Introduction
Hypoxic microenvironments exist in various malignant
carcinomas, leading to angiogenesis, tumourigenesis, and metastasis
of neoplasms. One of the mechanisms is that hypoxia can induce
epithelial-mesenchymal transition (EMT), a crucial step in
embryological development and cancer progression. In EMT, the
epithelial cells lose cell-to-cell tight junctions and baso-apical
polarity, adopt mesenchymal cell phenotypes and plasticity, and
become mobile (1,2). Loss of the epithelial markers,
E-cadherin and β-catenin, and acquisition of the mesenchymal
markers, vimentin, N-cadherin, and fibronectin, are the molecular
hallmarks of EMT.
Hypoxia-induced EMT was found in a number of
carcinomas, including colon, breast, pancreatic, and ovarian
carcinomas, and hepatoblastoma (3).
Hypoxia inducible factors (HIFs) regulate hypoxic responses,
including malignant biological behaviour in tumour cells. HIF-1 is
the most important factor, and is composed of two subunits, HIF-1α
and HIF-1β. The HIF-1α subunit is regulated by oxygen tension,
whereas HIF-1β is constitutively expressed. HIF-1 targets the
hypoxia-responsive genes (HRE), including the transcriptional
regulators (Snail, Slug, Twist, ZEB) of E-cadherin (4). It has been proposed that long-term
hypoxia drives a perpetual EMT through upregulation of the zinc
finger, E-box binding, homeobox protein, ZEB2, whereas short-term
hypoxia induces a reversible EMT that requires Twist1 (5). Hypoxia also is involved in
EMT-triggering pathways of TGF-β, Notch, NFκB, Wnt, and PI3K
(6–10). However, the underlying mechanisms of
how these multigene interactive cascades are triggered are still
unknown.
Signal transducer and activator of transcription 3
(STAT3), a member of the STAT family of transcription factors that
is activated through various mitogenic signalling pathways of cell
growth, migration, and angiogenesis, is overexpressed and
constitutively activated in a variety of human malignancies
(11). Studies have demonstrated
that STAT3 is required for EMT. TGF-β induced EMT in pancreatic
cancer requires STAT3 activity and Smad4-dependent signalling
regulates this process (12).
Activation of EGFR or IL-6R results in high levels of STAT3 and
activation of the downstream JAK2/STAT3 pathway, inducing a
fibroblast-like morphology and migratory phenotype, consistent with
the upregulation of mesenchyme-associated N-cadherin and vimentin,
and the nuclear translocation of β-catenin (13). EGF promotes EMT of cancer cells
through a ROS/STAT3/HIF-1α/TWIST1/N-cadherin signalling cascade
(14). STAT3 directly regulates the
expression of genes encoding the transcription factors TWIST, ZEB1,
and Snail (15–17). STAT3 and associated EMT may
contribute to chemoresistance, by involving EGFR signalling
(18,19).
Overexpression of HIF-1α and STAT3 has been detected
in numerous cancers, and these proteins play important roles in
cancer progression and migration. Activated STAT3 increases HIF-1α
protein levels by enhancing HIF-1α stability and accelerating its
de novo synthesis (20).
STAT3 activates HIF1 target genes by binding to their promoters,
interacting with HIF1α, and recruiting the coactivators, CREB
binding protein (CBP) and p300, and RNA polymerase II (Pol II) to
form enhanceosome complexes (21).
Stat3 regulates expression of Akt, which is required for growth
signal-induced HIF-1 upregulation (22). However, the role of STAT3 in
hypoxia-induced EMT has not been reported.
Oesophageal carcinoma is the fourth most prevalent
malignancy in China and the sixth cause of cancer-related death in
the world. Oesophageal squamous cell carcinoma (ESCC) is one of the
major histopathological subtypes of oesophageal cancer. Metastasis
and chemo/radio resistance of ESCC are closely related to hypoxic
conditions and EMT. This study investigated the mechanisms of
hypoxia-induced EMT in ESCC and the roles of STAT3 in this
process.
Materials and methods
Cell culture
Human ESCC cell lines TE-1 and EC-1 were purchased
from the Shanghai Institute of Biochemistry and Cell Biology
(Shanghai, China). The cells were grown in RPMI-1640 supplemented
with 10% foetal bovine serum (FBS) and 1% penicillin/streptomycin.
The cells were incubated under the normoxic condition of 20%
O2, or in the presence of cobalt chloride
(CoCl2; Sigma, St. Louis, USA), a known hypoxia-inducing
agent (23).
Reverse transcription-PCR (RT-PCR) and
real-time PCR (qPCR)
Total RNA Extraction, RT-PCR, and qPCR were
performed as previously described (3). Total cellular RNA was extracted with
the Ultrapure RNA kit (CW Biotech Co., Beijing, China).
First-strand cDNA was synthesized with oligo (dT) primers (Sangon
Biotech. Shanghai, China) and the Super RT cDNA kit (CW Biotech
Co.). The primer sequences were as follows: E-cadherin forward:
5′-GAGAACGCATTGCCACATACAC-3′, reverse:
5′-GAGCACCTTCCATGACAGACCC-3′; vimentin forward:
5′-ATGTGGATGTTTCCAAGCCTGAC-3′, reverse:
5′-GAGTGGGTATCAACCAGAGGGAGT-3′; HIF-1α forward:
5′-CACTGCACAGGCCACATTCACGT-3′, reverse:
5′-GAGCACCTTCCATGACAGACCC-3′; STAT3 forward:
5′-GAGAACGCATTGCCACATACAC-3′, reverse: 5′-TCTGGCCGACAATACTTTCC-3′;
and β-actin forward: 5′-GTCCACCGCAAATGCTTCTA-3′, reverse:
5′-TGCTGTCACCTTCACCGTTC-3′. Products from RT-PCR were monitored by
polyacrylamide gel electrophoresis (PAGE). qPCR was performed using
the UltraSYBR Mixture (CW Biotech Co.) and ABI 7300 Real-time PCR
system. To normalize the amount of input RNA, PCR was performed
with probe and primers for β-actin.
Immunoblotting
Cells were harvested and incubated with RIPA buffer
for 10 min to collect the cell lysates. The total cellular protein
was separated using 10% SDS-PAGE and transferred to PVDF membranes
(Millipore Corp., Bedford, MA, USA). PVDF membranes were blocked
with TRIS buffered saline (TBS) containing 5% non-fat milk, and
then incubated with the primary antibody at 4°C overnight, followed
by incubation with goat anti-rabbit/mouse secondary antibodies
(1:10000, Odessey, China). The primary antibodies included
anti-HIF-1α (1:1000, Abcam), anti-E-cadherin (1:2000, Abcam),
anti-β-actin (1:10000, Sigma), anti-STAT3 (1:2000, Santa Cruz
Biotechnology, Santa Cruz, CA, USA), anti-p-STAT3 (Tyr705) (1:2000,
Santa Cruz Biotechnology), and anti-vimentin (1:2000, Abcam),
anti-GAPDH (1:1000, Santa Cruz Biotechnology). Protein-antibody
complexes were visualised using the ECL detection system (Mbchem,
Shanghai, China) following the manufacturer's protocol. β-actin or
GAPDH was used as an internal control.
Immunofluorescence
The cells were grown on coverslips in 24-well
plates. Before staining, the cells were fixed with 4% formaldehyde
for 10 min, followed by 0.2–0.3% Triton X-100 for 10 min and then
were blocked in 1% BSA for 1 h. For staining, the cells were
incubated with the primary antibodies monoclonal anti-E-cadherin
(1:200, Abcam), anti-vimentin (1:200, Abcam) or anti-HIF-1α (1:200,
Abcam) for 1 h and then with goat anti-rabbit/mouse secondary
antibodies (1:1000, Invitrogen, Carlsbad, CA, USA) for 1 h at room
temperature. The nuclei of the cells were counterstained with DAPI
(1:10000, Invitrogen). After each step, cells were washed with PBS.
The images were acquired by laser scanning microscopy (Zeiss, Jena,
Germany).
Immunohistochemistry
E-cadherin, vimentin, HIF-1α, or pSTAT3 protein
expression in tissue was detected by immunohisto chemistry.
Sections (4 µm) were 10% formalin-fixed, paraffin-embedded,
deparaffinized in xylene, and then rehydrated through graded
ethanol solutions. The slides were incubated in 10 mM citrate
buffer (pH 6.0) at 95°C for 20 min, followed by 3% hydrogen
peroxide in methanol for 10 min, and then were blocked with 10%
bull serum albumin for 10 min in room temperature. The tissue
slides were incubated with anti-E-cadherin (1:100, Santa Cruz
Biotechnology), anti-vimentin (1:100, Santa Cruz Biotechnology),
anti-HIF-1α (1:100, Santa Cruz Biotechnology), or anti-pSTAT3
antibody (1:100, Santa Cruz Biotechnology) at 4°C overnight, and
with goat anti-rabbit/mouse secondary antibody (1:200, Santa Cruz
Biotechnology) for 30 min at room temperature. The sites of
antibody binding were visualized by 0.05% diaminobenzidine (DAB)
and stained with Mayer's hematoxylin. The slides were scored in a
blinded manner by two independent pathologists. The intensity of
protein expression was calculated by summing the percentage of
positively stained cells and the staining intensity.
Small interfering RNA (siRNA) and Small
hairpin RNA (shRNA) transfection
Human HIF-1α siRNA (Santa Cruz Biotechnology) and
STAT3 shRNA-expressing plasmid vector were used in the study. The
oligonucleotide encoding an shRNA targeting
5′-TGTTCTCTATCAGCACAAT-3′, starting at position 466 in the STAT3
sequence (GenBank Accession number: NM 39276) was cloned between
the HpaI and XhoI restriction sites of the plasmid
pGCL-GFP, which containing a CMV-driven GFP reporter (24), to constructed a vector mediating
RNAi (RNA interference) against STAT3 (pGCL-GFP-STAT3siRNA). HIF-1α
siRNA, empty vector PGCL-GFP, or pGCL-GFP-STAT3siRNA was
transfected into cells using Lipofectamine 2000 (Invitrogen)
following the manufacturer's instructions.
Chromatin immunoprecipitation (ChIP)
TE-1 and EC-1 cells were cultured in 100-mm dishes
at a density of 1×106/dish, serum-starved overnight, and
then cross-linked with 1% formaldehyde. Chromatin preparation and
immunoprecipitation were as previously described (15,25).
Chromatin was fragmented by sonication and was incubated with
anti-STAT3 (Santa Cruz Biotechnology) for the experiment, anti-RNA
polymerase II (Pol II) (Santa Cruz Biotechnology) for the positive
control, or rabbit polyclonal IgG (Santa Cruz Biotechnology) for
the negative control for 3 h at 4°C. Immunoprecipitation complex
was collected by vortexing with protein A/G agarose and extensive
washed, eluted, and purified. Then the DNA was used directly as
template for PCR detection. The possible binding sites of HIF-1α
were analysed using Primer Premier 5.0, DNA star and BLAST. Eight
pairs of primers for fragments covering the 3 kbp of the promoter
of the HIF-1α gene were designed: region 1 forward:
5′-GATATACTTACATAGTAATAA-3′, reverse: 5′-TGGATCATTTCAAAAG TTTAA-3′;
region 2 forward: 5′-GACTTAGATTTACTGTTACAA-3′, reverse:
5′-TTTTAGTAGAGACGAGGTTTC-3′; region 3 forward:
5′-AAAATTATCCGAGTGTGGTG-3′, reverse: 5′-CTTTTTAGTTTAACATTCTCA-3′;
region 4 forward: 5′-GTATTCTTTCCACAATTAAAA-3′, reverse:
5′-TAGTGGCTTTCAATTGTATTA-3′; region 5 forward:
5′-GATATTTATATTTACAACCCA-3′, reverse: 5′-ATTACC TAACCATGGGTCAAT-3′;
region 6 forward: 5′-TAGGTA ATCTGGTAAGGAAA-3′, reverse:
5′-CTCTCAGCCAATC AGGAGGC-3′; region 7 forward: 5′-GGACTTGCCGCC
TGCGTCGC-3′, reverse: 5′-TCCCTCCACACGCGGAG AAGA-3′; region 8
forward: 5′-GAACCCGCCTCCACCTC AGGT-3′, reverse:
5′-ATGTCTTCACGGCGGGCGGCC-3′.
Wound healing migration assay
TE-1 and EC-1 were seeded in 6-well plates, and
wounds were made by scratching the plates with sterile 200
µl pipet tips, five scratches per well. Cells were allowed
to continue growing in normoxia or with CoCl2 (100
µmol/l) in 5% CO2 at 37°C. Images were taken at
different times. The widths of the scratches were measured at five
different regions and are presented as mean ± SD.
Invasion assay
Twenty-four-well invasion chambers (Corning Inc.,
Corning, NY, USA) coated with Matrigel (BD Biocoat) were used as
described previously (25). Starved
cells (cultured in serum-free medium for 12 h before use) were
added to the rehydrated inserts with serum-free RPMI-1640, and 10%
FBS RPMI-1640 was added into the wells. One well of each plate was
left as a blank. The chambers were incubated in normoxia or with
CoCl2 (100 µmol/l) for 12 h. The cells that
passed through the filters to the lower surface of the insert
chambers were stained with 0.1% crystal violet, and then
counted.
Tumour xenografts
All animal work was done in accordance with a
protocol approved by the Laboratory Animal Management Committee of
Zhengzhou University. Six-week-old male, athymic, nude mice were
used for the xenografting experiment. EC-1 (1×106)
cells, mixed with an equal volume of Matrigel, were implanted
subcutaneously into the dorsal regions of mice. Mice were treated
with normal saline (as control), empty vector pGCL-GFP, or STAT3
siRNA by tail vein injection from day 7 after tumour implantation,
twice a week. PGCL-GFP or STAT3 siRNA was mixed with lipofecter
(Beyotime, Jiangsu, China) before injection (26,27).
Four weeks after implantation, mice were euthanized and the tumour
sizes were measured. Expression of pSTAT3, HIF-1α, E-cadherin, and
vimentin in the xenografts tumour was detected by
immunohistochemistry.
Statistical analyses
Statistical analyses were performed using IBM SPSS
Statistics 21.0 (SPSS Inc. IBM, Armonk, NY, USA). Differences
between two groups were assessed using the Student's t-test.
Differences among three or more groups were evaluated using one-way
ANOVAs. Data are shown as mean ± SD from three independent
experiments. P-values <0.05 were considered statistically
significant.
Results
Hypoxia induces EMT and activates STAT3
in ESCC cells
Hypoxia induces several kinds of tumour cells to
transform from epithelium to mesenchyme, both morphologically and
biologically. The tumours include pancreatic, breast, ovarian,
colon and hepatocellular cancers (28,29).
However, there are few studies focusing on hypoxia-induced EMT in
ESCC. To explore the effects of hypoxia on ESCC cells, we observed
the morphological changes and the migration and invasion abilities
of the ESCC cells, EC-1 and TE-1, in normoxia or hypoxia. The ESCC
cells had the morphology of mesenchymal cells: spindle shaped with
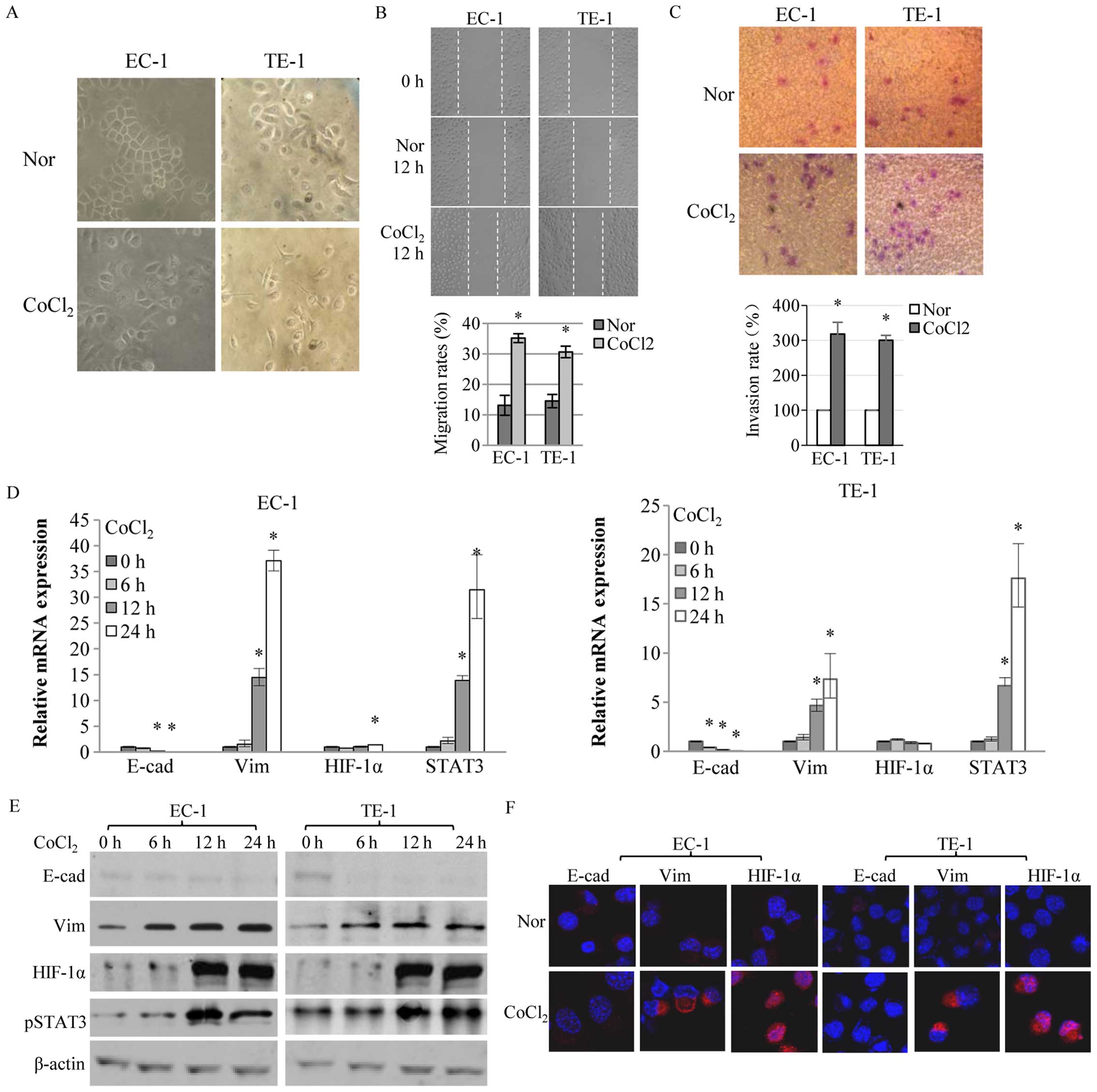
more pseudopodia in hypoxia or with CoCl2 (Fig. 1A). The migration and invasion rates
of ESCC cells were significantly enhanced in the hypoxia mimic,
CoCl2, than in normoxia (P<0.05; Fig. 1B and C). The EMT biomarkers,
E-cadherin and vimentin, and the hypoxic-response marker, HIF-1α,
were detected in ESCC cells in normoxia and hypoxia. When exposed
to CoCl2, mRNA and protein expression of E-cadherin
decreased and vimentin increased to levels higher than those in
normoxia. HIF-1α protein, but not mRNA, increased more sharply with
CoCl2 than in normoxia. Phosphorylated STAT3, the active
state of STAT3, increased in hypoxia (Fig. 1D and E), which was consistent with
earlier studies (30,31). Changes in EMT protein and HIF-1α
expression in CoCl2 were confirmed by immunofluorescence
assay (Fig. 1F). Previous studies
have shown that protein levels of HIF-1α and pSTAT3 increase with
CoCl2 as they do in 'real' hypoxia, and that tumour
cells performed similar EMTs in the hypoxia mimic,
CoCl2. Thus, the above molecular changes were not only
due to CoCl2 treatment, but also were reflective of
changes in 'real' hypoxia.
STAT3 regulates HIF-1α and EMT in
hypoxia
STAT3 plays important roles in EMT induced by TGFβ,
EGF, or IL-6 (32,33). To determine the effects of STAT3 on
EMT induced by hypoxia, we used STAT3 siRNA to knockdown STAT3 in
ESCC EC-1 cells. STAT3 mRNA and phosphorylated protein were
effectively silenced by STAT3 siRNA, both in normoxia and in
hypoxia. STAT3 siRNA downregulated HIF-1α mRNA and protein
expression in normoxia and hypoxia. The EMT markers E-cadherin and
vimentin were upregulated and downregulated (P<0.05),
respectively, and the tendencies were more evident in hypoxia, when
transfected with STAT3 siRNA (Fig.
2A–C). The invasion and migration rates of the cells declined
when treated with STAT3 siRNA (P<0.05), and this effect was
stronger in hypoxia (Fig. 2D and
E). The above results indicated that STAT3 knockdown regulates
HIF-1α and obliterates EMT induced by hypoxia. To observe the
effect of HIF-1α in hypoxia-induced EMT, we used HIF-1α siRNA to
block the expression of HIF-1α, and found that, in hypoxia,
inhibiting the expression of HIF-1α increases E-cadherin expression
and reduces vimentin expression. Based on the results of blocking
HIF-1α, inhibition of STAT3 by STAT3 siRNA can further increase
E-cadherin level and reduce vimentin expression (Fig. 2F and G).
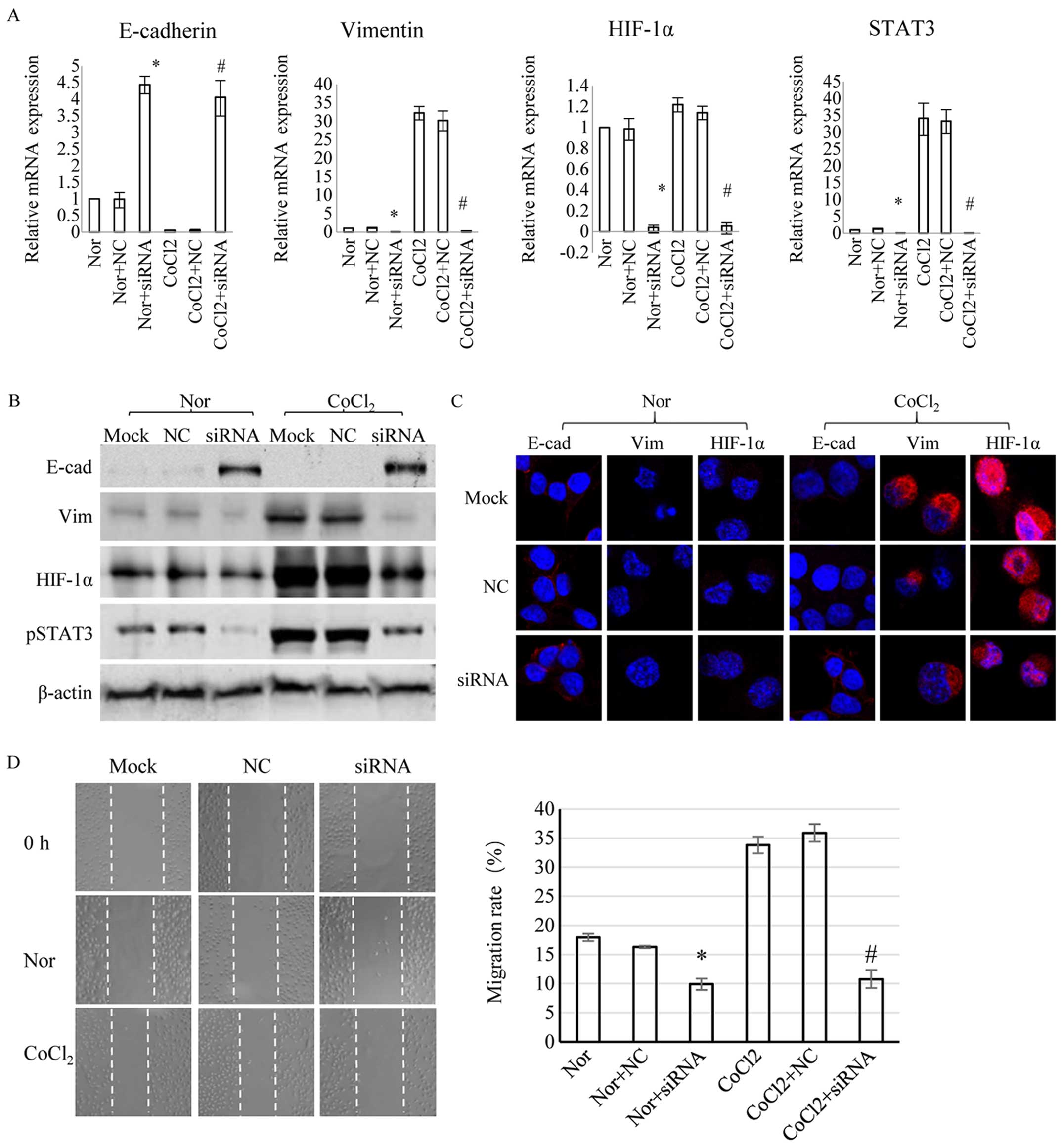 | Figure 2STAT3 regulates EMT induced by
hypoxia in ESCC. (A–D) PGCL-GFP-STAT3 siRNA (siRNA) or pGCL-GFP
(NC) was transfected into EC-1 cells with Lipofectamine 2000 for 24
h and incubated in normoxia (nor), or with CoCl2 (100
µmol/l). (A) Relative mRNA expression levels of E-cadherin
(E-cad), vimentin (vim), HIF-1α, and STAT3 in ESCC cells, with or
without CoCl2 (100 µmol/l) for 24 h. mRNA
expression was significantly different from that in the blank
control group in normoxia (*P<0.05). mRNA expression
was significantly different from that in the blank control group in
hypoxia (#P<0.05). (B) Western blot analysis of
E-cadherin, vimentin, HIF-1α, or pSTAT3 proteins, with or without
CoCl2 (100 µmol/l) for 24 h. Results were
normalized against β-actin. (C) IF of E-cadherin, vimentin, and
HIF-1α expression, with or without CoCl2 (100
µmol/l) for 24 h. (D) Cell migration was prevented by STAT3
inhibition. Scratches were made after transfections and the cells
were incubated in normoxia, or with CoCl2 (100
µmol/l), for 12 h. The cell migration rate was significantly
lower than in the blank control group in normoxia
(*P<0.05). The cell migration rate was significantly
lower than in the blank control group in hypoxia
(#P<0.05). STAT3 regulates EMT induced by hypoxia in
ESCC. (E and G) PGCL-GFP-STAT3 siRNA (siRNA) or pGCL-GFP (NC) was
transfected into EC-1 cells with Lipofectamine 2000 for 24 h and
incubated in normoxia (nor), or with CoCl2 (100
µmol/l). (E) Cells invasion was suppressed when STAT3 was
inhibited. Transfected cells seeded into invasion chambers and
incubated with or without CoCl2 (100 µmol/l) for
12 h, then were stained and detected. The cell invasion rate was
significantly lower than in the blank control group in normoxia
(*P<0.05). The cell invasion rate was significantly
lower than in the blank control group in hypoxia
(#P<0.05). The results represent means ± SD from
three independent experiments. (F) HIF-1α protein expression of
EC-1 cells with CoCl2 (100 µmol/l for 24 h) was
inhibited by HIF-1α siRNA, analyzed using western blot method. (G)
Western blot analysis of E-cadherin and vimentin protein expression
of EC-1 with HIF-1α siRNA and STAT3 siRNA with CoCl2
(100 µmol/l for 24 h). GAPDH served as a loading control in
(F and G). |
STAT3 binds the promoter of HIF-1α
gene
STAT3 is a transcription factor that regulates gene
expression and exerts its physiologic functions by binding to DNA.
Previous studies indicated that STAT3 increases HIF-1α protein
stability and interacts with HIF-1α to bind to HIF1 target gene
promoters. We noted that STAT3 affected mRNA expression of HIF-1α
in ESCC cell lines in normoxia and hypoxia conditions (Fig. 2A). To determine the specific
mechanisms of HIF-1α regulation by STAT3, we used in vivo
chromatin immunoprecipitation (ChIP) assays to detect STAT3 binding
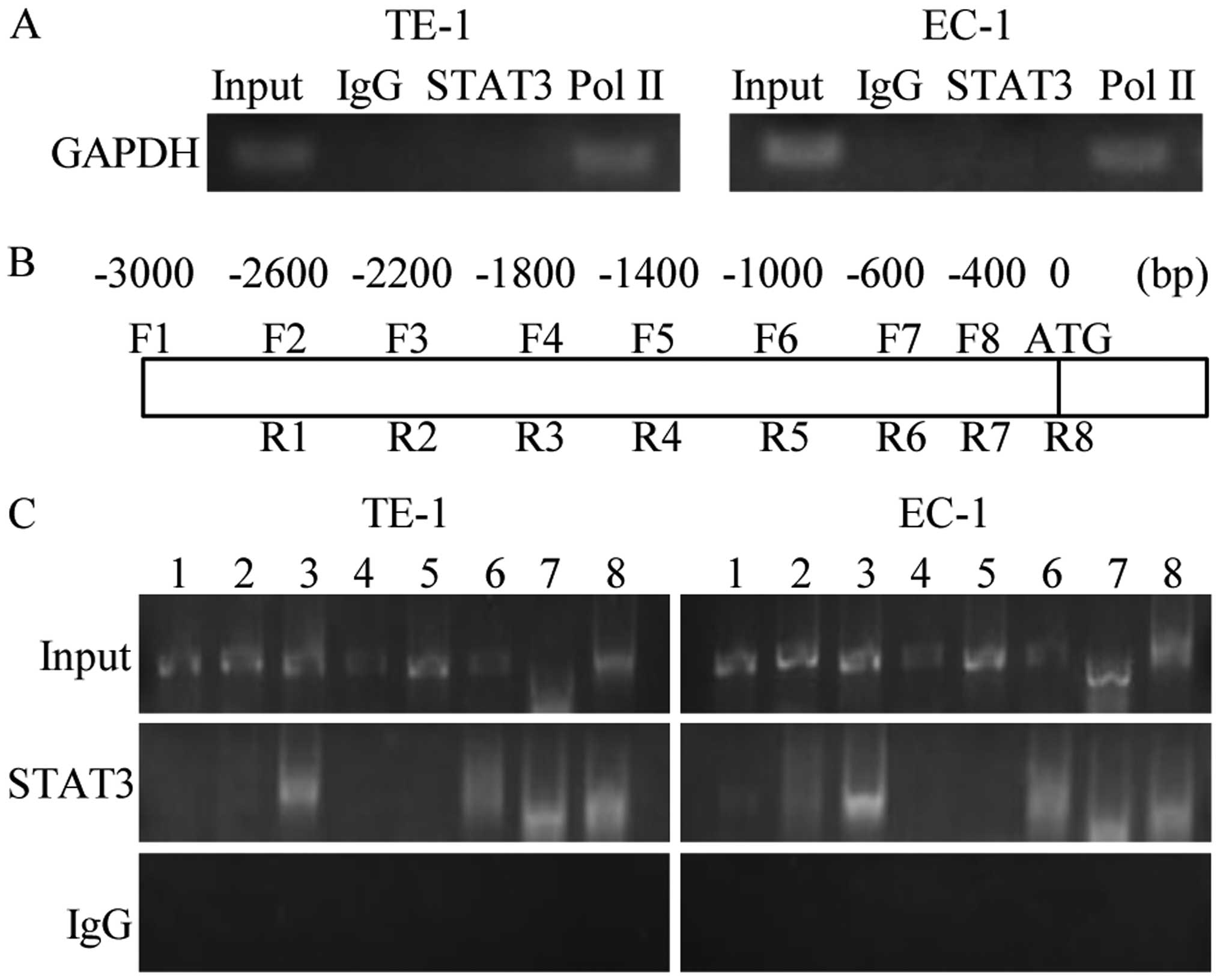
to the HIF-1α promoter (Fig. 3).
The results in Fig. 3A indicated
the efficiency of the immunoprecipitation, and that STAT3 did not
bind non-specifically to DNA. Eight pairs of primers were designed,
covering 3 kbp upstream of the HIF-1α open reading frame (Fig. 3B). As shown in Fig. 3C primer pairs 3, 6, 7, and 8
produced visible bands, suggesting that STAT3 binds to regions on
the HIF-1α promoter. The result shows that nuclear STAT3 binds to
the HIF-1α promoter of ESCC cells.
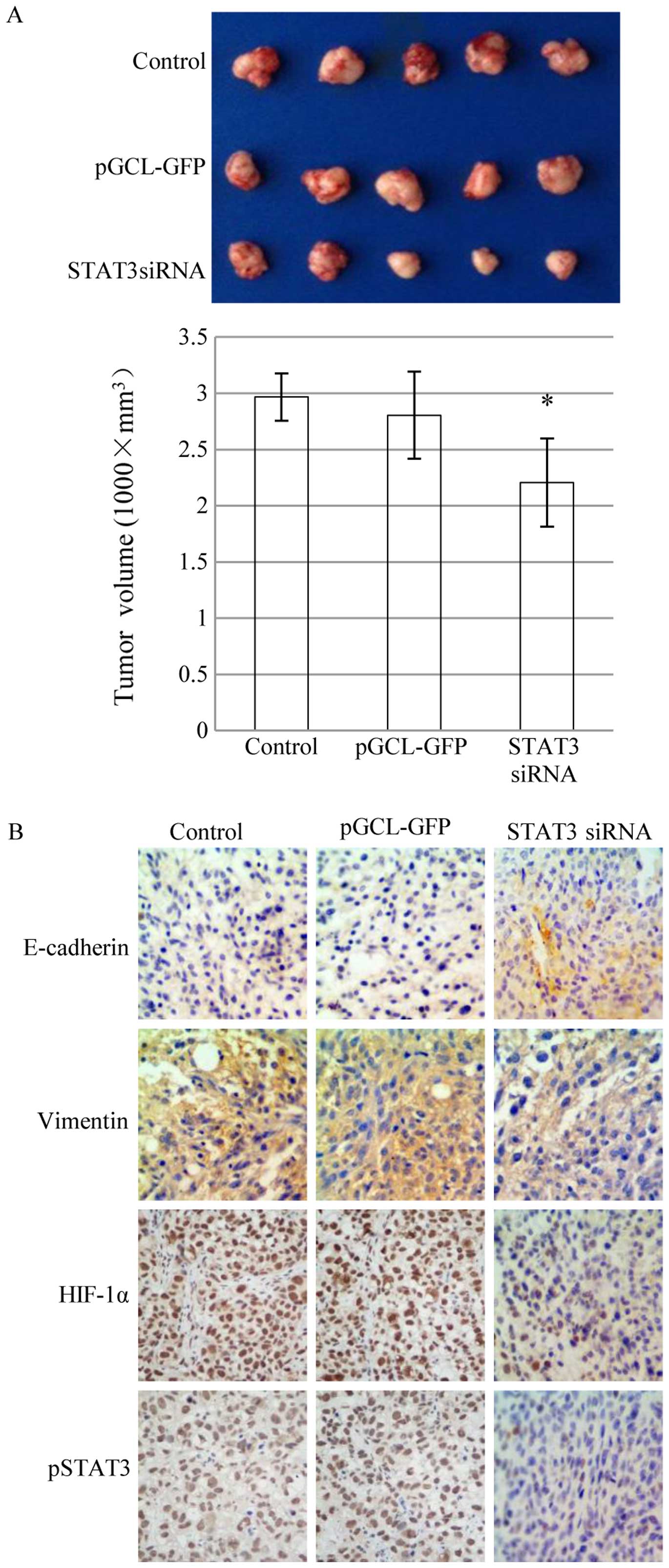
STAT3 siRNA inhibits EMT and tumour
progression of ESCC in vivo
STAT3 regulates HIF-1α expression and
hypoxia-induced EMT in ESCC in vitro. For in vivo
experiments, athymic nude mice were implanted with xenografted
human ESCC tumours. Mice were treated with STAT3 siRNA, or with the
controls, empty vector pGCL-GFP or saline. Tumour sizes were
smaller in the STAT3 siRNA treated group than in control groups
(P<0.05) (Fig. 4A). Lower
expression of p-STAT3, HIF-1α and vimentin, and higher expression
of E-cadherin, were detected in the treatment group than in the
control groups. The target proteins did not show differences in the
two control groups (Fig. 4B). Taken
together, these results indicated that STAT3 influences ESCC tumour
proliferation by regulating EMT via HIF-1α in ESCC in
vivo.
Discussion
Hypoxia is one of the driving factors of EMT, and
HIF-1α is the key factor of this process. Previous studies have
demonstrated hypoxia-induced EMT in breast, pancreatic, prostate,
and colon cancers. This study demonstrated hypoxia-induced EMT in
ESCC for the first time, which occurs through the upregulation of
HIF-1α. When HIF-1α expression is inhibited, hypoxia can no longer
induce EMT. In our previous study, we found that STAT3 regulates
EMT by targeting the transcription factors TWIST, SLUG, and SNAIL,
which regulate E-cadherin expression. Does STAT3 play a role in
hypoxia-induced EMT. In this study, we found that EMT in hypoxia is
reduced when STAT3 expression is silenced, indicating that STAT3 is
important in hypoxia-induced EMT. ESCCs undergo EMT in normoxic
conditions, as well, although weaker than in hypoxic condition. The
EMT in normoxic condition also was attenuated by inhibiting STAT3,
whereas HIF-1α at a low level. This phenomenon indicated that STAT3
regulates EMT in normoxic and in hypoxic conditions through
different pathways. The specific signalling networks involved in
STAT3-regulated EMT need further investigation.
HIF-1α and Stat3 are overexpressed in diverse
cancers and both play key roles in cancer progression. STAT3
affects HIF-1α expression and works with HIF-1α to adjust hypoxic
reactions. It was found that Stat3 is required for both basal and
growth signal-induced expression of HIF-1α. STAT3 binds to the
promoters of HIF1 target genes, interacting with HIF-1α protein and
recruiting a DNA binding complex to promote transcription. Our
study revealed the potential binding of STAT3 to the HIF-1α
promoter in ESCC cells, indicating that STAT3 transcriptionally
regulates HIF-1α expression. This is consistent with the results of
previous studies (34). The
difference is, our study found three potential binding sites in the
HIF-1α promoter, in addition to the Stat3 binding site at
position-373, which may play different roles in HIF-1α
transcription. HIF-1α mRNA levels do not increase in hypoxia, but
when STAT3 was silenced, HIF-1α mRNA declined significantly.
Perhaps the role of STAT3 is to maintain normal levels of HIF-1α
mRNA. Does STAT3 interact with HIF-1α in EMT? Our experiment showed
that HIF-1α and EMT were downregulated by STAT3 siRNA. This
suggests that STAT3 affects EMT induced by hypoxia by regulating
HIF-1α, which was previously shown to regulate transcriptional
regulators of E-cadherin. Taken together with prior studies, STAT3
might adjust EMT by targeting transcriptional regulators of
E-cadherin, directly or indirectly.
In conclusion, our study demonstrated, for the first
time, a key role of STAT3 in hypoxia-induced EMT in ESCC. Tumour
cells acquire stem cell-like and chemoresistant phenotypes from
EMT. Our findings may explain the radio/chemo resistance of ESCC
and the values of therapy that targets STAT3. Not all tumour cells
undergo EMT, and the process rarely occurs during the entire
process of tumour progression. When EMT cells arrive at their
destination, these mesenchymal cells may undergo the reverse
process, mesenchymal-epithelial transition (MET), and then adhere
to and proliferate in the new organs. The heterogeneity of
malignant tumour cells with mixed epithelial and mesenchymal traits
may enhance the ability of these cells to adapt changes in the
micro-environments, and to survive and evolve quickly. EMT is not
the complete story of tumour progression. Further studies are
required to clarify the precise mechanisms and signals that
regulate the transition between epithelial and mesenchymal
phenotypes, and the involvement of STAT3 and hypoxia in these
processes.
References
|
1
|
Książkiewicz M, Markiewicz A and Zaczek
AJ: Epithelial-mesenchymal transition: A hallmark in metastasis
formation linking circulating tumor cells and cancer stem cells.
Pathobiology. 79:195–208. 2012. View Article : Google Scholar
|
|
2
|
Tania M, Khan MA and Fu J: Epithelial to
mesenchymal transition inducing transcription factors and
metastatic cancer. Tumour Biol. 35:7335–7342. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Imai T, Horiuchi A, Wang C, Oka K, Ohira
S, Nikaido T and Konishi I: Hypoxia attenuates the expression of
E-cadherin via up-regulation of SNAIL in ovarian carcinoma cells.
Am J Pathol. 163:1437–1447. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Philip B, Ito K, Moreno-Sánchez R and
Ralph SJ: HIF expression and the role of hypoxic microenvironments
within primary tumours as protective sites driving cancer stem cell
renewal and metastatic progression. Carcinogenesis. 34:1699–1707.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yoo YG, Christensen J, Gu J and Huang LE:
HIF-1α mediates tumor hypoxia to confer a perpetual mesenchymal
phenotype for malignant progression. Sci Signal. 4:pt42011.
View Article : Google Scholar
|
|
6
|
Sahlgren C, Gustafsson MV, Jin S,
Poellinger L and Lendahl U: Notch signaling mediates
hypoxia-induced tumor cell migration and invasion. Proc Natl Acad
Sci USA. 105:6392–6397. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Copple BL: Hypoxia stimulates hepatocyte
epithelial to mesenchymal transition by hypoxia-inducible factor
and transforming growth factor-beta-dependent mechanisms. Liver
Int. 30:669–682. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li GQ, Zhang Y, Liu D, Qian YY, Zhang H,
Guo SY, Sunagawa M, Hisamitsu T and Liu YQ: PI3 kinase/Akt/HIF-1α
pathway is associated with hypoxia-induced epithelial-mesenchymal
transition in fibroblast-like synoviocytes of rheumatoid arthritis.
Mol Cell Biochem. 372:221–231. 2013. View Article : Google Scholar
|
|
9
|
Zhang Q, Bai X, Chen W, Ma T, Hu Q, Liang
C, Xie S, Chen C, Hu L, Xu S, et al: Wnt/β-catenin signaling
enhances hypoxia-induced epithelial-mesenchymal transition in
hepatocellular carcinoma via crosstalk with hif-1α signaling.
Carcinogenesis. 34:962–973. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Percio S, Coltella N, Grisanti S, Bernardi
R and Pattini L: A HIF-1 network reveals characteristics of
epithelial-mesenchymal transition in acute promyelocytic leukemia.
Genome Med. 6(84)2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brierley MM and Fish EN: STATs:
Multifaceted regulators of transcription. J Interferon Cytokine
Res. 25:733–744. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhao S, Venkatasubbarao K, Lazor JW,
Sperry J, Jin C, Cao L and Freeman JW: Inhibition of STAT3 Tyr705
phosphorylation by Smad4 suppresses transforming growth factor
beta-mediated invasion and metastasis in pancreatic cancer cells.
Cancer Res. 68:4221–4228. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Colomiere M, Ward AC, Riley C, Trenerry
MK, Cameron-Smith D, Findlay J, Ackland L and Ahmed N: Cross talk
of signals between EGFR and IL-6R through JAK2/STAT3 mediate
epithelial-mesenchymal transition in ovarian carcinomas. Br J
Cancer. 100:134–144. 2009. View Article : Google Scholar
|
|
14
|
Cho KH, Choi MJ, Jeong KJ, Kim JJ, Hwang
MH, Shin SC, Park CG and Lee HY: A ROS/STAT3/HIF-1α signaling
cascade mediates EGF-induced TWIST1 expression and prostate cancer
cell invasion. Prostate. 74:528–536. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lo HW, Hsu SC, Xia W, Cao X, Shih JY, Wei
Y, Abbruzzese JL, Hortobagyi GN and Hung MC: Epidermal growth
factor receptor cooperates with signal transducer and activator of
transcription 3 to induce epithelial-mesenchymal transition in
cancer cells via up-regulation of TWIST gene expression. Cancer
Res. 67:9066–9076. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang C, Yang G, Jiang T, Zhu G, Li H and
Qiu Z: The effects and mechanisms of blockage of STAT3 signaling
pathway on IL-6 inducing EMT in human pancreatic cancer cells in
vitro. Neoplasma. 58:396–405. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xiong H, Hong J, Du W, Lin YW, Ren LL,
Wang YC, Su WY, Wang JL, Cui Y, Wang ZH, et al: Roles of STAT3 and
ZEB1 proteins in E-cadherin down-regulation and human colorectal
cancer epithelial-mesenchymal transition. J Biol Chem.
287:5819–5832. 2012. View Article : Google Scholar :
|
|
18
|
Hu QD, Chen W, Yan TL, Ma T, Chen CL,
Liang C, Zhang Q, Xia XF, Liu H, Zhi X, et al: NSC 74859 enhances
doxorubicin cytotoxicity via inhibition of epithelial-mesenchymal
transition in hepatocellular carcinoma cells. Cancer Lett.
325:207–213. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yue P, Zhang X, Paladino D, Sengupta B,
Ahmad S, Holloway RW, Ingersoll SB and Turkson J: Hyperactive EGF
receptor, Jaks and Stat3 signaling promote enhanced colony-forming
ability, motility and migration of cisplatin-resistant ovarian
cancer cells. Oncogene. 31:2309–2322. 2012. View Article : Google Scholar
|
|
20
|
Jung JE, Lee HG, Cho IH, Chung DH, Yoon
SH, Yang YM, Lee JW, Choi S, Park JW, Ye SK, et al: STAT3 is a
potential modulator of HIF-1-mediated VEGF expression in human
renal carcinoma cells. FASEB J. 19:1296–1298. 2005.PubMed/NCBI
|
|
21
|
Pawlus MR, Wang L and Hu CJ: STAT3 and
HIF1α cooperatively activate HIF1 target genes in MDA-MB-231 and
RCC4 cells. Oncogene. 33:1670–1679. 2014. View Article : Google Scholar
|
|
22
|
Xu Q, Briggs J, Park S, Niu G, Kortylewski
M, Zhang S, Gritsko T, Turkson J, Kay H, Semenza GL, et al:
Targeting Stat3 blocks both HIF-1 and VEGF expression induced by
multiple oncogenic growth signaling pathways. Oncogene.
24:5552–5560. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ardyanto TD, Osaki M, Tokuyasu N, Nagahama
Y and Ito H: CoCl2-induced HIF-1alpha expression
correlates with proliferation and apoptosis in MKN-1 cells: A
possible role for the PI3K/Akt pathway. Int J Oncol. 29:549–555.
2006.PubMed/NCBI
|
|
24
|
Yang G, Huang C, Cao J, Huang KJ, Jiang T
and Qiu ZJ: Lentivirus-mediated shRNA interference targeting STAT3
inhibits human pancreatic cancer cell invasion. World J
Gastroenterol. 15:3757–3766. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lakka SS, Rajan M, Gondi C, Yanamandra N,
Chandrasekar N, Jasti SL, Adachi Y, Siddique K, Gujrati M, Olivero
W, et al: Adenovirus-mediated expression of antisense MMP-9 in
glioma cells inhibits tumor growth and invasion. Oncogene.
21:8011–8019. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Santel A, Aleku M, Röder N, Möpert K,
Durieux B, Janke O, Keil O, Endruschat J, Dames S, Lange C, et al:
Atu027 prevents pulmonary metastasis in experimental and
spontaneous mouse metastasis models. Clin Cancer Res. 16:5469–5480.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang HB, Yang X, Cao J, Li S, Liu YN, Suo
ZW, Cui HB, Guo Z and Hu XD: cAMP-dependent protein kinase
activated Fyn in spinal dorsal horn to regulate NMDA receptor
function during inflammatory pain. J Neurochem. 116:93–104. 2011.
View Article : Google Scholar
|
|
28
|
Cannito S, Novo E, Compagnone A, Valfrè di
Bonzo L, Busletta C, Zamara E, Paternostro C, Povero D, Bandino A,
Bozzo F, et al: Redox mechanisms switch on hypoxia-dependent
epithelial-mesenchymal transition in cancer cells. Carcinogenesis.
29:2267–2278. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang L, Huang G, Li X, Zhang Y, Jiang Y,
Shen J, Liu J, Wang Q, Zhu J, Feng X, et al: Hypoxia induces
epithelial-mesenchymal transition via activation of SNAI1 by
hypoxia-inducible factor-1α in hepatocellular carcinoma. BMC
Cancer. 13:1082013. View Article : Google Scholar
|
|
30
|
Matsuyama T, Nakanishi K, Hayashi T,
Yoshizumi Y, Aiko S, Sugiura Y, Tanimoto T, Uenoyama M, Ozeki Y and
Maehara T: Expression of hypoxia-inducible factor-1alpha in
esophageal squamous cell carcinoma. Cancer Sci. 96:176–182. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
You Z, Xu D, Ji J, Guo W, Zhu W and He J:
JAK/STAT signal pathway activation promotes progression and
survival of human oesophageal squamous cell carcinoma. Clin Transl
Oncol. 14:143–149. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang Y, Pan X, Lei W, Wang J, Shi J, Li F
and Song J: Regulation of transforming growth factor-beta 1-induced
apoptosis and epithelial-to-mesenchymal transition by protein
kinase A and signal transducers and activators of transcription 3.
Cancer Res. 66:8617–8624. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sullivan NJ, Sasser AK, Axel AE, Vesuna F,
Raman V, Ramirez N, Oberyszyn TM and Hall BM: Interleukin-6 induces
an epithelial-mesenchymal transition phenotype in human breast
cancer cells. Oncogene. 28:2940–2947. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Niu G, Briggs J, Deng J, Ma Y, Lee H,
Kortylewski M, Kujawski M, Kay H, Cress WD, Jove R, et al: Signal
transducer and activator of transcription 3 is required for
hypoxia-inducible factor-1alpha RNA expression in both tumor cells
and tumor-associated myeloid cells. Mol Cancer Res. 6:1099–1105.
2008. View Article : Google Scholar : PubMed/NCBI
|