA expression levels and
catalytic activities in the CRC tissues (Fig. 1F and G). The transfer enzymes
responsible for 2-AG were further detected in the same manner. The
mRNA levels and catalytic activities of both synthetase DGL-α and
hydrolase MGL exhibited no significant differences between the
tumor and the control tissues. Moreover, to investigate the
different variation trends of long-chain ceramides, the mRNA levels
of four abundant CerSs in the human colon were also measured, that
is CerS1, CerS2, CerS5 and CerS6. The results showed that all CerSs
revealed 2- to 4-fold upward trends (Fig. 2B). CerS1, a synthase responsible for
C18 ceramide synthesis, unexpectedly increased, and the same trends
were also observed for CerS2 and CerS6, which partly synthesize C20
and C18 ceramides. Yet, CerS5 and CerS6 for C16 ceramide showed
consistent upregulation, as did CerS2 which is partly responsible
for C24 ceramide.
Expression levels of cannabinoid
receptors in human CRC tissues
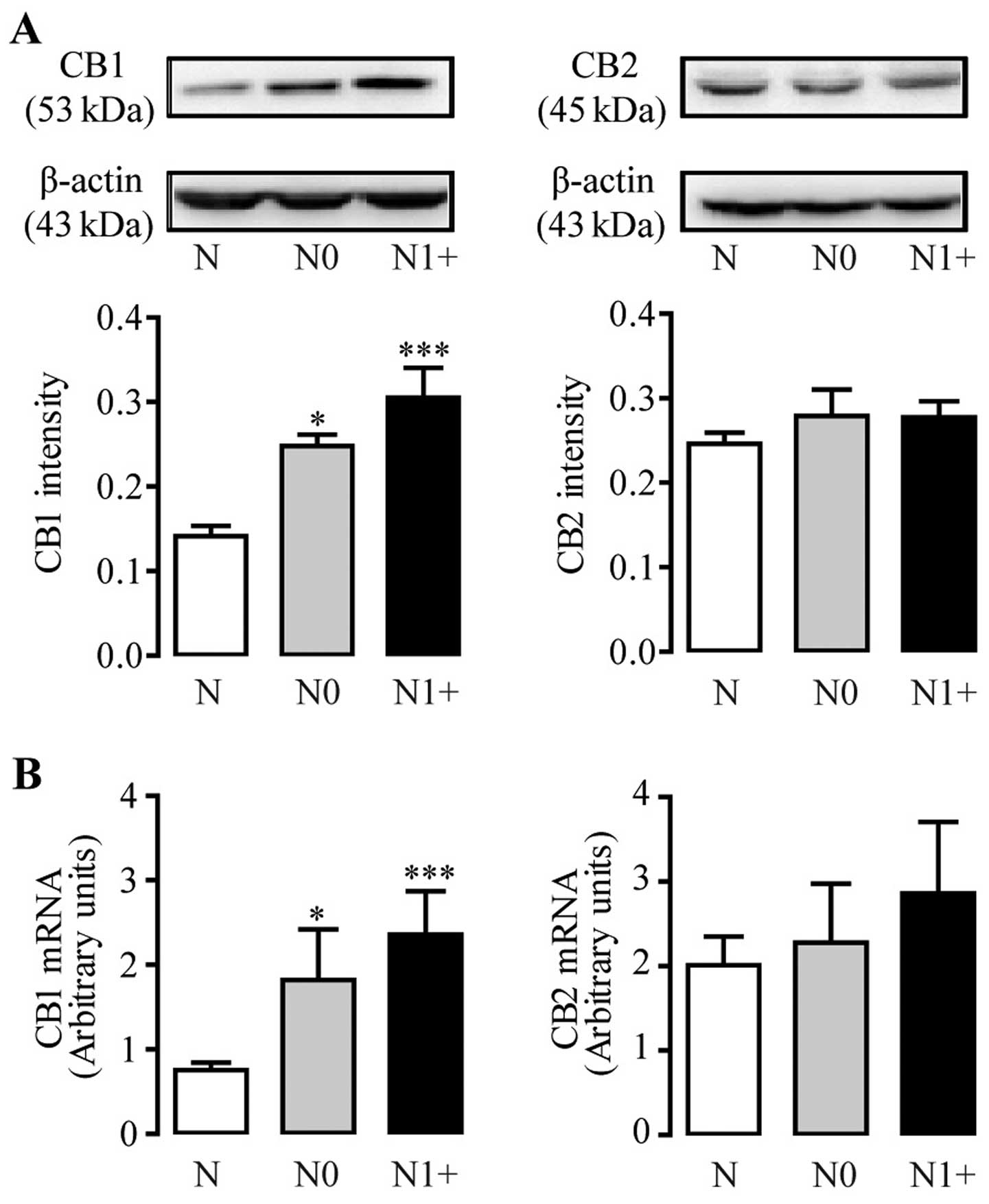
Based on the alteration of endocannabinoid levels in
the CRC tissues, we further analyzed the protein expression levels
of the cannabinoid receptors via western blot analysis in the same
tissue samples. CB1 expression was elevated 1.5-fold in the N0
group (0.25±0.01), and 2-fold in the N1+ group (0.31±0.04), which
were compared with adjacent non-tumor samples (0.14±0.01) (Fig. 6A). However, protein expression of
CB2 showed no obvious alteration between the CRC tissues and
non-tumor tissues (Fig. 6A). The
mRNA levels of cannabinoid receptors were further measured.
Consistent with the protein levels, the mRNA level of CB1 was also
upregulated in the tumor samples, and the gene expression of the
CB2 receptor was not altered (Fig.
6B). The detailed result of the CB1 mRNA levels was as follows:
0.8±0.1 in the N group; 1.8±0.5 in the N0 group; 2.4±0.5 in the N1+
group.
Discussion
Endocannabinoids have been demonstrated to act with
multi-function in the gastrointestinal tract, particularly in the
colon. D’Argenio et al reported that in human colitis and
inflammatory bowel diseases (IBDs), upregulation of AEA levels
exerted a protective effect against the inflammatory response
mainly via the CB1 receptor subtype (28). Since chronic inflammation is a
recognized risk factor for CRC, the key role of the cannabinoid
system in CRC progression warrants further investigation. It was
verified that increased endocannabinoid levels reduced the
development of precancerous lesions in the mouse colon (29). In addition, endocannabinoids were
even found to inhibit the proliferation of cancer cells, such as
breast cancer, prostate cancer and colorectal cancer cells, mostly
via CB1 receptors (15,17,30).
In the present study, we found that the level of AEA
was increased in the CRC tissues, particularly in those with lymph
node metastasis, while 2-AG levels showed no significant
difference. The result of AEA levels was consisting with data
reported by Ligresti et al (15). In contast, 2-AG levels in their
study were also upregulated, which may have been induced by
differences in the sample source, population distribution and
dietary habits. To further verify the detailed pathway of
endocannabinoid alterations, we measured the expression of relevant
metabolic enzymes. In agreement with the marked upward trends for
AEA, the mRNA levels and activities of synthase NPLD were highly
elevated. The same tendency was also observed for FAAH, which acts
as an AEA hydrolase. In the present study, high expression of NPLD
was thought to be the primary reason for elevation of AEA, thereby
FAAH was enhanced as a positive feedback due to substrate
accumulation. As a result, arachidonic acid, the main metabolite of
AEA, was found to increase in the CRC tissues particularly in those
associated with lymph node metastasis. Regarding the metabolic
pathway of 2-AG, there was no significant change in the activities
and the expression of synthase DGL-α and hydrolase MGL between the
tumor and the control tissues, which may be a reasonable
explanation for the consistent levels of 2-AG. Detailed mechanisms
of the variation in endocannabinoid levels still require further
study.
AEA elevation in CRC was suggested to be
self-protection against further tumor progression. It is known that
multiple pharmacological functions of AEA are accomplished through
activation of cannabinoid receptors, particular CB1 (12,13).
In the present study, the mRNA level of CB1 was increased in the
CRC tissues and even higher in the lymph node metastatic tumor
tissues. Western blot analysis further supported the above results,
which was similar to a research study by Cianchi et al
(31). A report by Ligresti et
al demonstrated that activation of the CB1 receptor resulted in
inhibition of CRC cell proliferation in vitro (15), while the CB2 receptor did not
display an obvious change in CRC cells, which is also consistent
with our findings. Based on the present data, we suggest that
strong activity of NPLD produced abundant endogenic AEA, which
provided much more substrates to induce higher levels of FAAh,
thereby sufficient agonist AEA further activated preferable CB1
receptor to prevent tumor progression.
It has been reported that CB1 receptor activation
caused sphingomyelin hydrolysis and rapid ceramide production in
astrocytes and C6 glioblastoma cells (20,32).
Tumor apoptosis induced by cannabinoid receptor activation was
verified to be mediated by ceramide de novo synthesis
(31). Growing evidence suggests
that as a ubiquitous lipid messenger, ceramides play an important
role in the control of tumor cell fate (33). Yet there is limited research to
further identify the role of ceramides in CRC progression and
treatment. In the present study, we found that the levels of
ceramides C16:0, C24:0 and C24:1 were obviously elevated in the CRC
tissues. Increased levels of the C16 and C24 ceramides were also
observed in human head and neck squamous cell carcinoma (HNSCC)
(34). As we observed, the C16
ceramide is the most abundant ceramide in the colon, followed by
the C24 ceramide. Both two types of ceramides were reported to
induce apoptosis in hepatocellular cancer, prostate cancer and
colon cancer cells (35–37). In another report, the C16 ceramide
was also considered to play an anti-apoptotic role in HNSCC via
selective regulation of the ER-stress response pathway (38). In contrary, levels of C18 and C20
ceramides in the CRC tumors were significantly decreased compared
to these levels in the non-tumor tissues. A reduced level of C18
ceramide in tumors was also reported in HNSCC (34), which was thought to contend for
tumor growth via telomerase activity and mitochondrial
dysfunction.
We further measured ceramide synthases (CerSs) in
human CRC tissues, which are important enzymes for ceramide
synthesis. Since specificity of each CerS subtype for ceramides
with fatty acyl chain, four abundant CerS subtypes in human colon
were evaluated. Unexpectedly, the mRnA levels of all the detected
CerSs were sharply increased in the CRC tissues. Upregulated levels
of CerS2, CerS5 and CerS6 were explained to synthesize and elevate
enough C16 and C24 ceramides to decelerate tumorigenesis. CerS1
showed a contrary upward trend against the reduced levels of its
specific C18 ceramide, which may be a defensive mechanism to
compensate enough C18 ceramides. Overlapping enzyme activity of
CerS2 and CerS6, partly responsible for synthesis of C18 and C20
ceramides, made it difficult to study CerSs and ceramides in a
one-to-one relationship. The complex regulatory mechanism of CerSs
and ceramides with different carbon chains of fatty acyl still
require further in-depth research. Yet, consistent alteration of
C16/C18/C24 ceramides in several tumors revealed that they may be
potential indicators in tumor monitoring.
In addition to AA, a metabolite of AEA, the
expression of other free fatty acids was also detected by LC-MS/MS.
MUFAs with carbon chains from C14 to C24 and most PUFAs showed a
high unitive increase, while among the SFAs, only C18:0 and C24:0
FFAs were increased in the CRC tissues. Increased MUFAs provide
enough resource to build more phospholipids, diacylglycerols and
triacylglycerols, to serve as a continuous nutrient fuel for
rapidly proliferating cancer cells (39,40).
As we know, regular cell growth needs a stabilized balance between
SFAs and unsaturated fatty acids (UFAs) (41). The augmentation of MUFAs and PUFAs
demonstrated the rapid proliferation and aggressiveness of human
CRC.
In conclusion, the present study demonstrated an
increase in AEA in human CRC tissues along with lymph node
metastasis, which could be an endogenous bio-indicator for the
malignant degrees of CRC. Overexpression of its preferred receptor,
CB1, aided in further understanding the important role of the
endocannabinoid system in the treatment of human CRC. Moreover, the
variation in levels of long chain ceramides and UFAs revealed that
lipid dysfunction plays a vital role in CRC progression, and these
levels may also be utilized as important monitoring biomarkers and
novel drug targets for colorectal cancer.
Acknowledgments
The authors thank the Eye institute of xiamen
University for assistance and Mrs Jin Fu for help with the design
of this research. This study was supported by grants from the
National Natural Sciences Foundation of China (no. 81373273),
Fujian Provincial Natural Science Foundation (no. 2013J05122), and
xiamen Southern Ocean Research Center Project (no.
14GYY018NF18).
References
|
1
|
Veldkamp R, Gholghesaei M, Bonjer HJ,
Meijer DW, Buunen M, Jeekel J, Anderberg B, Cuesta MA, Cuschierl A,
Fingerhut A, et al: European Association of Endoscopic Surgery
(EAES): Laparoscopic resection of colon Cancer: Consensus of the
European Association of Endoscopic Surgery (EAES). Surg Endosc.
18:1163–1185. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tsujimoto H, Tsukioka S, Ono S, Sakamoto
E, Sakamoto K, Tsuta K, Nakagawa F, Saito H, Uchida J, Kiniwa M, et
al: Effect of leucovorin on the antitumor efficacy of the 5-FU
prodrug, tegafur-uracil, in human colorectal cancer xenografts with
various expression levels of thymidylate synthase. Oncol Lett.
1:973–980. 2010.PubMed/NCBI
|
|
3
|
Yang SY, Miah A, Sales KM, Fuller B,
Seifalian AM and Winslet M: Inhibition of the p38 MAPK pathway
sensitises human colon cancer cells to 5-fluorouracil treatment.
Int J Oncol. 38:1695–1702. 2011.PubMed/NCBI
|
|
4
|
Mechoulam R, Ben-Shabat S, Hanus L,
Ligumsky M, Kaminski NE, Schatz AR, Gopher A, Almog S, Martin BR,
Compton DR, et al: Identification of an endogenous
2-mono-glyceride, present in canine gut, that binds to cannabinoid
receptors. Biochem Pharmacol. 50:83–90. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Devane WA, Breuer A, Sheskin T, Järbe TU,
Eisen MS and Mechoulam R: A novel probe for the cannabinoid
receptor. J Med Chem. 35:2065–2069. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Di Carlo G and Izzo AA: Cannabinoids for
gastrointestinal diseases: Potential therapeutic applications.
Expert Opin Investig Drugs. 12:39–49. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
De Petrocellis L, Cascio MG and Di Marzo
V: The endocannabinoid system: A general view and latest additions.
Br J Pharmacol. 141:765–774. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Piomelli D: The molecular logic of
endocannabinoid signalling. Nat Rev Neurosci. 4:873–884. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cravatt BF, Giang DK, Mayfield SP, Boger
DL, Lerner RA and Gilula NB: Molecular characterization of an
enzyme that degrades neuromodulatory fatty-acid amides. Nature.
384:83–87. 1996. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pertwee RG: Cannabinoid Receptors.
Academic Press; New York, NY: 1995
|
|
11
|
Pertwee RG: Pharmacology of cannabinoid
CB1 and CB2 receptors. Pharmacol Ther. 74:129–180. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Di Marzo V, Melck D, De Petrocellis L and
Bisogno T: Cannabimimetic fatty acid derivatives in cancer and
inflammation. Prostaglandins Other Lipid Mediat. 61:43–61. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gonsiorek W, Lunn C, Fan X, Narula S,
Lundell D and Hipkin RW: Endocannabinoid 2-arachidonyl glycerol is
a full agonist through human type 2 cannabinoid receptor:
Antagonism by anandamide. Mol Pharmacol. 57:1045–1050.
2000.PubMed/NCBI
|
|
14
|
Guzmán M: Cannabinoids: Potential
anticancer agents. Nat Rev Cancer. 3:745–755. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ligresti A, Bisogno T, Matias I, De
Petrocellis L, Cascio MG, Cosenza V, D’argenio G, Scaglione G,
Bifulco M, Sorrentini I, et al: Possible endocannabinoid control of
colorectal cancer growth. Gastroenterology. 125:677–687. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sarnataro D, Grimaldi C, Pisanti S,
Gazzerro P, Laezza C, Zurzolo C and Bifulco M: Plasma membrane and
lysosomal localization of CB1 cannabinoid receptor are dependent on
lipid rafts and regulated by anandamide in human breast cancer
cells. FEBS Lett. 579:6343–6349. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
De Petrocellis L, Melck D, Palmisano A,
Bisogno T, Laezza C, Bifulco M and Di Marzo V: The endogenous
cannabinoid anandamide inhibits human breast cancer cell
proliferation. Proc Natl Acad Sci USA. 95:8375–8380. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Contassot E, Wilmotte R, Tenan M, Belkouch
MC, Schnüriger V, de Tribolet N, Burkhardt K and Dietrich PY:
Arachidonylethanolamide induces apoptosis of human glioma cells
through vanilloid receptor-1. J Neuropathol Exp Neurol. 63:956–963.
2004.PubMed/NCBI
|
|
19
|
Galve-Roperh I, Sánchez C, Cortés ML,
Gómez del Pulgar T, Izquierdo M and Guzmán M: Anti-tumoral action
of cannabinoids: Involvement of sustained ceramide accumulation and
extracellular signal-regulated kinase activation. Nat Med.
6:313–319. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Morad SA and Cabot MC:
Ceramide-orchestrated signalling in cancer cells. Nat Rev Cancer.
13:51–65. 2013. View
Article : Google Scholar
|
|
21
|
Grösch S, Schiffmann S and Geisslinger G:
Chain length-specific properties of ceramides. Prog Lipid Res.
51:50–62. 2012. View Article : Google Scholar
|
|
22
|
Selzner M, Bielawska A, Morse MA, Rüdiger
HA, Sindram D, Hannun YA and Clavien PA: Induction of apoptotic
cell death and prevention of tumor growth by ceramide analogues in
metastatic human colon cancer. Cancer Res. 61:1233–1240.
2001.PubMed/NCBI
|
|
23
|
Garzotto M, White-Jones M, Jiang Y,
Ehleiter D, Liao WC, Haimovitz-Friedman A, Fuks Z and Kolesnick R:
12-O-tetradecanoylphorbol-13-acetate-induced apoptosis in LNCaP
cells is mediated through ceramide synthase. Cancer Res.
58:2260–2264. 1998.PubMed/NCBI
|
|
24
|
Chmura SJ, Nodzenski E, Beckett MA, Kufe
DW, Quintans J and Weichselbaum RR: Loss of ceramide production
confers resistance to radiation-induced apoptosis. Cancer Res.
57:1270–1275. 1997.PubMed/NCBI
|
|
25
|
Verheij M, van Blitterswijk WJ and
Bartelink H: Radiation-induced apoptosis - the ceramide-SAPK
signaling pathway and clinical aspects. Acta Oncol. 37:575–581.
1998. View Article : Google Scholar
|
|
26
|
Chen L, Xie B, Li L, Jiang W, Zhang Y, Fu
J, Guan G and Qiu Y: Rapid and sensitive LC-MS/MS analysis of fatty
acids in clinical amples. Chromatographia. 77:1241–1247. 2014.
View Article : Google Scholar
|
|
27
|
Wu X, Han L, Zhang X, Li L, Jiang C, Qiu
Y, Huang R, Xie B, Lin Z, Ren J, et al: Alteration of
endocannabinoid system in human gliomas. J Neurochem. 120:842–849.
2012. View Article : Google Scholar
|
|
28
|
D’Argenio G, Valenti M, Scaglione G,
Cosenza V, Sorrentini I and Di Marzo V: Upregulation of anandamide
levels as an endogenous mechanism and a pharmacological strategy to
limit colon inflammation. FASEB J. 20:568–570. 2006.
|
|
29
|
Izzo AA, Aviello G, Petrosino S, Orlando
P, Marsicano G, Lutz B, Borrelli F, Capasso R, Nigam S, Capasso F,
et al: Endocannabinoid Research Group: Increased endocannabinoid
levels reduce the development of precancerous lesions in the mouse
colon. J Mol Med Berl. 86:89–98. 2008. View Article : Google Scholar
|
|
30
|
Melck D, De Petrocellis L, Orlando P,
Bisogno T, Laezza C, Bifulco M and Di Marzo V: Suppression of nerve
growth factor Trk receptors and prolactin receptors by
endocannabinoids leads to inhibition of human breast and prostate
cancer cell proliferation. Endocrinology. 141:118–126. 2000.
|
|
31
|
Cianchi F, Papucci L, Schiavone N, Lulli
M, Magnelli L, Vinci MC, Messerini L, Manera C, Ronconi E,
Romagnani P, et al: Cannabinoid receptor activation induces
apoptosis through tumor necrosis factor α-mediated ceramide de novo
synthesis in colon cancer cells. Clin Cancer Res. 14:7691–7700.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sánchez C, Galve-Roperh I, Rueda D and
Guzmán M: Involvement of sphingomyelin hydrolysis and the
mitogen-activated protein kinase cascade in the
Δ9-tetrahydrocannabinol-induced stimulation of glucose
metabolism in primary astrocytes. Mol Pharmacol. 54:834–843.
1998.
|
|
33
|
Ogretmen B and Hannun YA: Biologically
active sphingolipids in cancer pathogenesis and treatment. Nat Rev
Cancer. 4:604–616. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Koybasi S, Senkal CE, Sundararaj K,
Spassieva S, Bielawski J, Osta W, Day TA, Jiang JC, Jazwinski SM,
Hannun YA, et al: Defects in cell growth regulation by
C18:0-ceramide and longevity assurance gene 1 in human head and
neck squamous cell carcinomas. J Biol Chem. 279:44311–44319. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Seumois G, Fillet M, Gillet L, Faccinetto
C, Desmet C, François C, Dewals B, Oury C, Vanderplasschen A,
Lekeux P, et al: De novo C16- and C24-ceramide generation
contributes to spontaneous neutrophil apoptosis. J Leukoc Biol.
81:1477–1486. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
White-Gilbertson S, Mullen T, Senkal C, Lu
P, Ogretmen B, Obeid L and Voelkel-Johnson C: Ceramide synthase 6
modulates TRAIL sensitivity and nuclear translocation of active
caspase-3 in colon cancer cells. Oncogene. 28:1132–1141. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Osawa Y, Uchinami H, Bielawski J, Schwabe
RF, Hannun YA and Brenner DA: Roles for C16-ceramide and
sphingosine 1-phosphate in regulating hepatocyte apoptosis in
response to tumor necrosis factor-alpha. J Biol Chem.
280:27879–27887. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Senkal CE, Ponnusamy S, Bielawski J,
Hannun YA and Ogretmen B: Antiapoptotic roles of
ceramide-synthase-6-generated C16-ceramide via selective regulation
of the ATF6/CHOP arm of ER-stress-response pathways. FASEB J.
24:296–308. 2010. View Article : Google Scholar :
|
|
39
|
Cairns RA, Harris IS and Mak TW:
Regulation of cancer cell metabolism. Nat Rev Cancer. 11:85–95.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rysman E, Brusselmans K, Scheys K,
Timmermans L, Derua R, Munck S, Van Veldhoven PP, Waltregny D,
Daniëls VW, Machiels J, et al: De novo lipogenesis protects cancer
cells from free radicals and chemotherapeutics by promoting
membrane lipid saturation. Cancer Res. 70:8117–8126. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nashed M, Chisholm JW and Igal RA:
Stearoyl-CoA desaturase activity modulates the activation of
epidermal growth factor receptor in human lung cancer cells. Exp
Biol Med. 237:1007–1017. 2012. View Article : Google Scholar
|