Introduction
The secreted frizzled-related protein 2 (SFRP2)
belongs to a large family of SFRPs consisting of SFRP1-5, and is
homologous to the extracellular portion of the Wingless-type (Wnt)
receptor Frizzled (1). The Wnt
signaling pathway plays important roles in carcinogenesis,
embryonic development and neurodegenerative disease (2), and has been extensively studied in
human cancer. It is reported to be aberrantly activated in various
human cancers, such as non-small cell lung cancer (NSCLC) (3), colorectal cancer (4), melanoma (5) and leukemia (6).
The secreted ligands of Wnt pathway possesses 19
highly conserved and hydrophobic glycoproteins (7), and the receptor of this signaling is a
protein complex consisting of Frizzled and lipoprotein
receptor-related protein (LRP) (1).
There are canonical and non-canonical Wnt signaling (1). The former is dependent on β-catenin,
while the latter is not and may proceed through calcium flux, G
proteins, and JNK (8). In the
activated state of canonical pathway, the ligand binds to
Frizzled/LRP receptor complex and released cytosolic β-catenin from
a degradation complex consisting of adenomatous polyposis coli
(APC), Axin and glycogen synthase kinase-3β (GSK-3β) (1,9).
Consequently, β-catenin accumulates in the cytoplasm and
subsequently translocates to the nucleus, where it interacts with
T-cell factor/lymphoid enhancer factor (TCF/LEF) transcription
factors and turns on the downstream genes, including CCND1,
Axin2, c-myc and MMP-7 (10).
Considering the homology of SFRP2 and extracellular
portion of Frizzled, it is reported that SFRP2 is an antagonist to
Wnt pathway by competing for Wnt binding to Frizzled (11). However, there are reports claiming
that SFRP2 is an agonist of the Wnt pathway (12–17).
Recently, investigators have shown that SFRP2 inhibition reduced
tumor volume of breast cancer to 46% compared with control mice
(18), suggesting that SFRP2 is a
tumor-promoting protein. Consistent with this, another recent
report showed that SFRP2 intensity increased with tumor size in
murine angiosarcoma (19).
For the purpose of clarifying whether SFRP2 is an
oncoprotein or a tumor suppressor in lung cancer, we assessed the
effects of SFRP2 on Wnt signaling pathway as well as on lung cancer
cell proliferation, apoptosis, migration and invasion. The
molecular mechanisms underlying the functions of SFRP2 in lung
cancer were also investigated.
Materials and methods
Cell lines and culture conditions
Human lung cancer cell lines 95-D, SPCA-1, and A549
were purchased from the Cell Bank of Type Culture Collection of
Chinese Academy of Sciences (Shanghai, China). All these cells were
cultured in RPMI-1640 medium supplemented with 10% fetal bovine
serum, 100 U/ml penicillin and 100 µg/ml streptomycin (Life
Technologies, Carlsbad, CA, USA).
Cell transfection
Three pairs of siRNAs were synthesized by Life
Technologies (Shanghai, China). The expression plasmid containing
SFRP2 coding region was purchased from ViGene BioSciences
(Shandong, China) and confirmed by DNA sequencing. The day before
transfection, cells were plated into 6-well plates and cultured
overnight. Cell transfection experiments were performed according
to the transfection reagent manufacturer's instructions. In brief,
2 µg of SFRP2 siRNA or expression plasmid and their
respective control DNAs were re-suspended in Opti-MEM (Life
Technologies, Carlsbad, CA, USA) to 100 µl in a tube;
meanwhile, 10 µl X-tremeGENE siRNA transfection reagent
(Roche, Indianapolis, IN, USA) was added to 90 µl Opti-MEM
in another tube. Transfection reagent in the second tube was then
added into the first tube containing DNA, and mixed and incubated
at room temperature for 20 min. After that, the complex was added
to the cells in 6-well plates and the cells were cultured for an
additional 24–48 h.
RNA isolation and real-time RT-qPCR
Total RNA was isolated by TRIzol (Life Technologies,
Carlsbad, CA, USA), and then quantified using the NanoDrop ND-1000
Spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA).
For RT-qPCR, 2 µg of total RNA and primers were mixed with
regents supplied by Quant One Step RT-qPCR (SYBR Green I) kit
(Tiangen, Beijing, China) according to the instructions. Specific
primers for mRNA detection are indicated in Table I. The qPCR was carried out using the
Bio-Rad CFX96 Real-Time PCR system (Bio-Rad, Hercules, CA, USA),
using the following reaction conditions: the first step at 95°C for
5 min, followed by 35 amplification cycles composed of 95°C for 10
sec, 59°C for 20 sec and 72°C for 20 sec. All reactions were
performed in triplicate and included negative controls (sterilized
water as template). Results were processed and exported using the
Bio-Rad CFX Manager software, version 1.1 (Bio-Rad). The expression
of target mRNA was normalized to glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) mRNA, and determined using the
2−ΔΔCT method. All the experiments were repeated at
least three times.
 | Table IPrimers for real-time RT-qPCR. |
Table I
Primers for real-time RT-qPCR.
| Gene | Forward primers
(5′→3′) | Reverse primers
(5′→3′) |
|---|
| CDK4 |
CACAGTTCGTGAGGTGGCTTTA |
GATGTGGCACAGACGTCCATC |
| CDK6 |
TCATTCAAAATCTGCCCAACC |
TCCTGGAAGTATGGGTGAGACA |
| CCNE1 |
GACTGCCTTGAATTTCCTTATGGT |
TCGCACCACTGATACCCTGAA |
| CCND1 |
GCATCTACACCGACAACTCCA |
CCAGGTTCCACTTGAGCTTG |
| p27 |
AGAAGCCTGGCCTCAGAAGAC |
CCATTCCATGAAGTCAGCGATA |
| GAPDH |
GAAGGTCGGAGTCAACGGAT |
CTGGAAGATGGTGATGGGATT |
| SFRP2 |
ATGATGATGACAACGACATAATG |
GAGCCACAGCACCGATTT |
| WIF-1 |
AATGCCAATGTCAAGAAGG |
GATGTCGGAGTTCACCAGA |
| APC |
CCAACAAGGCTACGCTAT |
CTGCTCGCCAAGACAAAT |
| Wnt5a |
GAGTGCTCGCATCCTCAT |
GCATGTCTTCAGGCTACA |
Protein extraction and western
blotting
Cell lysates were obtained in lysis buffer (20 mM
Tris-HCl, pH 7.5, 1% NP-40, 5 mM sodium pyrophosphate, 5 mM EDTA,
pH 8.0, 1 mM sodium vanadate, 150 mM NaCl, 10 µg/ml
aprotinin, 10 µg/ml leupeptin and 1 µM PMSF). After
quantification and denaturation, equal proteins of each sample were
subjected to 10% SDS-PAGE and transferred to a polyvinylidene
fluoride (PVDF) membrane (Bio-Rad). The membrane was first blocked
with 5% non-fat dry milk for 1 h at room temperature, and then
incubated with specific primary antibodies overnight at 4°C, such
as antibodies against SFRP2, Cyclin D1, Cyclin E1, CDK4, CDK6, p27,
Vimentin, E-cadherin, β-catenin, P-β-catenin and β-actin (Abcam,
Cambridge, MA, USA, 1:1,000). Next, the membrane was washed and
incubated with the secondary antibodies against rabbit or mouse for
1 h at room temperature. Lastly, the membrane was exposed to
enhanced electrochemiluminescence (ECL) following the detection kit
(KeyGen Biotechnology, Nanjing, Jiangsu, China) and photographed by
the ChemiDoc XRS gel documentation system (Bio-Rad). All the
experiments were repeated at least three times.
Cell proliferation assay
The cells were seeded at 8×103 cells/well
into 96-well plates and incubated overnight prior to transfection.
At 0, 24, 48 or 72 h after transfection, MTT (5 mg/ml in
phosphate-buffered saline) was added to each well and incubated for
4 h, and then the supernatants were removed from the wells by
aspiration. The cells were solubilized in 200 µl of dimethyl
sulfoxide (DMSO) and the absorbance was recorded on a microplate
reader at the wavelength of 490 nm. This assay was repeated at
least three times.
Wound scratch assay
After transfection, cells were allowed to recover
for 24 h, and then cells were cultured in FBS-free RPMI-1640 medium
for starvation overnight. The next day, the cell monolayers were
gently scratched with a pipette tip across the diameter of the
well, rinsed with media to remove cellular debris, and photographed
by microscopy as 0 h timepoint. After 24 h of culture, images were
taken at the same position as 0 h time-point. The healing distances
between the wound was established by calculating the ratio of the
wound surface distance change from 0 to 24 h over the wound surface
distance at 0 h. This assay was repeated at least three times.
Transwell assays
Cell suspension at 1×105 cells in 100
µl RPMI-1640 medium was added to the top chambers with
8-µm pore sized filter inserts of 24-well Transwell plates
(Costar, Cambridge, MA, USA). For invasion assay, the top surface
of filter membranes were coated with 30% of BD Matrigel matrix (BD
Biosciences, San Jose, CA, USA) before cell seeding, and placed for
4 h in an incubator. For migration assay, the top surface of filter
membranes was not coated with Matrigel. For both cell migration and
invasion assays, the following procedure was the same. The bottom
chambers were soaked by 600 µl RPMI-1640 medium supplemented
with 10% FBS. After overnight incubation, the non-migrated or
non-invaded cells on top surface of the filter membrane were
removed by cotton swabs, and the migrated or invaded cells under
the filter membrane were kept. Then the filter membranes were
submerged in methanol for 15 min at room temperature to fix the
cells. Subsequently, the cells were stained with crystal violet for
another 15 min and then PBS was used to remove the excess stain.
After the filter membranes were dried, the numbers of migrated or
invaded cells in more than three fields were counted under a
microscope, and the mean of the migrated or invaded cells for each
membrane was determined. The experiments were repeated for a
minimum of three times.
Flow cytometry
Cells at 1×106 were prepared, washed, and
resuspended in PBS. Then cells were fixed in 70% ice-cold ethanol
overnight at 4°C. After washing, cells were incubated for 15 min at
37°C in the dark in 200 µl of PI staining solution
containing 20 µg/ml PI, 100 U/ml DNase-free RNase, and 0.1%
v/v Triton X-100. All the samples were analyzed by the BD Accuri C6
(BD Biosciences).
Statistical analysis
All experimental data are presented as mean ±
standard deviation (SD). Statistical significance was evaluated
using the Student's t-test, and the significance was set at P-value
<0.05, and highly significant at ≤0.01.
Results
SFRP2 expression profiles in lung cancer
cells
To reveal SFRP2 expression profiles in lung cancer
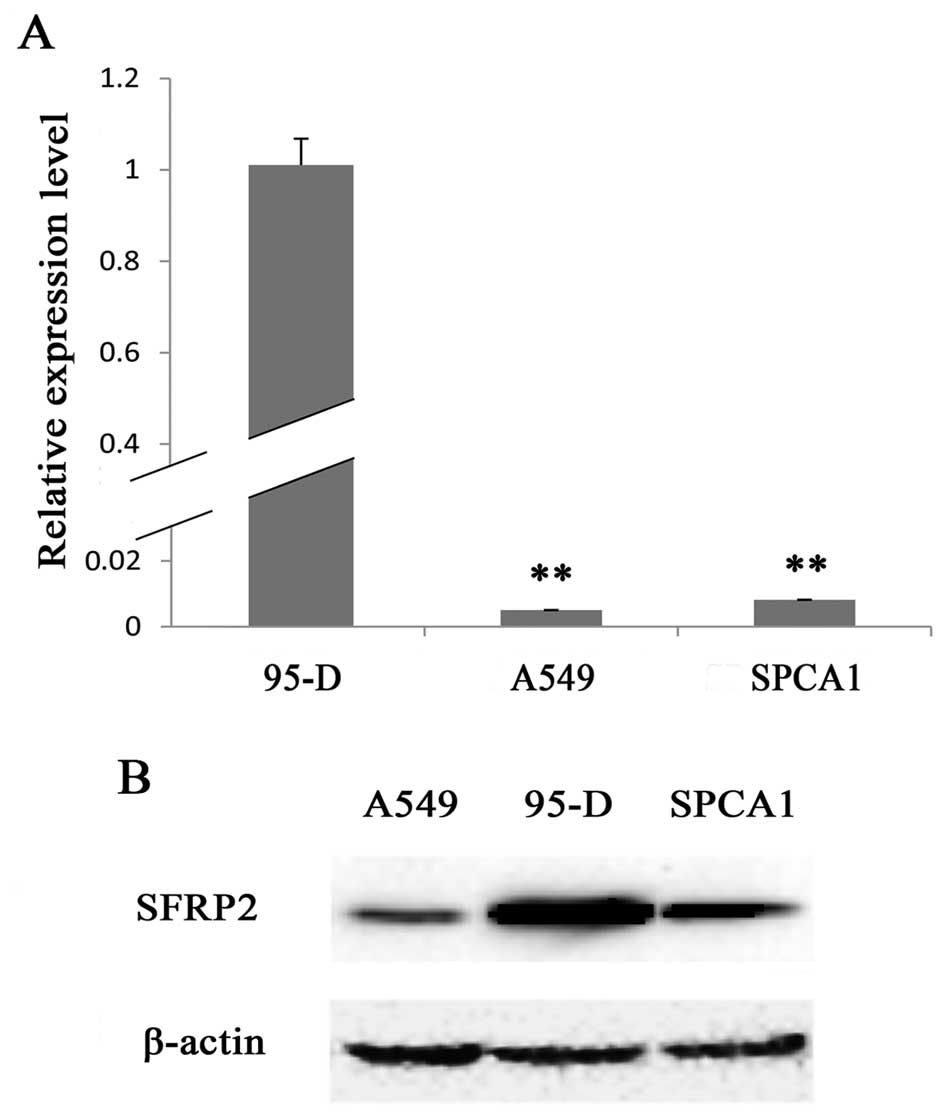
cells, we examined the lung cancer cell lines 95-D, A549 and SPCA1.
By real-time RT-PCR, we found that 95-D cells expressed the highest
level of SFRP2 mRNA, while A549 and SPCA1 cells expressed SFRP2
mRNA at significantly lower level as compared with 95-D cells
(5.01±0.00 or 8.14±0.00 vs. 100.00±6.80%, respectively; both pairs
P<0.01) (Fig. 1A). These results
were further confirmed by the data of western blotting for SFRP2
protein (Fig. 1B), which showed
extremely strong band in 95-D cell lysate, and the weakest band in
A549 cell lysate. Thus, A549 and 95-D cells were subsequently
chosen as cell models based on their remarkable difference in SFRP
expression at both transcriptional and translational levels in
order to investigate the cellular functions of SFRP2.
Cell models for SFRP2 cellular
function
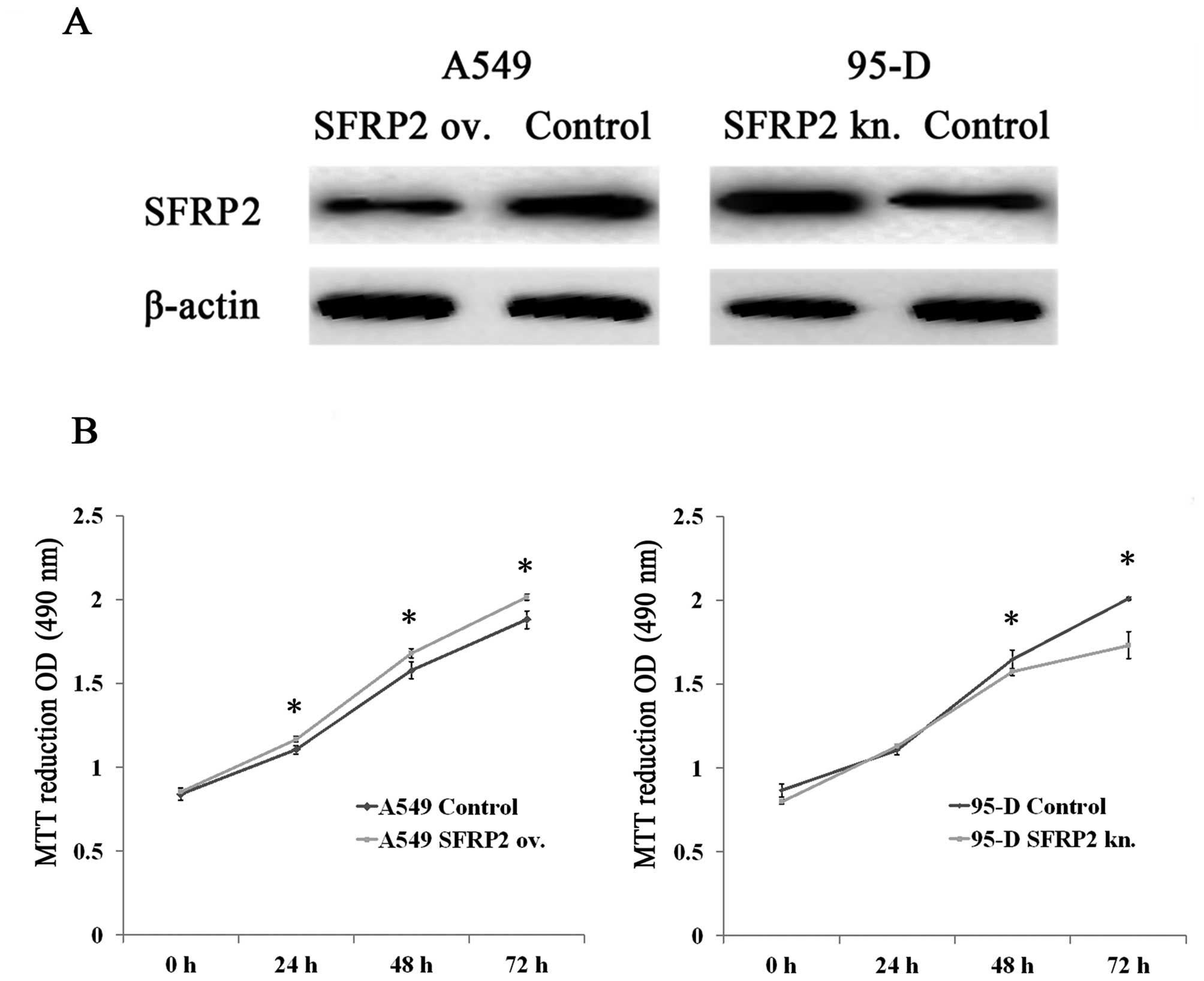
We employed A549 cells transfected with SFRP2
expressing plasmid and 95-D cells transfected with SFRP2 siRNA as
two cell models for the following study on SFRP2 cellular
functions. After transfection for 48 h, cells were lysed and
subjected to western blotting for detecting SFRP2 protein. As
indicated in Fig. 2A, SFRP2 protein
was upregulated in A549 cells after transfection with the SFRP2
expressing plasmid in comparison with the vector control cells
(left panel), whereas the level of SFRP2 protein was remarkably
reduced in 95-D cells with siRNA transfection as compared with
controls (right panel). These data suggest that two cell models for
the overexpression and knockdown of SFRP2 were established and
ready to use for subsequent experiments.
Effects of SFRP2 up- or down-regulation
on lung cancer cell proliferation
The two cell models established were applied to MTT
assay, which showed significantly higher OD490 values at 24h, 48h
and 72h after SFRP2 induction in A549 cells, compared with controls
(Fig. 2B, left panel). In contrast,
the readouts of OD490 declined significantly in 95-D cells with
SFRP2 downregulation at 48h and 72h compared with 95-D control
cells (Fig. 2B, right panel). These
results indicated that SFRP2 had the potential of promoting cell
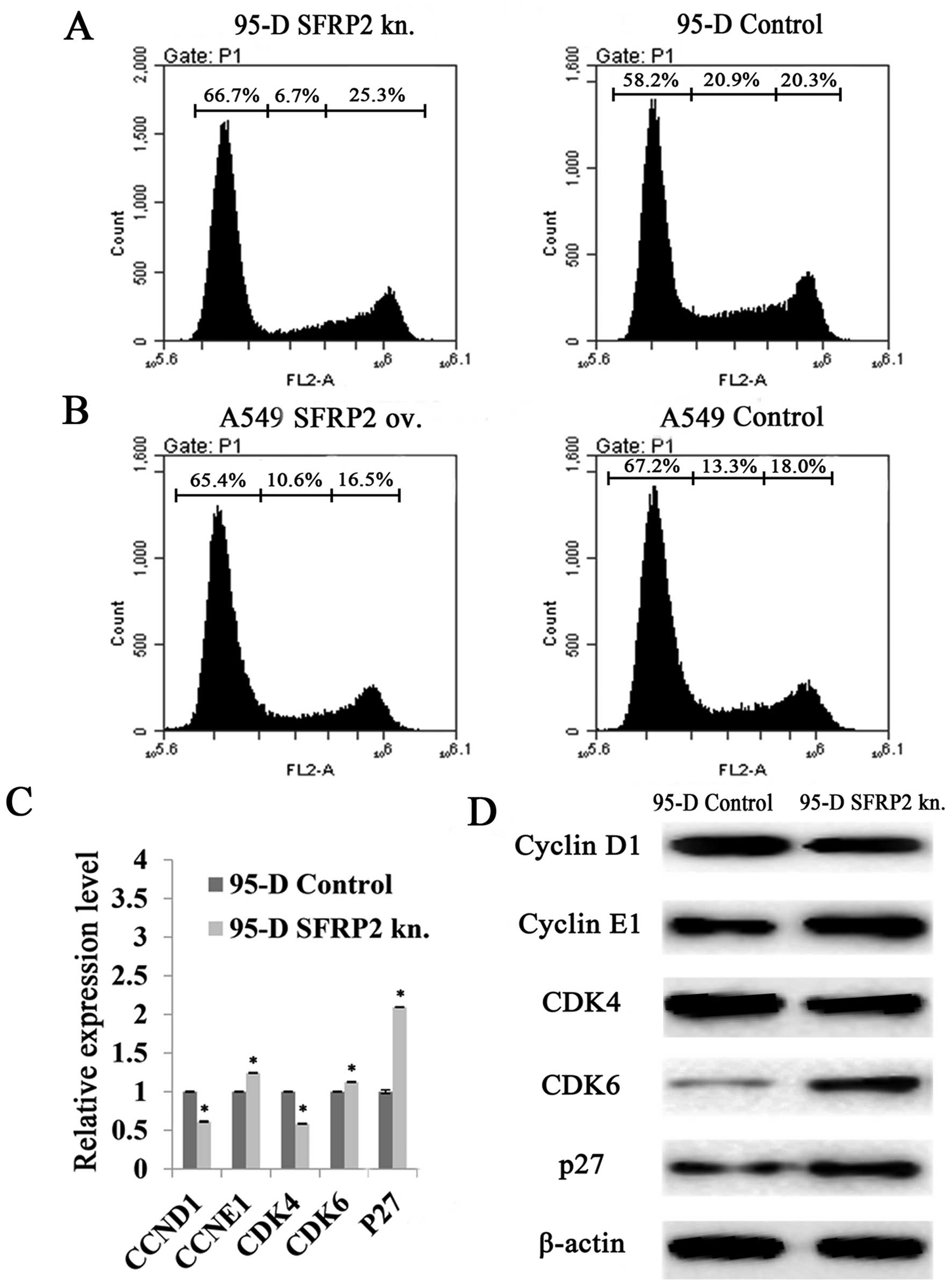
proliferation in lung cancer cells. Sequentially, flow cytometry
analysis demonstrated that the cell cycle was retarded in SFRP2
downregulated 95-D cells. The percentage of cells in G1 phase rose
to 66.7% after SFRP2 knockdown, with an increase of 14.60% compared
to 95-D controls (Fig. 3A), while
no significant change was found in SFRP2 overexpressing A549 cells
(Fig. 3B). In addition, the
molecules related to G1 phase processing were further investigated
by real-time RT-PCR. In SFRP2 downregulated 95-D cells, the mRNA
levels of cyclin E1 (CCNE1), CDK6 and p27 were
significantly increased compared with 95-D control cells
(124.26±0.49 vs. 100.00±0.45%, 112.51±0.53 vs. 100.00±0.49%, and
209.46±0.37 vs. 100.00±2.32%, respectively; all pairs P<0.05)
(Fig. 3C). The mRNA levels of
cyclin D1 (CCND1) and CDK4 were reduced when compared
with 95-D control cells (61.27±0.80 vs. 100.00±0.63% and 58.51±0.45
vs. 100.00±0.51%, respectively; all pairs P<0.05) (Fig. 3C). The protein levels of these G1
phase-related molecules were also evaluated by western blotting. As
shown in Fig. 3D, cyclin D1 and
CDK4 exhibited much weaker bands after SFRP2 knocked down in 95-D
cells, while cyclin E1, CDK6 and p27 with stronger bands (Fig. 3D).
Influences of SFRP2 on lung cancer cell
migration and invasion
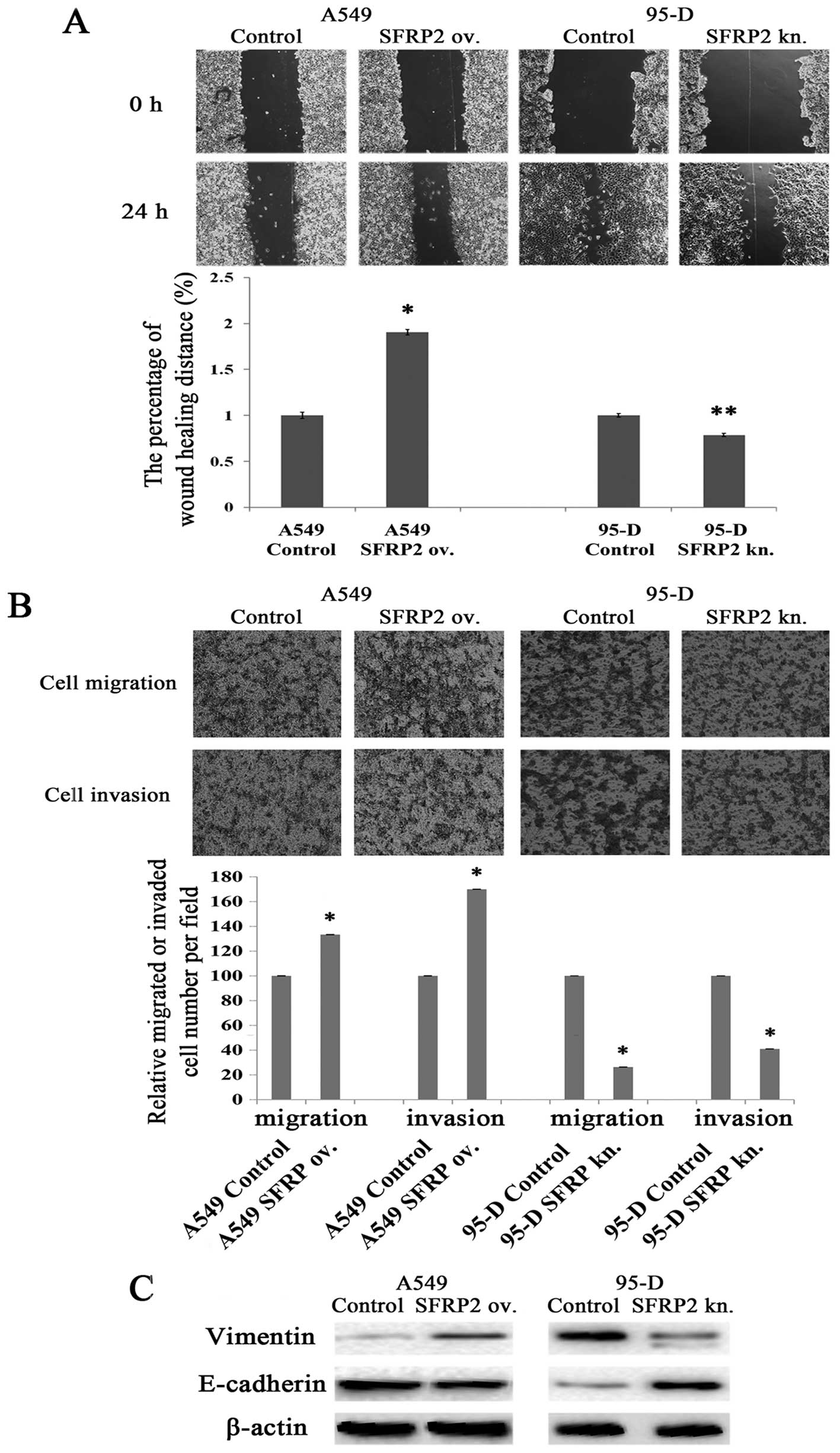
Wound scratch assays and Transwell assays were
performed to evaluate the influences of SFRP2 on lung cancer cell
migration and invasion. As shown in Fig. 4A, the distance changes between 0 and
24 h was calculated by defining the change of control cells as
100%, as presented Fig. 4A. A549
cells migrated faster after SFRP2 overexpression (190.34±3.04 vs.
100.00±3.53%; P<0.05) (Fig. 4A),
which indicates that SFRP2 promotes lung cancer cell migration.
There was also significant difference on migration distance between
95-D cells with and without SFRP2 knockdown (78.62±1.66 vs.
100.00±1.97%; P<0.001) (Fig.
4A). Furthermore, Transwell assays with or without Matrigel
were used to evaluate lung cancer cell properties of invasion and
migration. As shown in Fig. 4B, the
numbers of migrated and invaded A549 cells significantly increased
after SFRP2 overexpression compared with controls (133.33±0.12 vs.
100.00±0.19%, 170.00±0.08 vs. 100.00±0.19%, respectively; both
pairs P<0.05). While the numbers of migrated and invaded 95-D
cells were reduced after the knockdown of SFRP2 by siRNA
transfection as compared to controls (26.32±0.12 vs. 100.00±0.01%
and 40.94±0.10 vs. 100.00±0.08%, respectively; both pairs
P<0.05). Taken together, these findings demonstrated the
involvement of SFRP2 in promoting lung cancer cell migration and
invasion. We next probed the proteins related to the epithelial to
mesenchymal transition (EMT). As shown in Fig. 4C, cytoskeletal protein vimentin
remarkably increased, but E-cadherin significantly decreased in
SFRP2 overexpressing A549 cells (left panel). In the model of 95-D
cells with SFRP2 knockdown, vimentin was weaker, while E-cadherin
increased (Fig. 4C, right
panel).
Modulation of Wnt signaling pathway by
SFRP2 in lung cancer cells
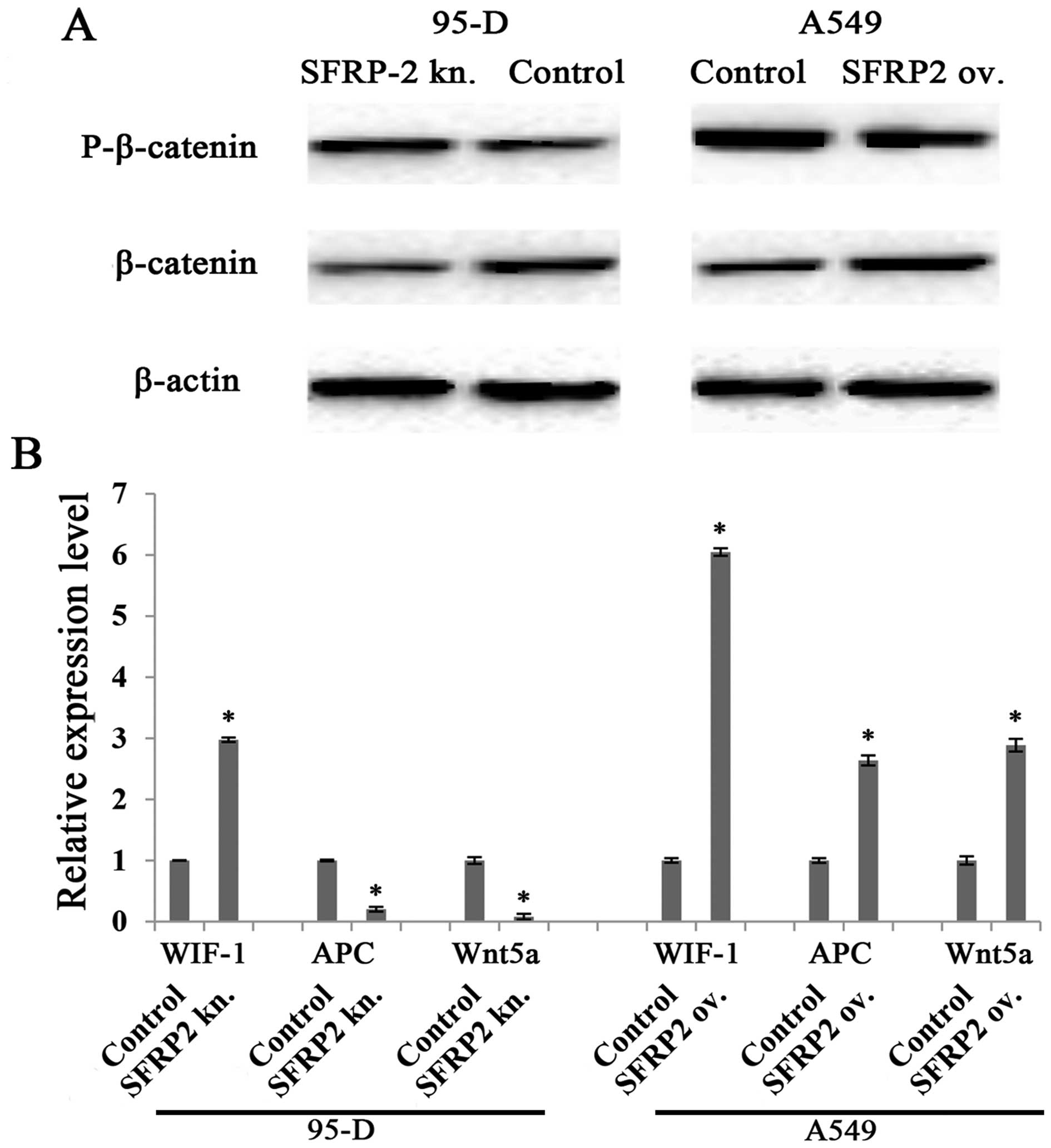
Given that SFRP2 has been reported to be an
antagonist of Wnt signaling pathway, we next investigated the
pathway variation in our cell line models. We found that
phosphorylated β-catenin (degeneration form of β-catenin) was
negatively correlated to SFRP2 level in both 95-D and A549 cell
models, while β-catenin was positively correlated to SFRP2 level.
These observations were supported by the data shown in Fig. 5A, which demonstrated stronger
phosphorylated β-catenin band and much fainter β-catenin band in
SFRP2 downregulated 95-D cell lysates, whereas the opposite data
were revealed in SFRP2 upregulated A549 cells (Fig. 5A, right panel). These findings
implied that SFRP2 serves as an agonist for the canonical Wnt
signaling pathway. The mRNA levels of several mediators of the Wnt
pathway were also checked. In both cell models, WIF-1 increased
significantly, while APC and Wnt5a were specifically reduced in
95-D cells with SFRP2 knockdown (Fig.
5B).
Discussion
There is still disagreement on whether SFRP2
possesses cell proliferation promoting or suppressive function.
Roth et al observed SFRP2 expressing glioma xenografts were
significantly larger than xenografts consisting of control cells
(17). In line with this, Sathi
et al found that SFRP2 promotes cell proliferation in
ameloblastoma (20). In contrast,
the overexpression of SFRP2 in oral squamous cell carcinoma cell
lines suppressed cell proliferation and arrested the cell cycle in
the G1 phase (21). In this study,
we investigated SFRP2 functions in two lung cancer cell models. Our
data with MTT assay showed a significant inhibition of
proliferation at 48h and 72h after SFRP2 knockdown in 95-D cells
(Fig. 2B). In contrast, SFRP2
overexpression in A549 cells promoted the lung cancer cell
proliferation (Fig. 2B). When SFRP2
was downregulated by siRNA in 95-D cells, the cell proliferation
was inhibited at G1 phase of the cell cycle (Fig. 3A) by inducing p27 expression,
accompanied by decreasing CDK4 and cyclin D1 expression on both
mRNA and protein levels (Fig. 3C and
D). It is considered that cyclin D1 binds to its activator CDK4
in order to make cell cycle G1 checkpoint go through. In our SFRP2
knockdown cell model, both CDK4 and cyclin D1 mRNAs and proteins
were reduced profoundly (Fig. 3C and
D). Therefore, our data suggest that SFRP2 is a protein
promoting lung cancer cell proliferation.
The controversy on the tumor functions of SFRP2
still exist. Some researchers considered SFRP2 as a
tumorsuppressive protein based on its highly methylated status in
gene promoter region in various cancer cells, such as breast cancer
(22), oral squamous cell carcinoma
(23) and hepatocellular carcinoma
(24). Others suggested it is an
antagonist to the Wnt pathway as it is homologous to extracellular
portion of Frizzled, the Wnt receptor (12–17).
However, it has been believed that SFRP2 serves as an oncogene to
promote carcinoma development and progression. For example, a
recent report declaimed that SFRP2 inhibition reduced tumor volume
of breast cancer to 46% compared to control mice (18). As we previously discussed, our
findings support the latter hypothesis that SFRP2 is a tumor
promoting protein for lung cancer. Metastasis is an ominous feature
of malignant solid tumors, and EMT, is a key step of the
progression of tumor cell invasion. The speed, site and severity of
tumor metastasis vary in different type of cancers. For example,
breast tumor undergoes EMT at an early stage (25), and it can be detected with the
metastasis to bone, lung or liver for years or decades after the
diagnosis or surgery (26). In case
of lung adenocarcinoma, however, it has the potential of rapidly
spreading to multiple organs, and causes high rate of mortality
(27). The high metastatic feature
of lung cancer calls for investigators to find the molecules
contributing to it. To further investigate the influences of SFRP2
on lung cancer cell invasion, we performed Transwell assays and
western blotting, and found that SFRP2 is critical in lung cancer
invasion for the observation that both the migration and invasion
abilities were positively correlated to the expression level of
SFRP2 (Fig. 4A and B). In addition,
the expression pattern of EMT related protein, such as E-cadherin
and Vimentin, changed according to the variation of cell invasion
potential (Fig. 4C). Interestingly,
β-catenin, one of the EMT related proteins, also changed in line
with alteration of cell invasion ability.
As previously described, in SFRP2 downregulated 95-D
cells, the cell proliferation was arrested at the G1 phase
(Fig. 3A) along with the decreased
cyclin D1 on both mRNA and protein levels (Fig. 3C and D). We also found that the
ability of cell invasion was weakened in the same cell model,
accompanied by reduced protein level of β-catenin (Fig. 5A). Moreover, β-catenin is also the
pivotal molecule in the canonical Wnt signaling pathway, in which,
it could be activated and translocated into the nucleus where it
binds TCF/LEF-1 to enhance the transcription of specific target
genes, such as cyclin D1, c-myc and Axin2 (1). Therefore, these two phenomena were
connected as CCND1 is the downstream target of β-catenin.
These results implied that SFRP2 knockdown downregulated the
canonical Wnt signaling pathway by decreasing β-catenin, and
ultimately reduced the transcriptional targets of β-catenin and
TCF/LEF-1 complex, such as CCND1. Thus, our data suggest
that SFRP2 is an agonist for the Wnt pathway. We also found the
changes of several other molecules related to Wnt pathway, such as
the agonist Wnt5a, and the antagonist WIF-1 and APC (Fig. 5B). However, whether these molecules
function synergistically with SFRP2 is still unknown, and needs to
be further investigated.
In the present study, we examined SFRP2 expression
status in several lung cancer cell lines, and studied its roles in
two lung cancer cell line models containing SFRP2 overexpression
and knockdown, respectively. Our data indicate that SFRP2 is a
cancer promoting protein that promotes lung cancer cell
proliferation and invasion. Moreover, SFRP2 serves as an agonist
for Wnt signaling pathway in lung cancer. These observations are
likely to have implications for our understanding of SFRP2
biological function as a critical molecule underlying lung cancer
development and progression, and the findings suggest that SFRP2
may serve as a potential target for lung cancer therapy.
Acknowledgments
This study was supported by grants from Guangzhou
Medical University (B147048, to J. Tang), the Science and
Information Technology Bureau of Guangzhou (2011Y1-00022 and
[2012]224-8, to J. Tang), and the Science and Information
Technology Bureau of Guangzhou Haizhu District (2012-ZD-02, to J.
Tang).
References
|
1
|
Shi Y, He B, You L and Jablons DM: Roles
of secreted frizzled-related proteins in cancer. Acta Pharmacol
Sin. 28:1499–1504. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Klaus A and Birchmeier W: Wnt signalling
and its impact on development and cancer. Nat Rev Cancer.
8:387–398. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Uematsu K, He B, You L, Xu Z, McCormick F
and Jablons DM: Activation of the Wnt pathway in non small cell
lung cancer: Evidence of dishevelled overexpression. Oncogene.
22:7218–7221. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Clevers H and Batlle E: EphB/EphrinB
receptors and Wnt signaling in colorectal cancer. Cancer Res.
66:2–5. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Weeraratna AT, Jiang Y, Hostetter G,
Rosenblatt K, Duray P, Bittner M and Trent JM: Wnt5a signaling
directly affects cell motility and invasion of metastatic melanoma.
Cancer Cell. 1:279–288. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lu D, Zhao Y, Tawatao R, Cottam HB, Sen M,
Leoni LM, Kipps TJ, Corr M and Carson DA: Activation of the Wnt
signaling pathway in chronic lymphocytic leukemia. Proc Natl Acad
Sci USA. 101:3118–3123. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Takebe N, Harris PJ, Warren RQ and Ivy SP:
Targeting cancer stem cells by inhibiting Wnt, Notch, and Hedgehog
pathways. Nat Rev Clin Oncol. 8:97–106. 2011. View Article : Google Scholar
|
|
8
|
Veeman MT, Axelrod JD and Moon RT: A
second canon. Functions and mechanisms of beta-catenin-independent
Wnt signaling. Dev Cell. 5:367–377. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Clevers H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Arce L, Yokoyama NN and Waterman ML:
Diversity of LEF/TCF action in development and disease. Oncogene.
25:7492–7504. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kawano Y and Kypta R: Secreted antagonists
of the Wnt signalling pathway. J Cell Sci. 116:2627–2634. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Courtwright A, Siamakpour-Reihani S,
Arbiser JL, Banet N, Hilliard E, Fried L, Livasy C, Ketelsen D,
Nepal DB, Perou CM, et al: Secreted frizzle-related protein 2
stimulates angiogenesis via a calcineurin/NFAT signaling pathway.
Cancer Res. 69:4621–4628. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mirotsou M, Zhang Z, Deb A, Zhang L,
Gnecchi M, Noiseux N, Mu H, Pachori A and Dzau V: Secreted frizzled
related protein 2 (Sfrp2) is the key Akt-mesenchymal stem
cell-released paracrine factor mediating myocardial survival and
repair. Proc Natl Acad Sci USA. 104:1643–1648. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee JL, Chang CJ, Wu SY, Sargan DR and Lin
CT: Secreted frizzled-related protein 2 (SFRP2) is highly expressed
in canine mammary gland tumors but not in normal mammary glands.
Breast Cancer Res Treat. 84:139–149. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gehmert S, Sadat S, Song YH, Yan Y and Alt
E: The anti-apoptotic effect of IGF-1 on tissue resident stem cells
is mediated via PI3-kinase dependent secreted frizzled related
protein 2 (Sfrp2) release. Biochem Biophys Res Commun. 371:752–755.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee JL, Chang CJ, Chueh LL and Lin CT:
Secreted frizzled related protein 2 (sFRP2) decreases
susceptibility to UV-induced apoptosis in primary culture of canine
mammary gland tumors by NF-kappaB activation or JNK suppression.
Breast Cancer Res Treat. 100:49–58. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Roth W, Wild-Bode C, Platten M, Grimmel C,
Melkonyan HS, Dichgans J and Weller M: Secreted Frizzled-related
proteins inhibit motility and promote growth of human malignant
glioma cells. Oncogene. 19:4210–4220. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fontenot E, Rossi E, Mumper R, Snyder S,
Siamakpour-Reihani S, Ma P, Hilliard E, Bone B, Ketelsen D, Santos
C, et al: A novel monoclonal antibody to secreted frizzled-related
protein 2 inhibits tumor growth. Mol Cancer Ther. 12:685–695. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tsuruta JK, Klauber-DeMore N, Streeter J,
Samples J, Patterson C, Mumper RJ, Ketelsen D and Dayton P:
Ultrasound molecular imaging of secreted frizzled related protein-2
expression in murine angiosarcoma. PLoS One. 9:e866422014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sathi GA, Inoue M, Harada H, Rodriguez AP,
Tamamura R, Tsujigiwa H, Borkosky SS, Gunduz M and Nagatsuka H:
Secreted frizzled related protein (sFRP)-2 inhibits bone formation
and promotes cell proliferation in ameloblastoma. Oral Oncol.
45:856–860. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xiao C, Wang L, Zhu L, Zhang C and Zhou J:
Secreted frizzled-related protein 2 is epigenetically silenced and
functions as a tumor suppressor in oral squamous cell carcinoma.
Mol Med Rep. 10:2293–2298. 2014.PubMed/NCBI
|
|
22
|
Suzuki H, Toyota M, Carraway H, Gabrielson
E, Ohmura T, Fujikane T, Nishikawa N, Sogabe Y, Nojima M, Sonoda T,
et al: Frequent epigenetic inactivation of Wnt antagonist genes in
breast cancer. Br J Cancer. 98:1147–1156. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sogabe Y, Suzuki H, Toyota M, Ogi K, Imai
T, Nojima M, Sasaki Y, Hiratsuka H and Tokino T: Epigenetic
inactivation of SFRP genes in oral squamous cell carcinoma. Int J
Oncol. 32:1253–1261. 2008.PubMed/NCBI
|
|
24
|
Takagi H, Sasaki S, Suzuki H, Toyota M,
Maruyama R, Nojima M, Yamamoto H, Omata M, Tokino T, Imai K, et al:
Frequent epigenetic inactivation of SFRP genes in hepatocellular
carcinoma. J Gastroenterol. 43:378–389. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hüsemann Y, Geigl JB, Schubert F, Musiani
P, Meyer M, Burghart E, Forni G, Eils R, Fehm T, Riethmüller G, et
al: Systemic spread is an early step in breast cancer. Cancer Cell.
13:58–68. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schmidt-Kittler O, Ragg T, Daskalakis A,
Granzow M, Ahr A, Blankenstein TJ, Kaufmann M, Diebold J, Arnholdt
H, Muller P, et al: From latent disseminated cells to overt
metastasis: Genetic analysis of systemic breast cancer progression.
Proc Natl Acad Sci USA. 100:7737–7742. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hoffman PC, Mauer AM and Vokes EE: Lung
cancer. Lancet. 355:479–485. 2000. View Article : Google Scholar : PubMed/NCBI
|