Introduction
Considerable interest has been drawn to the
possibility of preventing or controlling cancer using
phytochemicals since phytochemicals can safely modulate cancer cell
biology and induce cancer cell death. In addition, natural herbs
have become more popular in preventing or controlling cancer as
alternative therapy due to the following reasons. Firstly, the high
intake of fruit and vegetables containing abundant polyphenols is
associated with a low incidence of cancer (1,2).
Secondly, phytochemicals from natural products or herbs show
anticancer effects without noticeable toxicities (3). Thirdly, the number of elderly cancer
patients who cannot tolerate increasingly intense conventional
chemotherapy is increasing (4,5). The
Korean prostrate spurge Euphorbia supina (E. supina)
belongs to the Euphorbiaceae family. It has been used in folk
medicine in Korea against a variety of ailments such as bronchitis,
hemorrhage, jaundice and multiple gastrointestinal diseases
(6). It contains a number of
biologically active substances (7,8). Among
them, polyphenols have attracted our interest due to their
beneficial effects on human health. Literature has shown that
polyphenols reduce the risk of chronic diseases and possess
antioxidant and anti-aging properties (9). It was reported that Korean E.
supine is abundant in polyphenols. Song et al (6) isolated and identified nine polyphenols
from Korean E. supine (PES) which included quercetin and
kaempferol derivatives which have anticancer properties.
Evidence suggests that the major mechanism for the
anticancer effects of polyphenols is apoptosis, a type I programmed
cell death showing a distinctive phenotype: cytoplasmic blebbing,
nuclear condensation and fragmentation, and DNA fragmentation
(10,11). Polyphenol- or phytochemical-induced
apoptosis are mostly caspase-dependent. This caspase-induced
apoptosis is carried out through the intrinsic pathway and/or
extrinsic pathway (12–15).
The intrinsic apoptotic pathway involves
non-receptor-mediated caspase activation through intracellular
signals which increase permeability of the mitochondria and
facilitate release of cytochrome c into the cytoplasm. The
cytochrome c then initiates activation of the caspase
cascade through caspase-9 (16).
The extrinsic apoptotic pathway involves death receptor-mediated
caspase activation. The death receptors such as tumor necrosis
factor receptor 1 (TNFR1) and FAS receptor (Fas)/CD95 are on the
plasma membrane (17,18). Through increase of ligands or their
receptors, the death-inducing signaling complex (DISC) is formed as
ligands bind to these receptors, and then the caspase cascade is
activated through caspase-8 (19).
The Bcl-2 family consists of pro-apoptotic and
anti-apoptotic proteins involved in permeability of the
mitochondrial membrane (16,20).
In addition, the signaling pathways of mitogen-activated protein
kinases (MAPKs) and protein kinase B (PKB or Akt) are involved in
directing cellular responses to a diverse array of stimuli, and
regulate cell survival and apoptosis (21,22).
However, the underlying mechanisms of apoptosis induced by
polyphenols from PES against human leukemic cells have not been
reported yet.
Hence, the aim of the present study was to
investigate the mechanisms involved in the anticancer effects of
PES on U937 human leukemic cells. Our data suggest that PES induced
apoptosis via both the intrinsic and extrinsic apoptotic signaling
pathways; PES upregulated Fas, activated Bid, and suppressed
cFLIPL and Bcl-xL modulation of the Bcl-2 family and the
inhibitor of apoptosis protein (IAP) family proteins, finally
inducing apoptosis through activation of caspases. In addition, the
PES-induced apoptosis was at least in part associated with
extracellular signal-regulated kinase (ERK) activation. This is the
first study demonstrating the anticancer effects of PES against
U937 human leukemic cells.
Materials and methods
Preparation of PES
PES were extracted and purified by Professor S.C.
Shin (6). Briefly, the lyophilized
E. supina tissue (10 g) was extracted in ethyl acetate (300
ml) at 80°C for 20 h, and eluted using a mixture of
methanol:dichloromethane (1:5, 25 ml). The isolated polyphenol
mixtures were identified by HPLC-MS/MS. The nine PES were
identified: gallic acid, protocatechuic acid, nodakenin,
quercetin-3-O-hexoside, quercetin-3-O-pentoside,
kaempferol 3-O-hexoside, kaempferol 3-O-pentoside,
quercetin and kaempferol. Quercetin and kaempferol derivatives
accounted for 84.8% of the total polyphenols.
Cells and reagents
The U937 human leukemic cells obtained from the
American Type Culture Collection (ATCC; Manassas, VA, USA) were
cultured in RPMI-1640 medium (Invitrogen life Technologies.
Carlsbad, CA, USA) supplemented with 10% (v/v) fetal bovine serum
(FBS) (Gibco BRL, Grand Island, NY, USA), 1 mM l-glutamine, 100
U/ml penicillin, and 100 µg/ml streptomycin at 37°C in a
humidified atmosphere of 95% air and 5% CO2. Antibodies
against Bcl-2 (N-19), Bcl-xL, Bax, BAD, Bid, tBid, Fas, Fas ligand
(FasL), Fas-associated protein with death domain (FADD), FLICE-like
inhibitory protein long (c-FLIPL), FLICE-like inhibitory protein
short (c-FLIPS), X-linked inhibitor of apoptosis protein (XIAP),
cellular inhibitor of apoptosis protein-1 (cIAP-1), cIAP-2,
procaspase-3, procaspase-8, procaspase-9, cytochrome c, CoX
IV, Akt 1/2/3 (H-136), ERK and phospho-ERK (E-4) were purchased
from Santa Cruz Biotechnology Inc. (Dallas, TX, USA). Antibodies
against phospho-Akt (Ser473) and phospho-Akt (Thr 308) were
purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA).
Antibody against poly(ADP-ribose) polymerase (PARP) was purchased
from BD Biosciences Pharmingen (San Diego, CA, USA). An antibody
against β-actin was purchased from Sigma (Beverly, MA, USA).
Peroxidase-labeled donkey anti-rabbit and sheep anti-mouse
immunoglobulin, and an enhanced chemiluminescence (ECL) kit were
purchased from Amersham (Waltham, MA, USA). All other chemicals not
specifically cited here were purchased from Sigma-Aldrich (St.
Louis, MO, USA). All these solutions were stored at −20°C.
Cell viability assay
The cell viability was assessed by an MTT assay.
Briefly, U937 cells were treated with PES for 24 h, and then
incubated in 0.5 mg/ml
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (0.5
mg/ml) solution for 3 h at 37°C in the dark. The absorbance of each
well was measured at 540 nm with an enzyme-linked immunosorbent
assay (ELISA) reader (Molecular Devices, LLC, Sunnyvale, CA,
USA).
Nuclear staining
U937 cells treated with PES at the indicated
concentrations for 24 h were harvested, washed with
phosphate-buffered saline (PBS), and fixed with 3.7%
paraformaldehyde in PBS for 10 min at room temperature. After being
washed with PBS, the cells were stained with 2.5 µg/ml
4′,6-diamidino-2-phenylindole (DAPI) solution for 10 min at room
temperature. The cells were washed two more times with PBS, and
observed under a fluorescence microscope.
Agarose gel electrophoresis for DNA
fragmentation test
The cells treated by PES at the indicated
concentrations for 24 h were harvested, and lysed in a buffer
containing 10 mM Tris-HCl (pH 7.4), 150 mM NaCl, 5 mM EDTA, and
0.5% Triton X-100 for 1 h at room temperature. The lysates were
vortexed and centrifuged at 14,000 rpm for 30 min at 4°C. A 25:24:1
(v/v/v) equal volume of neutral phenol:chloroform:isoamyl alcohol
was used for extraction of the DNA from the supernatant.
Electrophoretic analysis was performed on 1.5% agarose gels
containing 0.1 µg/ml ethidium bromide (EtBr).
Flow cytometric analysis for cell cycle
distribution
The cells treated with PES at the indicated
concentrations for 24 h were collected, washed with cold PBS, and
then centrifuged. The pellet was fixed in 75% (v/v) ethanol for 1 h
at 4°C. The cells were washed once with PBS, and resuspended in
cold PI solution (50 µg/ml) containing Rnase A (0.1 mg/ml)
in PBS (pH 7.4) for 30 min in the dark. Flow cytometric analyses
were performed using Beckman Coulter Cytomics FC 500 (Beckman
Coulter, Inc., San Jose, CA, USA). The sub-G1 population was
calculated to estimate the apoptotic cell population.
Measurement of mitochondrial membrane
potential (MMP, ΔΨm)
MMP (ΔΨm) in living U937 cells treated with PES at
the indicated concentrations for 24 h was measured by flow
cytometry with the lipophilic cationic probe JC-1, a ratiometric,
dual-emission fluorescent dye. There are two excitation
wavelengths, 527 nm (green) for the monomer form and 590 nm (red)
for the J-aggregate form. The cells were harvested and resuspended
in 500 µl of PBS and incubated with 10 µM JC-1 for 20
min at 37°C. The green fluorescence reflecting the amount of
damaged mitochondria was quantified by a FACS flow cytometer.
Western blot analysis
The U937 cells treated with PES at the indicated
concentrations for 24 h were harvested and lysed. Their proteins
were quantified using the Bio-Rad protein assay kit (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). We separated the
mitochondrial fraction from whole cell lysates using a mitochondria
isolation kit for cultured cells from Thermo Fisher Scientific
(Waltham, MA USA). The proteins of the extracts were resolved by
electrophoresis, electrotransferred to a polyvinylidene difluoride
membrane from Millipore Corp. (Bedford, MA, USA), and then the
membrane was incubated with the primary antibodies followed by a
conjugated secondary antibody to peroxidase. Blots were developed
under an ECL detection system.
Assay of caspase activity
Caspase activity was measured in PES-treated U937
cells after 24 h by colorimetric assay kits, which utilized the
following synthetic tetra-peptides, labeled with p-nitroaniline
(pNA): Asp-Glu-Val-Asp (DEAD) for caspase-3, Ile-Glu-Thr-Asp (IETD)
for caspase-8 and Leu-Glu-His-Asp (LEHD) for caspase-9. The cells
were lysed in the supplied lysis buffer. The supernatants were
collected and incubated with the supplied reaction buffer
containing dithiothreitol with or without substrates at 37°C. The
caspase activities were determined by absorbance at 405 nm using a
microplate reader.
Statistical analysis
Each experiment was performed in triplicate. The
results are expressed as mean ± standard error (SEM). Significant
differences were determined using the one-way analysis of variance
(ANOVA) with post hoc test Newman-Keuls in the case of at least
three treatment groups and Student's t-test for a two group
comparison. Statistical significance was defined as P<0.05.
Results
PES inhibits cell proliferation, and
induces the apoptosis of U937 human leukemic cells
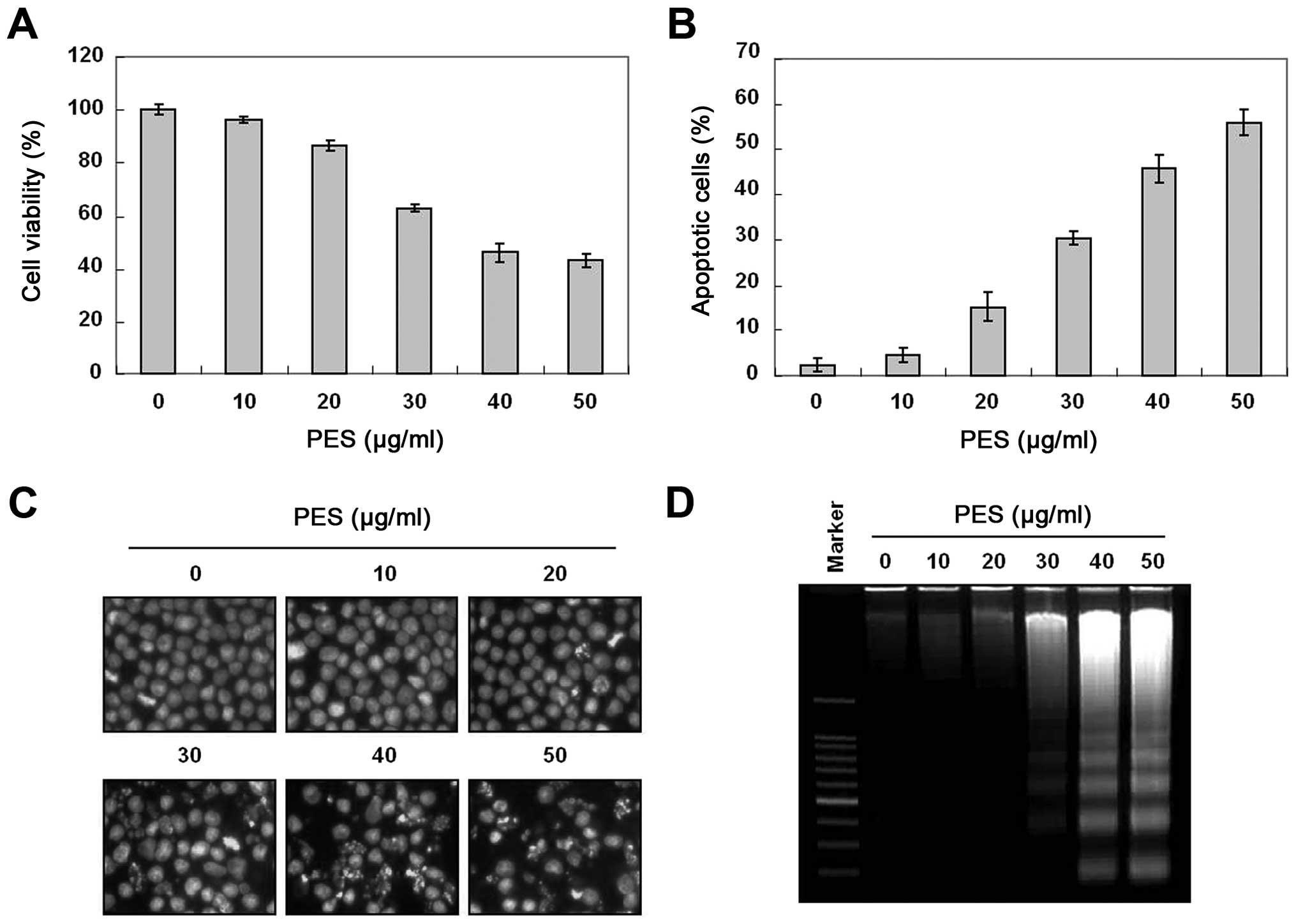
To investigate the anticancer activity of PES, U937
human leukemic cells were treated with the indicated concentrations
(up to 50 µg/ml) of PES for 24 h. The growth of U937 cells
was inhibited by PES treatment in a dose-dependent manner, and the
IC50 value was ~38 µg/ml (Fig. 1A). To investigate further the
mechanism of cell death of the U937 cells, we performed cell cycle
analysis to assess the effects of PES on cell death as well as the
cell cycle. As shown in Fig. 1B,
PES induced significant accumulation of cells with sub-G1 DNA
content (apoptotic cell population) in a dose-dependent manner.
DAPI staining for nuclear morphological changes revealed that PES
induced nuclear condensation and fragmentation in a dose-dependent
manner (Fig. 1C). In addition, DNA
fragmentation test also revealed a typical ladder pattern of DNA
fragmentation which indicates internucleosomal cleavage associated
with apoptosis (Fig. 1D). These
findings suggest that PES induced the apoptosis of U937 the human
leukemic cells.
PES induces caspase activation, and
subsequent cleavage in PARP
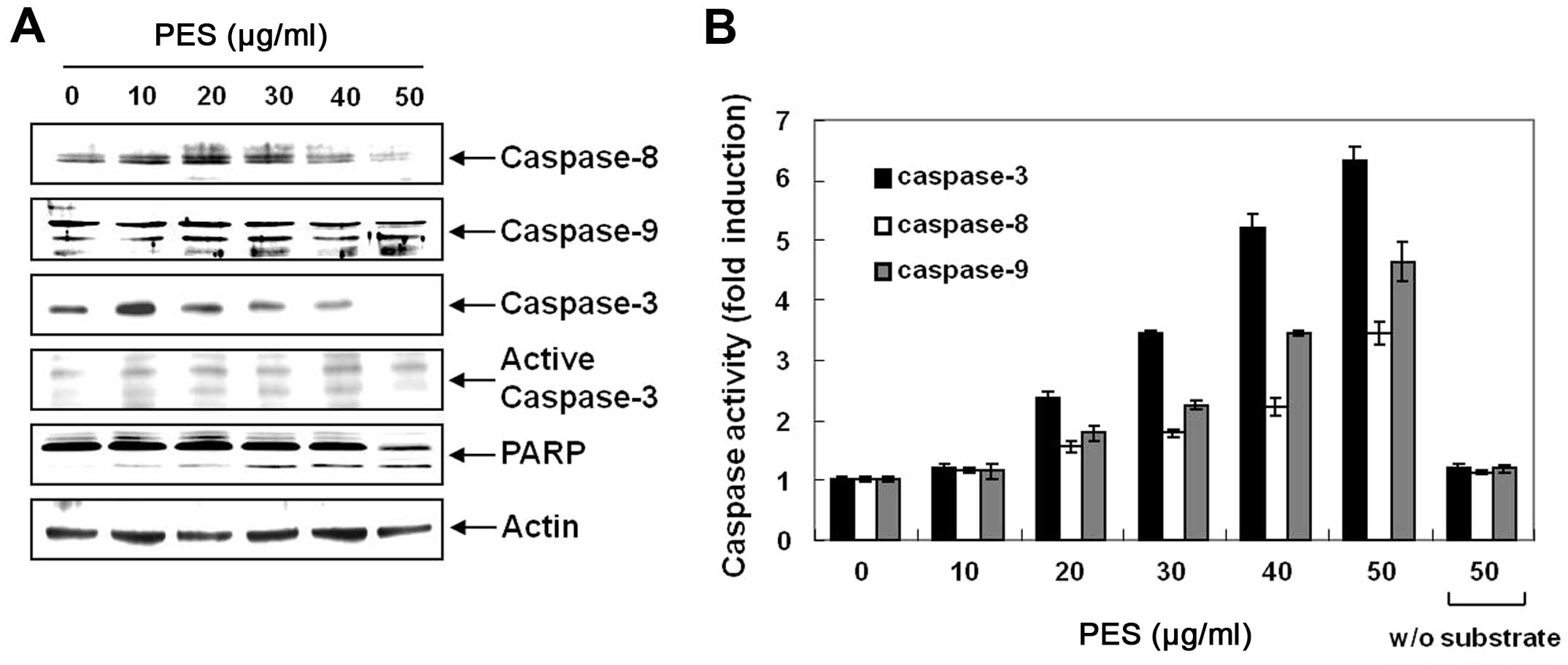
Next, we determined whether PES-induced apoptosis
was caspase-dependent. Western blot analysis revealed that PES
activated procaspase-8, and procaspase-3 as well as procaspase-9,
and cleaved PARP in a dose-dependent manner (Fig. 2A). In addition, caspase activity
assay revealed that PES activated caspase-3, -8 and -9 in a
dose-dependent manner (Fig. 2B). In
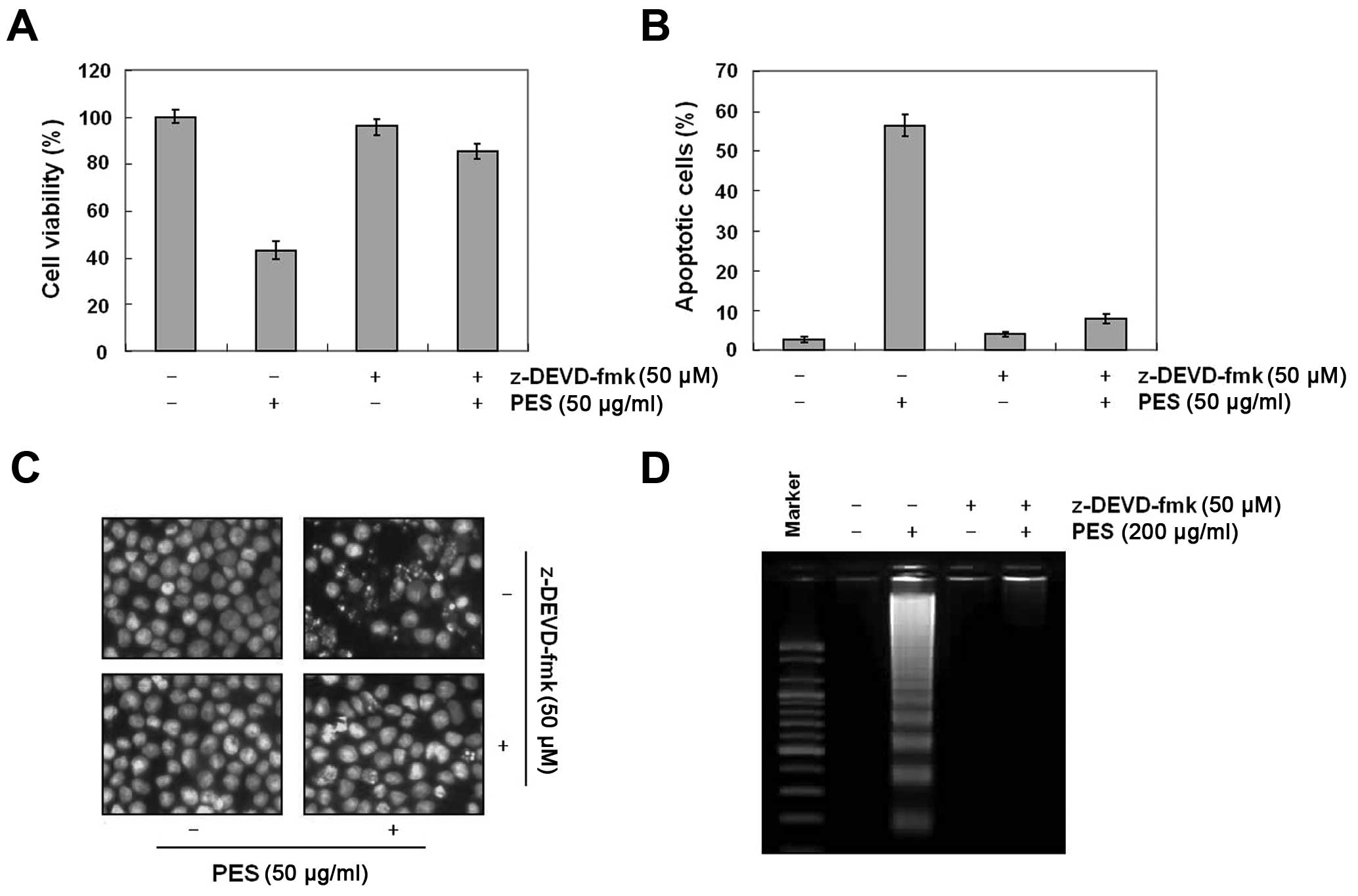
addition, as shown in Fig. 3A–D,
caspase-3 inhibitor (z-DEVD-fmk, 50 µM) suppressed the
PES-induced apoptosis. These findings suggest that PES induced
apoptosis at least in part through the extrinsic pathway of
apoptosis in the U937 cells.
PES induces apoptosis by upregulating
death receptor, Fas and suppressing anti-apoptotic protein, c-FLIP,
and augmented apoptosis by modulating Bcl-2 and IAP family
members
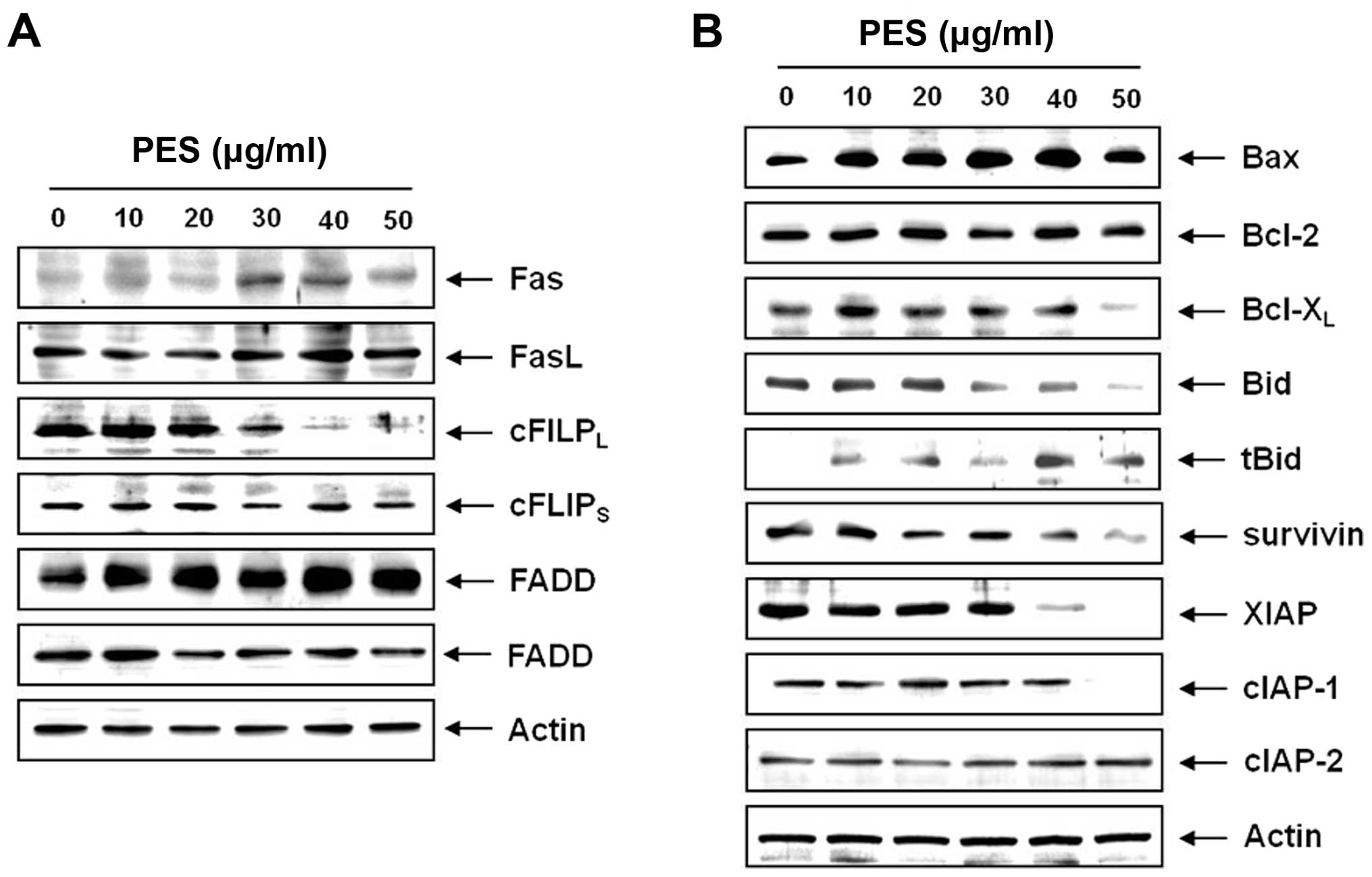
To determine how the extrinsic apoptotic pathway is
involved in the PES-induced apoptosis, we measured the expression
of Fas, FasL, FADD and c-FlIPL. Western blot analysis
revealed that PES upregulated Fas in a dose-dependent manner, and
suppressed c-FLIPL without suppressing c-FLIPS (Fig. 4A). These findings indicate that PES
also induced the death receptor-mediated apoptosis through the
extrinsic pathway. We next assessed the levels of Bcl-2 family
members by western blot analysis, which revealed that PES activated
Bid protein and downregulated anti-apoptotic protein, Bcl-xL
(Fig. 4B). In addition, we tested
the expression of IAP family members which also play an important
role in caspase-dependent apoptosis. Western blot analysis revealed
that PES significantly suppressed survivin, XIAP and cIAP-1 in a
dose-dependent manner (Fig. 4B).
These findings suggest that PES induced apoptosis by upregulating
death receptor Fas and suppressing anti-apoptotic protein cFLIP,
and augmented apoptosis by modulating Bcl-2 and IAP family
members.
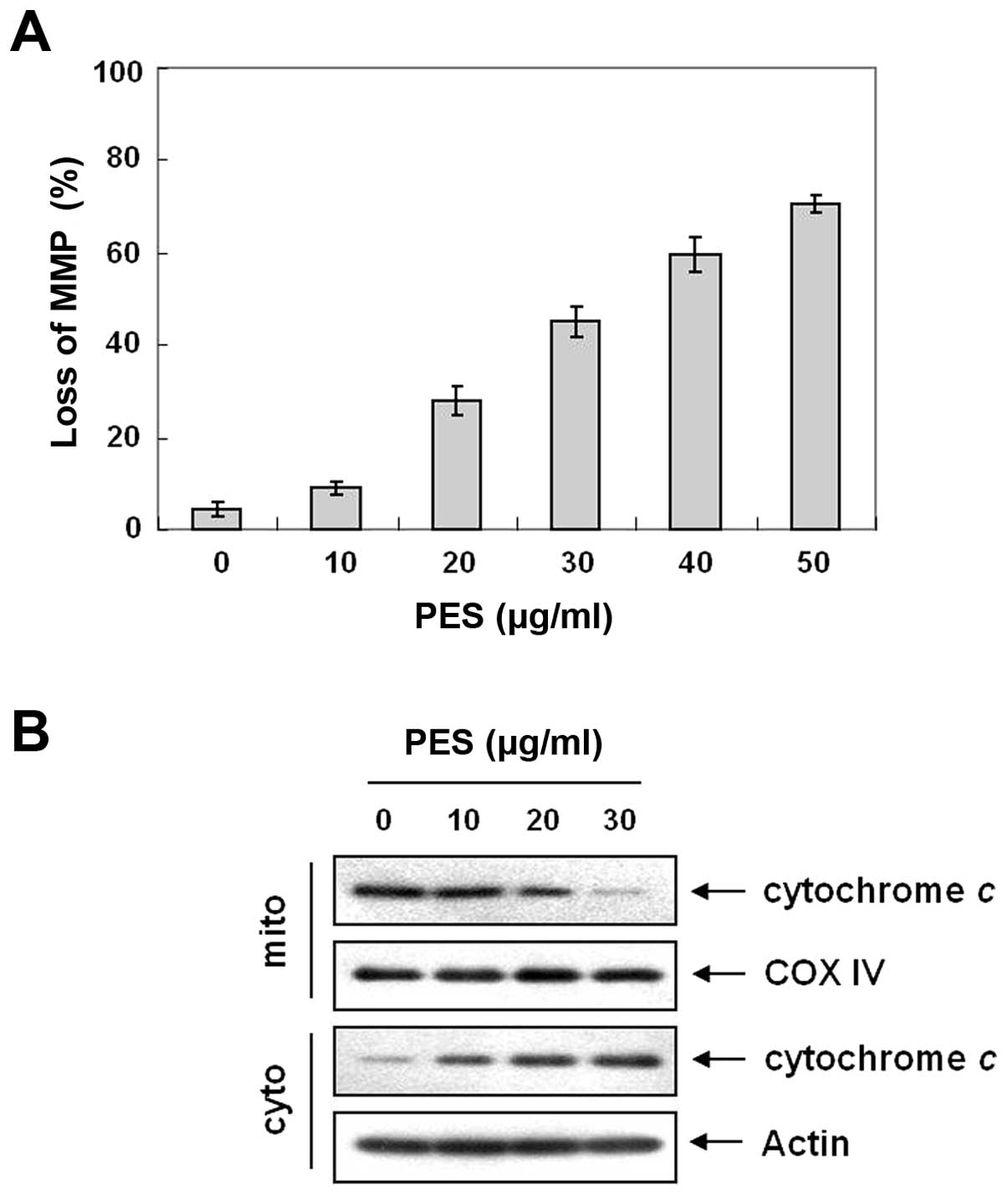
PES induces loss of MMP (ΔΨm) and
cytochrome c release
Bcl-2 family members play an important role in
inducing apoptosis by disrupting the MMP (ΔΨm) in response to
apoptotic signaling (23). We next
measured changes in MMP (ΔΨm) after PES treatment. As shown in
Fig. 5A, PES induced loss of MMP
(ΔΨm) in a dose-dependent manner. The loss of MMP (ΔΨm) causes the
opening of the mitochondrial voltage-dependent anion channel (VDAC)
(24), which results in the release
of cytochrome c, and activation of caspases. Western blot
analysis showed that PES induced cytochrome c release from
mitochondria (Fig. 5B). These
findings suggest that the mitochondrial pathway is also important
in PES-induced apoptosis.
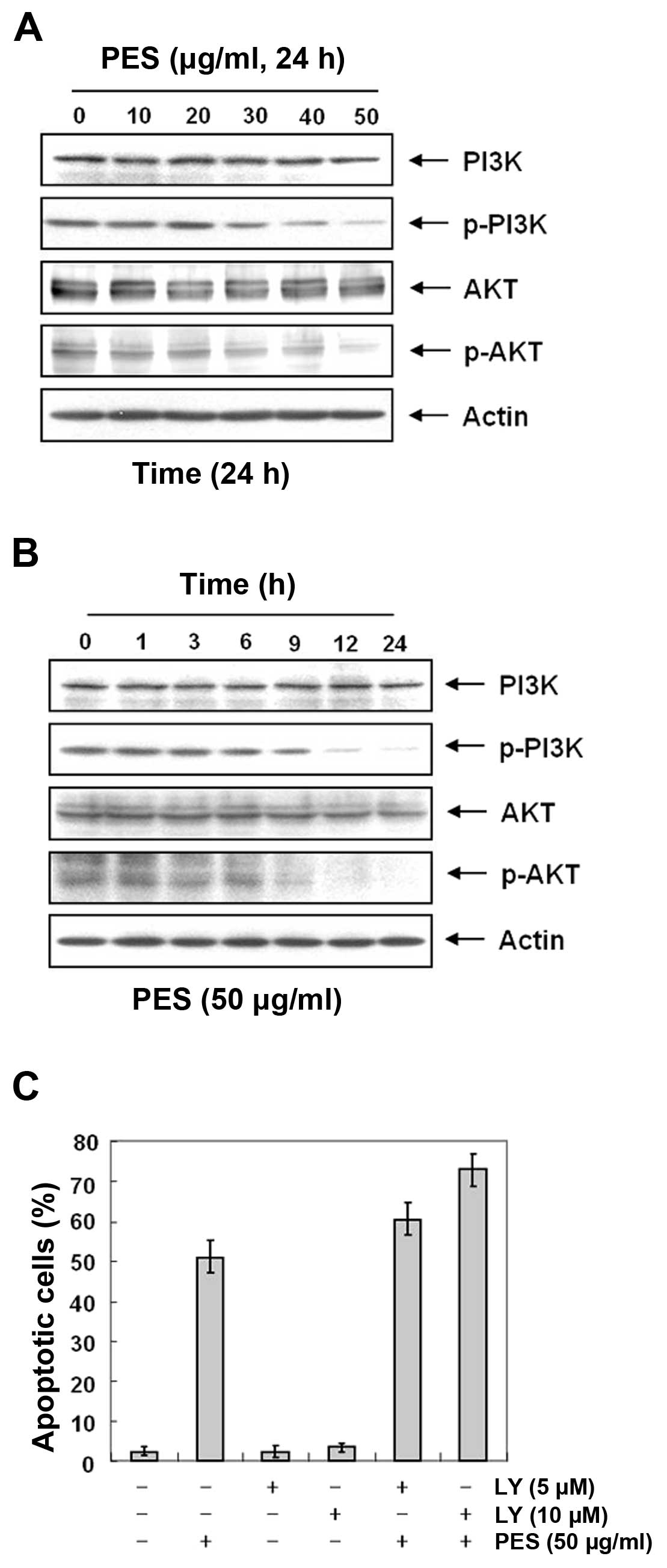
PES induces apoptosis at least in part by
inhibiting the phosphatidylinositol 3-kinase (PI3K)/Akt signaling
pathway
The PI3K/Akt pathway plays an important role in
regulating apoptosis and cell death. In addition, the expression
levels of Fas and cFLIP are regulated by Akt activity (25,26).
Hence, we investigated the effects of PES on Akt activity in the
U937 cells. Since the activity of Akt is regulated by
phosphorylation, we assessed the effects of PES on the levels of
phosphorylated Akt in the PES-treated U937 cells. Western blot
analysis revealed that PES suppressed the phosphorylation of Akt in
a dose- and time-dependent manner (Fig.
6A and B). To confirm this finding, we evaluated the effects of
PES with and without treatment of the the PI3K/Akt inhibitor
(LY294002). Expectedly, flow cytometric analysis for the sub-G1
fraction revealed that LY294002 augmented the effect of PES on
apoptosis (Fig. 6C). These findings
suggest that PES induced apoptosis at least in part by inhibiting
the PI3K/Akt signaling pathway.
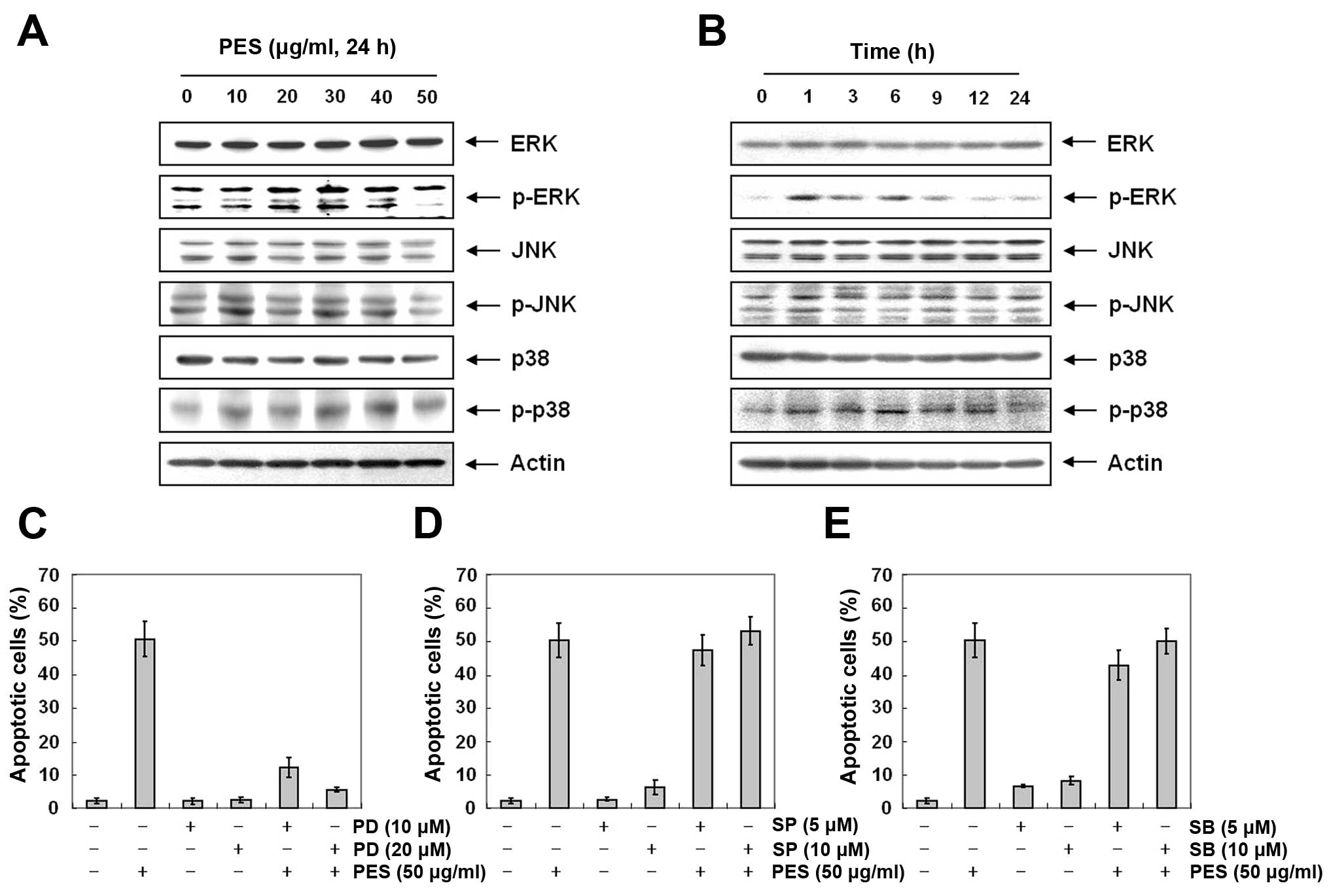
Activation of ERK and MAPK contributes to
PES-induced apoptosis to a greater extent than suppression of
PI3K/Akt signaling
MAPK pathways are also deeply involved in cell
proliferation, differentiation and apoptosis (27). Next, to further investigate the
underlying mechanism responsible for PES-induced apoptosis, we
assessed the changes in phosphorylation of MAPK at various
concentrations of PES for 24 h. Western blot analysis revealed that
PES increased p-ERK and p-p38 MAPK in the U937 cells (Fig. 7A and B). Then we assessed the
effects of MAPK inhibitors [ERK inhibitor (PD98059), JNK inhibitor
(SP600125), and p38 inhibitor (SB203580)] on PES-induced apoptosis
in the U937 cells. As shown in Fig.
7C–E, ERK inhibitor (PD98059) suppressed the PES-induced
apoptosis, while JNK inhibitor (SP600125), and p38 inhibitor
(SB203580) did not. This finding suggests that PES induced
apoptosis at least in part by activation of ERK.
Finally, using DAPI staining (Fig. 8A) and DNA fragmentation test
(Fig. 8B), and PARP cleavage
(Fig. 8C), we also confirmed the
effects of PI3K/Akt and MAPK on PES-induced apoptosis of the U937
cells. This finding suggests that although PES suppressed PI3K/Akt
signaling, the activation of ERK contributed to a greater extent to
PES-induced apoptosis rather than suppression of PI3K/Akt
signaling.
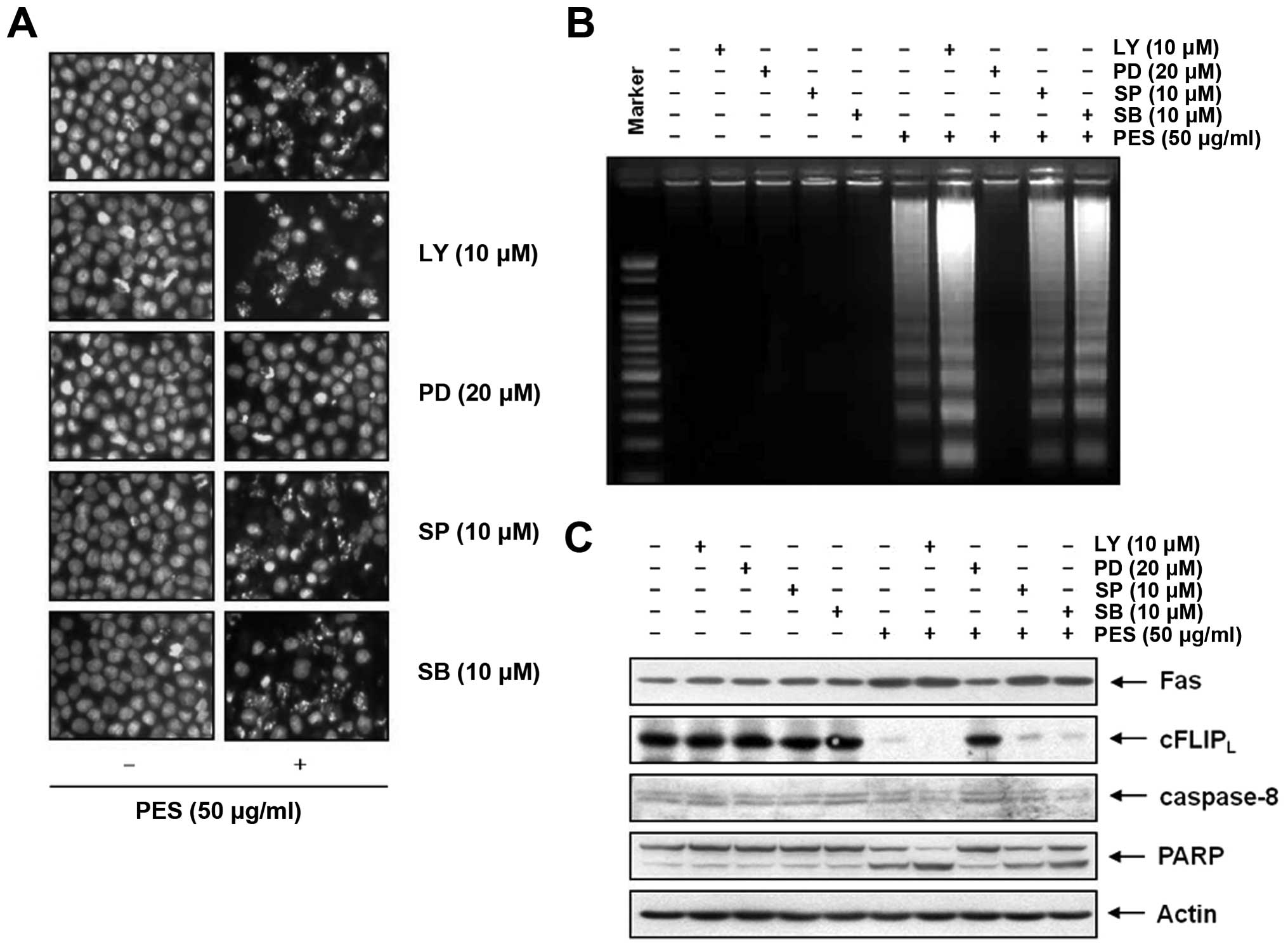 | Figure 8Activation of ERK and MAPK
contributes to PES-induced apoptosis to a greater extent than
suppression of PI3K/Akt signaling. u937 cells were incubated at
indicated concentrations of PES with or without the PI3K/Akt
(Ly294002) inhibitor, the ERK inhibitor (PD98059), the JNK
inhibitor (SP600125), and the p38 inhibitor (SB203580). The results
showed that although PES suppressed PI3K/Akt signaling, the
activation of ERK contributed more to PES-induced apoptosis rather
than PI3K/Akt signaling suppression. (A)
4′,6-Diamidino-2-phenylindole (DAPI) staining, (B) DNA
fragmentation test and (C) western blot analysis. The results are
from one representative of three independent experiments that
showed similar patterns. PES, polyphenols from Korean E.
supina; Ly, Ly294002; PD, PD98059; SP, SP600125; SB,
SB203580. |
Discussion
The present study was conducted to elucidate the
molecular mechanisms of PES-induced apoptosis in U937 human
leukemic cells. The results of this study suggest that PES induced
apoptosis via both the intrinsic and extrinsic pathways. Firstly,
PES strongly inhibited the growth of U937 human leukemic cells in a
dose-dependent manner (Fig. 1A).
Furthermore, PES induced caspase activation and subsequent cleavage
of PARP. The capase-dependent cell death was involved in Fas
upregulation and suppression of c-FLIPS. These two molecules are
closely associated with PI3K/Akt activity (25,26).
In addition, PES suppressed the phosphorylation of PI3K and Akt
which meant suppression of PI3K/Akt activity. In addition, we
demonstrated that the PI3K/Akt inhibitor, LY294002, augmented
PES-induced apoptosis in the U937 cells. These findings suggest
that PES can induce apoptosis at least in part by inhibiting the
PI3K/Akt signaling pathway. MAPK pathways are also deeply involved
in cell proliferation, differentiation and apoptosis (27). Our results showed that PES increased
p-ERK and p-p38 MAPK in the U937 cells. Among MAPKs, the ERK
inhibitor (PD98059) suppressed the PES-induced apoptosis. These
findings also suggest that activation of ERK was involved in
PES-induced apoptosis in the U937 cells. Furthermore, PES-induced
caspase-8 activation led to activation of pro-apoptotic protein Bid
(Fig. 4B). In turn, this activated
tBid translocation from the cytoplasm to the mitochondria, which
increased mitochondrial permeability, and finally released
cytochrome c which then activated the caspase cascade for
apoptosis (28). Consistent with
the above results, loss of MMP was significantly increased in a
dose-dependent manner (Fig. 5A).
Furthermore, PES decreased the expression of Bcl-xL, XIAP, and
cIAP-1 (Fig. 4B) which are direct
inhibitors of activated caspases (29). This finding that PES suppressed
anti-apoptotic factors such as Bcl-xL, XIAP and cIAP-1 suggest that
the intrinsic pathway of apoptosis is also important in PES-induced
apoptosis. These findings are similar to those in our previous
study demonstrating that polyphenols extracted from Allium
cepa induced apoptosis in human leukemia cells U937, through
the extrinsic pathway by upregulating DR5 as well as through the
intrinsic pathway by modulating Bcl-2 and IAP family members
(30). Altogether, our study
suggests that PES induce caspase-dependent apoptosis via both the
intrinsic and extrinsic pathways. Finally, we tested whether
PI3K/Akt and/or MAPK are involved in PES-induced apoptosis. These
results suggest that although PES suppressed PI3K/Akt signaling,
the activation of ERK contributed more to PES-induced apoptosis
rather than suppression of PI3K/Akt signaling.
In conclusion, this study demonstrated the
mechanisms involved in the anticancer effects of PES on U937 human
leukemic cells. PES induced apoptosis via both the intrinsic and
extrinsic apoptotic signaling pathways; PES upregulated Fas,
activated Bid, suppressed cFLIPL, and modulated the
Bcl-2 family and the IAP family proteins, finally inducing
apoptosis through activation of caspases. In addition, the
PES-induced apoptosis was at least in part associated with ERK
activation. Our findings provide the underlying molecular
mechanisms for the anticancer activity of PES that can be useful in
the treatment of leukemia.
Acknowledgments
This study was supported by grants from the National
R&D Program for Cancer Control, Ministry of Health &
Welfare, Republic of Korea (no. 0820050).
References
|
1
|
Lee JE, Männistö S, Spiegelman D, Hunter
DJ, Bernstein L, van den Brandt PA, Buring JE, Cho E, English DR,
Flood A, et al: Intakes of fruit, vegetables, and carotenoids and
renal cell cancer risk: a pooled analysis of 13 prospective
studies. Cancer Epidemiol Biomarkers Prev. 18:1730–1739. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gandini S, Merzenich H, Robertson C and
Boyle P: Meta-analysis of studies on breast cancer risk and diet:
the role of fruit and vegetable consumption and the intake of
associated micronutrients. Eur J Cancer. 36:636–646. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hatcher H, Planalp R, Cho J, Torti FM and
Torti SV: Curcumin: from ancient medicine to current clinical
trials. Cell Mol Life Sci. 65:1631–1652. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jung KW, Won YJ, Kong HJ, Oh CM, Lee DH
and Lee JS: Cancer statistics in Korea: incidence, mortality,
survival, and prevalence in 2011. Cancer Res Treat. 46:109–123.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jung KW, Won YJ, Kong HJ, Oh CM, Lee DH
and Lee JS: Prediction of cancer incidence and mortality in Korea,
2014. Cancer Res Treat. 46:124–130. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Song Y, Jeong SW, Lee WS, Park S, Kim YH,
Kim GS, Lee SJ, Jin JS, Kim CY, Lee JE, et al: Determination of
polyphenol components of Korean prostrate spurge (Euphorbia supina)
by using liquid chromatography-tandem mass spectrometry: overall
contribution to antioxidant activity. J Anal Methods Chem.
2014:4186902014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rice EL: Inhibition of nitrogen-fixing and
nitrifying bacteria by seed plants: VI. Inhibitors from Euphorbia
supina. Physiol Plant. 22:1175–1183. 1969. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cai WH, Matsunami K and Otsuka H:
Supinaionosides A and B: megastigmane glucosides and
supinanitrilosides A–F: hydroxynitrile glucosides from the whole
plants of Euphorbia supina Rafinesque. Chem Pharm Bull (Tokyo).
57:840–845. 2009. View Article : Google Scholar
|
|
9
|
Le Marchand L: Cancer preventive effects
of flavonoids - a review. Biomed Pharmacother. 56:296–301. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Buendia B, Santa-Maria A and Courvalin JC:
Caspase-dependent proteolysis of integral and peripheral proteins
of nuclear membranes and nuclear pore complex proteins during
apoptosis. J Cell Sci. 112:1743–1753. 1999.PubMed/NCBI
|
|
11
|
Kerr JF, Wyllie AH and Currie AR:
Apoptosis: a basic biological phenomenon with wide-ranging
implications in tissue kinetics. Br J Cancer. 26:239–257. 1972.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Danial NN and Korsmeyer SJ: Cell death:
critical control points. Cell. 116:205–219. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Armstrong JS: Mitochondrial membrane
permeabilization: the sine qua non for cell death. BioEssays.
28:253–260. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lemasters JJ, Qian T, He L, Kim JS, Elmore
SP, Cascio WE and Brenner DA: Role of mitochondrial inner membrane
permeabilization in necrotic cell death, apoptosis, and autophagy.
Antioxid Redox Signal. 4:769–781. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Boya P, Cohen I, Zamzami N, Vieira HL and
Kroemer G: Endoplasmic reticulum stress-induced cell death requires
mitochondrial membrane permeabilization. Cell Death Differ.
9:465–467. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li P, Nijhawan D, Budihardjo I,
Srinivasula SM, Ahmad M, Alnemri ES and Wang X: Cytochrome c and
dATP-dependent formation of Apaf-1/caspase-9 complex initiates an
apoptotic protease cascade. Cell. 91:479–489. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wajant H: The Fas signaling pathway: more
than a paradigm. Science. 296:1635–1636. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen G and Goeddel DV: TNF-R1 signaling: a
beautiful pathway. Science. 296:1634–1635. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Walczak H and Krammer PH: The CD95
(APO-1/Fas) and the TRAIl (APO-2l) apoptosis systems. Exp Cell Res.
256:58–66. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Murphy KM, Ranganathan V, Farnsworth ML,
Kavallaris M and Lock RB: Bcl-2 inhibits Bax translocation from
cytosol to mitochondria during drug-induced apoptosis of human
tumor cells. Cell Death Differ. 7:102–111. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mitsiades CS, Mitsiades N and Koutsilieris
M: The Akt pathway: molecular targets for anti-cancer drug
development. Curr Cancer Drug Targets. 4:235–256. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pearson G, Robinson F, Beers gibson T, Xu
BE, Karandikar M, Berman K and Cobb MH: Mitogen-activated protein
(MAP) kinase pathways: regulation and physiological functions.
Endocr Rev. 22:153–183. 2001.PubMed/NCBI
|
|
23
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Timmer JC and Salvesen GS: Caspase
substrates. Cell Death Differ. 14:66–72. 2007. View Article : Google Scholar
|
|
25
|
Maddigan A, Truitt L, Arsenault R,
Freywald T, Allonby O, Dean J, Narendran A, Xiang J, Weng A, Napper
S, et al: EphB receptors trigger Akt activation and suppress Fas
receptor-induced apoptosis in malignant T lymphocytes. J Immunol.
187:5983–5994. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang X, Chen W, Zeng W, Bai L, Tesfaigzi
Y, Belinsky SA and Lin Y: Akt-mediated eminent expression of c-FlIP
and Mcl-1 confers acquired resistance to TRAIL-induced cytotoxicity
to lung cancer cells. Mol Cancer Ther. 7:1156–1163. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Robinson MJ and Cobb MH: Mitogen-activated
protein kinase pathways. Curr Opin Cell Biol. 9:180–186. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kelekar A and Thompson CB: Bcl-2-family
proteins: the role of the BH3 domain in apoptosis. Trends Cell
Biol. 8:324–330. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hunter AM, LaCasse EC and Korneluk RG: The
inhibitors of apoptosis (IAPs) as cancer targets. Apoptosis.
12:1543–1568. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Han MH, Lee WS, Jung JH, Jeong JH, Park C,
Kim HJ, Kim G, Jung JM, Kwon TK, Kim GY, et al: Polyphenols
isolated from Allium cepa L. induces apoptosis by suppressing IAP-1
through inhibiting PI3K/Akt signaling pathways in human leukemic
cells. Food Chem Toxicol. 62:382–389. 2013. View Article : Google Scholar : PubMed/NCBI
|