Introduction
Pancreatic cancer, one of the most lethal types of
cancer, lacks effective therapeutic methods and exhibits a low
5-year survival rate of <5% (1).
Although surgical resection is the most important and curative
treatment methods for cancer, pancreatic cancer is usually
unresectable at the time of diagnosis (2,3).
Therefore, radiotherapy alone or as an adjuvant treatment has been
used as major therapeutic methods for pancreatic cancer (4). However, the radioresistance of
pancreatic cancer severely affects the efficacy and outcomes of
radiotherapy in clinical treatment (5,6).
Accordingly, overcoming radioresistance has become crucial in
cancer radiotherapy in recent years (7).
Autophagy, a catabolic process for the degradation
of cytoplasmic proteins and organelles as an adaptive response to
cell stress such as nutrient starvation or metabolic stress, is
considered a potential mechanism for the radioresistance of cancer
cells (8). Autophagy is a membrane
trafficking process in which the autophagosome formation is
triggered by class III phosphoinositide 3-kinase and beclin-1 (also
known as the mammalian homologue of the yeast autophagy-related
gene 6) (9). Beclin-1 is a critical
gene for autophagosome formation that shows high expression levels
during autophagy (10). Conversion
of the microtubule-associated protein light chain 3 (LC3) from
LC3-I (cytosolic form) to LC3-II (autophagic membrane form) is
another critical process during autophagy and the levels of LC3I/II
have been considered a classic marker for the detection of
autophagy (11). Mounting evidence
has demonstrated that inhibition of autophagy by 3-methyladenine
and chloroquine effectively enhanced the radiosensitivity of cancer
cells (12–14). Therefore, targeting autophagy to
inhibit radioresistance of pancreatic cancer cells is a promising
research direction for improving clinical outcomes.
MicroRNAs (miRNAs), the post transcriptional
regulators of gene expression, have been identified as an important
regulator in a variety of cell processes (15). They are 18–24 nucleotides in length
and bind to the 3′-untranslated region (UTR) of the target mRNA
leading to mRNA destabilization and thereby protein translational
inhibition (16,17). The role of miRNAs in tumorigenesis
and cancer treatment has been well characterized. miRNAs are
associated with patient survival and are useful predictors and
modificators for anticancer treatment (18–20).
However, the potential underlying mechanisms of miRNAs in
regulating radioresistance of pancreatic cancer cells remains
largely unknown.
Previous findings have indicated that miR-216a was
markedly decreased in pancreatic cancer (21–23),
suggesting an important role of miR-216a in pancreatic cancer. In
the present study, using bioinformatic algorithms we found that
beclin-1, an important regulator of autophagy, was a putative
target gene of miR-216a. Consequently, we investigated whether
miR-216a targeted beclin-1-mediated autophagy and played a critical
role in the radioresistance of pancreatic cancer cells. In the
present study, we found that miR-216a was inhibited in
radioresistant pancreatic cancer cells and that beclin-1 as well as
autophagy activity were highly upregulated. We also found that
forced expression of miR-216a significantly suppressed beclin-1 and
autophagy activity in radioresistant pancreatic cancer cells, which
enhanced the radiosensitivity of pancreatic cancer cells. Thus,
miR-216a is a promising target that can be used to sensitize
pancreatic cancer cells to irradiation by abrogating
irradiation-induced autophagy.
Materials and methods
Cell culture and mice
The human pancreatic cancer cell line, PANC-1, was
obtained from the Chinese Academy of Sciences (Shanghai, China) and
cultured in Dulbecco's modified Eagle's medium (DMEM;
Invitrogen-Life Technologies, Carlsbad, CA, USA) supplemented with
10% fetal bovine serum (Invitrogen-Life Technologies) containing
penicillin/streptomycin. The cells were grown in a humidified 5%
CO2 at 37°C in an incubator. The irradiation of PANC-1
cells was performed according to a previously reported method
(24). PANC-1 cells grown in
complete medium were subjected to 2 Gy 60Co radiation at
2 Gy/min using an X-ray machine (X-RAD 320, Precision X-ray) at the
Institute of Radiation Medicine of Affiliated Cancer Hospital of
Guangzhou Medical University (Guangdong, China). The irradiated
cells were then sub-cultured in new plates and irradiated with
increasing doses of irradiation (4, 6, 8, and 10 Gy) for subsequent
experiment.
Female 6-week-old BALB/c nude mice (25–30 g) were
obtained from the Medical Experimental Animal Center (Guangdong,
China) and housed under pathogen-free conditions with free access
to water and food. The animal experimental procedures were approved
and reviewed by the Institutional Animal Care and Use Committee of
Guangzhou Medical University.
Cell viability and colony formation
assays
Following irradiation or miR-216a treatment, cell
growth and viability was evaluated using a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT;
Sangon, Shanghai, China) assay. Briefly, the cells were seeded in
96-well plates at a density of 5×103 cells/200
µl. After treatment, fresh medium containing MTT [5 mg/ml
diluted in phosphate-buffered saline (PBS), 20 µl/well] was
added and incubated for an additional 4 h. The formed formazan was
resuspended with dimethyl sulfoxide (200 µl/well).
Absorbance was determined at 490 nm using an ELISA reader (Bio-Tek,
Winooski, VT, USA). For detection of the colony formation ability,
the cells following treatment were grown in 6-well plates and
cultured for 15 days. The old medium was discarded and cell
colonies stained with crystal violet were counted using a
dissecting microscope. The experiments were performed in
quadruplicate and repeated three times.
TUNEL assay
Apoptotic cells were stained using the TUNEL
(terminal deoxynucleotidyl transferase dUTP nick end-labeling)
apoptosis kit (Genmed Scientifics, Arlington, MA, USA) according to
the supplier's instructions. Briefly, the cells were fixed with 4%
paraformaldehyde followed by incubation with TUNEL reaction
mixtures for 1 h at 37°C. The stained cells were visualized and
counted using a fluorescence microscope (Olympus, Tokyo, Japan).
Five fields (magnification, ×400) were randomly selected for the
measurement of apoptotic cells in a blinded manner.
Dual-luciferase reporter assay
The beclin-1 3′-UTR and mutated 3′-UTR constructs
were amplified and subcloned into pGL3 Luciferase Promoter Vector
(Promega, Madison, WI, USA) with XbaI and NotI
restriction sites. Using Lipofectamine transfection reagent
(Invitrogen-Life Technologies) the pGL3 vector containing beclin-1
3′-UTR or mutated forms was co-transfected with or without 20
nmol/l miR-216a mimic (GenePharma, Shanghai, China) into PANC-1
cells according to the manufacturer's instructions. The cells were
collected after 48-h transfection and the luciferase activity was
measured using the Dual-Luciferase Reporter Assay kit (Promega).
The relative protein expression levels of beclin-1 and LC3-II were
quantified using Image-Pro Plus 6.0 software. The relative
luciferase activities were normalized with GAPDH to that of the
control cells.
Reverse transcriptase-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated by using TRIzol
(Invitrogen-Life Technologies) and small RNAs were extracted by
using mirVana kits (Ambion Inc, Austin, TX, USA) according to the
manufacturer's instructions. Corresponding cDNA was generated by
using M-MLV reverse transcriptase (Clontech, Palo Alto, CA, USA)
and the TaqMan miRNA Reverse Transcription kit (Applied Biosystems,
Foster City, CA, USA) according to the manufacturer's instructions.
To analyze the gene expression levels, the RT-qPCR mixture system
containing cDNA templates, primers and SYBR-Green qPCR Master Mix
were subjected to RT-qPCR quantification.
Glyceraldehyde-3-phosphate dehydrogenase (GAPDH; for beclin-1) and
U6 SnRNA (for miR-216a) were used as an internal reference and
relative gene expression was quantified by 2−ΔΔCt
method.
Western blot analysis
Total cell lysates were separated by 10% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis and transferred
to nitrocellulose membranes (Amersham, Little Chalfont, UK).
Non-fat dry milk (2.5%) was used for blocking the membrane and
primary antibodies including anti-LC3 antibody (ab63817) and
anti-beclin-1 (ab62557) (both from Abcam, Cambridge, UK),
anti-cleaved caspase-3 (PC679-50UG) (from Millipore, Boston, MA,
USA) and anti-GAPDH antibody (bs-2188R) (Bioss, Beijing, China)
were used for detection of the target protein. The target protein
was visualized by using an enhanced chemiluminescence (ECL)
detection system (Amersham).
Tumorigenicity assay
Cells (2×106) diluted in 200 µl
PBS were injected subcutaneously into the right groin of BALB/c
nude mice. The tumor volume was measured daily and the tumor was
irradiated with a dose of irradiation (10 Gy) when the volume
reached ~500 mm3. The length and width were measured and
the volume was calculated using the formula: length ×
width2 × π/6.
Statistical analysis
Data were presented as the mean ± standard deviation
(SD) of three or more independent experiments. Levels of
significance between or among groups were analyzed by the
two-tailed Student's t-test or the one-way ANOVA, respectively.
Results were considered statistically significant at P<0.05.
Results
Irradiation induces miR-216a inhibition
and autophagy activation in PANC-1 cells
To investigate the potential role of miR-216a in
radioresistance of pancreatic cancer cells, we first examined the
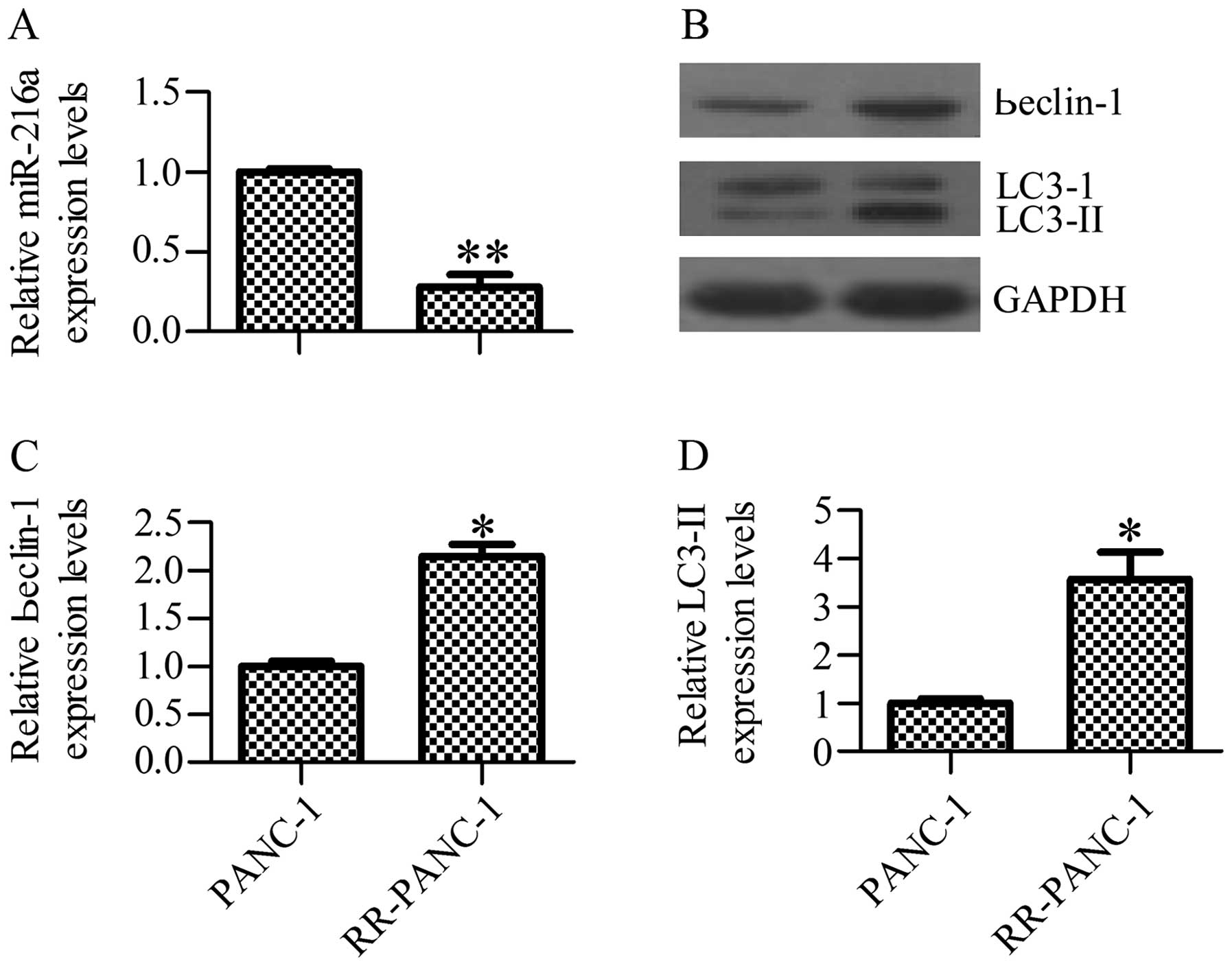
miR-216a levels in radioresistant cells using RT-qPCR. The results
showed that miR-216a expression levels were significantly inhibited
in radioresistant (RR)-PANC-1 cells as compared with the control
PANC-1 cells (Fig. 1A). We also
observed an increased level of autophagy activity as indicated by
upregulation of the beclin-1 and LC3-II protein levels in RR-PANC-1
cells in comparison with control cells as determined by western
blot analysis (Fig. 1B–D). The data
suggested that irradiation contributed to miR-216a inhibition and
autophagy activation.
Forced expression of miR-216a inhibits
the irradiation-induced upregulation of beclin-1 and autophagy
activity
To investigate whether the inhibited miR-216a
expression has a certain correlation with the increased autophagy
activity, we treated the RR-PANC-1 cells with miR-216a mimics and
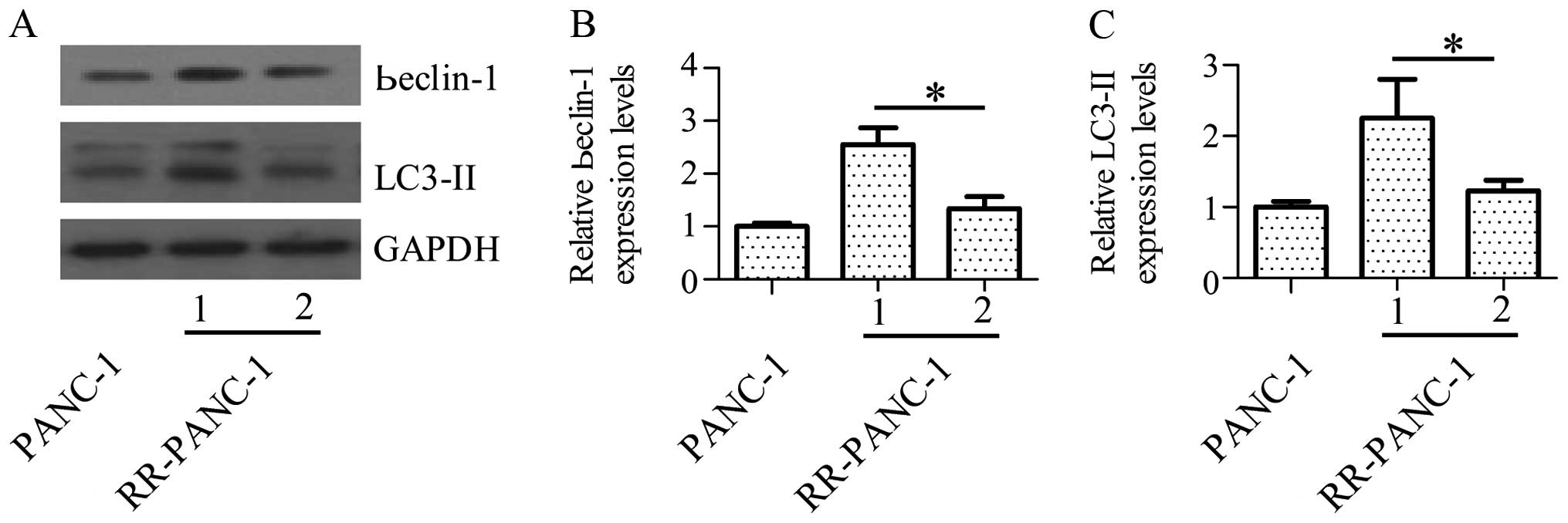
determined the alterations of autophagy activity. The results
exhibited that forced expression of miR-216a significantly
suppressed beclin-1 and LC3-II protein expression levels (Fig. 2A and B), suggesting that miR-216a
played an important role in the regulation of autophagy activity.
Fig. 2C shows that forced
expression of miR-216a had suppressed LC3-II protein expression
levels significantly too.
miR-216a directly targets beclin-1 to
regulate autophagy activity
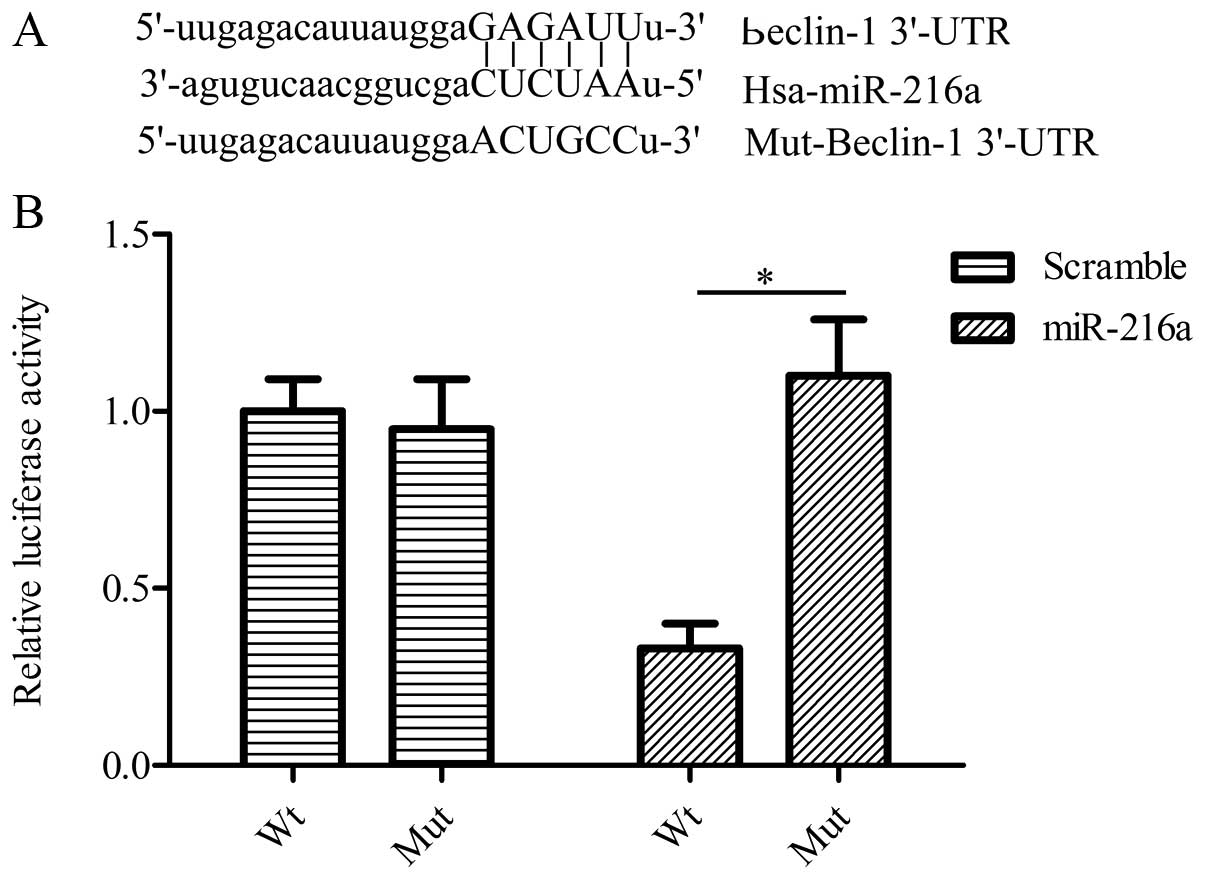
To investigate the relationship between miR-216a and
autophagy, we screened whether miR-216a has a direct interaction
with autophagy-related genes using bioinformatics analysis. As
expected, we found that beclin-1 was a putative target gene of
miR-216a (Fig. 3A). To determine
whether beclin-1 3′-UTR was responsive to miR-216a, we performed
the dual-luciferase reporter assay by using pGL3-beclin-1-3′-UTR
and miR-216a mimics. Co-transfection of pGL3-beclin-1-3′-UTR with
miR-216a mimics in PANC-1 cells significantly downregulated the
relative luciferase activity in comparison with cells
co-transfected with miR-216a, with pGL3-beclin-1-mut-3′-UTR
containing a mutation in the predicted consensus sequences for
miR-216a (Fig. 3B). These findings
suggested that beclin-1 is a direct target gene of miR-216a.
Forced expression of miR-216a reduces
cell growth and colony formation in RR-PANC-1 cells in response to
irradiation
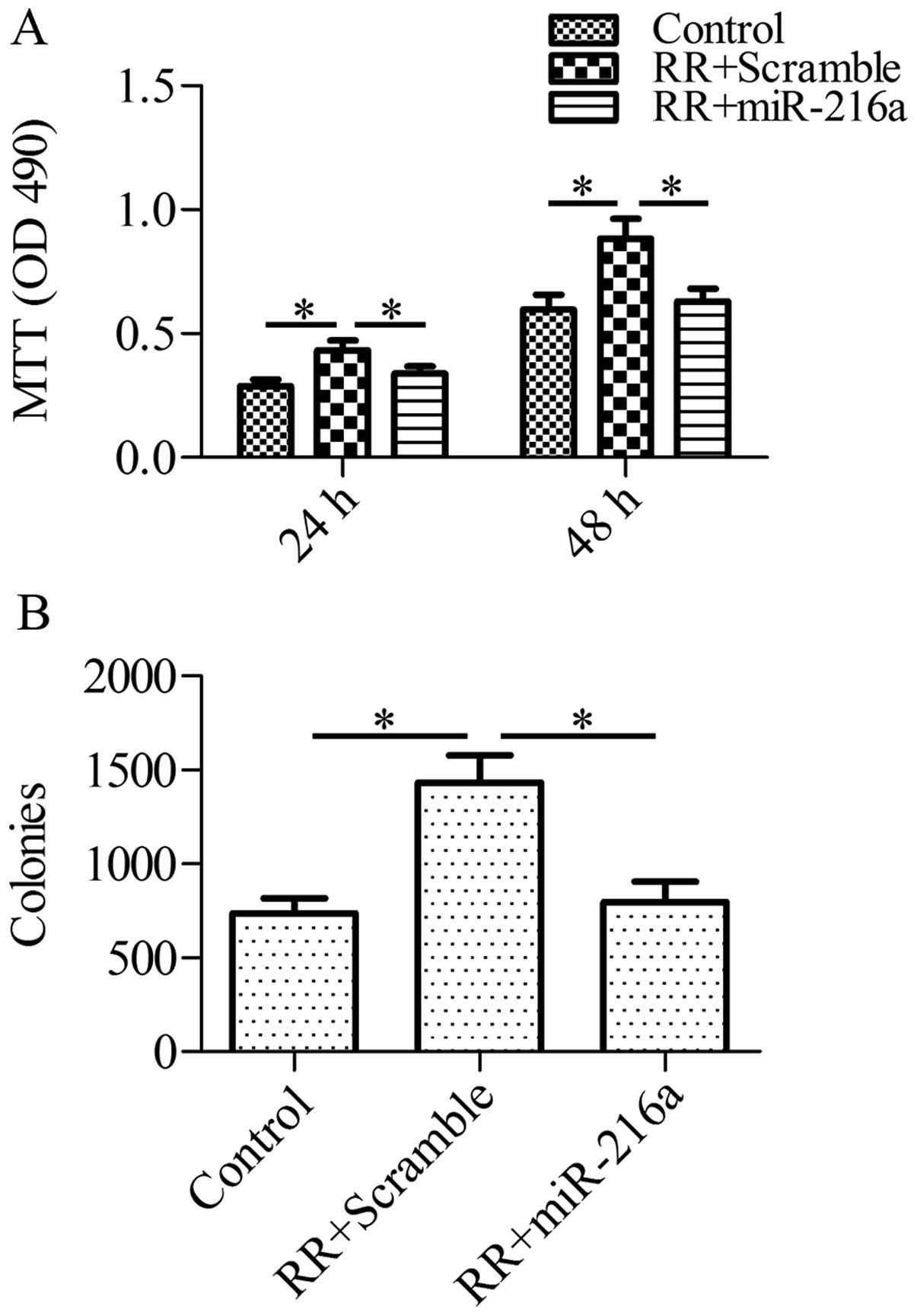
To assess whether miR-216a-mediated autophagy
inhibition via beclin-1 plays an important role in the growth of RR
cells in response to irradiation, we forced the expression of
miR-216a in RR cells and analyzed the cell growth and colony
formation after irradiation. RR-PANC-1 cells were pretreated with
miR-216a mimics or control scramble miRNA for 1 h and then
subjected to irradiation (2 Gy/min). Using the MTT assay, we found
that the forced expression of miR-216a markedly sensitized RR cells
to cell death in response to irradiation, as compared with the
control group (Fig. 4A).
Furthermore, the colony formation ability of RR-PANC-1 cells was
inhibited by miR-216a (Fig. 4B).
These data suggested that miR-216a sensitized RR-PANC-1 cells to
irradiation.
Forced expression of miR-216a sensitizes
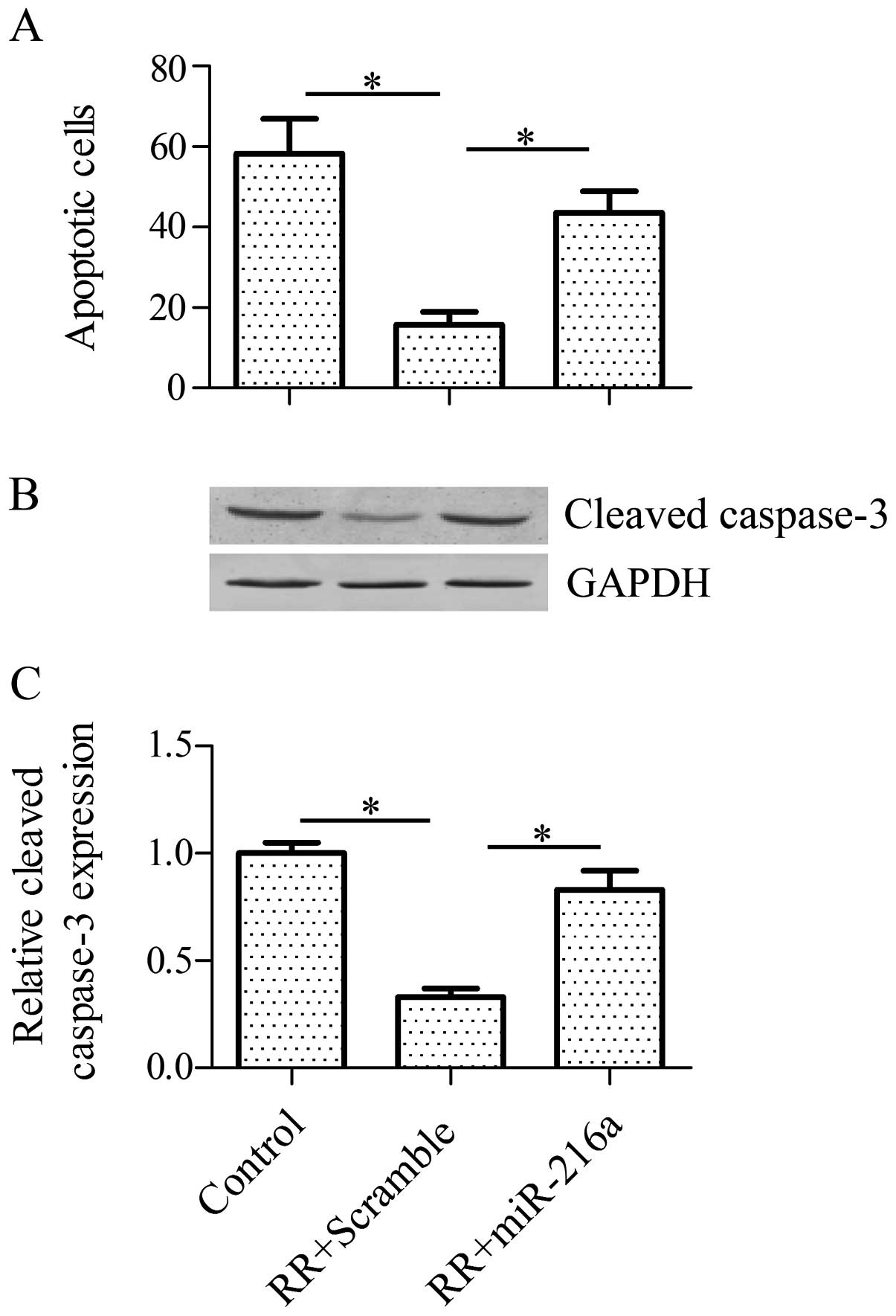
RR-PANC-1 cells to irradiation by elevating cell apoptosis
To study the role of miR-216a on RR cells, we
detected the effect of miR-216a on cell apoptosis in RR cells in
response to irradiation. The results of the TUNEL assay indicated
that a forced expression of miR-216a significantly increased the
cell apoptosis of RR-PANC-1 cells in comparison with the control
(Fig. 5A). In addition, the
pro-apoptotic protein expression level of cleaved caspase-3 was
also increased by miR-216a treatment (Fig. 5B and C). Collectively, these results
suggested that miR-216a enhanced the radiosensitivity of RR-PANC-1
cells via activation of cell apoptosis.
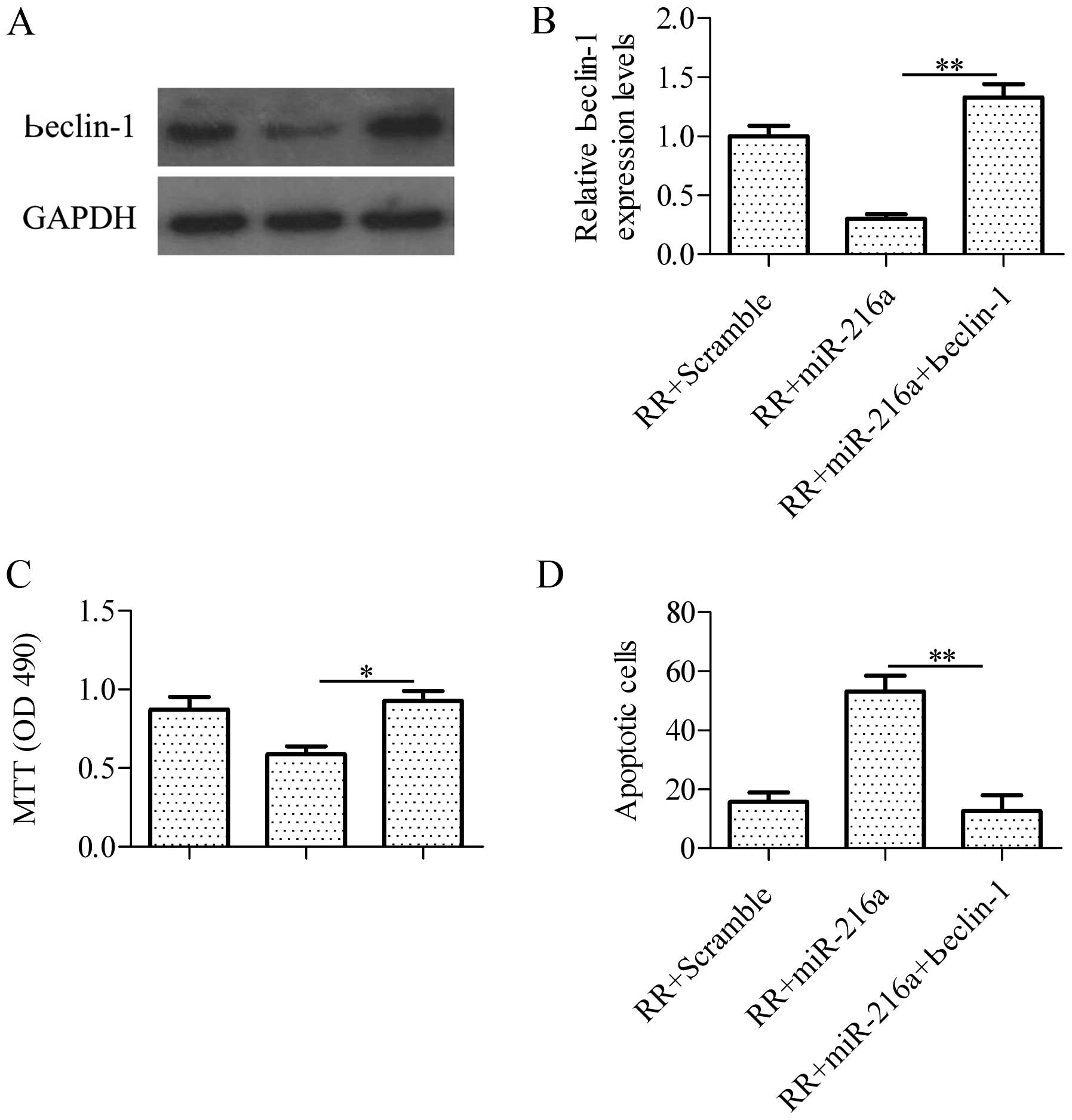
Overexpression of beclin-1 abrogates
miR-216a-induced irradiation sensitivity
To verify whether miR-216a sensitized RR-PANC-1
cells to irradiation by regulating beclin-1, we co-transfected
miR-216a with beclin-1 overexpression vectors harboring no specific
miR-216a binding specific sequences in 3′-UTR in RR-PANC-1 cells.
The results showed that over-expression of beclin-1 (Fig. 6A and B) significantly blocked
miR-216a-induced cell growth inhibition in RR-PANC-1cells in
response to irradiation detected by the MTT assay (Fig. 6C). Furthermore, overexpression of
beclin-1 abrogated cell apoptosis induced by miR-216a in RR-PANC-1
cells as indicated by the TUNEL assay (Fig. 6D). These findings suggested that
beclin-1 is important in regulating the irradiation sensitivity in
RR cells, further confirming that miR-216a targeted beclin-1 to
sensitized RR-PANC-1 cells to irradiation.
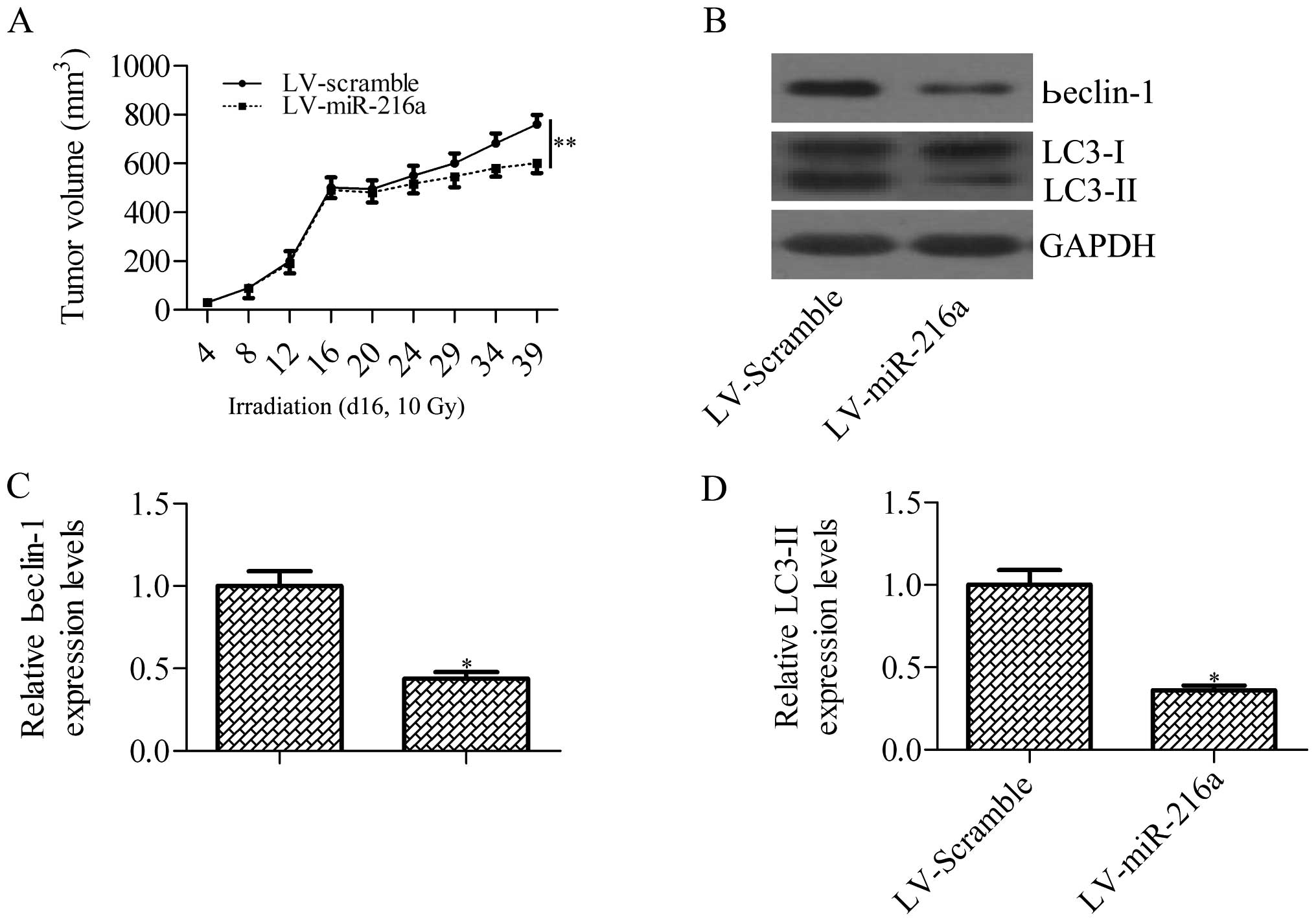
miR-216a sensitizes pancreatic cancer
cells to irradiation treatment by inhibiting irradiation-induced
autophagy in xenograft tumor
To assess whether miR-216a can increase the
efficiency of irradiation in killing implanted tumors in nude mice,
nude mice were subcutaneously injected with PANC-1 cells
pre-transfected with lentiviral vector expressing miR-216a or
scramble miRNA control. When the tumors reached ~500 mm3
on day 16, the tumors received a single dose of 10-Gy irradiation.
The results showed that force expression of miR-216a significantly
increased the radiosensitivity of PANC-1-derived tumors on
irradiation treatment (Fig. 7A). We
further analyzed beclin-1 expression and autophagy activity in the
xenograft samples after irradiation treatment. Consistent with the
above results, miR-216a inhibited irradiation-induced beclin-1
expression and autophagy activity (Fig.
7B–D). These results demonstrated that miR-216a sensitizes
pancreatic cancer cells to irradiation treatment by blocking
irradiation-induced beclin-1-mediated autophagy.
Discussion
The role of miR-216a in cancer has increasingly
drawn attention. The transcription of miR-216a has been found to be
stimulated by the androgen pathway which targets the tumor
suppressor in lung cancer-1 gene in the early stage of
carcinogenesis (25). miR-216a
targets phosphatase and tensin homolog and decapentaplegic homolog
7 to induce mesenchymal transition in hepatocellular carcinoma
(26). miR-216a seems to act as an
oncogene in hepatocellular carcinoma. However, miR-216a was
reported to be downregulated in pancreatic cancer (21,22).
The downregulation of miR-216a in feces has been suggested as a
biomarkers for pancreatic cancer (23). In pancreatic intraepithelial
neoplasms, miR-216a was also found to be decreased (27). The suppressed expression patterns of
miR-216a in pancreatic cancer suggests that miR-216a plays an
important role in the tumorigenesis of pancreatic cancers and is a
promising molecular target for the treatment of pancreatic
cancer.
As expected, we found that miR-216a was involved in
the regulation of radioresistance through beclin-1-mediated
autophagy. Autophagy, an adaptive response against cellular streak,
has been currently suggested to be involved in the radioresistance
of cancer cells (8). However, the
role of autophagy in the radioresistance of cancer cells remains
controversial. γ-radiation induces autophagy, which is responsible
for the radioresistance of glioma stem cells (28,29).
In breast cancer, inhibition of autophagy promoted
radio-sensitivity via the suppression of transforming growth
factor-activated kinase-1 (8).
Hypoxia-induced autophagy was suggested to attribute to the
radioresistance of breast cancer cells (30). Chaachouay et al have
demonstrated that radioresistant breast cancer cells exhibit a high
level of autophagy serving as a protective and survival mechanism
against irradiation (31).
Anticancer drugs, Akt inhibitors, were revealed to promote
radiosensitivity through induction of autophagy (32). Similarly, the inhibitors of the
mammalian target of rapamycin radiosensitized non-small cell lung
cancer cells harboring phosphatase and tensin homolog deficient and
epidermal growth factor receptor activating mutant (33). The apparent dual role of autophagy
in the irradiation of cancer cells remains elusive, suggesting that
autophagy has different biological functions due to the different
cell type or stimuli.
The role of autophagy in the regulation of
radioresistance of pancreatic cancer has been investigated.
Profilin1 has been indicated to be capable of sensitizing
pancreatic cancer cells to irradiation by inhibiting autophagy
(34). More recently, the
overexpression of miR-23b has been shown to decrease
radiation-induced autophagy and increase the radiosensitivity of
pancreatic cancer cells (35).
Consistently, the inhibition of autophagy by miR-216a was found to
enhance the radio-sensitivity of pancreatic cancer cells in our
study. We have demonstrated that miR-216a was significantly
downregulated by irradiation and the autophagy activity was
increased in pancreatic cancer cells in response to irradiation,
suggesting a potential association between autophagy and miR-216a.
As expected, we further identified and characterized that miR-216a
targeted a critical autophagic gene, beclin-1, to inhibit autophagy
activation in response to irradiation in pancreatic cancer cells.
Consistent with our findings, Menghini et al have
demonstrated that miR-216a targets beclin-1 to regulate autophagy
in endothelial cells contributing to the endothelial function in
cardiovascular diseases (36).
miR-30a sensitizes cancer cells to cis-platinum via
suppression of beclin 1-mediated autophagy (37). The down-regulation of
miR-17-5p-induced beclin-1 overexpression leads to paclitaxel
resistance in lung cancer cells (38). beclin-1 has been suggested to be
important in the chemoradiation which affects the overall survival
of patients with esophageal squamous cell carcinoma (39). Therefore, considering the role of
beclin-1 in anticancer therapy, targeting beclin-1 may have a
better outcome in the treatment of pancreatic cancer. In
conclusion, our results provide evidence that miR-216a inhibited
beclin-1 leading to the downregulation of autophagy induced by
irradiation, which enhanced the radiosensitivity of pancreatic
cancer cells.
Acknowledgments
The present study was supported by the Guangdong
Province Natural Science Foundation (S2013010016662), the Health
Bureau of Guangdong Province (A2014224 and B2014196), the Science
and Technology Planning Project of Guangdong Province
(2013B021800284) and the National Natural Science Foundation of
China (81201932 and 81372493).
Abbreviations:
|
miRNAs
|
microRNAs
|
|
UTR
|
untranslated region
|
|
LC3
|
microtubule-associated protein light
chain 3
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide
|
|
TUNEL
|
terminal deoxynucleotidyl transferase
dUTP nick end-labeling
|
References
|
1
|
Vincent A, Herman J, Schulick R, Hruban RH
and Goggins M: Pancreatic cancer. Lancet. 378:607–620. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li D, Xie K, Wolff R and Abbruzzese JL:
Pancreatic cancer. Lancet. 363:1049–1057. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Neoptolemos JP, Dunn JA, Stocken DD,
Almond J, Link K, Beger H, Bassi C, Falconi M, Pederzoli P,
Dervenis C, et al European Study Group for Pancreatic Cancer:
Adjuvant chemoradiotherapy and chemotherapy in resectable
pancreatic cancer: A randomised controlled trial. Lancet.
358:1576–1585. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Girard N, Mornex F, Bossard N, Ychou M,
Chauffert B and Wautot V: Estimating optimal dose of twice-weekly
gemcitabine for concurrent chemoradiotherapy in unresectable
pancreatic carcinoma: Mature results of GEMRT-01 Phase I trial. Int
J Radiat Oncol Biol Phys. 77:1426–1432. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Crane CH, Abbruzzese JL, Evans DB, Wolff
RA, Ballo MT, Delclos M, Milas L, Mason K, Charnsangavej C, Pisters
PW, et al: Is the therapeutic index better with gemcitabine-based
chemoradiation than with 5-fluorouracil-based chemoradiation in
locally advanced pancreatic cancer? Int J Radiat Oncol Biol Phys.
52:1293–1302. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chatterjee S, Willis N, Locks SM, Mott JH
and Kelly CG: Dosimetric and radiobiological comparison of helical
tomotherapy, forward-planned intensity-modulated radiotherapy and
two-phase conformal plans for radical radiotherapy treatment of
head and neck squamous cell carcinomas. Br J Radiol. 84:1083–1090.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Han MW, Lee JC, Choi JY, Kim GC, Chang HW,
Nam HY, Kim SW and Kim SY: Autophagy in hibition can overcome
radio-resistance in breast cancer cells through suppression of TAK1
activation. Anticancer Res. 34:1449–1455. 2014.PubMed/NCBI
|
|
9
|
Kondo Y, Kanzawa T, Sawaya R and Kondo S:
The role of autophagy in cancer development and response to
therapy. Nat Rev Cancer. 5:726–734. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Maiuri MC, Zalckvar E, Kimchi A and
Kroemer G: Self-eating and self-killing: Crosstalk between
autophagy and apoptosis. Nat Rev Mol Cell Biol. 8:741–752. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mizushima N, Ohsumi Y and Yoshimori T:
Autophagosome formation in mammalian cells. Cell Struct Funct.
27:421–429. 2002. View Article : Google Scholar
|
|
12
|
Liang B, Kong D, Liu Y, Liang N, He M, Ma
S and Liu X: Autophagy inhibition plays the synergetic killing
roles with radiation in the multi-drug resistant SKVCR ovarian
cancer cells. Radiat Oncol. 7:2132012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gewirtz DA, Hilliker ML and Wilson EN:
Promotion of autophagy as a mechanism for radiation sensitization
of breast tumor cells. Radiother Oncol. 92:323–328. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zois CE and Koukourakis MI:
Radiation-induced autophagy in normal and cancer cells: Towards
novel cytoprotection and radio-sensitization policies? Autophagy.
5:442–450. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mendell JT and Olson EN: MicroRNAs in
stress signaling and human disease. Cell. 148:1172–1187. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Winter J, Jung S, Keller S, Gregory RI and
Diederichs S: Many roads to maturity: microRNA biogenesis pathways
and their regulation. Nat Cell Biol. 11:228–234. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang P, Chen L, Zhang J, Chen H, Fan J,
Wang K, Luo J, Chen Z, Meng Z and Liu L: Methylation-mediated
silencing of the miR-124 genes facilitates pancreatic cancer
progression and metastasis by targeting Rac1. Oncogene. 33:514–524.
2014. View Article : Google Scholar
|
|
19
|
Oh JS, Kim JJ, Byun JY and Kim IA:
Lin28-let7 modulates radiosensitivity of human cancer cells with
activation of K-Ras. Int J Radiat Oncol Biol Phys. 76:5–8. 2010.
View Article : Google Scholar
|
|
20
|
Wang P, Zhuang L, Zhang J, Fan J, Luo J,
Chen H, Wang K, Liu L, Chen Z and Meng Z: The serum miR-21 level
serves as a predictor for the chemosensitivity of advanced
pancreatic cancer, and miR-21 expression confers chemoresistance by
targeting FasL. Mol Oncol. 7:334–345. 2013. View Article : Google Scholar
|
|
21
|
Hou B, Jian Z, Chen S, Ou Y, Li S and Ou
J: Expression of miR-216a in pancreatic cancer and its clinical
significance. Nan Fang Yi Ke Da Xue Xue Bao. 32:1628–1631. 2012.In
Chinese. PubMed/NCBI
|
|
22
|
Ali S, Banerjee S, Logna F, Bao B, Philip
PA, Korc M and Sarkar FH: Inactivation of Ink4a/Arf leads to
deregulated expression of miRNAs in K-Ras transgenic mouse model of
pancreatic cancer. J Cell Physiol. 227:3373–3380. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Link A, Becker V, Goel A, Wex T and
Malfertheiner P: Feasibility of fecal microRNAs as novel biomarkers
for pancreatic cancer. PLoS One. 7:e429332012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Skvortsov S, Jimenez CR, Knol JC,
Eichberger P, Schiestl B, Debbage P, Skvortsova I and Lukas P:
Radioresistant head and neck squamous cell carcinoma cells:
Intracellular signaling, putative biomarkers for tumor recurrences
and possible therapeutic targets. Radiother Oncol. 101:177–182.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen PJ, Yeh SH, Liu WH, Lin CC, Huang HC,
Chen CL, Chen DS and Chen PJ: Androgen pathway stimulates
microRNA-216a transcription to suppress the tumor suppressor in
lung cancer-1 gene in early hepatocarcinogenesis. Hepatology.
56:632–643. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xia H, Ooi LL and Hui KM:
MicroRNA-216a/217-induced epithelial-mesenchymal transition targets
PTEN and SMAD7 to promote drug resistance and recurrence of liver
cancer. Hepatology. 58:629–641. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu J, Li A, Hong SM, Hruban RH and Goggins
M: MicroRNA alterations of pancreatic intraepithelial neoplasias.
Clin Cancer Res. 18:981–992. 2012. View Article : Google Scholar :
|
|
28
|
Lomonaco SL, Finniss S, Xiang C,
Decarvalho A, Umansky F, Kalkanis SN, Mikkelsen T and Brodie C: The
induction of autophagy by gamma-radiation contributes to the
radioresistance of glioma stem cells. Int J Cancer. 125:717–722.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhuang W, Qin Z and Liang Z: The role of
autophagy in sensitizing malignant glioma cells to radiation
therapy. Acta Biochim Biophys Sin (Shanghai). 41:341–351. 2009.
View Article : Google Scholar
|
|
30
|
He WS, Dai XF, Jin M, Liu CW and Rent JH:
Hypoxia-induced autophagy confers resistance of breast cancer cells
to ionizing radiation. Oncol Res. 20:251–258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chaachouay H, Ohneseit P, Toulany M,
Kehlbach R, Multhoff G and Rodemann HP: Autophagy contributes to
resistance of tumor cells to ionizing radiation. Radiother Oncol.
99:287–292. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fujiwara K, Iwado E, Mills GB, Sawaya R,
Kondo S and Kondo Y: Akt inhibitor shows anticancer and
radiosensitizing effects in malignant glioma cells by inducing
autophagy. Int J Oncol. 31:753–760. 2007.PubMed/NCBI
|
|
33
|
Kim EJ, Jeong JH, Bae S, Kang S, Kim CH
and Lim YB: mTOR inhibitors radiosensitize PTEN-deficient
non-small-cell lung cancer cells harboring an EGFR activating
mutation by inducing autophagy. J Cell Biochem. 114:1248–1256.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cheng H, Li J, Liu C, Yao W, Xu Y, Frank
TS, Cai X, Shi S, Lu Y, Qin Y, et al: Profilin1 sensitizes
pancreatic cancer cells to irradiation by inducing apoptosis and
reducing autophagy. Curr Mol Med. 13:1368–1375. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang P, Zhang J, Zhang L, Zhu Z, Fan J,
Chen L, Zhuang L, Luo J, Chen H, Liu L, et al: MicroRNA 23b
regulates autophagy associated with radioresistance of pancreatic
cancer cells. Gastroenterology. 145:1133–1143. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Menghini R, Casagrande V, Marino A,
Marchetti V, Cardellini M, Stoehr R, Rizza S, Martelli E, Greco S,
Mauriello A, et al: miR-216a: A link between endothelial
dysfunction and autophagy. Cell Death Dis. 5:e10292014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zou Z, Wu L, Ding H, Wang Y, Zhang Y, Chen
X, Chen X, Zhang CY, Zhang Q and Zen K: microRNA-30a sensitizes
tumor cells to cis-platinum via suppressing beclin 1-mediated
autophagy. J Biol Chem. 287:4148–4156. 2012. View Article : Google Scholar :
|
|
38
|
Chatterjee A, Chattopadhyay D and
Chakrabarti G: miR-17-5p downregulation contributes to paclitaxel
resistance of lung cancer cells through altering beclin1
expression. PLoS One. 9:e957162014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen Y, Li X, Wu X, He C, Guo L, Zhang S,
Xiao Y, Guo W and Tan B: Autophagy-related proteins LC3 and
beclin-1 impact the efficacy of chemoradiation on esophageal
squamous cell carcinoma. Pathol Res Pract. 209:562–567. 2013.
View Article : Google Scholar : PubMed/NCBI
|