1. Introduction
Rapamycin, also known as sirolimus is an antifungal
agent that contains a macrocyclic lactone. It was isolated from a
strain of the soil bacterium, Streptomyces hygroscopicus,
found in Easter Island (known locally as Rapa Nui) and named as
rapamycin according to the name of its place of discovery (1,2). It
was also discovered to possess potent immunosuppressive,
antiproliferative and anticancer properties due to its ability to
arrest the cell cycle in the G1 phase (3).
The mammalian target of rapamycin (mTOR), also known
as FKBP12-rapamycin complex-associated protein (FRAP) is a
serine/threonine protein kinase that belongs to the
phosphoinositide-3-kinase (PI3K)-related kinase family (4,5).
Rapamycin binds with its intracellular receptor FK506-binding
protein (FKBP12) and forms a complex (mTOR) and exerts inhibitory
activity (6). mTOR is a large,
ubiquitously expressed multi-effector protein that in humans is
encoded by the MTOR gene. It is composed of 2,549 amino
acids with a molecular mass of ~250 kDa (7). The N-terminal half of the protein
contains 20 tandem HEAT repeats, each of which consists of 2α
helices of ~40 amino acids, and are implicated in protein-protein
interactions. The C-terminal half of mTOR contains the kinase
catalytic domain (KIN) that has sequence similarity with the
catalytic site in PI3Ks (6,7). It is reported to play a vital role in
controlling cell growth, cell survival, angiogenesis and autophagy
through the regulation of protein synthesis (5,8).
Deregulation of mTOR activity is associated with
numerous types of cancer. Oncogenic activation of mTOR signaling
through phosphorylation of PI3K or Akt significantly contributes to
the initiation and development of tumors. Over-activation of mTOR
activity supports tumor growth by promoting cell cycle progression,
increasing cell proliferation, and inhibiting autophagy through its
effect on protein synthesis (9,10).
Multimodal treatment strategies have been developed to target this
pathway resulting in the emergence of unique pharmacological
inhibitors that may provide the best therapeutic advantage for the
treatment of cancers (11,12).
Osteosarcoma (OS) is the most common primary
malignant tumor of bone occurring mostly in children and young
adults. It arises from the primitive transformed cells of
mesenchymal origin that mainly affects osteoblastic differentiation
and produces immature bone (13,14).
It mainly occurs in the tubular long bones; however also occurs in
other sites such as the femur, humerus, knee and pelvis. OS is the
eighth most common form of pediatric cancer with a higher incidence
in males (5.4/million/year) than in females (4.0/million/year)
(15). The typical signs and
symptoms of OS include pain followed by the localized swelling and
limitations of joint movement (14). The exact etiology of OS is unknown,
however it is evident that various risk factors are associated with
the development and pathogenesis of OS including age, gender,
ethnicity, genetic and familial factors (16). The current treatment options for OS
are surgery, chemotherapy and radiotherapy. The standard therapy of
OS is a combination of orthopedic surgery along with doxorubicin,
cisplatin, adriamycin, high-dose methotrexate with leucovorin
rescue and ifosfamide (14,17). Despite the multimodal therapy, the
conventional therapy of OS is still unsatisfactory due to the risk
of metastasis, recurrence and chemoresistance. Therefore,
alternative therapeutic strategies are urgently needed to improve
the efficacy and reduce the side-effects. Targeted therapies that
can identify targetable aberrations (specific genes or molecular
pathways) and can manipulate apoptosis and autophagy have become a
promising approach in OS treatment (18,19).
mTOR and Akt signaling have been reported to be
involved with the metastatic behavior of OS, and the most common
actionable aberrations found in the PI3K/Akt/mTOR pathway (19,20).
In the present review, we focused on the association of the mTOR
signaling pathway in the pathogenesis of OS, and the possible
effective treatment strategies by targeting this pathway.
2. The mTOR complex
mTOR is the catalytic subunit of two structurally
and functionally distinct multiprotein complexes: mTOR complex 1
(mTORC1) and mTOR complex 2 (mTORC2) (21). mTORC1 is sensitive to the effects of
rapamycin and is composed of mTOR itself, regulatory-associated
protein of mTOR (Raptor) and the non-core components of 40 kDa
pro-rich AKT substrate (PRAS40; also known as AKT1S1) (22). Raptor serves as the binding platform
where its association with mTOR forms a nutrient-sensitive complex
that signals to regulate cell growth (4,23).
Conversely, PRAS40 regulates mTORC1 kinase activity by functioning
as a direct inhibitor of mTORC1 substrate binding (24).
mTORC2 is composed of mTOR, rapamycin-insensitive
companion of mTOR (RICTOR), mammalian stress-activated map
kinase-interacting protein 1 (mSIN1; also known as MAPKAP1) and
protein observed with RICTOR (PROTOR). RICTOR is known as a
scaffold protein that contributes to the structural foundation of
mTORC2, and plays an essential role in embryonic growth and
development. However, the functions of mSIN1 and PROTOR are not
clear (8,25). Mammalian lethal with SEC13 protein 8
[mLST8; also known as G protein β subunit-like (GβL)] and DEP
domain-containing mTOR-interacting protein (DEPTOR) are the two
other proteins common in both complexes (26). The function of mLST8 within mTORC1
is not clear. However, it is essential for the activation of mTORC2
that regulates growth and development. DEPTOR functions as an
inhibitor of mTOR signaling that inhibits mTORC1 and mTORC2
activity by preventing substrate binding to mTORC1 and mTORC2
(27,28).
The activity of mTORC1 is stimulated by growth
factors, insulin, phosphatidic and amino acid (particularly
leucine) levels, energy status and oxidative stress (4,29).
Insulin receptor substrate (IRS) activates PI3K through stimulation
of growth factors. PI3K generates phosphatidylinositol
3,4,5-triphosphate (PIP3) upon phosphorylation. PIP3 then promotes
the phosphorylation of protein kinase (PKB/AKT) by
3-phosphoinositide-dependent protein kinase-1 (PDK1). AKT
phosphorylates tuberous sclerosis complex (TSC) which ultimately
leads to the activation of mTORC1 (22,30).
Activated mTORC1 phosphorylates its two downstream effectors, p70
ribosomal S6 kinase 1 (p70S6K1 or S6K1) and the eukaryotic
initiation factor 4E (eIF4E) binding protein 1 (4E-BP1), which are
thought to be the major regulators of protein translation, cell
proliferation, angiogenesis and autophagy. Autophagy is an
evolutionarily conserved, self-degradation system of cellular
components through an autophagosomal-lysosomal pathway dependent on
nutrient conditions and is reported to be deregulated in cancer.
During starvation, mTORC1 is inactivated and dissociates from the
ULK1 kinase complex (a protein complex composed of Atg1, Atg13,
Atg17 and Atg101) to induce autophagy (6,31,32).
Akt is the major substrate of mTORC2 and the full
activation of Akt requires two steps of phosphorylation: firstly,
PDK1-mediated phosphorylation at a threonine T308 residue and
secondly, mTORC2-mediated phosphorylation at a serine S473 residue.
The mTORC2 is activated by growth factors and functions as an
important regulator for the organization of the actin cytoskeleton,
cell survival and lipid metabolism (8,33).
3. Association of mTOR signaling with
OS
mTOR is a critical signaling pathway that is
potentially activated in OS. However, the mechanism by which the
mTOR signaling pathway is activated in OS is poorly understood.
Evidence shows that multiple elements of mTOR signaling are
involved in OS (Fig. 1). The
upstream regulators (for example, PI3K) of mTOR signaling are
deregulated through the overexpression of growth factor receptors,
such as human epidermal growth factor receptor 2 (HER-2) and
insulin-like growth factor receptor (IGFR), mutations in
PI3K and amplifications of AKT (34–36).
PTEN, the negative regulator of PI3K signaling is downregulated in
OS and contributes to bone proliferation, angiogenesis and
metastasis through several mechanisms, including mutation,
methylation or protein instability (34,37).
The downstream effectors of mTOR (such as, S6K1, 4EBP1 and eIF4E)
are overexpressed in OS and implicated in cellular transformation
and poor cancer prognosis (38).
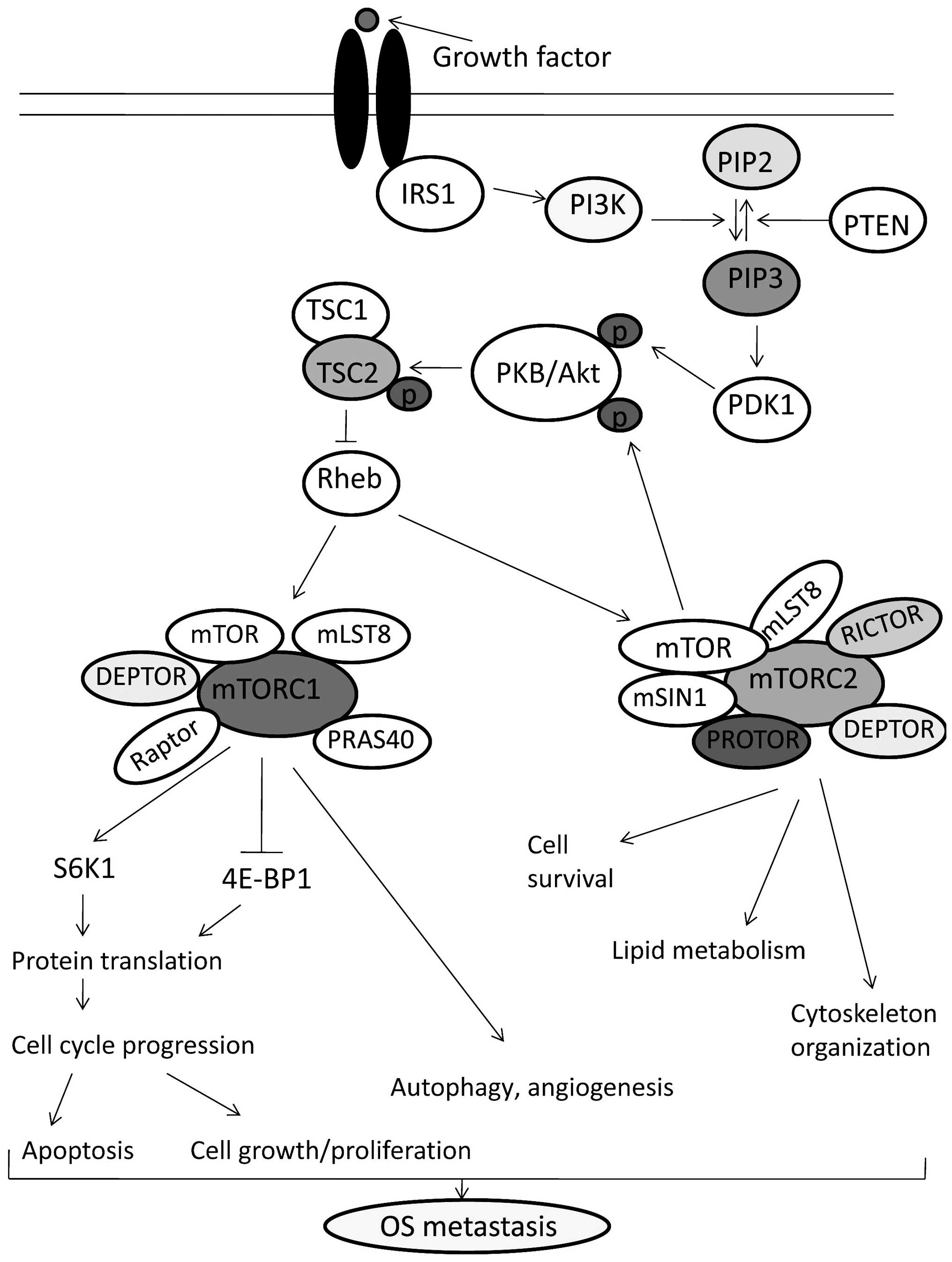 | Figure 1mTOR signaling pathway and its
association with osteosarcoma. Upon growth factor stimulation,
insulin receptor substrate (IRS) activates PI3K and PI3K generates
PIP3, which in turn activates PDK1 and Akt. After phosphorylation
by PDK1, Akt phosphorylates and inhibits the TSC complex which
ultimately leads to the activation of mTORC1 (22,30).
Activated mTORC1 phosphorylates S6K1 and 4E-BP1, which thereby
regulate protein translation, cell proliferation, angiogenesis and
autophagy (4,32). mTORC2 activates Akt through the two
steps of phosphorylation and functions as an important regulator
for the organization of the actin cytoskeleton, cell survival and
lipid metabolism (8,33). The oncogenic activation of the
components of the mTOR pathway through the overexpression of growth
factor receptors, mutations in PI3K, loss of PTEN and
amplifications of AKT contributes to bone proliferation,
angiogenesis, autophagy and inhibits apoptosis leading to OS
metastasis (34). |
Ezrin, a member of the ezrin/radixin/moesin (ERM)
family of proteins, is involved in intracellular signal
transduction (39). It is reported
that ezrin expression increases phosphorylation and expression of
S6K1 and 4E-BP-1 as well as S6K1 activity, which may induce
ezrin-mediated metastatic behavior in OS (40). Zhou et al investigated the
expression of mTOR and p70S6K in 65 patients with primary OS, and
found that overexpression both of mTOR and p70S6K are significantly
correlated with OS cell proliferation, survival and metastasis in
OS patients (38). Tenascin-C
(TN-C) is one of extracellular matrix glycoproteins found to be
overexpressed in OS. Tanaka et al reported that the
overexpression of TN-C has an effect of easier migration of OS
cells to promote distant metastases (41). Zheng et al reported that TN-C
in combination with its alternative spliced FNIII repeats with A1
subdomain (TN-C FNIIIA1) is overexpressed in MG-63 OS cells and
promote cellular migration. Their study also found that the
downstream molecules of mTOR such as, 4E-BP1 and S6K1 can
facilitate the expression of TN-C FNIIIA1 in MG-63 cells and
promote the metastasis of OS by facilitating MG-63 cell migration
(42).
4. Targeting the mTOR complex in
osteosarcoma
Conventional therapy for OS remains unsatisfactory
due to the resistance of OS to chemotherapy and radiotherapy. New
alternative therapies and therapeutic targets are needed to be
identified for the development of more effective target-specific
anti-OS agents for the treatment of OS. Targeting the mTOR
signaling pathway may represent an attractive potential therapeutic
approach for the prevention and treatment of OS (43,44).
Table I summarizes the potential
therapeutic strategies and their effects on OS by targeting the
mTOR signaling pathway.
 | Table IPotential therapeutic strategies of
OS by targeting the mTOR pathway. |
Table I
Potential therapeutic strategies of
OS by targeting the mTOR pathway.
| Therapeutic
agents | Experimental OS
samples | Effects | (Refs.) |
|---|
| Rapamycin | Murine model of OS
(K7M2) | Inhibited
ezrin-related metastatic behavior | (40) |
| Rapamycin | Human OS cells | Inhibited OS cell
proliferation, suppressed tumor growth, induced autophagy and cell
cycle arrest in the G1 phase | (43) |
| Rapamycin or
CDDP | MG63 OS cells | Enhanced the
effects of the activation of autophagy and induced apoptosis | (50) |
| Kinase inhibitor
(PP242) or rictor knockdown by siRNA | OS cell lines
(MG63/U2OS/SaOS-2) | Prevented cell
migration and induced apoptosis | (47) |
| NVP-BEZ235 | OS cell lines
(MG63, U2OS and SaOS-2) | Induced apoptosis
and cell cycle arrest, slowed tumor progression and reduced tumor
vasculature | (52,53) |
| Alisertib | U2OS and MG63 OS
cells | Promoted autophagy
and induced apoptosis | (54) |
| Sorafenib plus
everolimus | OS cell lines
(MNNG-HOS, HOS, KHOS/NP, MG63, U2OS, SJSA-1 and SaOS-2), patients
with unresectable high-grade OS | Enhanced
antiproliferative and proapoptotic effects impaired tumor growth,
potentiated antiangiogenesis, reduced migratory and metastatic
potential | (20,56) |
| Ridaforolimus | Patients with
OS | Achieved confirmed
partial response | (57) |
| Lupeol | MNNG-HOS and MG63
OS cells | Decreased tumor
growth, induced apoptosis and cell cycle arrest | (58) |
| FIM-A | Human OS cells | Inhibited cell
growth and proliferation and reduced angiogenesis | (59) |
| Oleanolic acid | MG63 and SaOS-2
cells | Exhibited antitumor
activity | (60) |
| Geldanamycin plus
3-methyladenine | KTHOS cells | Induced autophagy
and apoptosis | (18) |
| MicroRNA-223 | MG63 cells | Inhibited cell
growth, exhibited significant G0/G1 arrest and increased
apoptosis | (61) |
| MicroRNA-101 | OS tissues and
SaOS-2 cells | Inhibited cell
proliferation and promoted apoptosis | (62) |
| Perifosine | Human OS cells | Induced cell
apoptosis and growth inhibition | (63) |
| RAD001 plus
zoledronate | Human and mouse OS
cells | Inhibited cell
proliferation and abolished drug resistance | (64) |
Perry et al used several methods to identify
genomic events contributing to OS, and suggested that targeting the
PI3K/mTOR pathway has central vulnerability for therapeutic
exploitation in OS (34). Rapamycin
itself is the inhibitor of the mTOR signaling pathway, and may be a
promising agent against OS. Zhao et al evaluated the effects
of rapamycin on human OS cells and reported that rapamycin
increased the expression of p27 and decreased the expression of
cyclin D1, inhibited OS cell proliferation, induced autophagy and
cell cycle arrest in the G1 phase. Their results also reported that
rapamycin suppressed the tumor growth in mouse xenograft models
(43). The use of mTOR inhibitors
blunted the p53 response to nucleolar stress by regulating ribosome
biogenesis and mRNA translation (45). p53 potently inhibited cell
proliferation, metastasis, and angiogenesis in OS cells through the
inhibition of the PI3K/AKT/mTOR pathway (46). Wang et al reported that
targeting of mTORC2 either by a kinase inhibitor or rictor
knockdown prevented OS cell migration and promoted
cisplatin-induced apoptosis (47).
Although mTOR is a novel promising oncological
target, unfortunately mTOR-targeted monotherapies have shown only
modest antitumor activity in OS. The combination with other
rationally selected therapeutic agents may improve the response
(48). Wagner et al
evaluated the activity of the combined inhibition of cixutumumab
[inhibitor of insulin-growth factor type 1 receptor (IGF-1R)] with
the mTOR inhibitor temsirolimus in preclinical models of OS and
other sarcoma patients. In the present study, 43 evaluable patients
received 6 mg/kg cixutumumab with 8 mg/m2 temsirolimus
intravenously once weekly in 4-week cycles and observed that 16% of
patients were progression-free at 12 weeks. The reported adverse
effects were only mucositis, electrolyte disturbances and
myelosuppression (49). In another
study, Xie et al reported that rapamycin enhanced the
effects of the activation of autophagy and the induction of
apoptosis of OS cells by cis-diamminedichloroplatinum
(cisplatin) (50). In another
study, Wan et al found that blockade of the mTOR pathway
with rapamycin inhibited ezrin-related metastatic behavior in a
murine model of OS (40). In a
recent study, the combination of rapamycin and an autophagy
inhibitor, specific and potent autophagy inhibitor-1 (spautin-1)
was found to effectively induce the apoptosis pathway, and was
suggested as a possible treatment option in OS (51).
Systematic screening has identified that dual
inhibition of the PI3K-mTOR pathway is a sensitive, druggable
target in OS (52). Zhu et
al studied the therapeutic potential of NVP-BEZ235, a novel
PI3K/mTOR dual inhibitor, on OS cells, and found that NVP-BEZ235
downregulated cyclin D1/B1 expression, induced apoptosis and cell
cycle arrest in the G0/G1 phase through inhibition of
PI3K-AKT-mTORC1 signaling in OS cells. An in vivo study also
showed that oral administration of NVP-BEZ235 inhibited OS
xenograft growth in SCID mice (53). In a murine pre-clinical model, Gobin
et al reported that NVP-BEZ235 significantly reduced tumor
progression, ectopic tumor bone formation and reduced tumor
vasculature (54). Alisertib (ALS;
MLN8237) is a selective aurora kinase-A inhibitor that displays
potent growth inhibitory, pro-apoptotic, pro-autophagic and EMT
inhibitory effects on OS cells. Niu et al reported that ALS
markedly downregulated the expression levels of cyclin D1/2 and B1,
induced G2/M arrest, suppressed EMT-like phenotypes as well as
promoted apoptosis and autophagy via inhibition of the
PI3K/Akt/mTOR signaling pathway in OS cells (55). MLN0128, an ATP-competitive mTOR
kinase inhibitor was reported to inhibit mTORC1/2 targets and show
potent anti-tumor activity in multiple sarcoma subtypes including
OS (56).
Treatment with the multikinase inhibitor sorafenib
showed disease stabilization and reduced drug resistance in
patients with unresectable advanced and metastatic OS (20,57).
Pignochino et al investigated the activity of sorafenib in
combination with everolimus in preclinical models of OS to overcome
the drawbacks. Their results observed enhanced anti-proliferative
and proapoptotic effects, impaired tumor growth, potentiated
antiangiogenesis as well as reduced migratory and metastatic
potential by complete inhibition of the mTOR pathway (20). A phase II clinical trial (ClinicalTrials.gov, no. NCT01804374) was conducted, in
which 38 patients with unresectable high-grade OS were enrolled.
Patients received 800 mg sorafenib plus 5 mg everolimus once a day
until disease progression and observed progression-free survival at
6 months in 17 patients. The most common clinical adverse events
were lymphopenia and hypophosphataemia, oral mucositis, diarrhoea,
hand and foot syndrome, thrombocytopenia, fatigue and anaemia. No
treatment-related death was reported in the present study (57). Another phase II trial was conducted
to assess the antitumor activity of ridaforolimus (an inhibitor of
mTOR) in patients with distinct subtypes of advanced bone and soft
tissue sarcomas. In this trial, a total of 212 patients were
enrolled and ridaforolimus (12.5 mg) was administered as a 30-min
intravenous infusion once daily for 5 days every 2 weeks. Two
patients with OS achieved confirmed partial response. Related
adverse events were stomatitis, mouth ulceration, mucosal
inflammation and fatigue (58).
Liu et al investigated the anticancer
activity of lupeol (a dietary triterpene present in many fruits and
medicinal plants) in human OS cells. Their results showed that
lupeol promoted downregulation of the protein expression levels of
PI3K, AKT, p70S6K and cyclin D1 and upregulation of the expression
levels of p21 and p27, which play a pivotal role in the regulation
of apoptosis and cell cycle arrest in G0/G1 phase of OS cells in
vitro. They also established tumor xenografts in female nude
BALB/c mice, and administered lupeol intravenously and found that
administration of lupeol decreased tumor growth, induced apoptosis
as well as cell cycle arrest of human OS cells through the
PI3K/AKT/mTOR signaling pathway (59). Phosphorus-containing sirolimus
(FIM-A) inhibited cancer cell growth and proliferation, reduced
angiogenesis, induced cell cycle arrest in the G1 phase of OS cells
by targeting mTOR. In in vivo mouse osteosarcoma xenografts,
FIM-A significantly decreased phosphorylation of p70S6K1 and
4E-BP1, and decreased the average tumor volume and the number of
intratumoral microvessels through the imhibition of mTORC1
signaling (60). Zhou et al
examined the anticancer activities of oleanolic acid (OA; a
pentacyclic triterpenoid) in OS cells, and found that OA inhibited
cell proliferation, induced G1 arrest in OS cells and regulated
protein translation through the inhibition of mTOR signaling
(61).
Heat shock protein 90 (Hsp90) plays a critically
important role in tumor cell growth, survival and autophagy. Mori
et al examined the effects of the Hsp90 inhibitor,
geldanamycin (GA) on OS cells and found that GA induced autophagy
and apoptosis in OS cells through inhibition of the Akt/mTOR
signaling pathway. They also reported that the combination of GA
and the autophagy inhibitor 3-methyladenine (3-MA) may be an
effective treatment for OS as this combination effectively
suppressed a protective mechanism induced by the Hsp90 inhibitor
and enhanced GA-induced apoptosis in OS cells (18). Hsp90B1 plays an oncogenic role and
is a target gene of miR-223. Overexpression of miR-223
downregulated Hsp90B1 and showed tumor-suppressor function in OS
through the inhibition of the PI3K/Akt/mTOR pathway (62). miR-101 functions as a tumor
suppressor, and is reported to be downregulated in OS tissues and
cell lines. mTOR gene is a direct target of miR-101 and
reintroduction of miR-101 in OS cell line inhibited the
proliferation and promoted apoptosis by downregulating mTOR
expression (63). Perifosine (Akt
inhibitor) is another possible anti-OS agent that induced cell
apoptosis and growth inhibition in human OS cells. Perifosine
promoted caspase-3, c-Jun N-terminal kinases (JNK) and p53
activation and inhibited survivin expression by disrupting its
association with HSP-90 through inhibition of Akt/mTORC1 signaling
(64). Moriceau et al
investigated the effects of RAD001 (everolimus, a new orally
available mTOR inhibitor) on the growth of human and mouse OS cells
in combination with zoledronate (ZOL; an anti-osteoporotic drug).
They found that the combination of RAD001 with ZOL augmented the
inhibition of cell proliferation and abolished the resistance of OS
cells to RAD001 through inhibition of PI3K/mTOR signaling (65). A recent interesting finding showed
that dual mTORC1/2 inhibition by the potent mTOR kinase inhibitor
INK-128 (MLN0128) exerted anti-OS activity in vitro and
in vivo (66). However,
there is not yet enough evidence. Yet, INK-128 may be further
investigated as a novel anti-OS agent.
5. Future direction
mTOR is a master regulator of cell growth,
proliferation, survival and autophagy through its ability to
stimulate mRNA translation, ribosome biogenesis and protein
stability. Oncogenic activation of mTOR signaling plays a crucial
role in the development of OS. A better understanding of the
functions of the mTOR pathway and the molecular mechanism by which
it promotes OS may provide the basis to develop a potential
therapeutic target for the treatment of OS. mTOR is a new promising
oncological target and blockade of the mTOR pathway with selective
inhibitors represents an attractive and potential target for the
treatment of OS. Development of the optimal dose, regimen and a
rationale for the use of mTOR inhibitors in combination with other
agents may provide a promising therapy for the treatment of OS and
could improve the survival rate of OS patients.
References
|
1
|
Vézina C, Kudelski A and Sehgal SN:
Rapamycin (AY-22,989), a new antifungal antibiotic. I Taxonomy of
the producing streptomycete and isolation of the active principle.
J Antibiot. 28:721–726. 1975. View Article : Google Scholar
|
|
2
|
Wullschleger S, Loewith R and Hall MN: TOR
signaling in growth and metabolism. Cell. 124:471–484. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Benjamin D, Colombi M, Moroni C and Hall
MN: Rapamycin passes the torch: A new generation of mTOR
inhibitors. Nat Rev Drug Discov. 10:868–880. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kim DH, Sarbassov DD, Ali SM, King JE,
Latek RR, Erdjument-Bromage H, Tempst P and Sabatini DM: mTOR
interacts with raptor to form a nutrient-sensitive complex that
signals to the cell growth machinery. Cell. 110:163–175. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Laplante M and Sabatini DM: mTOR signaling
in growth control and disease. Cell. 149:274–293. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hay N and Sonenberg N: Upstream and
downstream of mTOR. Genes Dev. 18:1926–1945. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hoeffer CA and Klann E: mTOR signaling: At
the crossroads of plasticity, memory and disease. Trends Neurosci.
33:67–75. 2010. View Article : Google Scholar
|
|
8
|
Zoncu R, Efeyan A and Sabatini DM: mTOR:
From growth signal integration to cancer, diabetes and ageing. Nat
Rev Mol Cell Biol. 12:21–35. 2011. View
Article : Google Scholar
|
|
9
|
Guertin DA and Sabatini DM: An expanding
role for mTOR in cancer. Trends Mol Med. 11:353–361. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu K, Liu P and Wei W: mTOR signaling in
tumorigenesis. Biochim Biophys Acta. 1846:638–654. 2014.PubMed/NCBI
|
|
11
|
Beauchamp EM and Platanias LC: The
evolution of the TOR pathway and its role in cancer. Oncogene.
32:3923–3932. 2013. View Article : Google Scholar
|
|
12
|
Fasolo A and Sessa C: Targeting mTOR
pathways in human malignancies. Curr Pharm Des. 18:2766–2777. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Luetke A, Meyers PA, Lewis I and Juergens
H: Osteosarcoma treatment - where do we stand? A state of the art
review Cancer Treat Rev. 40:523–532. 2014. View Article : Google Scholar
|
|
14
|
Bielack S and Carrle D: Osteosarcoma: ESMO
clinical recommendations for diagnosis, treatment and follow-up.
Ann Oncol. 20(Suppl 4): S137–S139. 2009. View Article : Google Scholar
|
|
15
|
Ottaviani G and Jaffe N: The epidemiology
of osteosarcoma. Cancer Treat Res. 152:3–13. 2009. View Article : Google Scholar
|
|
16
|
Ottaviani G and Jaffe N: The etiology of
osteosarcoma. Cancer Treat Res. 152:15–32. 2009. View Article : Google Scholar
|
|
17
|
Anderson PM and Pearson M: Novel
therapeutic approaches in pediatric and young adult sarcomas. Curr
Oncol Rep. 8:310–315. 2006. View Article : Google Scholar
|
|
18
|
Mori M, Hitora T, Nakamura O, Yamagami Y,
Horie R, Nishimura H and Yamamoto T: Hsp90 inhibitor induces
autophagy and apoptosis in osteosarcoma cells. Int J Oncol.
46:47–54. 2015.
|
|
19
|
Egas-Bejar D, Anderson PM, Agarwal R,
Corrales-Medina F, Devarajan E, Huh WW, Brown RE and Subbiah V:
Theranostic profiling for actionable aberrations in advanced high
risk osteosarcoma with aggressive biology reveals high molecular
diversity: The human fingerprint hypothesis. Oncoscience.
1:167–179. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pignochino Y, Dell'Aglio C, Basiricò M,
Capozzi F, Soster M, Marchiò S, Bruno S, Gammaitoni L, Sangiolo D,
Torchiaro E, et al: The combination of sorafenib and everolimus
abrogates mTORC1 and mTORC2 upregulation in osteosarcoma
preclinical models. Clin Cancer Res. 19:2117–2131. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Betz C and Hall MN: Where is mTOR and what
is it doing there? J Cell Biol. 203:563–574. 2013. View Article : Google Scholar :
|
|
22
|
Ashworth RE and Wu J: Mammalian target of
rapamycin inhibition in hepatocellular carcinoma. World J Hepatol.
6:776–782. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hara K, Maruki Y, Long X, Yoshino K,
Oshiro N, Hidayat S, Tokunaga C, Avruch J and Yonezawa K: Raptor, a
binding partner of target of rapamycin (TOR), mediates TOR action.
Cell. 110:177–189. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang L, Harris TE, Roth RA and Lawrence JC
Jr: PRAS40 regulates mTORC1 kinase activity by functioning as a
direct inhibitor of substrate binding. J Biol Chem.
282:20036–20044. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mendoza MC, Er EE and Blenis J: The
Ras-ERK and PI3K-mTOR pathways: Cross-talk and compensation. Trends
Biochem Sci. 36:320–328. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guertin DA, Stevens DM, Thoreen CC, Burds
AA, Kalaany NY, Moffat J, Brown M, Fitzgerald KJ and Sabatini DM:
Ablation in mice of the mTORC components raptor, rictor, or mLST8
reveals that mTORC2 is required for signaling to Akt-FOXO and
PKCalpha, but not S6K1. Dev Cell. 11:859–871. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Peterson TR, Laplante M, Thoreen CC,
Sancak Y, Kang SA, Kuehl WM, Gray NS and Sabatini DM: DEPTOR is an
mTOR inhibitor frequently overexpressed in multiple myeloma cells
and required for their survival. Cell. 137:873–886. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Laplante M and Sabatini DM: mTOR signaling
at a glance. J Cell Sci. 122:3589–3594. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fang Y, Vilella-Bach M, Bachmann R,
Flanigan A and Chen J: Phosphatidic acid-mediated mitogenic
activation of mTOR signaling. Science. 294:1942–1945. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sengupta S, Peterson TR and Sabatini DM:
Regulation of the mTOR complex 1 pathway by nutrients, growth
factors, and stress. Mol Cell. 40:310–322. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Finn RS: Current and future treatment
strategies for patients with advanced hepatocellular carcinoma:
Role of mTOR inhibition. Liver Cancer. 1:247–256. 2012. View Article : Google Scholar
|
|
32
|
Pyo JO, Nah J and Jung YK: Molecules and
their functions in autophagy. Exp Mol Med. 44:73–80. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bayascas JR and Alessi DR: Regulation of
Akt/PKB Ser473 phosphorylation. Mol Cell. 18:143–145. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Perry JA, Kiezun A, Tonzi P, Van Allen EM,
Carter SL, Baca SC, Cowley GS, Bhatt AS, Rheinbay E, Pedamallu CS,
et al: Complementary genomic approaches highlight the PI3K/mTOR
pathway as a common vulnerability in osteosarcoma. Proc Natl Acad
Sci USA. 111:E5564–E5573. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kuijjer ML, van den Akker BE, Hilhorst R,
Mommersteeg M, Buddingh EP, Serra M, Bürger H, Hogendoorn PC and
Cleton-Jansen AM: Kinome and mRNA expression profiling of
high-grade osteosarcoma cell lines implies Akt signaling as
possible target for therapy. BMC Med Genomics. 7:42014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pópulo H, Lopes JM and Soares P: The mTOR
signalling pathway in human cancer. Int J Mol Sci. 13:1886–1918.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Freeman SS, Allen SW, Ganti R, Wu J, Ma J,
Su X, Neale G, Dome JS, Daw NC and Khoury JD: Copy number gains in
EGFR and copy number losses in PTEN are common events in
osteosarcoma tumors. Cancer. 113:1453–1461. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhou Q, Deng Z, Zhu Y, Long H, Zhang S and
Zhao J: mTOR/p70S6K signal transduction pathway contributes to
osteosarcoma progression and patients' prognosis. Med Oncol.
27:1239–1245. 2010. View Article : Google Scholar
|
|
39
|
Di Cristofano C, Leopizzi M, Miraglia A,
Sardella B, Moretti V, Ferrara A, Petrozza V and Della Rocca C:
Phosphorylated ezrin is located in the nucleus of the osteosarcoma
cell. Mod Pathol. 23:1012–1020. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wan X, Mendoza A, Khanna C and Helman LJ:
Rapamycin inhibits ezrin-mediated metastatic behavior in a murine
model of osteosarcoma. Cancer Res. 65:2406–2411. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tanaka M, Yamazaki T, Araki N, Yoshikawa
H, Yoshida T, Sakakura T and Uchida A: Clinical significance of
tenascin-C expression in osteosarcoma: Tenascin-C promotes distant
metastases of osteosarcoma. Int J Mol Med. 5:505–510.
2000.PubMed/NCBI
|
|
42
|
Zheng L, Zhang D, Zhang Y, Wen Y and Wang
Y: mTOR signal transduction pathways contribute to TN-C FNIII A1
overexpression by mechanical stress in osteosarcoma cells. Mol
Cells. 37:118–125. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhao S, Lu N, Chai Y and Yu X: Rapamycin
inhibits tumor growth of human osteosarcomas. J BUON. 20:588–594.
2015.PubMed/NCBI
|
|
44
|
Zhang J, Yu XH, Yan YG, Wang C and Wang
WJ: PI3K/Akt signaling in osteosarcoma. Clin Chim Acta.
444:182–192. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Goudarzi KM, Nistér M and Lindström MS:
mTOR inhibitors blunt the p53 response to nucleolar stress by
regulating RPL11 and MDM2 levels. Cancer Biol Ther. 15:1499–1514.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Song R, Tian K, Wang W and Wang L: P53
suppresses cell proliferation, metastasis, and angiogenesis of
osteosarcoma through inhibition of the PI3K/AKT/mTOR pathway. Int J
Surg. 20:80–87. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang X, Lai P, Zhang Z, Huang M, Wang L,
Yin M, Jin D, Zhou R and Bai X: Targeted inhibition of mTORC2
prevents osteosarcoma cell migration and promotes apoptosis. Oncol
Rep. 32:382–388. 2014.PubMed/NCBI
|
|
48
|
Fleuren ED, Versleijen-Jonkers YM, Roeffen
MH, Franssen GM, Flucke UE, Houghton PJ, Oyen WJ, Boerman OC and
van der Graaf WT: Temsirolimus combined with cisplatin or
bevacizumab is active in osteosarcoma models. Int J Cancer.
135:2770–2782. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wagner LM, Fouladi M, Ahmed A, Krailo MD,
Weigel B, DuBois SG, Doyle LA, Chen H and Blaney SM: Phase II study
of cixutumumab in combination with temsirolimus in pediatric
patients and young adults with recurrent or refractory sarcoma: A
report from the Children's Oncology Group. Pediatr Blood Cancer.
62:440–444. 2015. View Article : Google Scholar
|
|
50
|
Xie ZG, Xie Y and Dong QR: Inhibition of
the mammalian target of rapamycin leads to autophagy activation and
cell death of MG63 osteosarcoma cells. Oncol Lett. 6:1465–1469.
2013.PubMed/NCBI
|
|
51
|
Horie R, Nakamura O, Yamagami Y, Mori M,
Nishimura H, Fukuoka N and Yamamoto T: Apoptosis and antitumor
effects induced by the combination of an mTOR inhibitor and an
autophagy inhibitor in human osteosarcoma MG63 cells. Int J Oncol.
48:37–44. 2016.
|
|
52
|
Gupte A, Baker EK, Wan SS, Stewart E, Loh
A, Shelat AA, Gould CM, Chalk AM, Taylor S, Lackovic K, et al:
Systematic Screening identifies dual PI3K and mTOR inhibition as a
conserved therapeutic vulnerability in osteosarcoma. Clin Cancer
Res. 21:3216–3229. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhu YR, Min H, Fang JF, Zhou F, Deng XW
and Zhang YQ: Activity of the novel dual phosphatidylinositol
3-kinase/mammalian target of rapamycin inhibitor NVP-BEZ235 against
osteosarcoma. Cancer Biol Ther. 16:602–609. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Gobin B, Battaglia S, Lanel R, Chesneau J,
Amiaud J, Rédini F, Ory B and Heymann D: NVP-BEZ235, a dual
PI3K/mTOR inhibitor, inhibits osteosarcoma cell proliferation and
tumor development in vivo with an improved survival rate. Cancer
Lett. 344:291–298. 2014. View Article : Google Scholar
|
|
55
|
Niu NK, Wang ZL, Pan ST, Ding HQ, Au GH,
He ZX, Zhou ZW, Xiao G, Yang YX, Zhang X, et al: Pro-apoptotic and
pro-autophagic effects of the Aurora kinase A inhibitor alisertib
(MLN8237) on human osteosarcoma U-2 OS and MG-63 cells through the
activation of mitochondria-mediated pathway and inhibition of p38
MAPK/PI3K/Akt/mTOR signaling pathway. Drug Des Devel Ther.
9:1555–1584. 2015.PubMed/NCBI
|
|
56
|
Slotkin EK, Patwardhan PP, Vasudeva SD, de
Stanchina E, Tap WD and Schwartz GK: MLN0128, an ATP-competitive
mTOR kinase inhibitor with potent in vitro and in vivo antitumor
activity, as potential therapy for bone and soft-tissue sarcoma.
Mol Cancer Ther. 14:395–406. 2015. View Article : Google Scholar
|
|
57
|
Grignani G, Palmerini E, Ferraresi V,
D'Ambrosio L, Bertulli R, Asaftei SD, Tamburini A, Pignochino Y,
Sangiolo D, Marchesi E, et al Italian Sarcoma Group: Sorafenib and
everolimus for patients with unresectable high-grade osteosarcoma
progressing after standard treatment: A non-randomised phase 2
clinical trial. Lancet Oncol. 16:98–107. 2015. View Article : Google Scholar
|
|
58
|
Chawla SP, Staddon AP, Baker LH, Schuetze
SM, Tolcher AW, D'Amato GZ, Blay JY, Mita MM, Sankhala KK, Berk L,
et al: Phase II study of the mammalian target of rapamycin
inhibitor ridaforolimus in patients with advanced bone and soft
tissue sarcomas. J Clin Oncol. 30:78–84. 2012. View Article : Google Scholar
|
|
59
|
Liu Y, Bi T, Dai W, Wang G, Qian L, Shen G
and Gao Q: Lupeol induces apoptosis and cell cycle arrest of human
osteosarcoma cells through PI3K/AKT/mTOR pathway. Technol Cancer
Res Treat. Oct 6–2015.(Epub ahead of print). pii:
1533034615609014.
|
|
60
|
Liu WN, Lin JH, Cheng YR, Zhang L, Huang
J, Wu ZY, Wang FS, Xu SG, Lin WP, Lan WB, et al: FIM-A, a
phosphorus-containing sirolimus, inhibits the angiogenesis and
proliferation of osteosarcomas. Oncol Res. 20:319–326. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zhou R, Zhang Z, Zhao L, Jia C, Xu S, Mai
Q, Lu M, Huang M, Wang L, Wang X, et al: Inhibition of mTOR
signaling by oleanolic acid contributes to its anti-tumor activity
in osteosarcoma cells. J Orthop Res. 29:846–852. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Li G, Cai M, Fu D, Chen K, Sun M, Cai Z
and Cheng B: Heat shock protein 90B1 plays an oncogenic role and is
a target of microRNA-223 in human osteosarcoma. Cell Physiol
Biochem. 30:1481–1490. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Lin S, Shao NN, Fan L, Ma XC, Pu FF and
Shao ZW: Effect of microRNA-101 on proliferation and apoptosis of
human osteosarcoma cells by targeting mTOR. J Huazhong Univ Sci
Technolog Med Sci. 34:889–895. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Yao C, Wei JJ, Wang ZY, Ding HM, Li D, Yan
SC, Yang YJ and Gu ZP: Perifosine induces cell apoptosis in human
osteosarcoma cells: New implication for osteosarcoma therapy? Cell
Biochem Biophys. 65:217–227. 2013. View Article : Google Scholar
|
|
65
|
Moriceau G, Ory B, Mitrofan L, Riganti C,
Blanchard F, Brion R, Charrier C, Battaglia S, Pilet P, Denis MG,
et al: Zoledronic acid potentiates mTOR inhibition and abolishes
the resistance of osteosarcoma cells to RAD001 (Everolimus):
Pivotal role of the prenylation process. Cancer Res.
70:10329–10339. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Jiang H and Zeng Z: Dual mTORC1/2
inhibition by INK-128 results in antitumor activity in preclinical
models of osteosarcoma. Biochem Biophys Res Commun. 468:255–261.
2015. View Article : Google Scholar : PubMed/NCBI
|














