Introduction
Osteochondromas are benign neoplasms that can be
subdivided into 2 groups. The more common tumors (also called
osteocartilaginous exostoses) consist of a cartilage-capped bony
projection arising on the external surface of bone and containing a
marrow cavity that is continuous with that of the underlying bone
(1). The second variant is the rare
extraskeletal (or soft tissue) osteochondroma. The latter is
defined as a single mass that is peripherally encased by mature
hyaline cartilage, has an osseous centre, shows an overall
organization similar to that of a conventional osteochondroma but
the lesion cannot be intra-articular or in any way be attached to
bone (2–4).
Many osteochondromas in bone are asymptomatic and
consequently are not detected and resected (1). Candidates for the cell of origin of
osteochondroma include growth-plate chondrocytes, perichondrial
cells, and cells of the Groove of Ranvier (5,6). The
latter is a fibrochondrosseous structure encircling the growth
plate and containing chondro- and osteoprogenitor cells. It was
long debated whether osteochon-droma was a developmental disorder
or a true neoplasm. The finding in a subset of osteochondromas of
cytogenetic aberrations, mainly 8q deletions, as well as biallelic
inactivation of the EXT1 (located in 8q24) or EXT2
(located in 11p11) gene in cells of the cartilage cap supports a
neoplastic nature of the tumors (1). Evidence has been provided that
EXT1 functions as a typical tumor suppressor gene, that is,
both copies are functionally inactivated in osteochondroma cells,
with both inactivating events being somatic in solitary,
non-hereditary osteochondromas (7,8). The
protein sequences EXT1 and EXT2 show structural similarities. They
accumulate in the Golgi apparatus where they catalyze the synthesis
of heparan sulphate (9,10), an essential component of cell
surface and matrix-associated proteoglycans (11). It interacts with numerous signaling
proteins and regulates their distribution and activity on target
cells. Many of these proteins are expressed in the growth plate of
developing skeletal elements, and several skeletal phenotypes are
caused by mutations affecting these proteins as well as in heparan
sulphate-synthesizing and modifying enzymes (11). Mutations of EXT1 and
EXT2 lead to heparan sulphate deficiency. The resulting
aberrant distribution of signaling factors as well as aberrant
responsiveness to them by target cells lead to exostosis formation
(11).
Extraskeletal osteochondroma is a slowly growing,
often painless tumor which is most commonly located in the hands,
feet, and knee joint (2,3,12–15).
Extraskeletal osteochondromas have also been found near the hip
(16), in the buttocks (17), and in the nape area (18). They may display cellular atypia but
they do not metastasize or undergo malignant transformation
(2–4). Although various mechanisms have been
suggested for the cause of extraskeletal osteochondroma (3,4), its
etiology and pathogenesis are unknown and there is not cytogenetic
or molecular genetic information about the disease.
In the present study, we describe the cytogenetic
rearrangement of chromosome band(s) 12q14~15 through a pericentric
inversion in two extraskeletal osteochondromas. The molecular
result of this was aberrant expression of HMGA2.
Materials and methods
Ethics statement
The study was approved by the Regional Ethics
Committee (Regional komité for medisinsk forskning-setikk Sør-Øst,
Norge, http://helseforskning.etikkom.no) and written informed
consent was obtained from the patients.
Patients
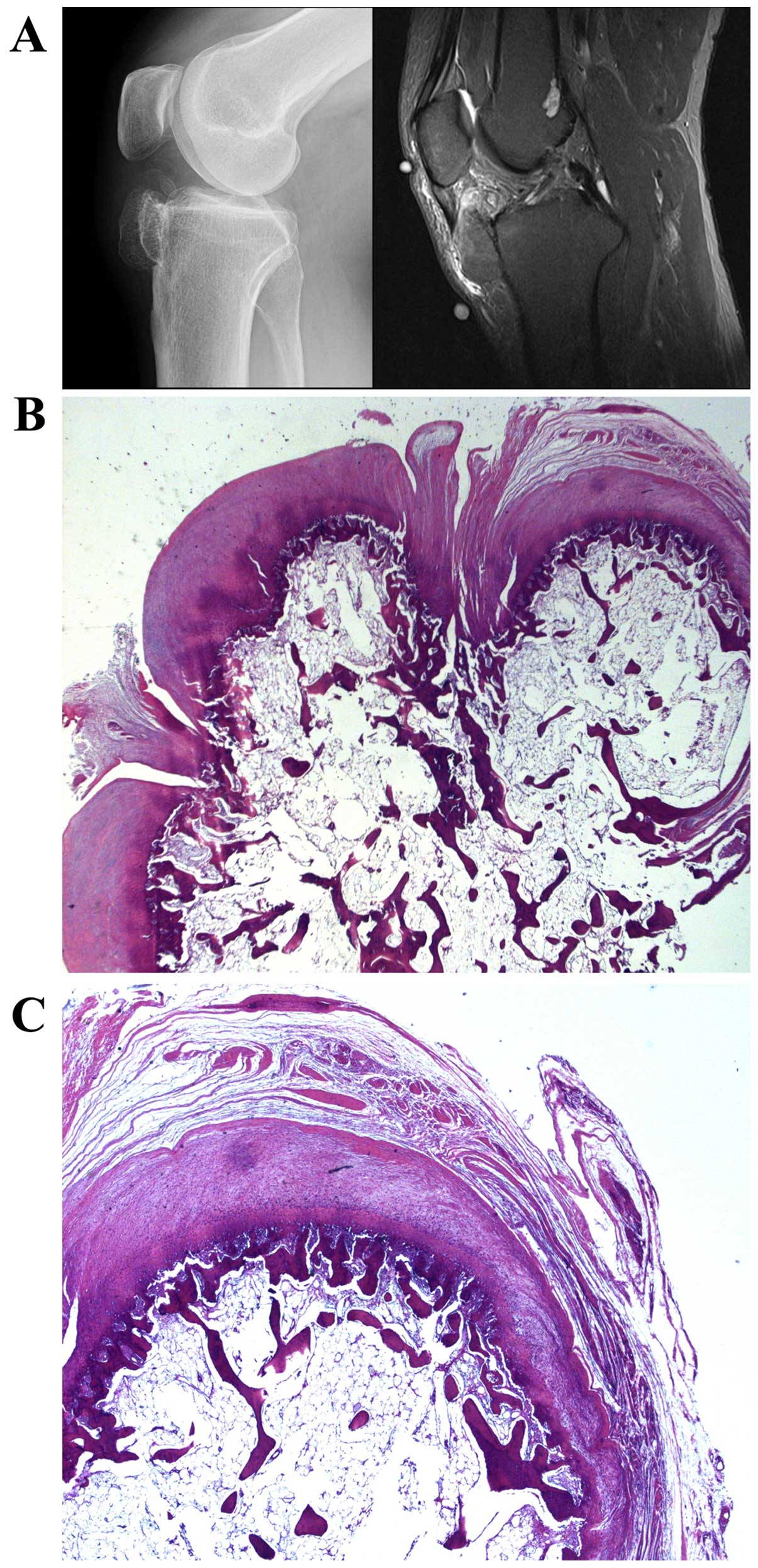
Case 1. A 43-year-old man had noticed a tumor in the
right knee for the last five years. For six months there had been
some pain. The plain radiograph showed a bony lesion adjacent to
the proximal tibial epiphysis and metaphysis. It was ovoid to
dumbbell-shaped, measured 4 cm, and consisted of trabecular bone
surrounded by a thin cortex. The MR signal was consistent with
fatty bone marrow covered by a thin cartilage cap. Thus, the lesion
resembled an osteochondroma, but the underlying anterior tibial
cortex was intact and there was no continuous marrow cavity with
the underlying bone (Fig. 1A).
Histological examination disclosed a bony lesion in which fatty
medullary bone in the middle was surrounded by thin cortical bone
covered by a cartilaginous cap. Around the cap there was some
fibrocartilage and connective tissue. No cellular atypia was
observed. The microscopic features were of an osteochondromatous
lesion with no connection to preexisting bone (Fig. 1B and C).
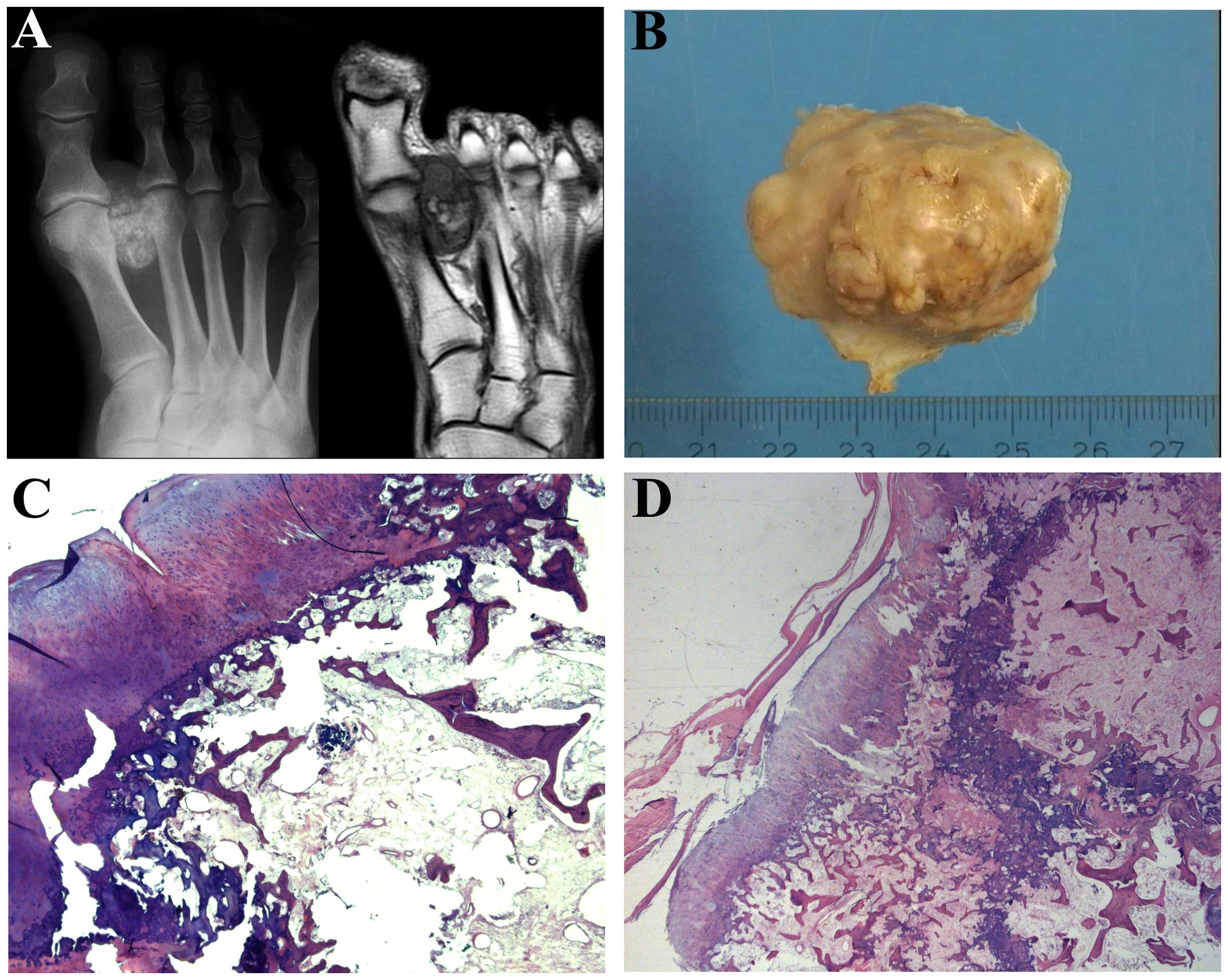
Case 2. A 45-year-old man had for at least five
years noticed a growing tumor in the foot. The radiograph revealed
a partly mineralized 4-cm lesion in the soft tissues between the
first and second metatarsal heads and extending distal to the
MCP-joints. There was no continuous cortex-like periphery, and the
calcifications were partly flocculent and comma-shaped as in
cartilage matrix, partly more trabecular resembling ossification.
On MR, the lesion was located adjacent to the surface of the first
metatarsal and the base of the proximal phalanx without involvement
of the bone marrow. There was fatty marrow in the central part,
while the periphery of the lesion gave signals as fibrous and
chondroid tissue (Fig. 2A). The
excised tumor consisted of fatty and fibrous bone marrow centrally
with small trabeculae and some more sclerotic bony tissue without
atypia (Fig. 2B). On the surface a
cartilage cap was noted, but there was no continuum of the
medullary with the underlying bone marrow. The microscopic features
were of an osteochondromatous lesion with no connection to the
pre-existing bone (Fig. 2C and
D).
Chromosome banding analysis and
fluorescence in situ hybridization (FISH)
Samples from the surgically removed tumors were
mechanically and enzymatically disaggregated and short-term
cultured as described elsewhere (19). The cultures were harvested and the
chromosomes G-banded using Wright stain. The subsequent cytogenetic
analysis and karyotype description followed the recommendations of
the ISCN (20).
FISH analysis was performed on metaphase plates. BAC
clones were retrieved from the Human genome high-resolution BAC
re-arrayed clone set (the ‘32K set’; BACPAC Resources, http://bacpac.chori.org/pHumanMinSet.htm). The ‘32K
set’ is mapped on the UCSC Genome Browser on Human Genome May 2004
(NCBI/hg17) assembly. Mapping data for the 32K human re-array are
available in an interactive web format (http://bacpac.chori.org/pHumanMinSet.htm, from the
Genomic Rearrays page) and are obtained by activation of the UCSC
browser track for the hg17 UCSC assembly from the ‘32K set’
homepage (http://bacpac.chori.org/genomicRear-rays.php). The BAC
clones were selected according to physical and genetic mapping data
on chromosome 12 as reported on the Human Genome Browser at the
University of California, Santa Cruz website (May 2004, http://genome.ucsc.edu/). In addition, FISH mapping of
the clones on normal controls was performed to confirm their
chromosomal location.
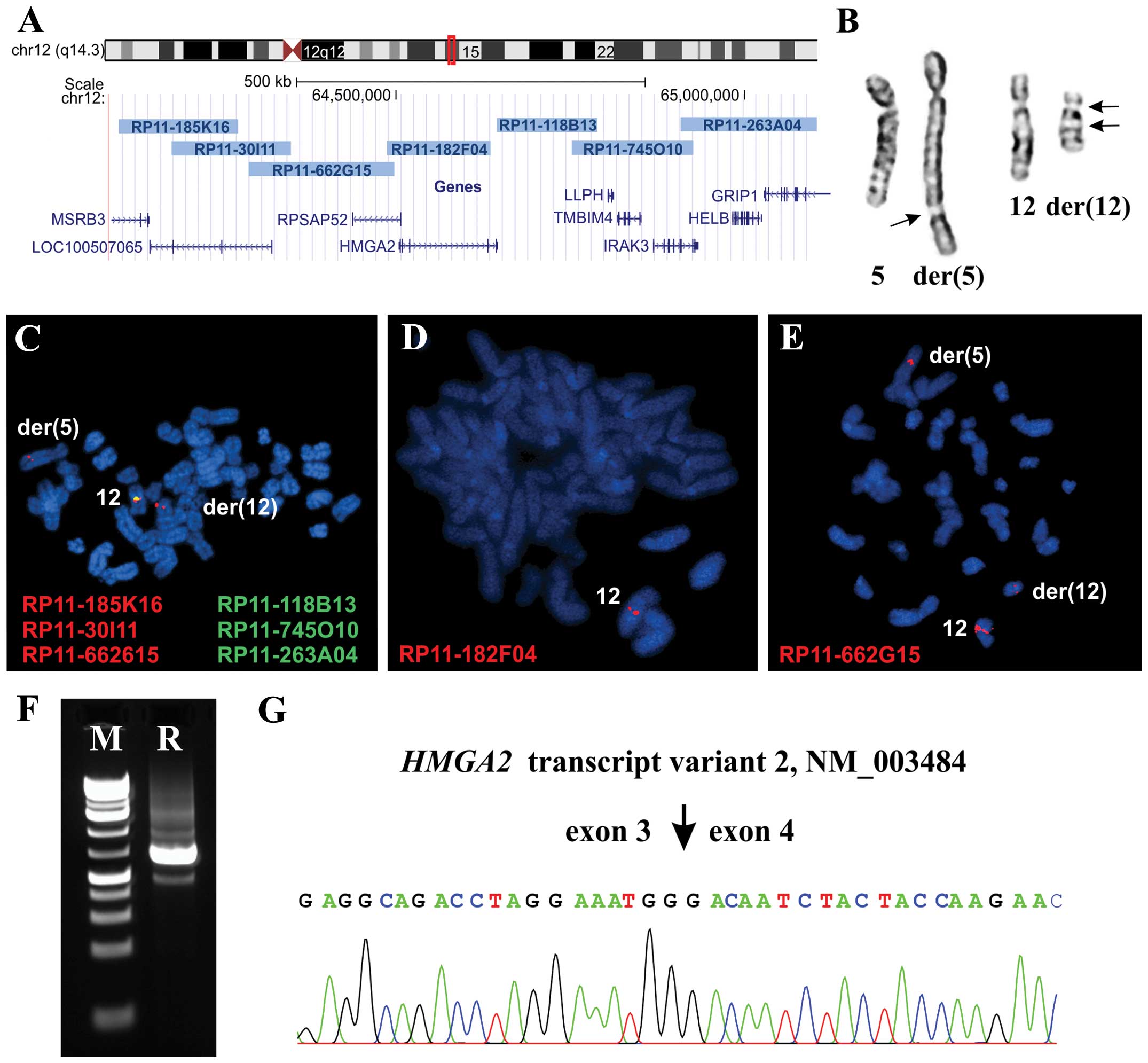
The clones used were RP11-185K16,
(chr12:64103524-642 74514), RP11-30I11 (chr12:64178505-64349708),
RP11-662 G15 (chr12:64288763-64498219), RP11-182F04 (chr12:644
86880-64635771), RP118B13 (chr12:64644 968-64789255), RP11-745O10
(chr12:64752327-64926193) and RP11-263A04
(chr12:64908453-65103538). All of them are mapped to chromosome
band 12q14.3 (Fig. 3A). DNA was
extracted and probes were labeled and hybridized according to
Abbott Molecular recommendations (http://www.abbottmolecular.com/home.html). Chromosome
preparations were counter-stained with 0.2 µg/ml DAPI and
overlaid with a 24×50 mm2 coverslip. Fluorescent signals
were captured and analyzed using the CytoVision system (Leica
Biosystems, Newcastle, UK).
Molecular genetic analysis
Total RNA was extracted using miRNeasy kit and
QIAcube according to the manufacturer’s recommendations (both from
Qiagen Nordic, Stockholm, Sweden). Human Universal Reference Total
RNA was used as control (Clontech Laboratories; Takara-Bio Group;
Europe/SAS, Saint-Germain-en-Laye, France). According to the
company’s information, it is a mixture of total RNAs from a
collection of adult human tissues chosen to represent a broad range
of expressed genes. Both male and female donors are represented.
Total RNA (400–500 ng) was reverse-transcribed in a 20-µl
reaction volume using iScript Advanced cDNA Synthesis kit for
RT-qPCR according to the manufacturer’s instructions (Bio-Rad
Laboratories, Oslo, Norway). The cDNA was diluted to 10 ng
equivalent of RNA/µl and 1 µl was used as template in
subsequent real-time PCR assays.
Real-time PCR was carried out to determine the
expression level of HMGA2. The TaqMan gene expression assays
(Applied Biosystems, Foster City, CA, USA), Hs00171569_m1 (HMGA2
exons 1–2), Hs00971725_m1 (HMGA2 exons 4–5), Hs00609162_m1 (EXT1
exon 8–9), and Hs00181158_m1 (EXT2 exon 8–9) were used. The
S100A10 gene, assay Hs00237010_ml, was used as endogenous
control since this gene is expressed in chondrocytes (21). The 20-µl reaction volume
contained 1× TaqMan Universal Master Mix II with UNG, lx of the 20X
TaqMan gene expression mix and 1 µl cDNA (10 ng equivalent
of RNA). Four replicates of each sample and the endogenous control
were performed. Real time PCR was run on a CFX96 Touch™ Real-Time
PCR Detection system (Bio-Rad). The thermal cycling included an
initial step at 50°C for 2 min, followed by 10 min at 95°C and 40
cycles of 15 sec at 95°C, and 1 min at 60°C. The data were analyzed
using the CFX Manager software (Bio-Rad).
For 3′-RACE, 100 ng of total RNA were
reverse-transcribed in a 20-µl reaction volume with the
A3RNV-RACE primer (5′-ATC GTT GAG ACT CGT ACC AGC AGA GTC ACG AGA
GAG ACT ACA CGG TAC TGG TTT TTT TTT TTT TTT-3′) using iScript
Select cDNA Synthesis kit according to the manufacturer’s
instructions (Bio-Rad). One microliter was used as template and
amplified using the outer primer combination HMGA2–846F1 (5′-CCA
CTT CAG CCC AGG GAC AAC CT-3′) and A3R-1New (5′-TCG TTG AGA CTC GTA
CCA GCA GAG TCA C-3′). One microliter of the amplified products was
used as template in nested PCR with the primers HMGA2–982F1 (5′-CAA
GAG TCC CTC TAA AGC AGC TCA-3′) and A3R-3 (5′-CGA GAG AGA CTA CAC
GGT ACT GGT-3′). For both PCRs the 25-µl reaction volume
contained 12.5 µl of Premix Taq (Takara-Bio) template, and
0.4 µM of each of the forward and reverse primers. PCR
cycling consisted of an initial step of denaturation at 94°C for 30
sec followed by 35 cycles of 7 sec at 98°C, 30 sec at 55°C, 90 sec
at 72°C, and a final extension for 5 min at 72°C.
Three microliters of the PCR products were stained
with GelRed (Biotium, Hayward, CA, USA), analyzed by
electrophoresis through 1.0% agarose gel, and photographed. The
rest of the amplified fragments were purified using the Thermo
Scientific GeneJET PCR purification kit (Fisher Scientific, Oslo,
Norway) and direct sequencing was performed using the Lightrun
sequencing service of GATC Biotech (http://www.gatc-biotech.com/en/sanger-services/lightrun-sequencing.html).
The BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi) and BLAT
(http://genome.ucsc.edu/cgi-bin/hgBlat) programs were
used for computer analysis of sequence data.
To verify the results obtained by 3′-RACE in case 2,
i.e., the presence of an HMGA2-SOX5 chimera transcript (see
below), PCRs were performed using the following primer
combinations: HMGA2-846F1/SOX5-Int1-R1 (5′-CAA CCA TAG CTG CAT CCC
GCT GT-3′), HMGA2-846F1/SOX5-634R1 (5′-AAG TTC CCC GAT CCC ATT GCA
AG-3′) and HMGA2-846F1/SOX5-481R1 (CGT TCA GGA GTT CCC AGG GCT GT).
The primers SOX5-634R1 and SOX5-481R1 correspond to nucleotides
634–656 (exon 4) and 481–503 (exon 3) in the SOX5 mRNA
sequence with accession NM_006940 version 4. The 25-µl PCR
volumes contained 12.5 µl of Premix Taq (Takara-Bio), 2
µl of diluted cDNA, and 0.2 µM of each of the forward
and reverse primers. The PCRs were run on a C-1000 Thermal cycler
(Bio-Rad). The PCR conditions were: an initial denaturation at 94°C
for 30 sec followed by 35 cycles of 7 sec at 98°C, 120 sec at 68°C,
and a final extension for 5 min at 68°C.
Immunohistochemistry
To detect the HMGA2 protein, immu-nostaining was
performed as previously described (22).
Results
Chromosome banding analysis and FISH
In case 1, the G-banding analysis yielded the
karyotype 46,XY,der(5)t(5;12)
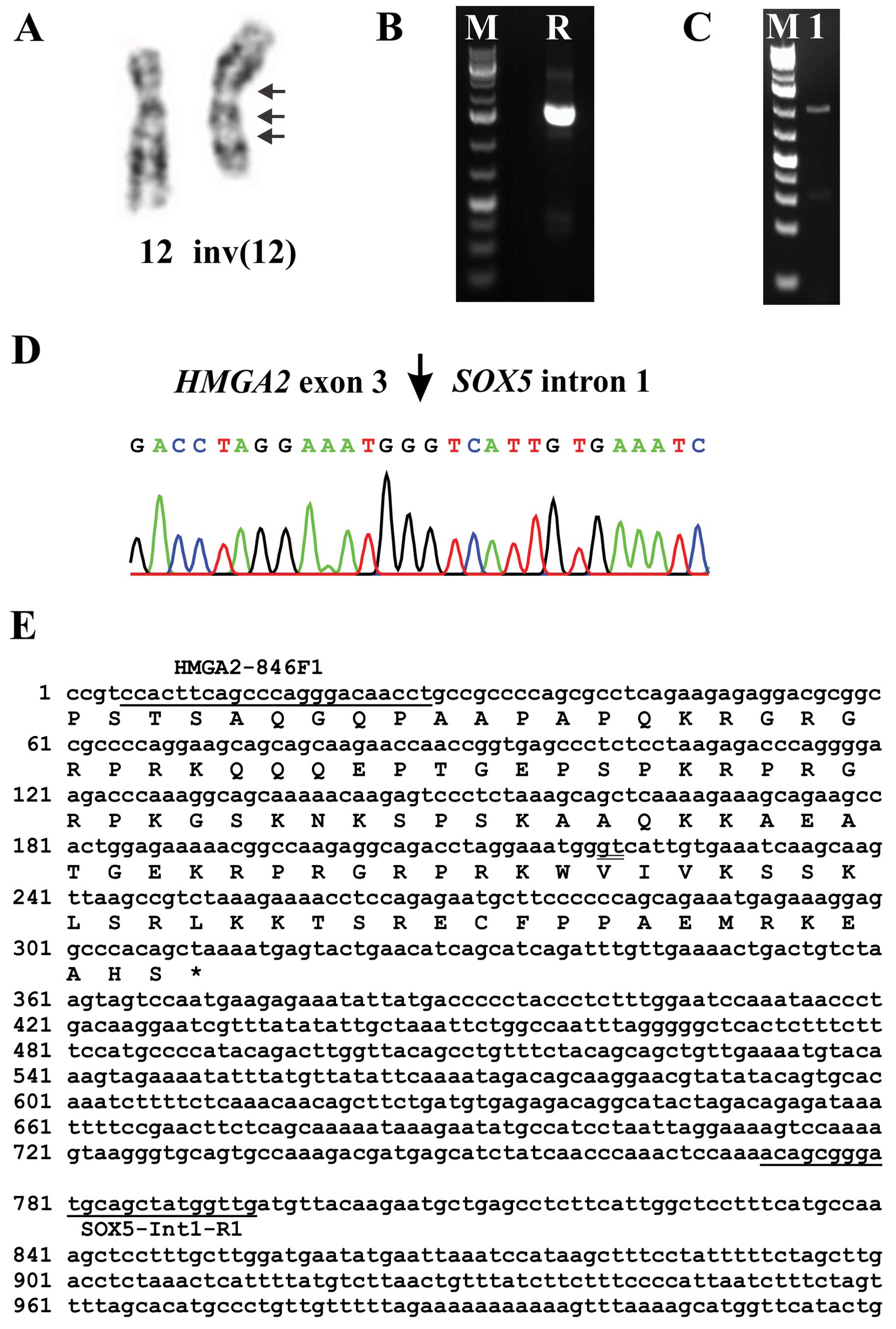
(q35;q14~15),der(12)t(5;12)inv(12)(p11q14~15)[8]/46,XY[3] (Fig. 3B). In case 2, the analysis yielded
the karyotype 46,XY,inv(12)(qter->q14~15::p11->q13::q14~15->q13::p11->pter)
[13]/46,XY,idem,t(5;13)(q13;p11)[2] (Fig. 4A).
The FISH experiments in case 1 showed that there was
only one copy of the HMGA2 gene in the metaphase cells with
the aberrant karyotype which was located on the normal chromosome
12 (Fig. 3C and D) indicating
heterozygous deletion of HMGA2. On the derivative
chromosomes, the probe which was a pool of the BACs RP11-118B13,
RP11-745O10 and RP11-263A04 (Fig.
3C) as well as the probe RP11-182F04 which covers the
HMGA2 gene (Fig. 3D) were
deleted. The probe containing the three BACs RP11-185K16,
RP11-30I11 and RP11-662G15 was split with one signal on
der(12) and the other on
der(5) (Fig. 3C). Further experiments showed that
the split signal was located on the BAC RP11-662G15 which is
upstream and outside the HMGA2 locus (Fig. 3E). Interphase FISH confirmed the
heterozygous deletion of HMGA2. In 98 nuclei, 76 had one
copy presumably corresponding to cells with an aberrant karyotype,
whereas 22 had 2 copies of HMGA2 and presumably representing
cells with a normal karyotype.
Molecular genetic analysis
3′-RACE in case 1 amplified a single fragment
(Fig. 3F) which by Sanger
sequencing was found to be the alternative transcript variant 2 of
HMGA2 with accession number NM_003484 (Fig. 3G).
3′-RACE in case 2 amplified a single fragment
(Fig. 4B). Sanger sequencing showed
that it was a chimeric cDNA fragment in which exon 3 of
HMGA2 was fused to a sequence in intron 1 of the SOX5
gene located in 12p12 (Fig. 4D and
E). PCR with the primers HMGA2-846F1/SOX5-Int1-R1 amplified a
cDNA fragment (Fig. 4C), direct
sequencing of which showed the same fusion point with the 3′-RACE
amplified fragment (Fig. 4D and E).
PCR with the forward HMGA2-846F1 primer and the reverse primers
SOX5-634R1 (located in exon 4) and SOX5-481R (located in exon 3 of
SOX5) did not amplify any other HMGA2-SOX5 fusion
transcripts.
By real-time PCR, the mean quantification cycle of
S100A10 (Cq Mean) was found to be 26.50, 22.23 and 26.75 for
case 1, case 2, and the human reference control sample,
respectively (Table I). The Cq Mean
for HMGA2 exons 1–2 was 34.24, 28.15 and 31.99, for case 1,
case 2 and the human reference control sample, respectively.
Expression of exons 4–5 of HMGA2 was noted in case 1 and in
the reference sample but not in case 2. The Cq Mean was 33.83 and
32.90 for case 2 and the reference sample, respectively. Thus, for
case 2, the data indicated the presence of an HMGA2
transcript in which exons 1 and 2 were present whereas exons 4 and
5 were lost. The expression of the EXT1 and EXT2
genes in case 1 was comparable to what was found in the human
reference control sample whereas their expression was very low in
case 2 (Table I).
 | Table IThe Cq values of expression of
HMGA2, EXT1, EXT2 and S100A10 in the
examined extraskeletal osteochondromas and human reference RNA. |
Table I
The Cq values of expression of
HMGA2, EXT1, EXT2 and S100A10 in the
examined extraskeletal osteochondromas and human reference RNA.
| Gene (assay) | Case 1 | Case 2 | Human
reference |
|---|
| S100A10
(Hs00237010_ml) | 26.50 | 22.23 | 26.75 |
| EXT1
(Hs00609162_m1) | 28.47 | 37.93 | 29.07 |
| EXT2
(Hs00181158_m1) | 28.78 | 32.39 | 27.79 |
| HMGA2 Exons
1–2 (Hs00171569_m1) | 34.24 | 28.15 | 31.99 |
| HMGA2 Exons
4–5 (Hs00971725_m1) | 33.83 | – | 32.90 |
Immunohistochemistry
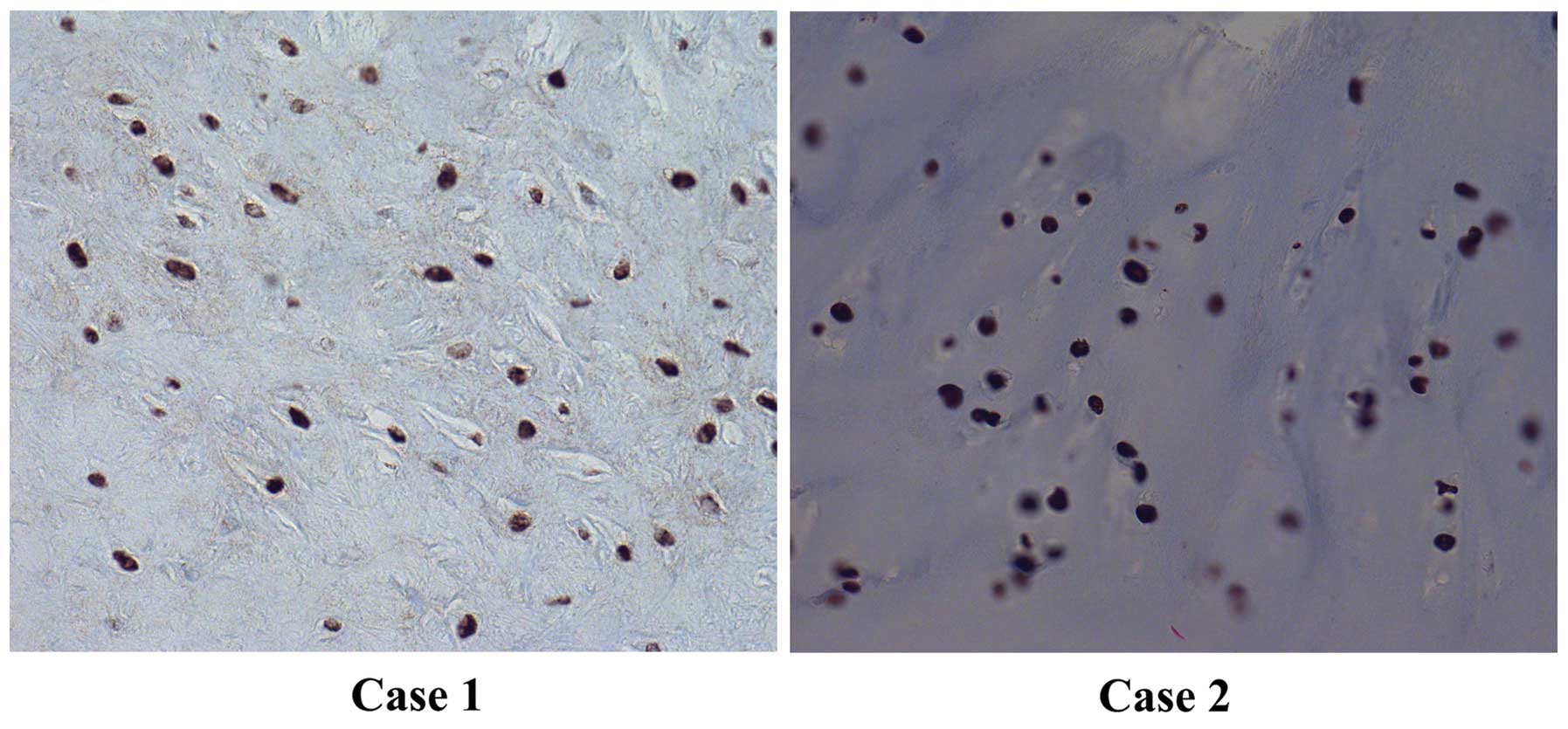
Strong and widespread immunohis-tochemical nuclear
staining for HMGA2 was noted in both tumors (Fig. 5).
Discussion
Cytogenetic information on both osteochondromas in
bone and extra skeletal osteochondromas is very limited. According
to the Mitelman Database of Chromosome Aberrations and Gene Fusions
in Cancer (http://cgap.nci.nih.gov/Chromosomes/Mitelman, Database
last updated on August 18, 2014), only 26 osteochondromas in bone
with clonal karyotypic aberrations have been published in
altogether 8 articles (23–30). Involvement of chromosome 8, mostly
deletions of 8q as cytogenetic evidence of EXT1 loss, was
observed in 18 of them. Four other tumors showed rearrangement of
11p as cytogenetic evidence of EXT2 involvement.
Additionally, breakpoints in 1p13~22 were noted in 5
osteochondromas (29) but no gene
has been associated with this cytogenetic change. No other
consistent pattern of aberrations has emerged.
Here, we present two cytogenetically analyzed
extraskeletal osteochondromas in which chromosome bands 12q14~15
was rearranged; both had inv(12)
but no microscopically detectable rearrangement of the long arm of
chromosome 8 let alone of band 8q24. Structural rearrangements
involving 12q were previously reported in 7 osteochondromas
(24). Aberration of 12q12~13 was
found to be clonal in two tumors whereas in another two it was
noted in a single metaphase only. Two other cases had abnormalities
mapped to 12q24 and a third had rearrangement of 12q11 (24). It is not possible for us to know
whether the breakpoint in some of these tumors might have been
reassigned to 12q14~15 if reviewed again.
Since cytogenetic change of bands 12q13~15 in benign
connective tissue tumors is almost always associated with
rearrangement and/or activation of HMGA2 (31), we decided to investigate whether
this gene is involved also in our two cases. The experiments by
3′-RACE, RT-PCR, and immunohictochemistry showed that HMGA2
was transcribed and translated into nuclear protein in both tumors.
In case 1, the data indicated that both transcript 1 (NM_003483,
real-time PCR experiments) and transcript 2 (NM_003484, 3′-RACE
experiments) were expressed. HMGA2 transcript 2 (assigned
with accession numbers AF533652, AY601867, and U29112) has been
found expressed in embryonic cells, cultured fibroblasts, as well
as leiomyomas; evidently, expression is not restricted to
neoplastic contexts (32,33).
The FISH experiments showed that, in the cells with
aberrant karyotype, there was only one copy of the HMGA2
gene which was located on the normal chromosome 12 and not on
der(5) or der(12). In addition, there were cells
carrying both copies of HMGA2 and had normal 46,XY
karyotype. Thus, the observed expression of HMGA2 could be
the result of an active HMGA2 allele on the normal 12 in
cells with abnormal karyo-type and/or active HMGA2 in cells
with a normal karyotype.
In case 2, only a chimeric HMGA2-SOX5 was
expressed in which exons 1–3 of HMGA2 were fused to an
intronic sequence of SOX5 from 12p12 (Fig. 4D and E). The ensuing
HMGA2-SOX5 fusion transcript codes for a putative protein
which contains amino acid residues 1–83 of HMGA2 protein (accession
number NP_003474.1) corresponding to exons 1–3 of the gene, and 30
amino acid residues from the intronic sequence of SOX5
(VIVKSSKLSRLKKTSRECFPPAEMRKEAHS). This pattern is similar to the
rearrangements of HMGA2 found in other connective tissue
tumor types, i.e., disruption of the HMGA2 locus leaves
intact exons 1–3 which encode the AT-hook domains and separates
them from the 3′-terminal part of the gene (34).
Recombinant HMGA2 protein was shown to significantly
increase the proliferative activity of chondrocytes in a
dose-dependent manner in an in vitro system utilizing cells
of porcine origin (35).
Application of a synthetic peptide comprising the functional
AT-hook motifs of the HMGA2 protein onto porcine hyaline cartilage
chondrocytes, grown in a monolayer cell culture, showed a
growth-promoting effect similar to the wild-type HMGA2 protein
(36). Moreover, HMGA2 can
influence the expression of genes involved in chondrogenesis such
as COL11A2 (37).
Overexpression of HMGA2-LPP fusion transcripts promotes
chondrogenesis by upregulating cartilage-specific collagen gene
expression through the N-terminal DNA binding domains. In the same
study, full-length HMGA2 was also shown to activate the
COL11A2 promoter when overexpressed indicating that
COL11A2 is a target gene of HMGA2 (37).
It should be emphasized that the role of
HMGA2 in chondromatous tumors is still studied only very
rudimentary in-as-much as only six soft tissue chondromas, two
skeletal chondromas, and three periosteal chondromas have been
subjected to this type of analysis (38,39).
HMGA2 expression was found in four soft tissue chondromas of
which three expressed a truncated transcript of HMGA2 and
one the full length transcript 1. Expression of HMGA2 was
found in both examined skeletal chondromas: a tumor with a
pericentric inv(12)(p12q13)
expressed a truncated transcript of HMGA2 whereas a tumor
carrying a t(2;11)(q37;q13) without visible involvement of 12q
expressed the full length HMGA2 transcript (38). On the other hand, neither
conventional RT-PCR nor real-time PCR showed expression of
HMGA2 in the examined periosteal chondromas, although two of
them had structural aberrations of chromosome bands 12q13~15
(39).
Our data on EXT1 and EXT2 from case 2
are in agreement with previously reported expression data from
osteochondromas in that they showed low expression levels (7,8). For
case 1, however, expression of EXT1 and EXT2 was at
the same low level as that found in the human reference control
sample (Table I). In bone
osteochondromas, homozygous deletion of the EXT1 gene is
only seen in chondrocytes of the cartilaginous cap, not in cells of
the perichondrium or from the bony stalk (8). A possible explanation of our results
for case 1 could be that the material used for expression analysis
of EXT1 and EXT2 contained more cells of the
perichondrium and bony stalk and less cells of the cartilaginous
cap.
In conclusion, our study showed that rearrangement
of chromosome bands 12q14~15 is recurrent in extraskeletal
osteochondromas. The cytogenetic change leads to expression of
HMGA2 or formation of HMGA2 chimeras, i.e., the same
pathogenetic motifs that are well known also from other benign
connective tissue tumors.
Acknowledgments
The authors thank Hege Kilen Andersen for technical
help. This work was supported by grants from the Norwegian Cancer
Society and the Norwegian Radium Hospital Foundation.
References
|
1
|
Fletcher CDM, Bridge JA, Hogendoorn PCW
and Mertens F: WHO Classification of Tumours of Soft Tissue and
Bone. 5. 4th edition. IARC; Lyon: 2013
|
|
2
|
Estil JCJ Jr, Yeo ED, Kim HJ, Cho WT and
Lee JJ: A large extraskeletal osteochondroma of the foot. J Foot
Ankle Surg. 52:663–665. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kho VK and Chen WC: Extraskeletal
osteochondroma of the foot. J Chin Med Assoc. 73:52–55. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sit YK and Lui TH: Extraskeletal
osteochondroma of the medial arch of the foot. Foot Ankle Spec.
5:397–400. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bovée JV: EXTra hit for mouse
osteochondroma. Proc Natl Acad Sci USA. 107:1813–1814. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jones KB, Piombo V, Searby C, Kurriger G,
Yang B, Grabellus F, Roughley PJ, Morcuende JA, Buckwalter JA,
Capecchi MR, et al: A mouse model of osteochondromagenesis from
clonal inactivation of Ext1 in chondrocytes. Proc Natl Acad Sci
USA. 107:2054–2059. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hameetman L, David G, Yavas A, White SJ,
Taminiau AH, Cleton-Jansen AM, Hogendoorn PC and Bovée JV:
Decreased EXT expression and intracellular accumulation of heparan
sulphate proteoglycan in osteochondromas and peripheral
chon-drosarcomas. J Pathol. 211:399–409. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hameetman L, Szuhai K, Yavas A,
Knijnenburg J, van Duin M, van Dekken H, Taminiau AH, Cleton-Jansen
AM, Bovée JV and Hogendoorn PC: The role of EXT1 in nonhereditary
osteochondroma: identification of homozygous deletions. J Natl
Cancer Inst. 99:396–406. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Busse-Wicher M, Wicher KB and
Kusche-Gullberg M: The exostosin family: proteins with many
functions. Matrix Biol. 35:25–33. 2014. View Article : Google Scholar
|
|
10
|
McCormick C, Duncan G, Goutsos KT and
Tufaro F: The putative tumor suppressors EXT1 and EXT2 form a
stable complex that accumulates in the Golgi apparatus and
catalyzes the synthesis of heparan sulfate. Proc Natl Acad Sci USA.
97:668–673. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huegel J, Sgariglia F, Enomoto-Iwamoto M,
Koyama E, Dormans JP and Pacifici M: Heparan sulfate in skeletal
development, growth, and pathology: the case of hereditary multiple
exostoses. Dev Dyn. 242:1021–1032. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Maheshwari AV, Jain AK and Dhammi IK:
Extraskeletal para-articular osteochondroma of the knee - a case
report and tumor overview. Knee. 13:411–414. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Maheshwari AV, Muro-Cacho CA and Pitcher
JD Jr: Extraskeletal para-articular osteochondroma of the posterior
knee. J Knee Surg. 22:30–33. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nogita T and Kawakami M: Extraskeletal
osteochondroma in the finger. Mimicking the fourth phalangeal bone.
Acta Derm Venereol. 72:287–288. 1992.PubMed/NCBI
|
|
15
|
Sheff JS and Wang S: Extraskeletal
osteochondroma of the foot. J Foot Ankle Surg. 44:57–59. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu ZJ, Zhao Q and Zhang LJ: Extraskeletal
osteochondroma near the hip: a pediatric case. J Pediatr Orthop B.
19:524–528. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lim SC, Kim YS, Kim YS and Moon YR:
Extraskeletal osteo-chondroma of the buttock. J Korean Med Sci.
18:127–130. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Singh R, Sharma AK, Magu NK, Kaur KP, Sen
R and Magu S: Extraskeletal osteochondroma in the nape of the neck:
a case report. J Orthop Surg (Hong Kong). 14:192–195. 2006.
|
|
19
|
Mandahl N: Methods in solid tumour
cytogenetics. Human Cytogenetics: Malignancy and Acquired
Abnormalities. Rooney DE: Oxford University Press; New York NY: pp.
165–203. 2001
|
|
20
|
Schaffer LG, Slovak ML and Campbell LJ:
ISCN 2009: An International System for Human Cytogenetic
Nomenclature. Karger, Basel: 2009
|
|
21
|
Song C, Zhou X, Dong Q, Fan R, Wu G, Ji B,
Meng Q and Zheng M: Regulation of inflammatory response in human
chondrocytes by lentiviral mediated RNA interference against
S100A10. Inflamm Res. 61:1219–1227. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gorunova L, Bjerkehagen B and Heim S:
Paratesticular leiomyoma with a der(14)t(12;14)(q15;q24). Cancer
Genet. 204:465–468. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bridge JA, Bhatia PS, Anderson JR and Neff
JR: Biologic and clinical significance of cytogenetic and molecular
cytogenetic abnormalities in benign and malignant cartilaginous
lesions. Cancer Genet Cytogenet. 69:79–90. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bridge JA, Nelson M, Orndal C, Bhatia P
and Neff JR: Clonal karyotypic abnormalities of the hereditary
multiple exostoses chromosomal loci 8q24.1 (EXT1) and 11p11–12
(EXT2) in patients with sporadic and hereditary osteochondromas.
Cancer. 82:1657–1663. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Buddingh EP, Naumann S, Nelson M, Neffa
JR, Birch N and Bridge JA: Cytogenetic findings in benign
cartilaginous neoplasms. Cancer Genet Cytogenet. 141:164–168. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Feely MG, Boehm AK, Bridge RS, Krallman
PA, Neff JR, Nelson M and Bridge JA: Cytogenetic and molecular
cytogenetic evidence of recurrent 8q24.1 loss in osteochondroma.
Cancer Genet Cytogenet. 137:102–107. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mertens F, Rydholm A, Kreicbergs A, Willén
H, Jonsson K, Heim S, Mitelman F and Mandahl N: Loss of chromosome
band 8q24 in sporadic osteocartilaginous exostoses. Genes
Chromosomes Cancer. 9:8–12. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sawyer JR, Swanson CM, Lukacs JL, Nicholas
RW, North PE and Thomas JR: Evidence of an association between
6q13–21 chromosome aberrations and locally aggressive behavior in
patients with cartilage tumors. Cancer. 82:474–483. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sawyer JR, Thomas EL, Lukacs JL, Swanson
CM, Ding Y, Parham DM, Thomas JR and Nicholas RW: Recurring
breakpoints of 1p13 approximately p22 in osteochondroma. Cancer
Genet Cytogenet. 138:102–106. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tallini G, Dorfman H, Brys P, Dal Cin P,
De Wever I, Fletcher CD, Jonson K, Mandahl N, Mertens F, Mitelman
F, et al: Correlation between clinicopathological features and
karyotype in 100 cartilaginous and chordoid tumours. A report from
the chromosomes and morphology (CHAMP) collaborative study Group. J
Pathol. 196:194–203. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Heim S and Mitelman F: Cancer
Cytogenetics. 3rd edition. Wiley-Blackwell; New York, NY: 2009
|
|
32
|
Hauke S, Leopold S, Schlueter C, Flohr AM,
Murua Escobar H, Rogalla P and Bullerdiek J: Extensive expression
studies revealed a complex alternative splicing pattern of the
HMGA2 gene. Biochim Biophys Acta. 1729:24–31. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Quade BJ, Weremowicz S, Neskey DM, Vanni
R, Ladd C, Dal Cin P and Morton CC: Fusion transcripts involving
HMGA2 are not a common molecular mechanism in uterine leiomyomata
with rearrangements in 12q15. Cancer Res. 63:1351–1358.
2003.PubMed/NCBI
|
|
34
|
Cleynen I and Van de Ven WJ: The HMGA
proteins: A myriad of functions (Review). Int J Oncol. 32:289–305.
2008.PubMed/NCBI
|
|
35
|
Richter A, Hauschild G, Murua Escobar H,
Nolte I and Bullerdiek J: Application of high-mobility-group-A
proteins increases the proliferative activity of chondrocytes in
vitro. Tissue Eng Part A. 15:473–477. 2009. View Article : Google Scholar
|
|
36
|
Richter A, Lübbing M, Frank HG, Nolte I,
Bullerdiek JC and von Ahsen I: High-mobility group protein
HMGA2-derived fragments stimulate the proliferation of chondrocytes
and adipose tissue-derived stem cells. Eur Cell Mater. 21:355–363.
2011.PubMed/NCBI
|
|
37
|
Kubo T, Matsui Y, Goto T, Yukata K and
Yasui N: Overexpression of HMGA2-LPP fusion transcripts promotes
expression of the alpha 2 type XI collagen gene. Biochem Biophys
Res Commun. 340:476–481. 2006. View Article : Google Scholar
|
|
38
|
Dahlén A, Mertens F, Rydholm A, Brosjö O,
Wejde J, Mandahl N and Panagopoulos I: Fusion, disruption, and
expression of HMGA2 in bone and soft tissue chondromas. Mod Pathol.
16:1132–1140. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Panagopoulos I, Gorunova L, Taksdal I,
Bjerkehagen B and Heim S: Recurrent 12q13–15 chromosomal
aberrations, high frequency of isocitrate dehydrogenase 1
mutations, and absence of high mobility group AT-hook 2 expression
in periosteal chondromas. Oncol Lett. 10:163–167. 2015.
|