Introduction
Thyroid cancers are the most common endocrine
malignancies, and represent ~1–1.5% of all tumor-related diseases
(1). The overall 5-year survival of
these tumors is ~85–90%, with the highest mortality rate reported
for undifferentiated histotypes. Thyroid cancers are classified as
papillary (PTC), follicular (FTC), medullary (MTC) and anaplastic
(ATC) carcinomas.
ATC is one of the most aggressive human
malignancies. These tumors are poorly defined, fleshy masses with
areas of necrosis and hemorrhage and there are no features of
thyroid differentiation (2). The
mechanisms underlying the development of ATCs are not completely
understood. Available therapies for ATCs include chemotherapy,
radiotherapy and surgery (3).
Nonetheless, patients with ATC have a median survival of 5 months
and less than 20% of patients survive 1 year post-diagnosis.
Mutations in genes encoding members of the
RAS-MAPK-ERK and PI3K-AKT-mTOR signaling pathways are usually
present in well-differentiated tumor components from which most
ATCs develop (4–6). These signaling pathways are mainly
involved in controlling cell survival, differentiation,
proliferation and metabolism (7).
Moreover, changes in the activity of these pathways can lead to
drug resistance (8), which is a
common feature of ATC. In addition, ATCs are characterized by other
genetic and epigenetic aberrations, which cause deregulation of
genes related to cell cycle regulation and its checkpoints, as well
as alteration of chromosome segregation and spindle structure
(9).
ATCs show frequent upregulation of cyclin-dependent
kinase (CDK) expression, mostly through inactivation of endogenous
cyclin-dependent kinase inhibitors (CDKIs) including
p27KIP1 (10,11). Since CDKs and endogenous CDKIs are
frequently deregulated in cancer cells, these have been considered
as valid drug targets. A large number of synthetic CDKIs have been
tested as anti-proliferative agents in cancer therapy, including
roscovitine, one of the first CDKIs produced (12). Inhibitors of the kinase components
of oncogenic pathways are also being currently explored as
anti-proliferative agents in several human neoplasms, including
thyroid cancer (13).
In the present study, we investigated, in three ATC
cell lines, the effects of a novel roscovitine derivate, named
BP-14, whose efficacy has already been evaluated in hepatocellular
carcinoma (12,14). For this purpose, we evaluated cell
viability, colony-forming capacity and expression of two genes
related to epithelial-mesenchymal transition, CDH1, a well
known marker of epithelial phenotype, and vimentin (VIM), a
type III intermediate filament (IF) protein that is expressed in
mesenchymal cells (15). We,
subsequently, studied the effects of a combined treatment using
BP-14 and the mTOR inhibitor everolimus (RAD-001) on ATC cells,
focusing on cell viability, migration and invasion abilities.
Materials and methods
Cell lines
The human thyroid cancer cell lines derived from
anaplastic thyroid cancer (ATC) used in this study were: FRO
(purchased from the European Collection of Cell Cultures,
Salisbury, UK), SW1736 (obtained from Cell Lines Service GmbH,
Eppelheim, Germany) and 8505C (purchased from Sigma-Aldrich), all
harboring a BRAF V600E mutation (16,17).
These cell lines were tested for being mycoplasma-free and
authenticated by short tandem repeat analysis to be appropriate
cell lines of thyroid cancer origin. FRO cells were grown in
Dulbecco's modified Eagle's medium (DMEM; EuroClone, Milan, Italy),
while SW1736 and 8505C cells were cultivated in Roswell Park
Memorial Institute (RPMI)-1640 medium (EuroClone), in a humidified
incubator (5% CO2 in air at 37°C; Eppendorf AG, Hamburg,
Germany). Both media were supplemented with 10% fetal bovine serum
(Gibco Invitrogen, Milan, Italy), 2 mM L-glutamine (EuroClone) and
50 mg/ml gentamicin (Gibco Invitrogen). Cultured cells were treated
with vehicle (DMSO; Sigma-Aldrich, St. Louis, MO, USA), BP-14
[prepared as described previously (12)] or RAD-001 (everolimus; Novartis,
Basel, Switzerland).
Cell viability
To test cell viability, a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay was performed, as previously described (18). Briefly, 3,000 cells/well were seeded
onto 96-well plates in 200 µl medium. On the following day,
the growth medium was replaced with vehicle-treated medium (NT,
untreated cultures) or with medium containing different doses of
BP-14 (5, 10, 25 and 50 nM) or RAD-001 (25, 50, 100 and 150 nM)
alone or in combination, as indicated in Table I. These dose ranges were selected
based on previous studies (12,19).
The plates were incubated for 0, 24, 48 and 72 h. All experiments
were run in quadruplicate, and cell viability was expressed as a
percentage relative to the vehicle-treated cells. The percentage of
cell viability was used to determine EC50 concentrations
from dose-response curves, after 72 h of treatment.
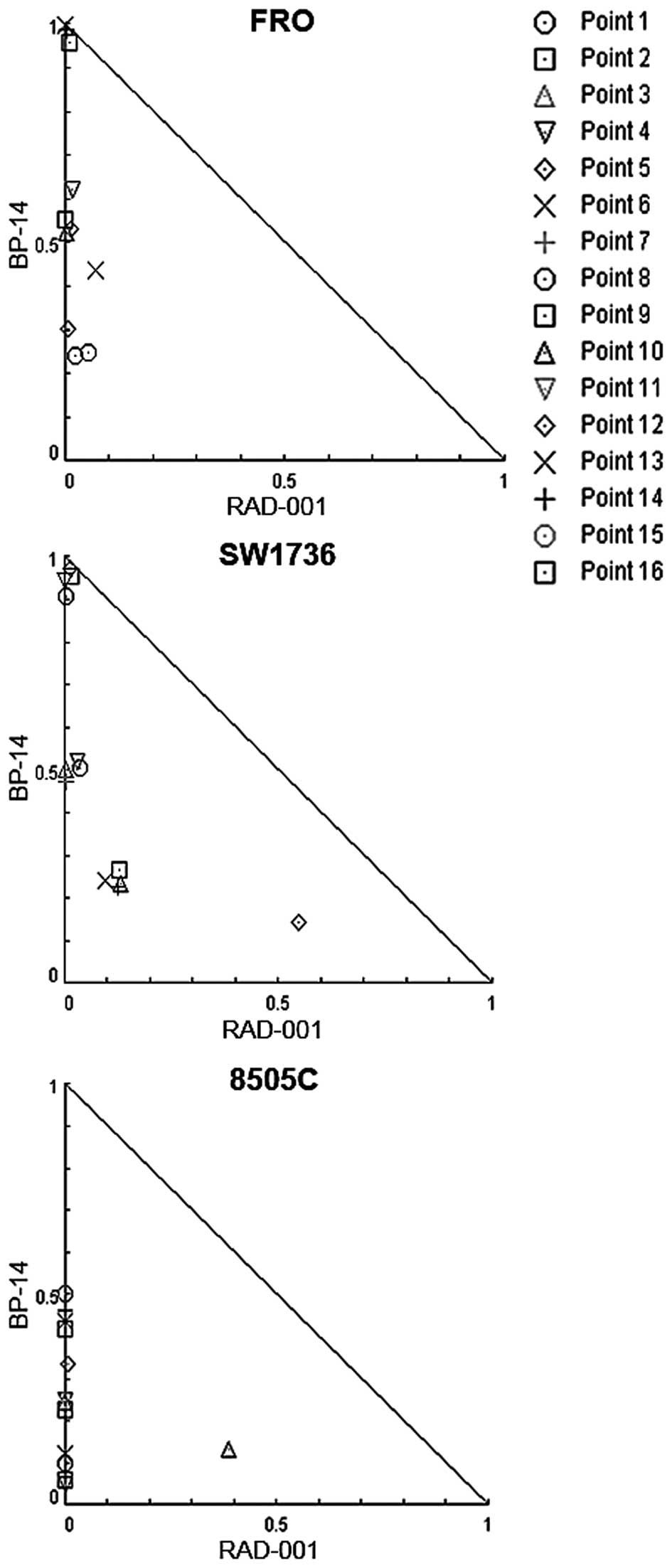
 | Table ICombination index (CI) data for the
combined treatment of BP-14 and RAD-001 in ATC cell lines. |
Table I
Combination index (CI) data for the
combined treatment of BP-14 and RAD-001 in ATC cell lines.
| BP-14 dose (nM) | RAD-001 dose
(nM) | CI in FRO | CI in SW1736 | CI in 8505C |
|---|
| Point 1 | 5.0 | 25.0 | 0.30202 | 1.097E8 | 18596.3 |
| Point 2 | 5.0 | 50.0 | 2.12135 | 0.39369 | 0.05979 |
| Point 3 | 5.0 | 100.0 | 3.73150 | 0.50684 | 0.52049 |
| Point 4 | 5.0 | 150.0 | 5.34166 | 0.94557 | 0.05011 |
| Point 5 | 10.0 | 25.0 | 0.31318 | 0.69159 | 305.585 |
| Point 6 | 10.0 | 50.0 | 0.50569 | 0.33884 | 0.12378 |
| Point 7 | 10.0 | 100.0 | 4.24271 | 0.47682 | 6.17151 |
| Point 8 | 10.0 | 150.0 | 0.26274 | 0.91167 | 0.10012 |
| Point 9 | 25.0 | 25.0 | 0.55719 | 15.6815 | 0.22520 |
| Point 10 | 25.0 | 50.0 | 0.52671 | 0.36697 | 0.24538 |
| Point 11 | 25.0 | 100.0 | 0.63991 | 0.55161 | 0.24814 |
| Point 12 | 25.0 | 150.0 | 0.54472 | 0.98968 | 0.34295 |
| Point 13 | 50.0 | 25.0 | 1.00195 | 3.44322 | 0.43611 |
| Point 14 | 50.0 | 50.0 | 0.98965 | 0.35021 | 0.46149 |
| Point 15 | 50.0 | 100.0 | 1.04564 | 0.54217 | 0.50288 |
| Point 16 | 50.0 | 150.0 | 0.96844 | 0.97197 | 0.41918 |
Colony-formation assay
The clonogenic activity of the ATC cell lines was
evaluated by colony-formation assay. Briefly, the cells were
treated with vehicle, BP-14 and RAD-001, alone or in combination
for 48 h. FRO, SW1736 and 8505C cells were then seeded in 10-cm
plates at a density of 1,000, 500 and 3,000/plate, respectively.
Colonies were stained with 0.1% Coomassie Blue solution
(Sigma-Aldrich) and counted using Gel Doc (Bio-Rad, Hercules, CA,
USA). Data are representative of three independent experiments.
Gene expression assays
Total RNA from the human cell lines, treated either
with vehicle, BP-14 at 25 nM or RAD-001 at 100 nM, alone or in
combination, was extracted with an RNeasy Mini kit according to the
manufacturer's instructions (Qiagen, Hilden, Germany). Total RNA
(500 ng) was reverse transcribed to cDNA using random exaprimers
and SuperScript III reverse transcriptase (Life Technologies,
Carlsbad, CA, USA). Real-time PCRs were performed using Platinum
SYBR Green qPCR SuperMix (Life Technologies) with the ABI Prism
7300 sequence detection systems (Applied Biosystems). The ∆∆CT
method, by means of SDS software (Applied Biosystems), was used to
calculate mRNA levels. Ol igonucleotide pr i mers for CDH1
(for wa rd, 5′-CAAATCGATGTGGATGTTTCCA-3′ and reverse,
5′-CTCGCCCCGTGTGTTAGTTC-3′); VIM (forward,
5′-AGCCTCAGGTCATAAACATCATTG-3′ and reverse,
5′-AGGTTCTTGGCAGCCACACT-3′); and β-actin (forward,
5′-TTGTTACAGGAAGTCCCTTGCC-3′ and reverse,
5′-ATGCTATCACCTCCCCTGTGTG-3′) were purchased from
Sigma-Aldrich.
Combination index (CI) value
Effects of the drug combination used in this study
were evaluated using the combination index (CI) equation based on
the multiple drug-effect equation of Chou and Talalay (20,21).
In all cases where the CI value could be determined, the following
diagnostic rule was applied: CI<1 indicates synergism, CI=1
indicates an additive effect and CI>1 indicates antagonism. The
analysis was carried out using CompuSyn software (ComboSyn Inc.,
Paramus, NJ, USA).
Statistical analysis
Cell viability, colony-forming capacity and
evaluation of mRNA levels were expressed as means ± SD, and
significances were analyzed with the Student's t-test, performed
with GraphPad Software for Science (San Diego, CA, USA).
Results
In a first set of experiments, we evaluated the
biological effects of BP-14, a novel roscovitine derivate, in three
human anaplastic thyroid cancer-derived cell lines (FRO, SW1736 and
8505C), in a time course of treatments with different doses of
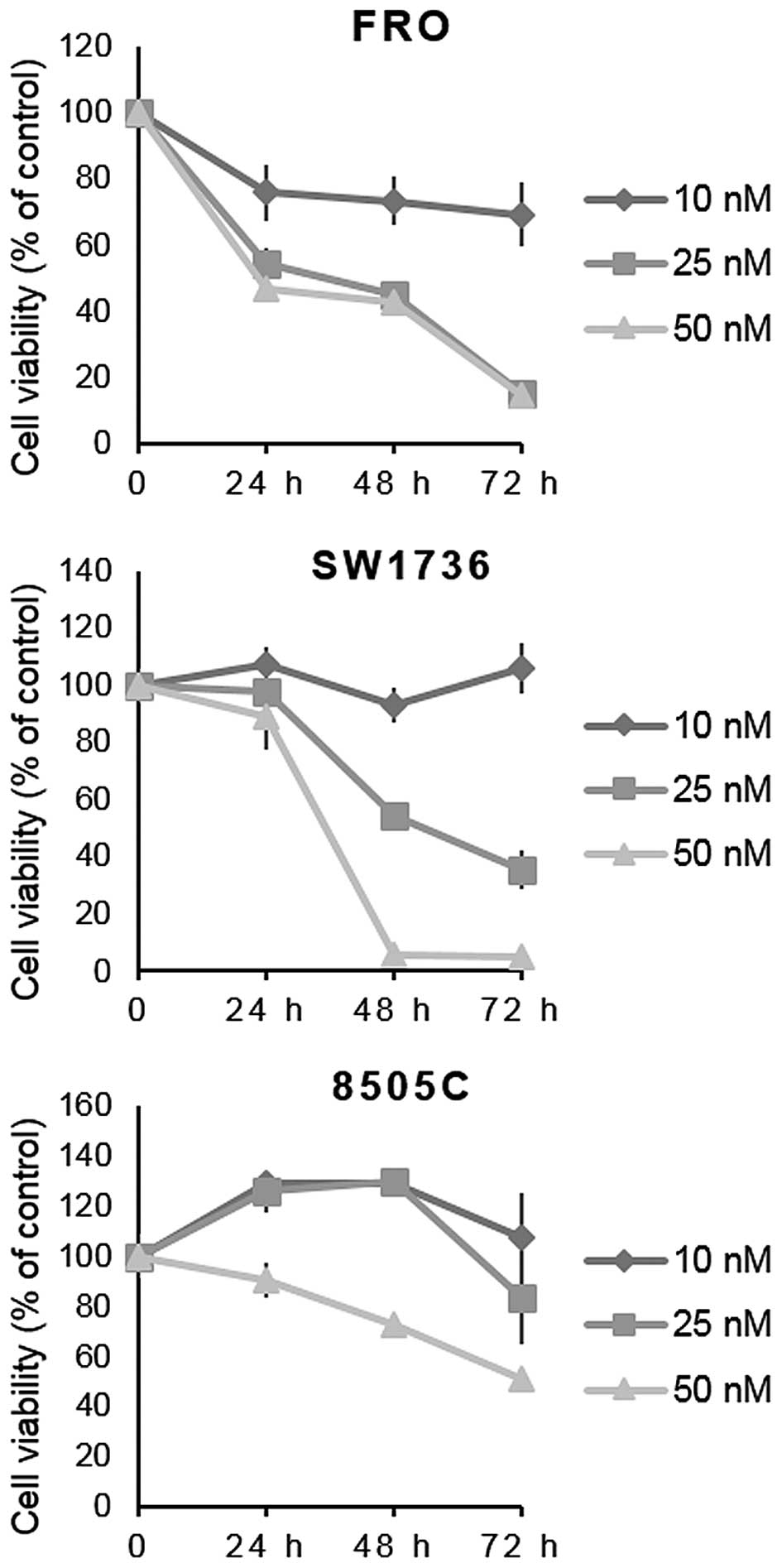
BP-14. As shown in Fig. 1,
incubation with different doses of BP-14 significantly reduced FRO
and SW1736 cell viability, at different time-points, while it
affected 8505C cell viability only at high doses.
Based on the obtained data, in further experiments,
we decided to use the median effective dose of 25 nM for a 48-h
treatment, i.e., dose and time required to achieve 50% of the
theoretical maximal effect in the FRO and SW1736 cell lines.
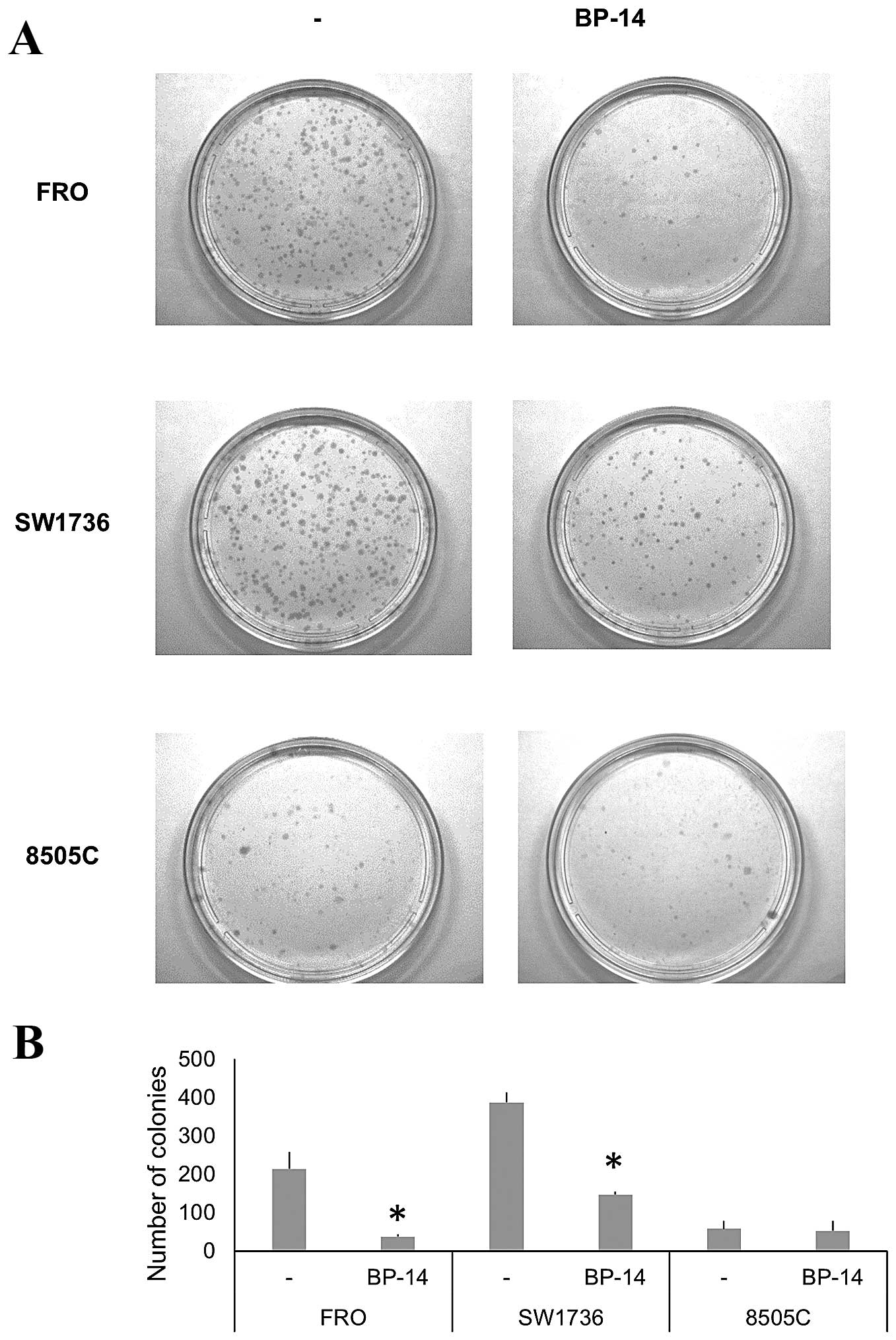
When focusing on clonogenic activity, a 48-h
treatment with BP-14 at 25 nM reduced the number of colonies in the
FRO and SW1736 cells when compared to the number of colonies in the
control cells. In particular, as shown in Fig. 2, we detected a 5.5- and 3-fold
reduction in the FRO and SW1736 cells, respectively. The 8505C cell
line displayed low colony formation efficiency that was not
affected by BP-14.
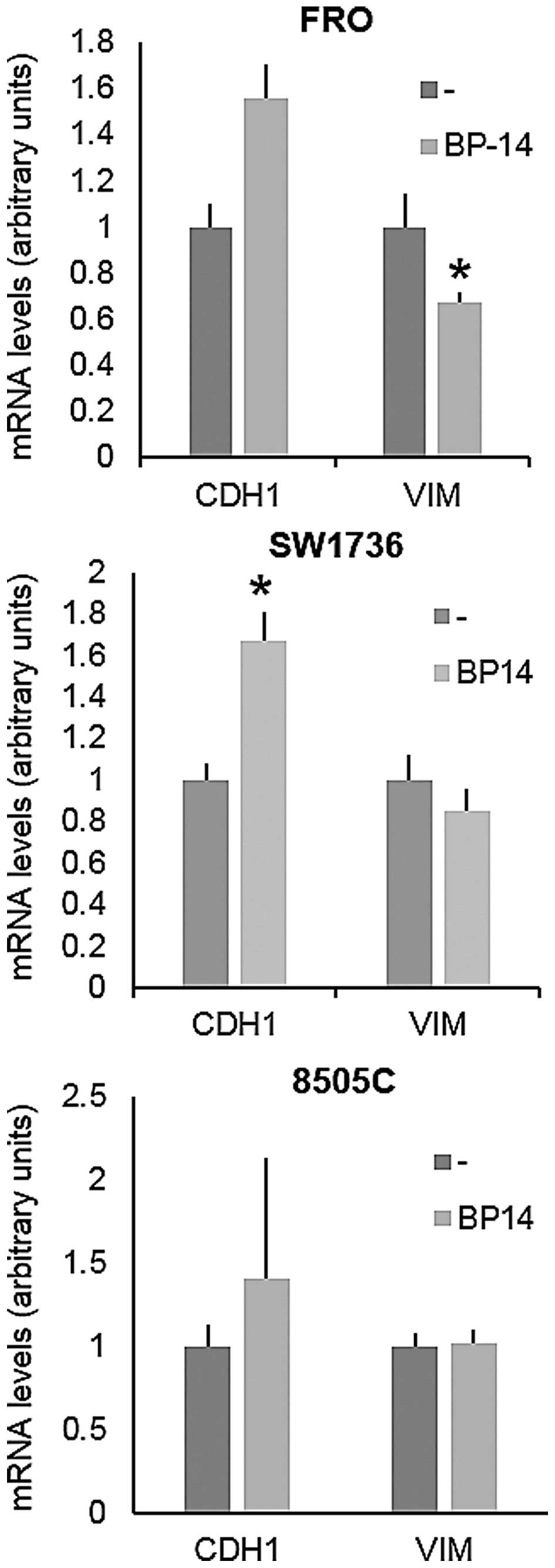
To further investigate the effects of BP-14 on
aggressiveness parameters, we analyzed the expression of various
genes involved in EMT and, in particular, we focused on CDH1
and vimentin (VIM). A significant increase in CDH1
gene expression was noted in the FRO and SW1736 cells (FRO,
P=0.005; SW1736, P=0.002), while 8505C cells showed no variation in
CDH1 expression levels. VIM expression levels were
significantly decreased only in FRO cells (P=0.007, Fig. 3).
Since a promising approach to cancer therapy is the
combined use of different drugs, we evaluated whether BP-14
interacts synergistically with other compounds already used in
cancer treatment, such as everolimus (RAD-001). For this purpose,
we evaluated the effect of a combined treatment of BP14 and RAD-001
by measuring the CI values, according to the Chou and Talalay
equation (20,21). The two-drug combination elicited a
strong decrease in cell viability, compared to the untreated cells
(CI values were <1) (Table I and
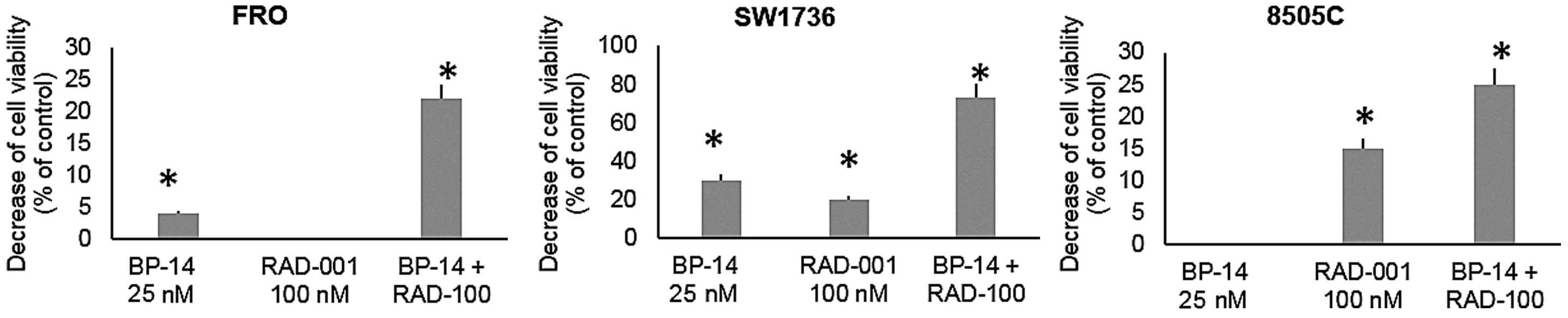
Figs. 4 and 5). Our results revealed that BP-14 and
RAD-001 exhibited a synergistic effect in decreasing cell
proliferation at a quite high range of doses in all three ATC cell
lines.
Then, we evaluated changes in EMT-related processes,
i.e. colony formation as well as CDH1 and VIM gene
expression, after a combined treatment with BP-14 at 25 nM and
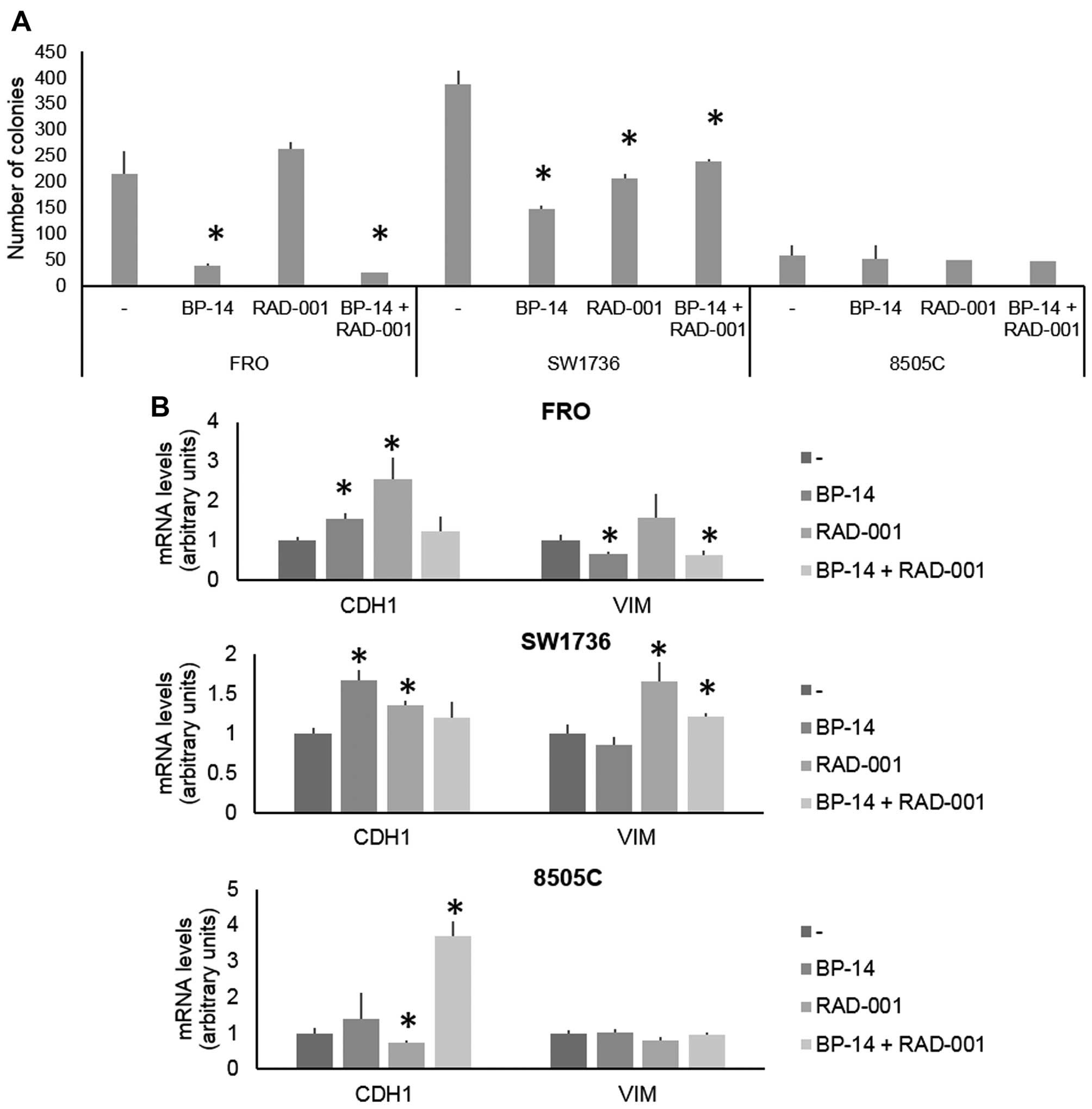
RAD-001 at 100 nM. A 48-h BP-14 treatment significantly reduced
colony formation in the FRO (P=0.0002) and SW1736 (P=0.004) cells
(comparison of Fig. 2 and 6A). The treatment with RAD-001 at 100 nM
reduced the clonogenic ability only in the SW1736 cells (P=0.027).
In contrast, the combined treatment significantly reduced the
clonogenic capacity of the FRO (P=0.0007) and SW1736 (P=0.0002)
cells. The 8505C cells displayed, again, no benefit after either
BP-14 or RAD-001 treatment, alone or in combination.
Following analysis of the expression of EMT-related
genes, treatment with BP-14 at 25 nM increased the CDH1 mRNA
level in the FRO (P=0.005) and SW1736 (P=0.002) cells and reduced
the VIM mRNA level only in the FRO cells (P=0.007)
(comparison of Fig. 3 and 6B). Treatment with RAD-001 at 100 nM
modestly modified CDH1 (FRO, P=0.006; SW1736, P=0.0047;
8505C, P=0.02) and VIM (FRO, P=ns; SW1736, P=0.007; 8505C,
P=ns) mRNA levels in all three cell lines. The synergistic
treatment with BP-14 and RAD-001 greatly increased the CDH1
mRNA level in the 8505C (P=0.0003), so far considered
'unresponsive' to single agent treatment.
Discussion
The search for new target therapies for ATC is
urgently needed. In fact, this neoplasm, although a rare histotype
of thyroid cancer, is characterized by an extremely poor prognosis
(22). The complete loss of
differentiation makes it unresponsive to radioiodine treatment
(23), and current treatments,
based on a combination of surgery, chemotherapy and external
radiotherapy, are not effective. The discovery of molecular
alterations occurring in such tumors, has permitted the selection
of a series of novel agents able to act against such molecular
targets. For an initial screening of these novel potential drugs,
several human ATC cell lines, which carry the same genetic and
epigenetic alterations of human neoplasias, have been largely
exploited.
Among the inhibitors affecting major oncogenic
pathways, the most attractive are those targeting the kinases
involved in cell cycle regulation (24). Since data indicate that CDKs and
CDKIs play a role in thyroid tumorigenesis (25–27),
we focused on the efficacy of a novel compound, BP-14, which is
able to antagonize CDK1/2/5/7 and CDK9 (14). Apart from 'in vitro' effects,
using cultured cell lines (12,14),
BP-14 has been previously investigated 'in vivo' using
xenografts derived from hepatoma cell lines and chemically induced
liver cancer (14). By these
approaches, it has been demonstrated that, at the dose of 1 mg/kg,
BP-14 significantly reduces tumor growth without any side effects.
We showed that BP-14 affected the viability of three different ATC
cell lines at very similar concentrations to those active in
hepatoma cells (14): 25 nM for FRO
and SW1736 cells and 50 nM for 8505C cells. Therefore, considering
data obtained 'in vitro' and 'in vivo' with hepatoma
cells, our findings suggest that BP-14 could be used 'in
vivo' for ATC treatment.
A major finding in our resarch was the demonstration
of synergy between BP-14 and the mTOR inhibitor everolimus. In
fact, in all three ATC cell lines analyzed, the combined use of
BP-14 and everolimus significantly decreased cell viability to a
greater extent than using the two compounds alone. The simultaneous
use of compounds targeting distinct signaling pathways appears to
be a very efficient anticancer strategy (28). Accordingly, previous research has
demonstrated 'in vivo' the synergy between mTOR and MAPK
inhibitors (29). Since the
combination of distinct molecular drugs appears to be quite
effective in ATC (30), our data
would indicate a novel strategy for treatment of this type of
tumor.
Acknowledgments
The present study was supported by grants to G.D.
from Associazione Italiana per la Ricerca sul Cancro (AIRC)
(project no. IG 10296) and to V.K. from the Czech Science
Foundation (grant no. 15-152645).
References
|
1
|
Pellegriti G, Frasca F, Regalbuto C,
Squatrito S and Vigneri R: Worldwide increasing incidence of
thyroid cancer: Update on epidemiology and risk factors. J Cancer
Epidemiol. 2013:9652122013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
O'Neill JP and Shaha AR: Anaplastic
thyroid cancer. Oral Oncol. 49:702–706. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kojic SL, Strugnell SS and Wiseman SM:
Anaplastic thyroid cancer: A comprehensive review of novel therapy.
Expert Rev Anticancer Ther. 11:387–402. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang HM, Huang YW, Huang JS, Wang CH, Kok
VC, Hung CM, Chen HM and Tzen CY: Anaplastic carcinoma of the
thyroid arising more often from follicular carcinoma than papillary
carcinoma. Ann Surg Oncol. 14:3011–3018. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Quiros RM, Ding HG, Gattuso P, Prinz RA
and Xu X: Evidence that one subset of anaplastic thyroid carcinomas
are derived from papillary carcinomas due to BRAF and p53
mutations. Cancer. 103:2261–2268. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu Z, Hou P, Ji M, Guan H, Studeman K,
Jensen K, Vasko V, El-Naggar AK and Xing M: Highly prevalent
genetic alterations in receptor tyrosine kinases and
phosphatidylinositol 3-kinase/akt and mitogen-activated protein
kinase pathways in anaplastic and follicular thyroid cancers. J
Clin Endocrinol Metab. 93:3106–3116. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mendoza MC, Er EE and Blenis J: The
Ras-ERK and PI3K-mTOR pathways: Cross-talk and compensation. Trends
Biochem Sci. 36:320–328. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
McCubrey JA, Steelman LS, Abrams SL, Lee
JT, Chang F, Bertrand FE, Navolanic PM, Terrian DM, Franklin RA,
D'Assoro AB, et al: Roles of the RAF/MEK/ERK and PI3K/PTEN/AKT
pathways in malignant transformation and drug resistance. Adv
Enzyme Regul. 46:249–279. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Russo D, Damante G, Puxeddu E, Durante C
and Filetti S: Epigenetics of thyroid cancer and novel therapeutic
targets. J Mol Endocrinol. 46:R73–R81. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tallini G, Garcia-Rostan G, Herrero A,
Zelterman D, Viale G, Bosari S and Carcangiu ML: Downregulation of
p27KIP1 and Ki67/Mib1 labeling index support the
classification of thyroid carcinoma into prognostically relevant
categories. Am J Surg Pathol. 23:678–685. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pita JM, Figueiredo IF, Moura MM, Leite V
and Cavaco BM: Cell cycle deregulation and TP53 and RAS mutations
are major events in poorly differentiated and undifferentiated
thyroid carcinomas. J Clin Endocrinol Metab. 99:E497–E507.
2014.PubMed/NCBI
|
|
12
|
Gucký T, Jorda R, Zatloukal M, Bazgier V,
Berka K, Řezníčková E, Béres T, Strnad M and Kryštof V: A novel
series of highly potent 2,6,9-trisubstituted purine
cyclin-dependent kinase inhibitors. J Med Chem. 56:6234–6247. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hersey P, Bastholt L, Chiarion-Sileni V,
Cinat G, Dummer R, Eggermont AM, Espinosa E, Hauschild A, Quirt I,
Robert C, et al: Small molecules and targeted therapies in distant
metastatic disease. Ann Oncol. 20(Suppl 6): vi35–vi40. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Haider C, Grubinger M, Řezníčková E, Weiss
TS, Rotheneder H, Miklos W, Berger W, Jorda R, Zatloukal M, Gucky
T, et al: Novel inhibitors of cyclin-dependent kinases combat
hepatocellular carcinoma without inducing chemoresistance. Mol
Cancer Ther. 12:1947–1957. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Polyak K and Weinberg RA: Transitions
between epithelial and mesenchymal states: Acquisition of malignant
and stem cell traits. Nat Rev Cancer. 9:265–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pilli T, Prasad KV, Jayarama S, Pacini F
and Prabhakar BS: Potential utility and limitations of thyroid
cancer cell lines as models for studying thyroid cancer. Thyroid.
19:1333–1342. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schweppe RE, Klopper JP, Korch C,
Pugazhenthi U, Benezra M, Knauf JA, Fagin JA, Marlow LA, Copland
JA, Smallridge RC, et al: Deoxyribonucleic acid profiling analysis
of 40 human thyroid cancer cell lines reveals cross-contamination
resulting in cell line redundancy and misidentification. J Clin
Endocrinol Metab. 93:4331–4341. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Baldan F, Lavarone E, Di Loreto C, Filetti
S, Russo D, Damante G and Puppin C: Histone post-translational
modifications induced by histone deacetylase inhibition in
transcriptional control units of NIS gene. Mol Biol Rep.
41:5257–5265. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Papewalis C, Wuttke M, Schinner S,
Willenberg HS, Baran AM, Scherbaum WA and Schott M: Role of the
novel mTOR inhibitor RAD001 (everolimus) in anaplastic thyroid
cancer. Horm Metab Res. 41:752–756. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chou TC and Talalay P: Analysis of
combined drug effects: A new look at a very old problem. Trends
Pharmacol Sci. 4:450–454. 1983. View Article : Google Scholar
|
|
21
|
Chou TC and Talalay P: Quantitative
analysis of dose-effect relationships: The combined effects of
multiple drugs or enzyme inhibitors. Adv Enzyme Regul. 22:27–55.
1984. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Denaro N, Nigro CL, Russi EG and Merlano
MC: The role of chemotherapy and latest emerging target therapies
in anaplastic thyroid cancer. Onco Targets Ther. 9:1231–1241. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schlumberger M, Lacroix L, Russo D,
Filetti S and Bidart JM: Defects in iodide metabolism in thyroid
cancer and implications for the follow-up and treatment of
patients. Nat Clin Pract Endocrinol Metab. 3:260–269. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Senderowicz AM: Targeting cell cycle and
apoptosis for the treatment of human malignancies. Curr Opin Cell
Biol. 16:670–678. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang S, Wuu J, Savas L, Patwardhan N and
Khan A: The role of cell cycle regulatory proteins, cyclin D1,
cyclin E, and p27 in thyroid carcinogenesis. Hum Pathol.
29:1304–1309. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Elisei R, Shiohara M, Koeffler HP and
Fagin JA: Genetic and epigenetic alterations of the
cyclin-dependent kinase inhibitors p15INK4b and
p16INK4a in human thyroid carcinoma cell lines and
primary thyroid carcinomas. Cancer. 83:2185–2193. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rocha AS, Paternot S, Coulonval K, Dumont
JE, Soares P and Roger PP: Cyclic AMP inhibits the proliferation of
thyroid carcinoma cell lines through regulation of CDK4
phosphorylation. Mol Biol Cell. 19:4814–4825. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li F, Zhao C and Wang L:
Molecular-targeted agents combination therapy for cancer:
Developments and potentials. Int J Cancer. 134:1257–1269. 2014.
View Article : Google Scholar
|
|
29
|
Carracedo A, Baselga J and Pandolfi PP:
Deconstructing feedback-signaling networks to improve anticancer
therapy with mTORC1 inhibitors. Cell Cycle. 7:3805–3809. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Perri F, Lorenzo GD, Scarpati GD and
Buonerba C: Anaplastic thyroid carcinoma: A comprehensive review of
current and future therapeutic options. World J Clin Oncol.
2:150–157. 2011. View Article : Google Scholar : PubMed/NCBI
|