Introduction
Ovarian cancer is regarded as the deadliest
gynecological cancer with the highest mortality rate worldwide. In
the year 2015, more than 21,290 new cases and 14,180 deaths were
reported all over the world (1).
Traditional methods with chemotherapy after surgery may cause
unwanted side-effects and drug resistance in patients. Therefore,
it is necessary to develop more effective drugs against ovarian
cancer. In recent studies, natural agents have been widely used for
cancer therapy. Grifolin (2-trans,
trans-farnesyl-5-methylresorcinol), a secondary metabolite
extracted from the mushroom Albatrellus confluens, is an
antibiotic belonging to basidiomycota (2,3).
Several mechanisms have been reported, including inhibition of G1
phase cell cycle (4–6), promotion of apoptosis proteins
(7,8) suppression of cell metastasis (9) which have been proposed to explain
grifolin's antitumor effects. However, the other mechanisms of the
drug's antitumor effects have remained unclear.
In the present study, we investigated the autophagic
effect of grifolin on human ovarian cancer cell lines A2780 and
SKOV3 in vitro. We found that grifolin inhibited cell growth
in the two types of cells through inducing autophagic cell death
via Akt/mTOR/S6K pathway. These results suggest that grifolin could
be a meaningful therapeutic for the treatment of human ovarian
cancer.
Materials and methods
Cell lines and culture
The human ovarian cancer cell line A2780 and SKOV3
were purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA) and were cultured in RPMI-1640 medium with 10%
fetal bovine serum (FBS). All cells were maintained at 37°C in a
humidified atmosphere containing 5% CO2.
Reagents and antibodies
Grifolin was kindly provided by Kunming Institute of
Botany, the Chinese Academy of Science and prepared at a
concentration of 100 mmol/l stock solution in dimethyl sulfoxide
(DMSO). Both chloroquine disphosphate (CQ) and acridine orange were
obtained from Abcam (Cambridge, UK). The antibodies to p-Akt, Akt,
p-mTOR, mTOR, p-p70S6K, p70S6K, p-S6, S6, p-4E-BP1 and 4E-BP1 were
all purchased from Cell Signaling Technology (Danvers, MA, USA),
the antibodies to LC3B, P62, Atg7, Beclin-1 and β-actin were
obtained from Abcam. The cell culture media and other reagents were
obtained from HyClone Laboratories (Logan, UT, USA).
Cell viability assay
Cell viability was measured by MTT assay. The A2780
and SKOV3 cells were seeded at a density of (4–5)×103 on 96-well plates and was
allowed to adhere overnight. The cells were then treated with
various concentration of the drugs (0, 20, 40, 60, 80 and 100
µM) to cells for 24, 48 and 72 h. At indicated time-points,
the cells in the 96-well plate were incubated with 20 µl MTT
and after 4 h at 37°C. The formazan product was dissolved in 100
µl DMSO and evaluated at 490 nm with a microplate reader
called infinite M200 PRO (Bio-Rad Laboratories, Hercules, CA,
USA.)
Flow cytometric analysis of acidic
vesicular organelles (AVOs)
The ovarian cancer cells were treated with grofolin
with different concentrations (0, 25, 50 and 75 µM) for 24 h
and then stained with acridine orange (1 µg/ml) in PBS at
37°C for 15 min in dark. Then the cells were washed with cold PBS
twice and re-suspended in PBS for analysis in 1 h. The data were
analyzed by CellQuest software.
Immunofluorescence staining
Analysis of LC3B protein localization: the cells
were plated in 24-well plates and treated with 50 µM
grifolin for 24 h. The cells were then fixed with 4%
paraformaldehyde for 15 min at room temperature and were
permeabilized with 0.2% Triton 150–200 µl in PBS for 10 min.
After that, the cells were blocked with indicated anti-LC3B
antibody (1:200) overnight and related secondary antibody (1:250)
for 1 h in dark and then stained with 4,6-diamidino-2-pheny-lindole
(DAPI) for 5 min in dark and at room temperature. At last, the
fluorescence images were observed using a DP71 fluorescence
microscope (Olympus, Tokyo, Japan).
Electron microscopy
The grifolin-treated cells were used to detect the
induction of autophagy in ovarian cancer. The cells were treated
with 50 µM grifolin for 48 h and harvested by trypsinization
and then fixed with cold fixative containing 2.5% glutaraldehyde
and 2% paraformaldehyde in 0.1 M cacodylate buffer. Then the
samples were post-fixed in 1% osmium tetroxide buffer
(OsO4). We observed the representative areas (90 nm thin
sections were cut) with a JEM-1010 transmission electron microscope
(JEOL USA, Inc., Peabody, MA, USA) at 80 kV.
Western blot analysis
Vehicle- or drug-treated cells were lysed in a mixed
buffer which contained RIPA, NaF, PMSF, and the supernatants were
collected and the protein levels were measured. Protein (20
µg) were resolved by 12 or 10% SDS-PAGE and transferred to
PVDF membranes (Immobilon P; Millipore, Bedford, MA, USA). After
blocking for 2 h using 5% non-fat milk, the stripes were incubated
with the indicated primary antibodies overnight at 4°C. This was
followed by incubation with secondary antibodies at room
temperature for 1–2 h. The protein signals were then detected by
ImageQuent LAS 4000.
GFP-LC3 puncta formation assay
Cells were plated in 6-well cell culture plates at
the confluence of 1×105 cells/well and incubated at 37°C
temperature with 5% CO2 overnight. Then the cells were
transfected with GFP-LC3 plasmid using Olifectamine and Opti-MEM
medium according to the manufacturer's protocol. shRNA
transfection. ShRNA was used to silence LC3B protein expression
in A2780 and SKOV3 cells. Establishment of A2780-plko.1-shLC3B,
A2780-plko.1-NC, SKOV3-plko.1-shLC3B and SKOV3-plko.1-NC cell lines
(shRNA-Forward primer
CCGGCGCTTACAGCTCAATGCTAATCTCGAGATTAGCATTGAGCTGTAAGCGTTTTTG and
shRNA-Reverse primer
AATTCAAAAACGCTTACAGCTCAATGCTAATCTCGAGATTAGCATTGAGCTGTAAGCG).
Statistical analysis
T-test and one-way ANOVA were performed to determine
significance by using the software SPSS 18.0. Statistical
significance was determined at P<0.05. All the experiments were
performed in triplicate.
Results
Grifolin induces cell proliferation in
human ovarian cancer cell lines
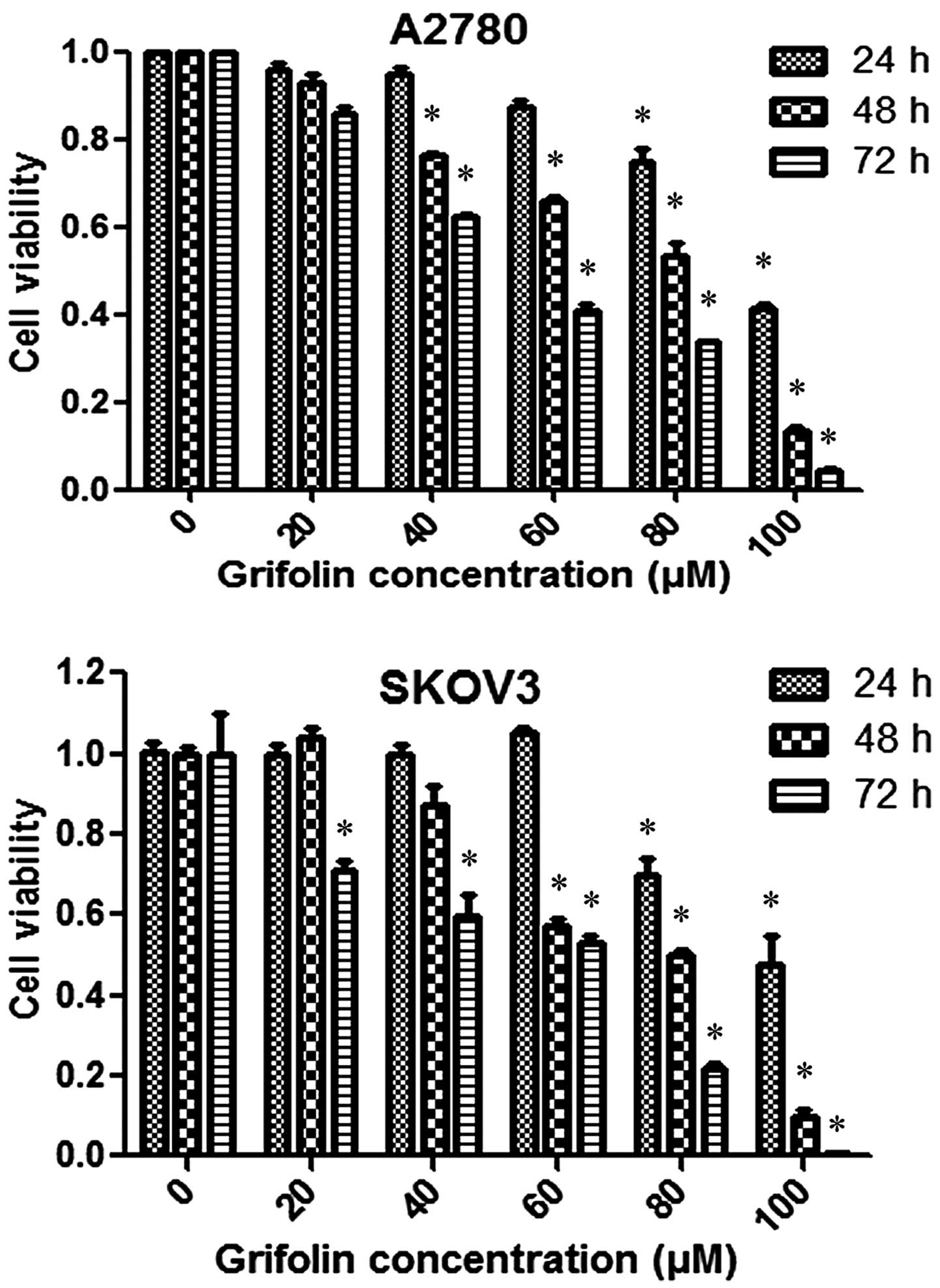
We examined the effect of grifolin on cell viability
of A2780 and SKOV3 cells which did decrease cell viability in the
two cell lines in a dose- and time-dependent manner, as shown in
Fig. 1.
Grifolin induces autophagy in ovarian
cancer cell lines
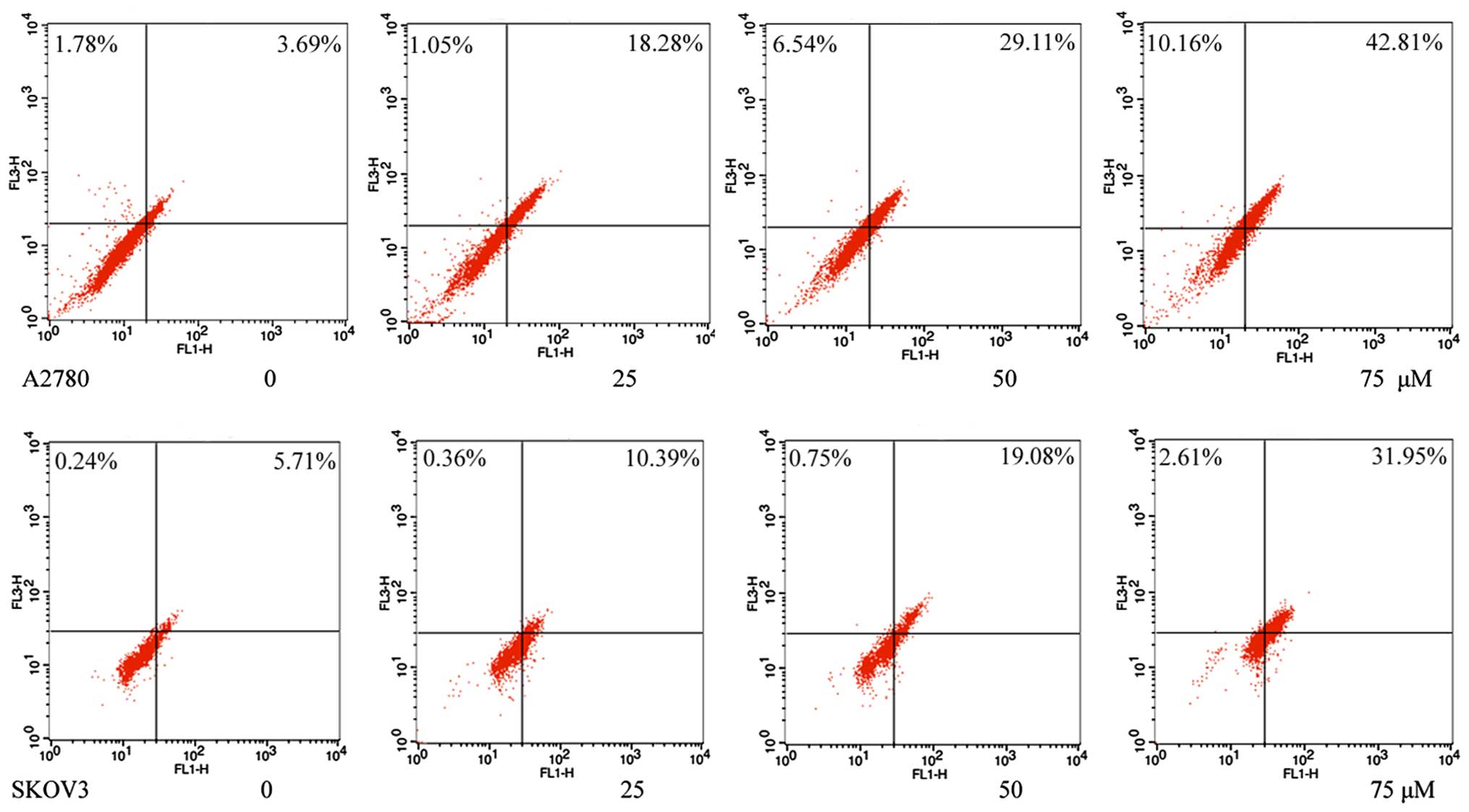
The induction of autophagy by grifolin was confirmed
by flow cytometry using acridine orange staining. As shown in
Fig. 2, grifolin-treated cells
increased formation of AVOs in a dose-dependent manner.
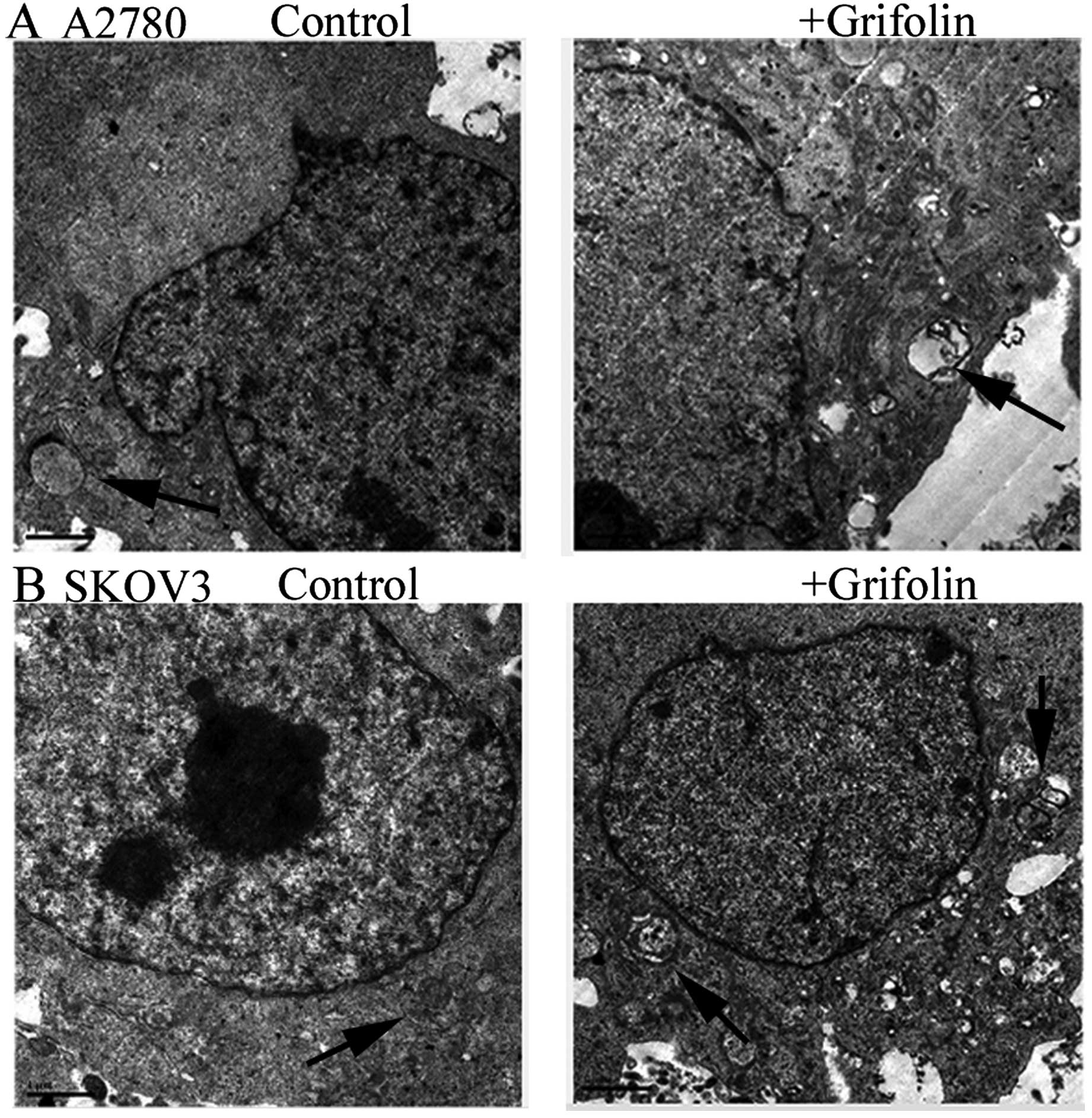
In the grifolin-treated cells, autophagy could cause
ultrastructural changes. In Fig. 3,
we observed autophagy in grifolin-treated cells by transmission
electron microscopy. The cells were treated with 50 µM
grifolin for 48 h, after which, we were able to observe the
characteristics of cells undergoing autophagy.
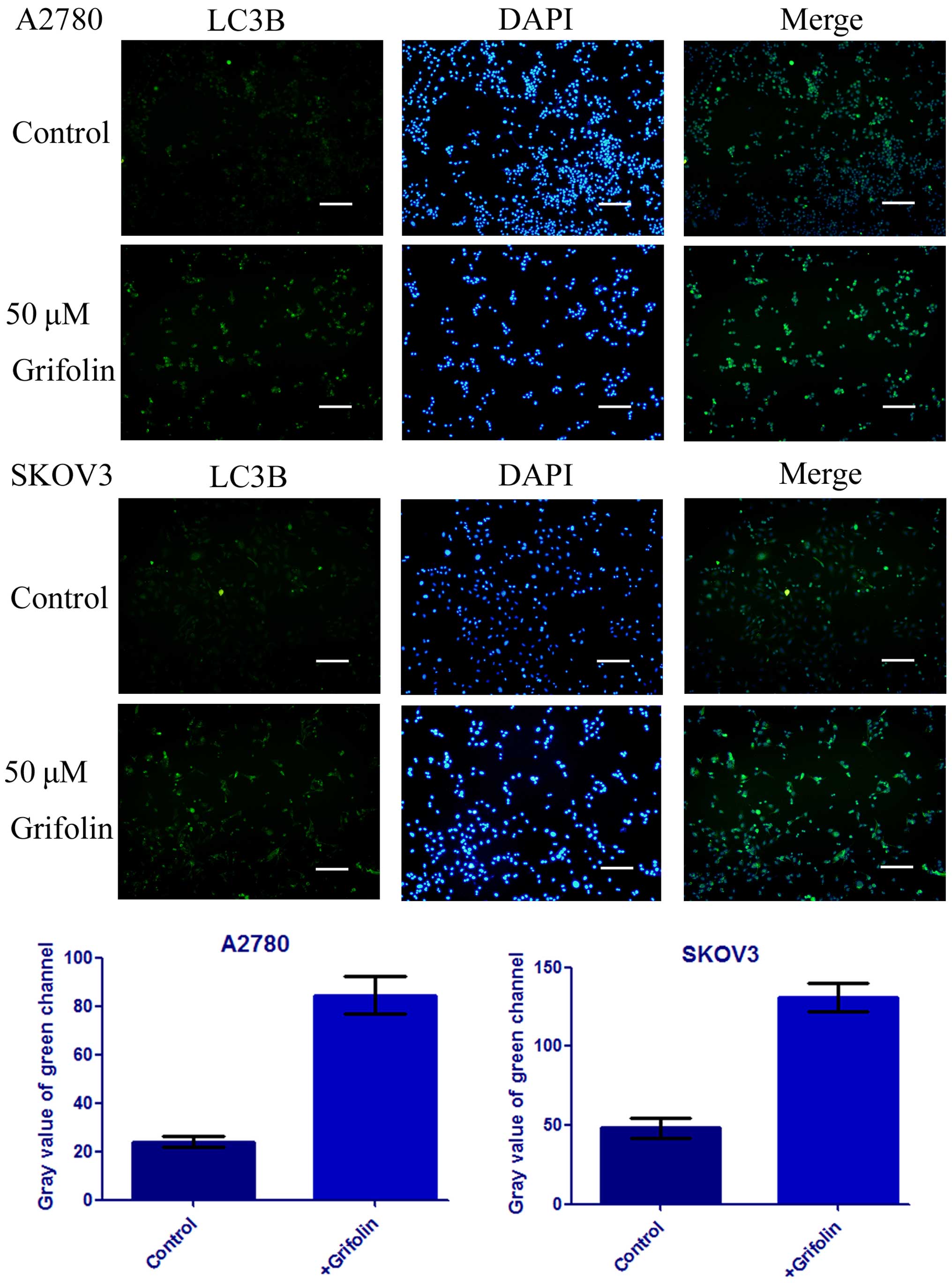
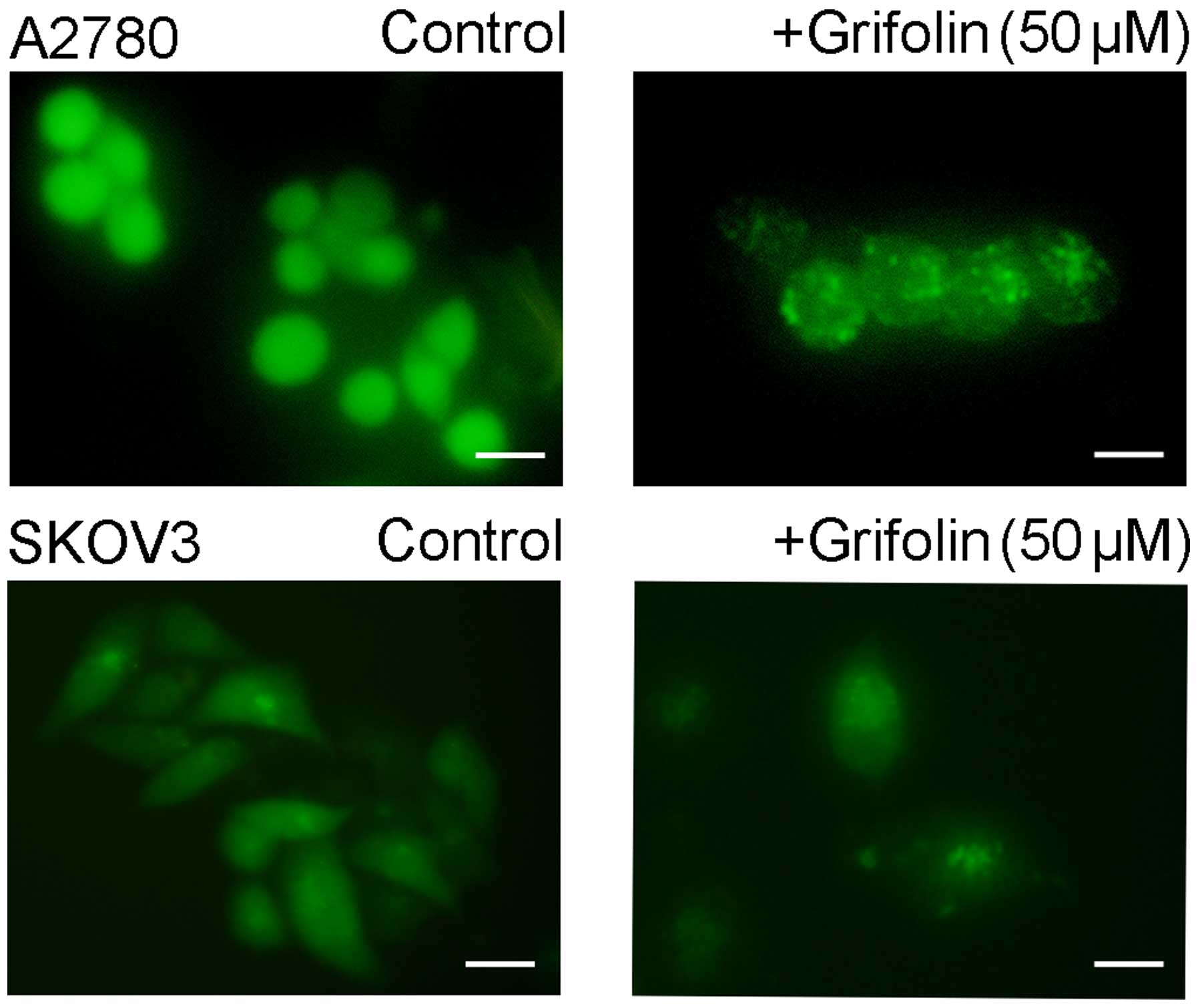
It has been well-established that the
microtubule-associated protein 1 light chain 3 (LC3) is a protein
associated with the autophagosomal membrane (10). After being treated with 50 µM
grifolin for 24 h, the number and intensity of punctuate LC3B
fluorescence increased as shown in Fig.
4. Next, we investigated the expression of autophagy-related
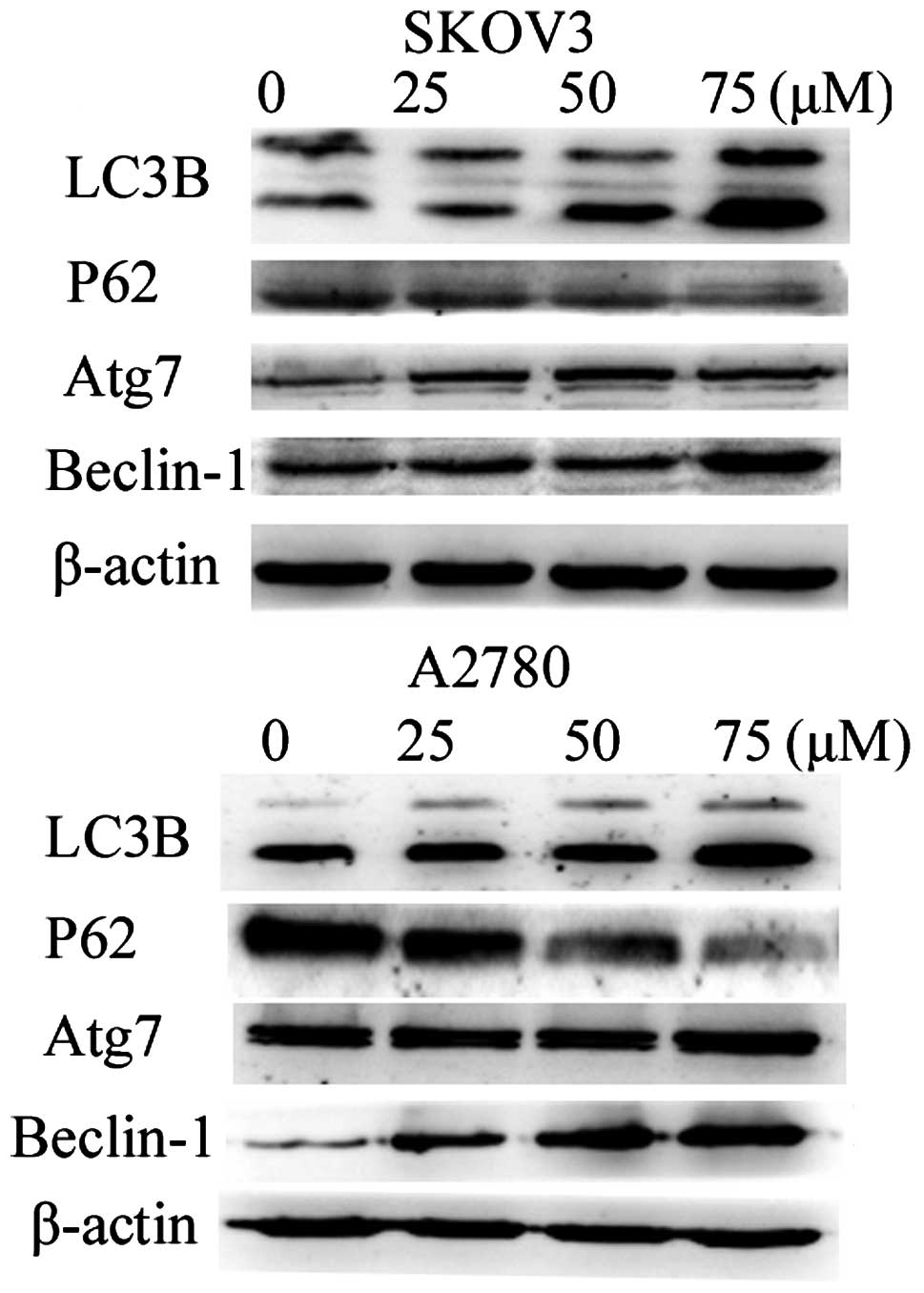
genes by western blot analysis. We found a significant increase in
LC3B, Atg7, Beclin-1 and a decrease in P62 in a dose-dependent
manner as shown in Fig. 5.
For further studying the role of grifolin in
inducing autophagy in human ovarian cancer cell lines, the effect
of grifolin on autophagy was confirmed by a GFP-LC3 puncta
formation assay. As shown in Fig.
6, we find increased number of GFP-LC3 puncta in the
grifolin-treated A2780 and SKOV3 cells.
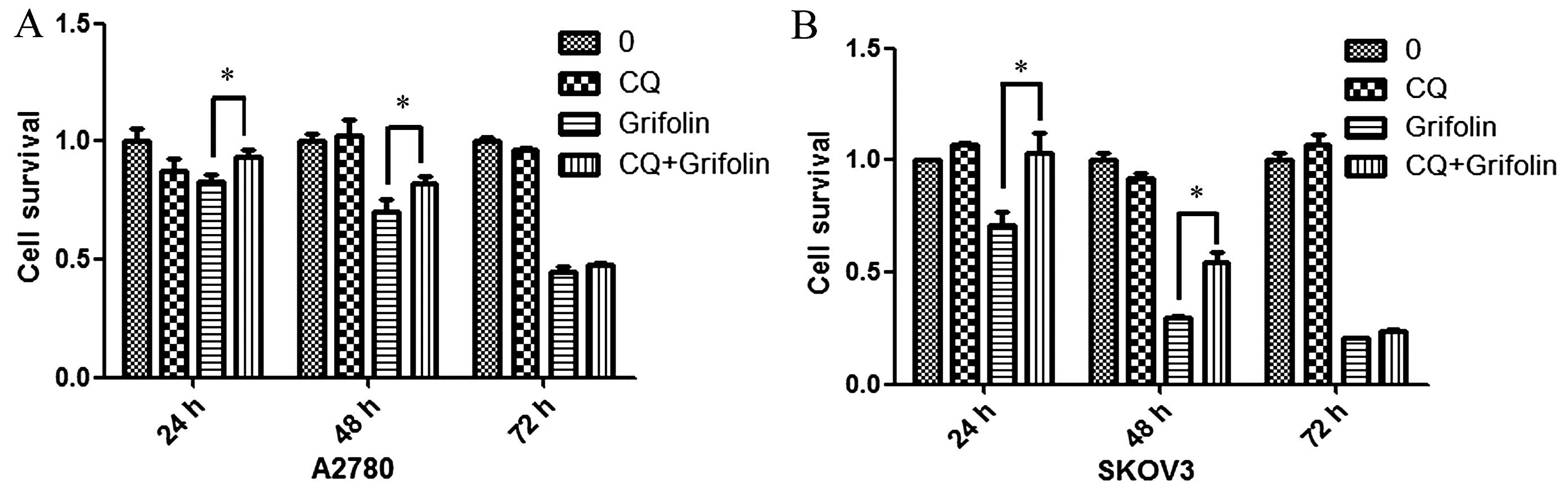
Next, we used the autophagy inhibitor chloroquine
(CQ) to investigate whether grifolin induced ovarian cancer cell
death through the induction of autophagy. CQ is a well-known
inhibitor of autophagy and inhibits lysosome acidification and
degradation (11). We used MTT
assay to detect cell viability (as shown in Fig. 7) using 10 µM of CQ which was
able to significantly reduce the cell death in A2780 and SKOV3 cell
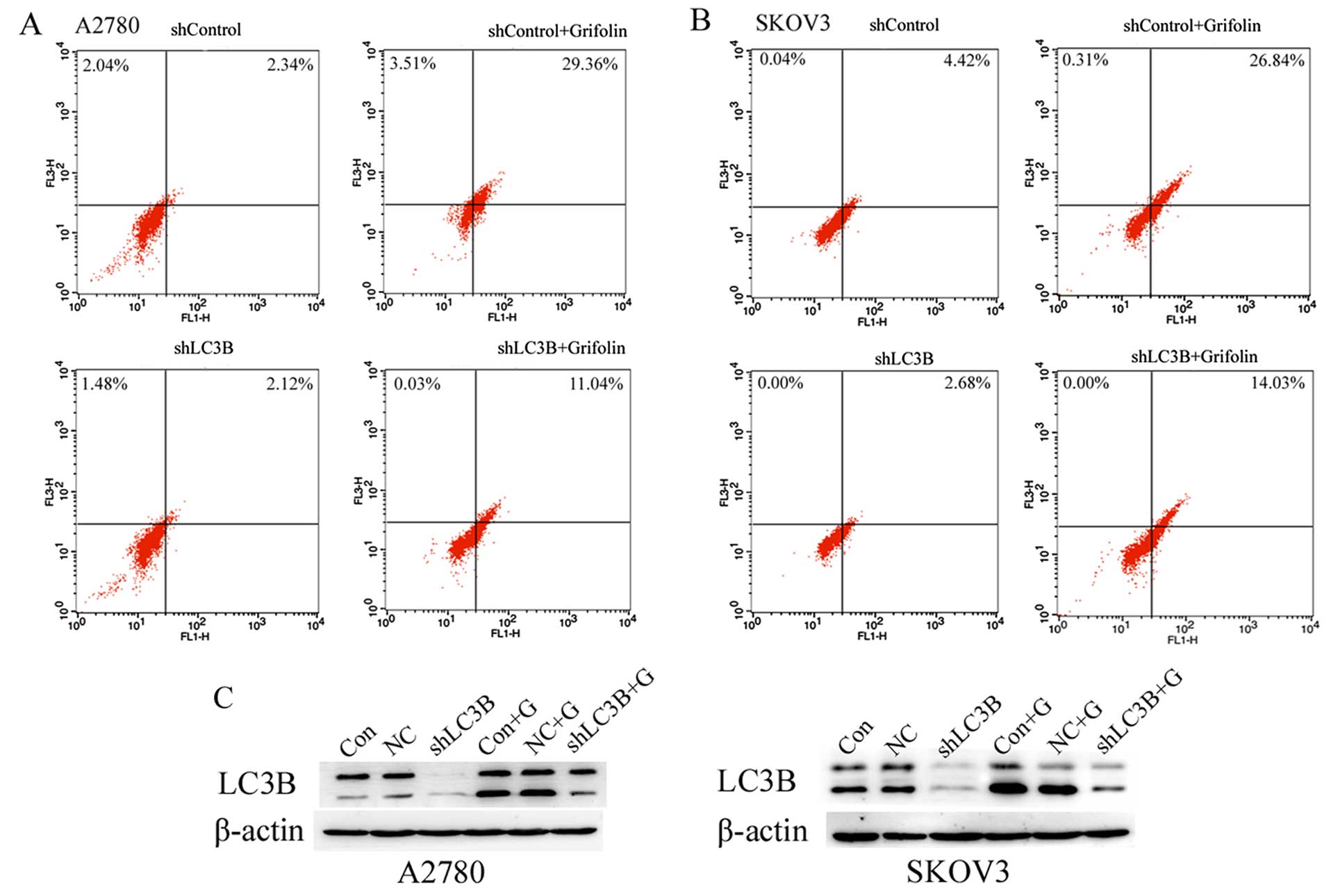
lines. To prove the role of LC3B on autophagy in the
grifolin-treated human ovarian cancer cells, we silenced the
expression of LC3B and measured the autophagy level by flow
cytometry-AVO analysis. As demonstrated in Fig. 8A and B, we found that transfection
with shLC3B blocked the autophagic effect of grifolin-treated A2780
and SKOV3 cells. Then we measured the level of LC3B by western blot
analysis. Fig. 8C demonstrates that
knockdown of LC3B in A2780 and SKOV3 cells significantly decreased
its expression level, and the expression level of LC3B protein in
grifolin-treated shLC3B cells expressed was higher than the
negative control group. These results suggested that grifolin could
reduce autophagic cell death in human ovarian cancer cell
lines.
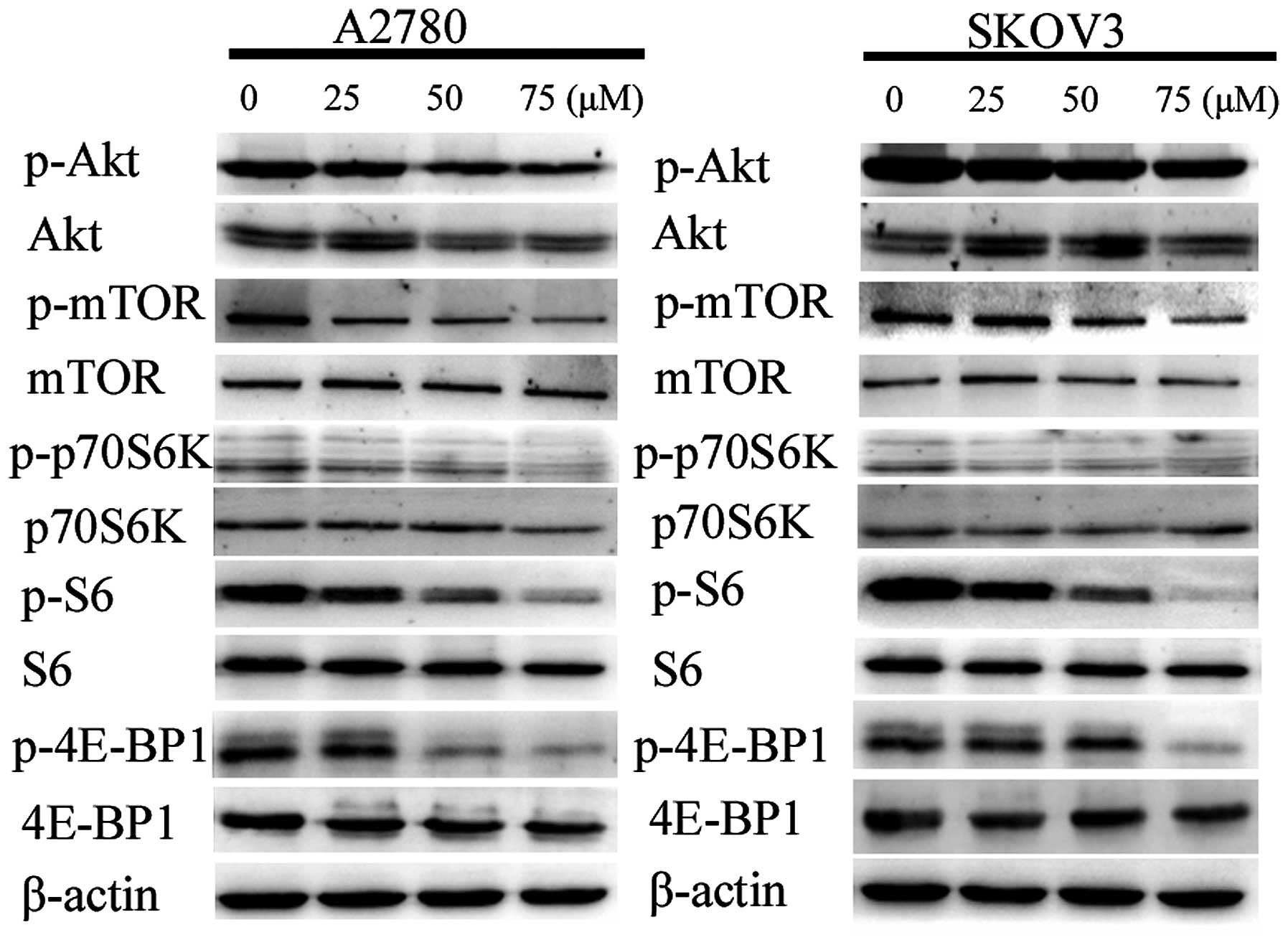
Grifolin inhibits the activity of the
Akt/mTOR/S6K pathway
Researchers have found that the inhibition of
Akt/mTOR/S6K pathway is associated with autophagy in various cancer
cells (12–14). Thus, we examined whether the pathway
is associated with grifolin-treated ovarian cancer cell autophagic
death using western blot analysis (Fig.
9). The results demonstrated that grifolin caused a decrease in
levels of the phosphorylated form of Akt, mTOR, p70S6K, S6 and
4E-BP1 while the total of these proteins remained unaffected by the
treatment. Those results suggested that grifolin could induce
autophagic cell death by inhibiting the activity of Akt/mTOR/S6K
pathway.
Discussion
In the present study, we investigated the role of
autophagy in human ovarian cancer cell lines A2780 and SKOV3 which
were treated with grifolin by inhibiting the Akt/mTOR/S6K pathway.
This is the first study on the role of grifolin in inducing
autophagy in ovarian cancer. A previous study showed that grifolin
induced cell cycle arrest in G1 phase via the ERK1/2 pathway
(6), and also induced apoptosis
through the Bax/Bcl-2 and caspase-3/-9 families via inhibition of
PI3K/Akt signalling pathway in human osteosarcoma cells (8). In addition, it has been reported that
grifolin upregulated death-associated protein kinase 1 DAPK1 via
p53 and mediated G1 phase arrest in nasopharyngeal carcinoma cells
(4,7). According to the latest reports,
grifolin suppressed cancer cell metastasis by inhibiting ERK1/2
pathway (9).
The Akt/mTOR/S6K pathway plays a significant role in
biological functions on various human cancers. A body of evidence
proved that Akt is a major component of the Akt/mTOR/S6K pathway
and its inactivation promotes cell proliferation and reduces cell
death (15). mTOR, a major
down-stream target of the pathway, is essential to regulate tumor
growth (16). In addition, S6K
possesses a key role in cell proliferation and survival (17). The Akt/mTOR/S6K pathway is becoming
an attractive therapeutic target for cancer therapy and its
inactivation is reported to occur in various tumors (18,19).
Kondo et al (20) found that
autophagy is induced mainly through the PI3K/Akt/mTOR pathway. In
recent years, many studies demonstrated that the pathway regulates
autophagy in various cancers such as breast cancer (21), human hepatocellular carcinoma
(22), ovarian cancer (12), melanoma (13), non-small cell lung (23), and nasopharyngeal cancer (14) and the mechanisms of autophagy are
well-known. The main mechanism is led by mTOR kinase that acts as
an upstream factor of all autophagy-associated genes and regulates
transcription and translation (24). The other important mechanism
involves the effect of mTOR on two main scaffold proteins the Atg11
and Atg 17, the changes in which lead to activation of Atg1, a key
autophagy kinase and then the activated Atg1 is able to affect the
output of autophagsome formation (25). In the present study, we investigated
that the main proteins of Akt/mTOR/S6K pathway in grifolin-treated
ovarian cancer cells. Our results show that grifolin treatment
downregulates phosphorylation of Akt and mTOR and also their
downstream targets, p70S6K, S6 and 4E-BP1 while the total proteins
have no obvious changes. Those findings prove that grifolin could
inhibit autophagy via inhibiting Akt/mTOR/S6K pathway on human
ovarian cancer cells.
In summary, we have shown for the first time that
grifolin induces autophagic cell death in human ovarian cancer
cells by inhibiting the Akt/mTOR/S6K pathway. We provide evidence
to prove grifolin could play a role as a novel antitumor agent for
human ovarian cancer through the induction of autophagy. However,
it is still crucial to explore the responsible molecular mechanisms
of grifolin inducing human ovarian cancer cell death.
Acknowledgments
The present study was partly funded by the National
Natural Science Foundation of China (no. 81072121 and no. 81372808
to J.J. and no. 81173614 to Q.T.L) and was also partly funded by
the Science and Technology Development Planning of Shandong (no.
2012G0021823 to J.J. and no. 2011GSF12122 to X.Z and J.J.), the
Science and Technology Development Planning of Jinan (201303035)
and the Science Foundation of Qilu Hospital of Shandong University
(2015QLMS44).
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Quang DN, Hashimoto T, Arakawa Y, Kohchi
C, Nishizawa T, Soma G and Asakawa Y: Grifolin derivatives from
Albatrellus caeruleoporus, new inhibitors of nitric oxide
production in RAW 264.7 cells. Bioorg Med Chem. 14:164–168. 2006.
View Article : Google Scholar
|
|
3
|
Liu XT, Winkler AL, Schwan WR, Volk TJ,
Rott MA and Monte A: Antibacterial compounds from mushrooms I: A
lanostane-type triterpene and prenylphenol derivatives from
Jahnoporus hirtus and Albatrellus flettii and their activities
against Bacillus cereus and Enterococcus faecalis. Planta Med.
76:182–185. 2010. View Article : Google Scholar
|
|
4
|
Luo X, Yu X, Liu S, Deng Q, Liu X, Peng S,
Li H, Liu J and Cao Y: The role of targeting kinase activity by
natural products in cancer chemoprevention and chemotherapy
(Review). Oncol Rep. 34:547–554. 2015.PubMed/NCBI
|
|
5
|
Luo XJ, Li W, Yang LF, Yu XF, Xiao LB,
Tang M, Dong X, Deng QP, Bode AM, Liu JK, et al: DAPK1 mediates the
G1 phase arrest in human nasopharyngeal carcinoma cells induced by
grifolin, a potential antitumor natural product. Eur J Pharmacol.
670:427–434. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ye M, Luo X, Li L, Shi Y, Tan M, Weng X,
Li W, Liu J and Cao Y: Grifolin, a potential antitumor natural
product from the mushroom Albatrellus confluens, induces cell-cycle
arrest in G1 phase via the ERK1/2 pathway. Cancer Lett.
258:199–207. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Luo XJ, Li LL, Deng QP, Yu XF, Yang LF,
Luo FJ, Xiao LB, Chen XY, Ye M, Liu JK, et al: Grifolin, a potent
antitumour natural product upregulates death-associated protein
kinase 1 DAPK1 via p53 in nasopharyngeal carcinoma cells. Eur J
Cancer. 47:316–325. 2011. View Article : Google Scholar
|
|
8
|
Jin S, Pang RP, Shen JN, Huang G, Wang J
and Zhou JG: Grifolin induces apoptosis via inhibition of PI3K/AKT
signalling pathway in human osteosarcoma cells. Apoptosis.
12:1317–1326. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Luo X, Yang L, Xiao L, Xia X, Dong X,
Zhong J, Liu Y, Li N, Chen L, Li H, et al: Grifolin directly
targets ERK1/2 to epigenetically suppress cancer cell metastasis.
Oncotarget. 6:42704–42716. 2015.PubMed/NCBI
|
|
10
|
Liang C, Feng P, Ku B, Dotan I, Canaani D,
Oh BH and Jung JU: Autophagic and tumour suppressor activity of a
novel Beclin1-binding protein UVRAG. Nat Cell Biol. 8:688–699.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mizushima N, Yoshimori T and Levine B:
Methods in mammalian autophagy research. Cell. 140:313–326. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zi D, Zhou ZW, Yang YJ, Huang L, Zhou ZL,
He SM, He ZX and Zhou SF: Danusertib induces apoptosis, cell cycle
arrest, and autophagy but inhibits epithelial to Mesenchymal
transition involving PI3K/Akt/mTOR signaling pathway in human
ovarian cancer cells. Int J Mol Sci. 16:27228–27251. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao G, Han X, Zheng S, Li Z, Sha Y, Ni J,
Sun Z, Qiao S and Song Z: Curcumin induces autophagy, inhibits
proliferation and invasion by downregulating AKT/mTOR signaling
pathway in human melanoma cells. Oncol Rep. 35:1065–1074. 2016.
|
|
14
|
Wang KF, Yang H, Jiang WQ, Li S and Cai
YC: Puquitinib mesylate (XC-302) induces autophagy via inhibiting
the PI3K/AKT/mTOR signaling pathway in nasopharyngeal cancer cells.
Int J Mol Med. 36:1556–1562. 2015.PubMed/NCBI
|
|
15
|
Blanco-Aparicio C and Renner O: PTEN, more
than the AKT pathway. Carcinogenesis. 28:1379–1386. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fingar DC, Salama S, Tsou C, Harlow E and
Blenis J: Mammalian cell size is controlled by mTOR and its
downstream targets S6K1 and 4EBP1/eIF4E. Genes Dev. 16:1472–1487.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Harada H, Andersen JS, Mann M, Terada N
and Korsmeyer SJ: p70S6 kinase signals cell survival as well as
growth, inactivating the pro-apoptotic molecule BAD. Proc Natl Acad
Sci USA. 98:9666–9670. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Feigin ME, Akshinthala SD, Araki K,
Rosenberg AZ, Muthuswamy LB, Martin B, Lehmann BD, Berman HK,
Pietenpol JA, Cardiff RD, et al: Mislocalization of the cell
polarity protein scribble promotes mammary tumorigenesis and is
associated with basal breast cancer. Cancer Res. 74:3180–3194.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vivanco I and Sawyers CL: The
phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat Rev
Cancer. 2:489–501. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kondo Y, Kanzawa T, Sawaya R and Kondo S:
The role of autophagy in cancer development and response to
therapy. Nat Rev Cancer. 5:726–734. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang X, Qi W, Li Y, Zhang N, Dong L, Sun
M, Cun J, Zhang Y, Lv S and Yang Q: Huaier extract induces
autophagic cell death by inhibiting the mTOR/S6K pathway in breast
cancer cells. PLoS One. 10:e01317712015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shi YM, Yang L, Geng YD, Zhang C and Kong
LY: Polyphyllin I induced-apoptosis is enhanced by inhibition of
autophagy in human hepatocellular carcinoma cells. Phytomedicine.
22:1139–1149. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li YR, Li S, Ho CT, Chang YH, Tan KT,
Chung TW, Wang BY, Chen YK and Lin CC: Tangeretin derivative,
5-Acetyloxy-6, 7, 8,4′-tetramethoxyflavone induces G2/M arrest,
apoptosis and autophagy in human non-small cell lung cancer cells
in vitro and In vivo. Cancer Biol Ther. 17:48–64. 2016. View Article : Google Scholar
|
|
24
|
Wang CW and Klionsky DJ: The molecular
mechanism of autophagy. Mol Med. 9:65–76. 2003.PubMed/NCBI
|
|
25
|
Kamber RA, Shoemaker CJ and Denic V: A
molecular switch for selective autophagosome formation. Autophagy.
11:2132–2133. 2015. View Article : Google Scholar : PubMed/NCBI
|