Introduction
Colorectal cancer is the third most common cancer in
males and the second in females, and is thus a severe health threat
worldwide (1). Several factors
including genetics, gender, ethnic origin, geographical region, and
environmental conditions influence the incidence of colorectal
cancer (2). Moreover, lifestyle
factors such as consumption of processed meat products, high-fat
and low-fiber diet, lack of physical activity, and obesity increase
the risk of this disease (3,4). Key
obstacles to long-term survival of colorectal cancer patients
include resistance of the tumors to chemotherapeutic agents and the
side-effects of prolonged chemotherapeutic treatments (5).
Non-thermal gas plasma (NTGP), a novel tool
successfully used in wound healing and surface sterilization,
promotes cell proliferation and increases transfection efficiency
(6,7). In addition, this method has recently
emerged as a promising approach for treating cancer. The anticancer
activity of NTGP has been demonstrated in both in vivo and
in vitro models including skin, liver, lung, and colon
cancers (8,9). Plasma, considered to be the fourth
stage of matter, consists of charged particles (electrons, ions),
excited atoms, and reactive oxygen species (ROS) (10). Several lines of evidence suggest
that plasma ROS induce apoptosis via oxidative stress (9). Induction of selective cell death in
colorectal cancer cells by NTGP treatment represents a promising
approach to colorectal cancer therapy that would both avoid the
deleterious side-effects of chemotherapy and circumvent
chemoresistance.
Cells can undergo apoptosis via three different
pathways, respectively mediated by death receptors, mitochondria,
or the endoplasmic reticulum (ER) (11–13).
The ER is the primary site for synthesis and folding of secreted
and membrane-bound proteins, as well as some organelle-targeted
proteins. This organelle is highly sensitive to stresses that
perturb cellular energy level, redox state, or Ca2+
concentration (14). Protein
chaperones such as glucose-related protein 78 (GRP78)/BiP and GRP94
maintain the correct folding of newly synthesized proteins in the
ER (15). High ROS levels disturb
ER function, leading to accumulation of unfolded proteins and a
state referred to as ER stress. In response to ER stress, the cell
activates signaling pathways including the unfolded protein
response (UPR) and ER-associated protein degradation (ERAD)
(16,17). The UPR, the primary defense
mechanism of the ER, restores cellular function by halting protein
synthesis and bolstering protein folding capacity, thereby
improving the cell's likelihood of survival (17). However, when ER stress is so severe
that these tactics cannot restore cellular homeostasis, the UPR
triggers apoptosis (16).
Because plasma generates ROS, resulting in oxidative
stress, we predicted that NTGP would induce ER stress in SNUC5
human colon cancer cells and thereby cause apoptosis. The effects
of the plasma exposure depend on the plasma source (e.g., plasma
jet or needle, surface or volume of plasma), exposure time, and
process gas (e.g., air, argon or helium) (18). For this study, we used a non-thermal
dielecteic barrier discharge (DBD) plasma source with a gas
consisting of 70% oxygen and 30% argon at atmospheric pressure.
Previously, we showed that a DBD plasma system can generate ROS and
induce apoptosis in human keratinocytes (19), and we used the same experimental
settings for this study.
Materials and methods
Reagents
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide (MTT), 2′,7′-dichlorodihydrofluorescein diacetate
(DCFH-DA), N-acetylcysteine (NAC), and anti-actin primary
antibody were purchased from Sigma-Aldrich (St. Louis, MO, USA).
ER-Tracker™ Blue-White DPX and Rhod-2 AM dyes were purchased from
Molecular Probes, Inc. (Eugene, OR, USA). Primary antibodies
against GRP78, phosphorylated eukaryotic initiation factor 2α
(p-eIF2α), phosphorylated PERK (p-PERK), and X-box binding protein
1 (XBP1) were purchased from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA), and anti-CHOP antibody was purchased from Cell
Signaling Technology, Inc. (Beverly, MA, USA). Primary antibody
against phosphorylated IRE1 (p-IRE1) was purchased from Thermo
Fisher Scientific, Inc. (Rockford, IL, USA).
Cell culture
The SNUC5 colon cancer cell line was obtained from
the Korean Cell Line Bank (Seoul, Republic of Korea). Cells were
cultured at 37°C in an incubator containing humidified air (95%)
and carbon dioxide (5%). The culture medium was RPMI-1640
(Invitrogen, Grand Island, NY, USA) containing 10% heat-inactivated
FBS (Sigma-Aldrich), streptomycin (100 µg/ml), and
penicillin (100 U/ml).
Plasma treatment
Non-thermal DBD was used as the plasma source, as
previously described (19). For DBD
plasma treatment, cells were trypsinized and counted to adjust the
density to 2×105 cells/ml, and 11 ml of cell suspension
was placed in a 60-mm cell culture dish. After exposure to DBD
plasma, cell suspensions were transferred to new cell culture
dishes or wells for subsequent experiments. Control samples were
subjected to all steps except plasma exposure.
Detection of intracellular ROS
Plasma-treated cells were seeded at a density of
1×105 cells/plate and incubated for 24 h at 37°C. Cells
were then harvested, washed, and re-suspended in PBS containing 25
µM DCFH-DA. After 15 min at 37°C, the cells were washed,
re-suspended in PBS, and analyzed by flow cytometry (Becton
Dickinson, Mountain View, CA, USA). For image analysis, cells were
loaded with DCFH-DA and incubated for 30 min at 37°C. The stained
cells were washed and mounted on microscope slides in mounting
medium (Dako, Carpinteria, CA, USA). Microscopic images were
obtained on a confocal laser-scanning microscope and analyzed using
LSM 5 PASCAl software (Carl Zeiss Jena GmbH, Jena, Germany).
Cell viability
Cell viability was assayed by MTT test. Cells were
suspended in 11 ml of media at a density of 2×105
cells/ml, and the suspensions were placed in 60-mm cell culture
dishes. With the lids removed from the dishes, the cells were
exposed to plasma for 2 min. Plasma-treated cells were transferred
to 24-well plates at a density of 1×105 cells/well.
Twenty-four hours later, 50 µl of MTT stock solution (2
mg/ml) were added to each well to yield a total reaction volume of
200 µl. After incubation for 4 h, the plate was centrifuged
at 800 × g for 5 min, and the supernatants were aspirated. Formazan
crystals in each well were dissolved in 150 µl of
dimethyl-sulfoxide (DMSO), and A540 was measured using a
scanning multi-well spectrophotometer (20).
Nuclear staining with Hoechst 33342
Cells were transferred into 24-well plates following
plasma exposure and incubated at 37°C for 24 h. The DNA-specific
fluorescent dye Hoechst 33342 was added to each well, and the cells
were incubated for 10 min at 37°C. The stained cells were
visualized under a fluorescence microscope equipped with a
CoolSNAP-Pro color digital camera. The degree of nuclear
condensation was evaluated by counting apoptotic cells in randomly
selected equally sized areas in each well.
ER staining
Plasma-exposed cells were seeded in chamber slides
(Nalge Nunc International Corp., Rochester, NY, USA) at a density
of 1×105 cells/ml and incubated for 24 h at 37°C.
ER-Tracker™ Blue-White DPX dye was added to the cells, and the
samples were incubated for an additional 30 min. The cells were
washed twice with PBS before the addition of mounting medium.
Microscopic images were collected using LSM 5 PASCAL software.
Western blotting
Harvested cells were lysed by incubation on ice for
30 min in 150 µl of lysis buffer (iNtRON Biotechnology,
Seoul, Republic of Korea). The resultant cell lysates were
centrifuged at 13,000 rpm for 5 min. Supernatants were collected,
and protein concentrations were determined. Aliquots were boiled
for 5 min and electrophoresed on 12% SDS-polyacrylamide gels.
Protein blots of the gels were transferred onto nitrocellulose
membranes. The membranes were incubated with the appropriate
primary antibodies (1:1,000) followed by horseradish
peroxidase-conjugated anti-IgG secondary antibodies (1:5,000)
(Pierce, Rockford, IL, USA). Protein bands were detected using an
enhanced chemiluminescence western blotting detection kit
(Amersham, Little Chalfont, UK).
Measurement of mitochondrial
Ca2+ levels
Mitochondrial Ca2+ levels were measured
using Rhod-2 AM (21).
Plasma-treated cells were seeded at a density of 1×105
cells/plate and incubated for 24 h at 37°C. Cells were harvested,
washed, and re-suspended in PBS containing Rhod-2 AM. After 15 min
at 37°C, the cells were washed, re-suspended in PBS, and analyzed
by flow cytometry. For image analysis, cells were loaded with
Rhod-2 AM and incubated for 30 min at 37°C. The stained cells were
washed and mounted on microscope slides in mounting medium.
Microscopic images were obtained under a confocal laser-scanning
microscope and analyzed using lSM 5 PASCAl software.
Statistical analysis
All measurements were made in triplicates, and all
values are expressed as means ± standard error of the mean (SEM).
The results were subjected to analysis of variance (ANOVA) using
the Tukey's test to analyze differences. P<0.05 was considered
statistically significant.
Results
NTGP induces ROS level in SNUC5
cells
Plasma generates ROS and induces oxidative stress in
cells (9). Therefore, we assessed
intracellular ROS generation in plasma-treated cells using DCFH-DA,
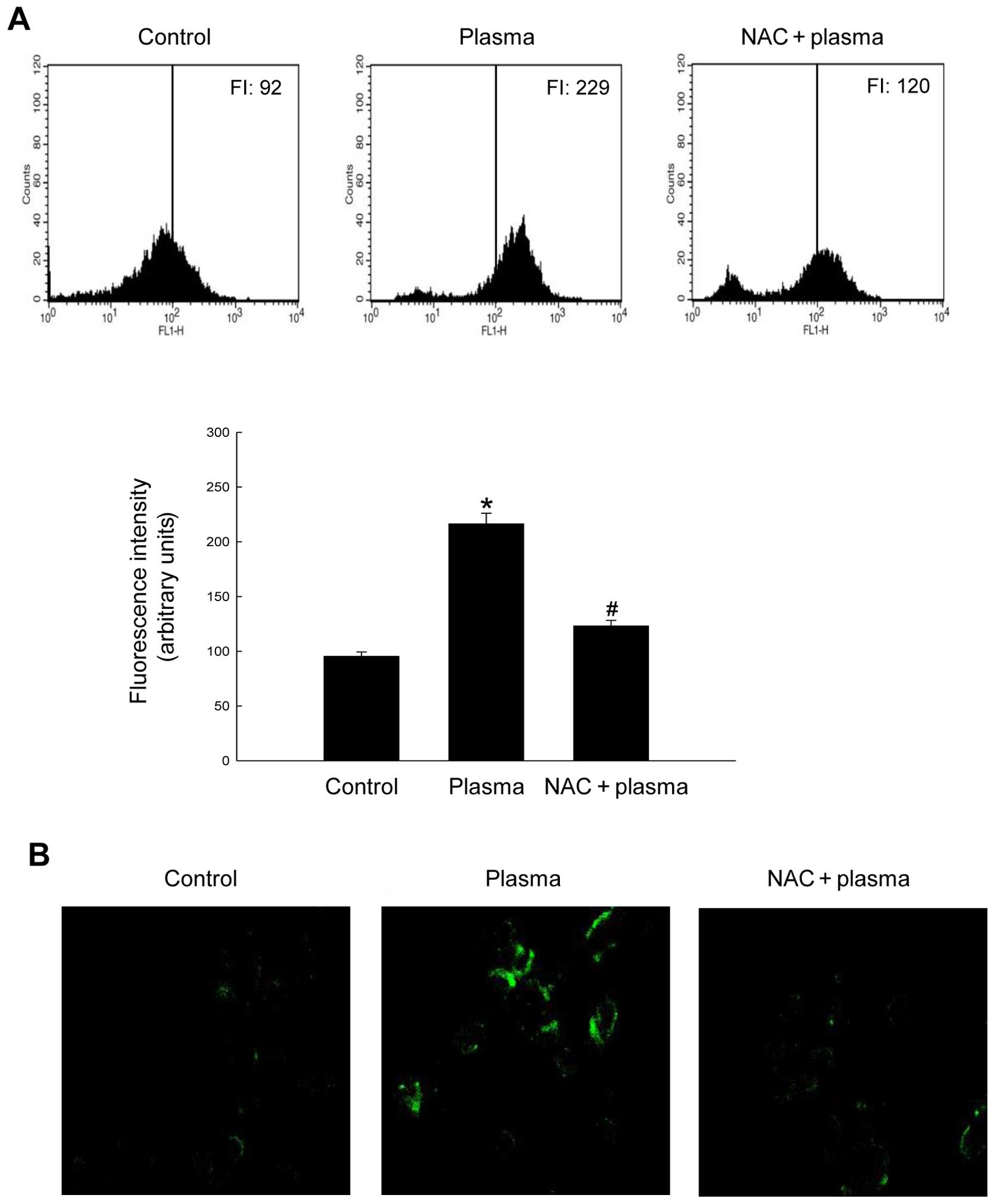
a ROS-sensitive fluorogenic dye. Flow cytometry revealed that 2 min
of plasma exposure increased DCFH-DA fluorescence to 229 (FI: 229)
vs. 92 in control cells not exposed to plasma (Fig. 1A). Samples pre-treated with NAC, a
well-known free radical scavenger, yielded an FI value of 120. The
flow cytometry results were consistent with those obtained by
confocal microscopy analysis of DCFH-DA-stained cells (Fig. 1B). These results indicate that NTGP
generates ROS in SNUC5 human colon carcinoma cells.
NTGP induces apoptosis in SNUC5
cells
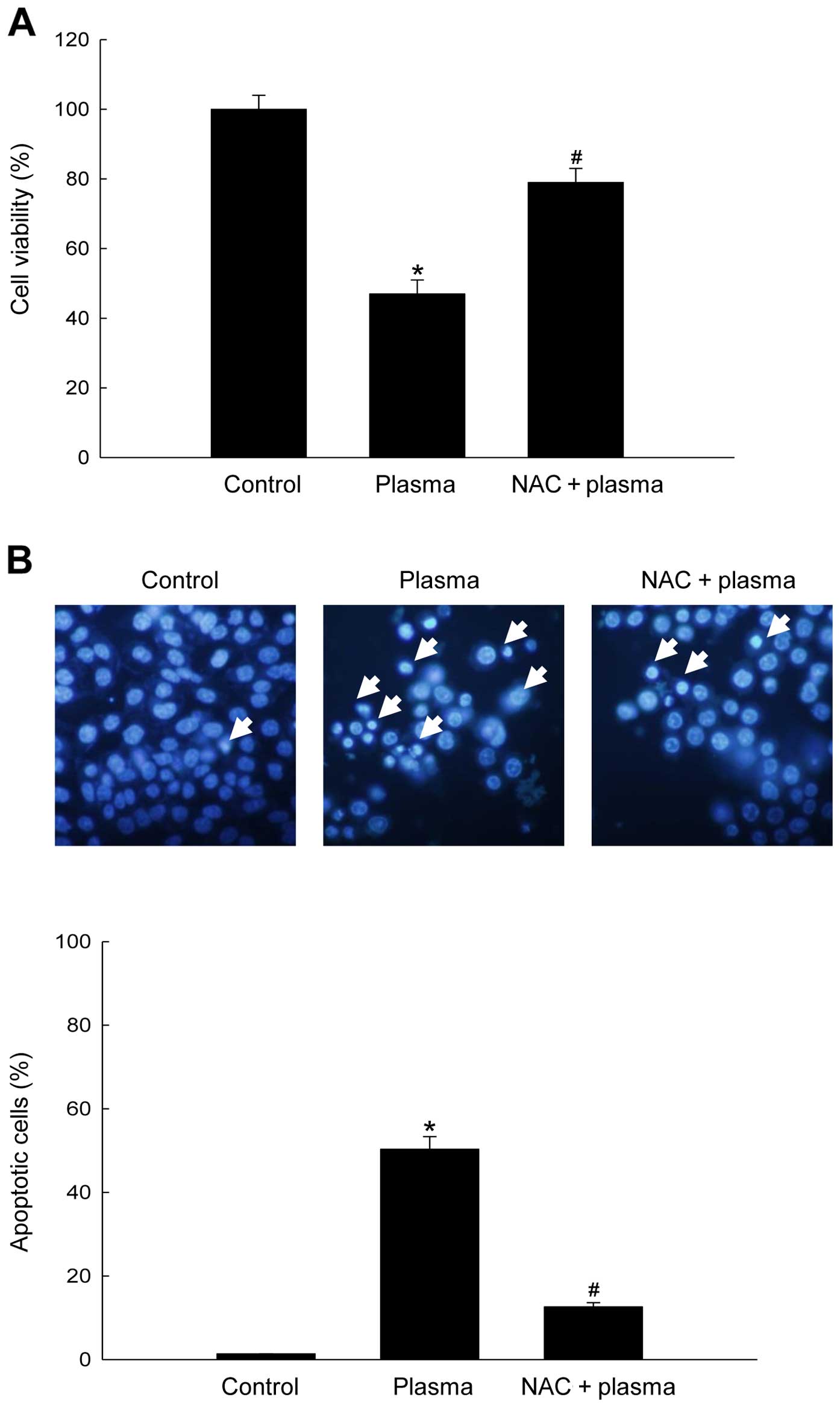
MTT assay results revealed that plasma treatment
markedly decreased cell viability (55% relative to untreated
controls). This reduction in viability was rescued by treatment of
NAC prior to plasma exposure (Fig.
2A). Next, we investigated whether cells would undergo
apoptosis following plasma exposure using the nuclear staining dye
Hoechst 33342. As shown in Fig. 2B,
>50% of plasma-exposed cells underwent apoptosis, as indicated
by the formation of apoptotic bodies. Again, NAC pre-treatment
diminished the formation of apoptotic bodies. These results suggest
that NTGP induces apoptosis via ROS generation in SNUC5 cells.
Plasma-induced apoptosis in SNUC5 cells
is mediated by ER stress
Overwhelming ROS levels cause ER stress (16). Therefore, we next investigated
whether plasma exposure can induce ER stress. To this end,
plasma-exposed cells were stained with ER-Tracker™ Blue-White DPX
dye, and were observed by confocal microscopy. Under these
conditions, plasma-exposed cells stained bright blue, indicative of
ER stress (Fig. 3A); as with the
loss of viability and apoptosis described above, pre-treatment with
NAC prior to plasma exposure decreased the brightness of staining,
demonstrating that ER stress in plasma-treated cells arises due to
high levels of ROS.
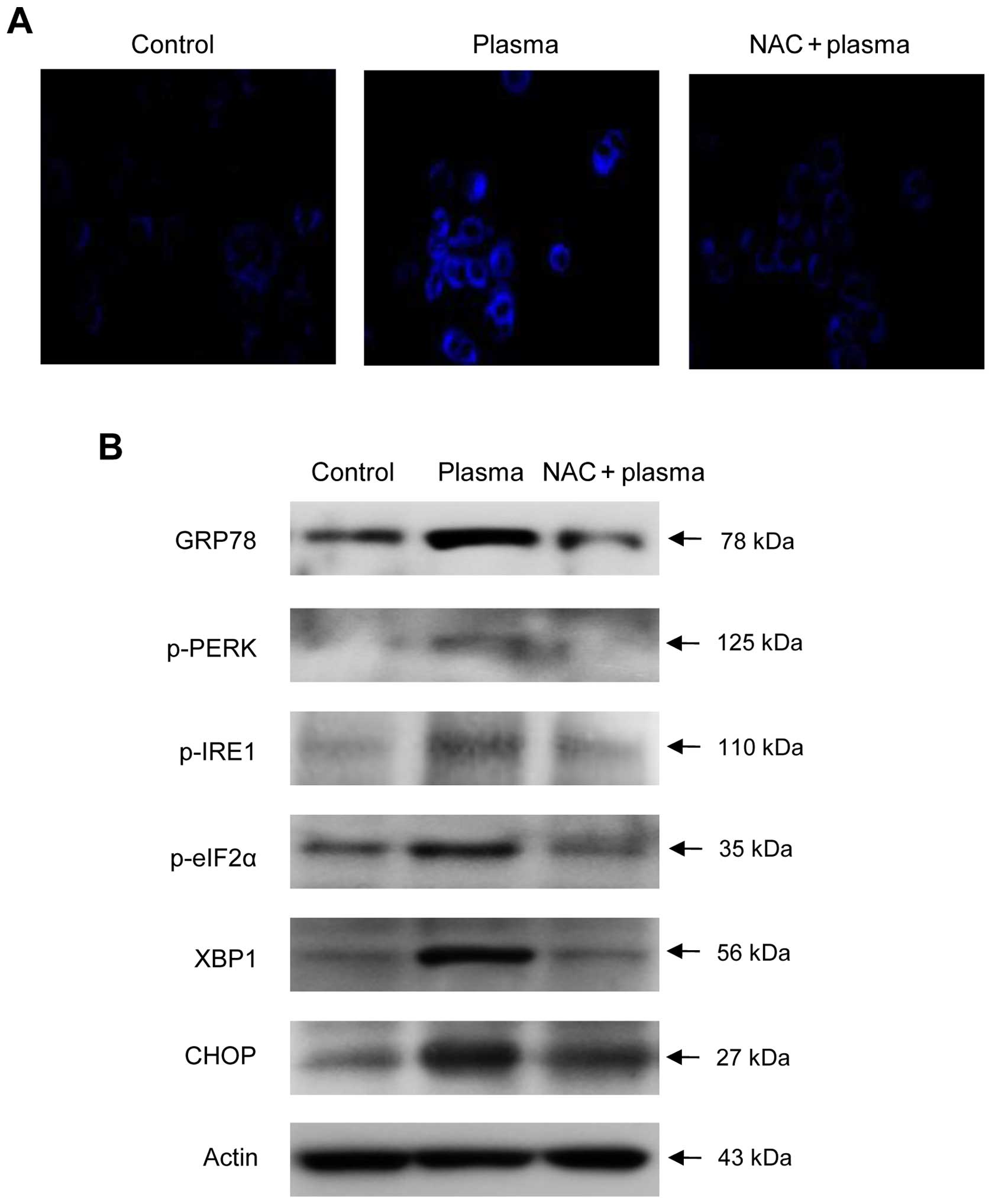 | Figure 3NTGP induces ER stress. (A) Cells
were exposed to plasma and incubated for 24 h at 37°C, and then
ER-Tracker™ Blue-White DPX fluorescence staining dye was added and
ER stress was analyzed by confocal microscopy. (B) SNUC5 cells were
exposed to plasma and incubated for 24 h at 37°C. Cells were
harvested, and the proteins were extracted. Expression levels of
GRP78, p-PERK, p-IRE1, p-eIF2α, XBP1, and CHOP proteins were
evaluated using western blotting. Actin, a housekeeping protein,
was used to confirm equal protein loading. NTGP, non-thermal gas
plasma; ER, endoplasmic reticulum; GRP78, glucose-related protein
78; p-PERK, phosphorylated protein kinase R (PKR)-like ER kinase;
p-IRE1, phosphorylated inositol-requiring enzyme 1; p-eIF2α,
phosphorylated eukaryotic initiation factor 2α; XBP1, X-box binding
protein 1; CHOP, CCAAT/enhancer-binding protein homologous
protein. |
To confirm the induction of ER stress upon plasma
exposure, we analyzed ER stress-related proteins. The UPR is a
suite of signaling pathways that cells activate to restore cellular
homeostasis once ER stress has occurred. Cells that are unable to
achieve homeostasis undergo apoptosis (22). The ER-resident transmembrane
receptors PERK (pancreatic eIF2-α kinase or protein kinase R
(PKR)-like ER kinase), inositol-requiring enzyme 1 (IRE1), and ATF6
are maintained in their inactive forms through association with
GRP78, which dissociates upon ER stress. Upon GRP78 dissociation,
the UPR receptors are activated by phosphorylation (15). Western blotting revealed that the
expression of GRP78, which is itself a transcriptional target of
the UPR, increased dramatically following plasma treatment
(Fig. 3B). Furthermore, levels of
p-IRE1 and p-PERK were significantly elevated following plasma
exposure, suggesting that plasma exposure induces the UPR in SNUC5
cells. In accordance with the data presented above, NAC
pre-treatment attenuated upregulation of GRP78 and phosphorylation
of both IRE1 and PERK. Activated PERK phosphorylates eIF2α, leading
to inhibition of general protein translation, a hallmark of the UPR
(23). Plasma exposure increased
the level of p-eIF2α, confirming the activation of the UPR; as with
the other indicators of UPR activity, the p-eIF2α level decreased
in NAC-pre-treated cells (Fig.
3B).
Next, we investigated whether the UPR restores ER
function or initiates apoptosis in SNUC5 cells. After
non-conventional splicing of the XBP1 mRNA by IRE1, XBP1 protein
translocates to the nucleus, where it induces transcription of
genes involved in protein degradation and inhibition of the
PERK-mediated translational block (24). At this stage, if the UPR has
restored homeostasis, the cell will survive; otherwise, it will be
driven to apoptosis. Expression of CCAAT/enhancer-binding protein
homologous protein (CHOP), also known as growth arrest and DNA
damage-inducible gene 153 (GADD153), is activated by the UPR; CHOP
downregulates the anti-apoptotic mitochondrial protein Bcl-2
(25), fostering a pro-apoptotic
environment and stimulating the mitochondria to release cytochrome
c and activate caspase-3. Our data revealed that plasma
exposure increased the expression of both XBP1 and CHOP, indicating
that ER stress exerted by NTGP causes apoptosis in SNUC5 cells. All
of the aforementioned effects were suppressed by the pre-treatment
with NAC prior to plasma exposure.
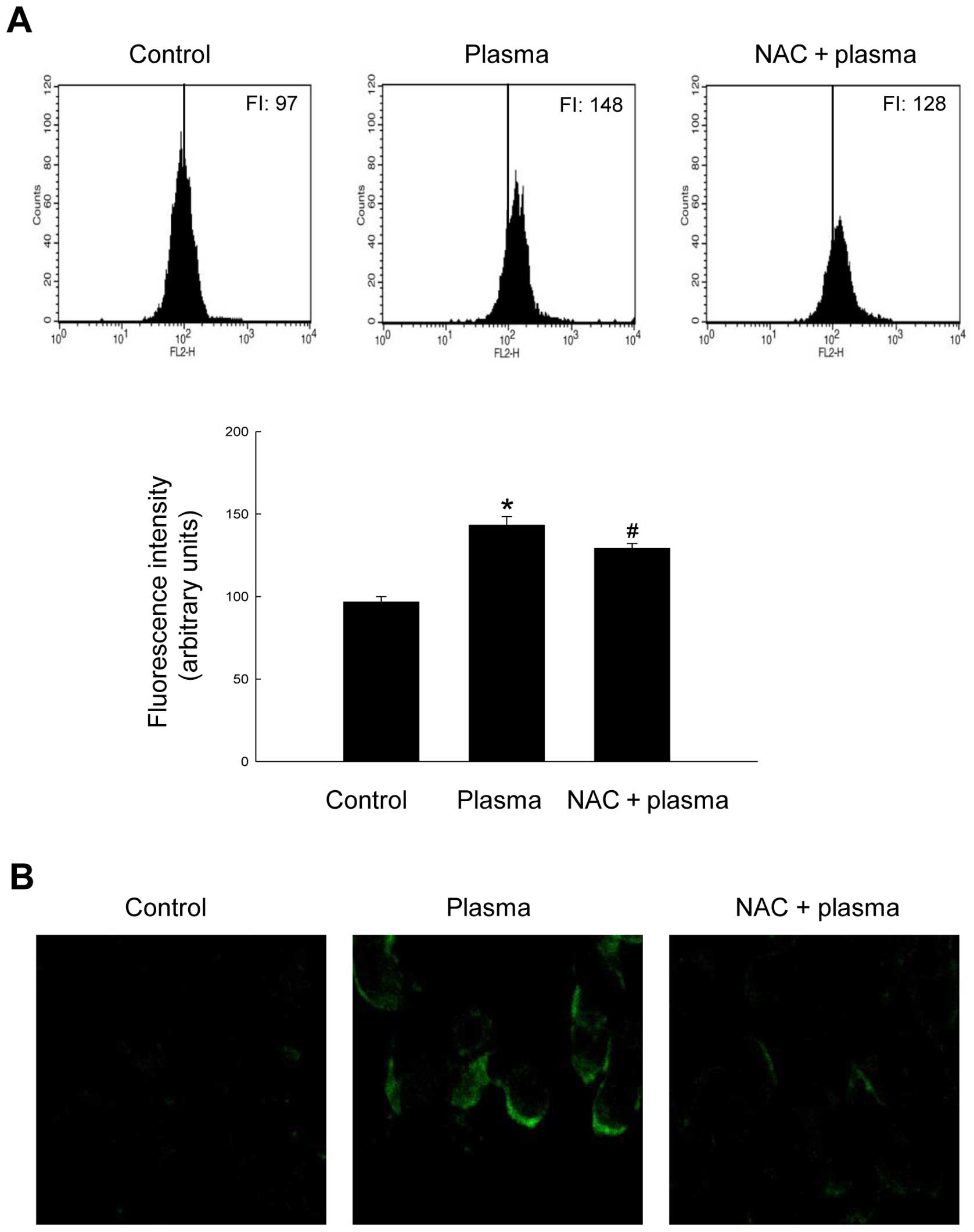
Mitochondrial Ca2+ overloading
is involved in plasma-mediated ER stress
ER stress is characterized by various molecular
markers; for example, depletion of Ca2+ from the ER and
accumulation of Ca2+ in mitochondria are hallmarks of
severe ER stress (26). Therefore,
we detected mitochondrial Ca2+ overload in
plasma-exposed cells using Rhod-2 AM fluorescent dye. Flow
cytometry revealed a significant increase in the mitochondrial
Ca2+ level in plasma-treated cells (FI: 148) relative to
that in controls (FI: 97) (Fig.
4A), which was attenuated (FI: 128) by pre-treatment with NAC
before plasma exposure. We also monitored mitochondrial
Ca2+ overload by confocal microscopy in cells stained
with Rhod-2 AM. As expected, the confocal microscopy results
(Fig. 4B) were consistent with the
flow cytometry data. These findings suggest that plasma treatment
triggers ER stress and mitochondrial Ca2+ accumulation,
which in turn promotes apoptosis, in SNUC5 colon carcinoma cells.
Because NAC pre-treatment could ameliorate these effects, it is
likely that the ER stress is induced by excess ROS.
Discussion
As a widespread type of cancer, colorectal cancer
represents a serious health threat throughout the world.
Chemoresistance and side-effects of prolonged chemical treatments
are substantial challenges for the treatment of colorectal cancer
(5). Therefore, novel non-chemical
strategies for treating colorectal cancer are urgently needed. NTGP
has been characterized in various clinical applications as a
promising tool for wound healing (27), plasma sterilization (28), blood coagulation (29), cell detachment (30), induction of apoptosis (31), and cancer therapy (32). Therefore, we investigated whether
NTGP induces apoptosis in SNUC5 human colon carcinoma cells by
generating ER stress.
For this study, we exposed cells to NTGP for 2 min
using a non-thermal DBD plasma source. First, we investigated
whether NTGP induces ROS. To this end, we stained plasma- exposed
cells with DCFH-DA and examined them by flow cytometry and confocal
microscopy (Fig. 1A and B). Data
obtained by both methods indicated that plasma treatment
significantly increased intracellular ROS generation. Furthermore,
NTGP decreased cell viability compared to the controls (Fig. 2A) by inducing apoptosis (Fig. 2B). Pre-treatment with NAC attenuated
these and all other consequences of plasma treatment, strongly
indicating that cell death and other effects of plasma exposure are
mediated by ROS.
Overwhelming ROS levels can trigger ER stress.
Staining of cells with ER-Tracker™ Blue-White DPX dye confirmed
that plasma treatment induced ER stress (Fig. 3A). A mean of combating the
detrimental effects of ER-stressed cells is the UPR, whose basic
purpose is to halt protein synthesis and accumulation of misfolded
proteins until proper ER function can be restored. If the UPR
cannot restore cellular homeostasis, it triggers apoptosis
(16). Under normal conditions,
GRP78 is bound to the luminal domains of PERK and IRE1, maintaining
them in their inactive states. Upon ER stress, GRP78 dissociates
from the receptors, allowing them to be phosphorylated (33). Once activated, the UPR halts protein
synthesis and upregulates production of ER-resident chaperones that
promote protein folding and help the cell recover from stress.
Plasma treatment increased the levels of GRP78, p-PERK, and p-IRE1,
and also activated PERK-mediated phosphorylation of p-eIF2α, which
imposes a translational block on ER protein synthesis (33) (Fig.
3B). These results confirmed that plasma exposure activates key
proteins involved in the UPR.
Persistent ER stress inhibits PERK and halts the
UPR, ultimately resulting in apoptosis (34). Cleaved ATF6 translocates to the
nucleus and initiates transcription of XBP1 and CHOP. XBP1 mRNA is
unconventionally spliced by p-IRE1, enabling translation of XBP1
protein (34), which translocates
to the nucleus and induces the expression of P58IPK, which inhibits
p-PERK and initiates protein degradation (23). Plasma exposure increased the levels
of both XBP1 and CHOP (Fig. 3B).
CHOP downregulates the anti-apoptotic mitochondrial protein Bcl-2,
increasing mitochondrial membrane permeability and releasing
cytochrome c into the cytoplasm to trigger apoptosis
(35). Ca2+ accumulation
in the mitochondria is a marker of the early and late stages of
apoptosis (26). Because Bcl-2
expression is blocked under severe ER stress by activation of CHOP,
Ca2+ can leak from the ER into the cytoplasm and
mitochondria. Thus, mitochondrial Ca2+ overload is a
hallmark of intensive ER stress (26). Consistent with this, plasma exposure
strongly increased the mitochondrial Ca2+ level in SNUC5
cells (Fig. 4A and B).
These data confirm our hypothesis that NTGP induces
apoptosis in SNUC5 human colon carcinoma cells via induction of ER
stress. The attenuation of the UPR and related phenomena by NAC
pre-treatment demonstrates that the effects of plasma exposure are
mediated by generation of ROS.
Acknowledgments
This study was supported by the R&D Program of
Plasma Advanced Technology for Agriculture and Food (Plasma
Farming) through the National Fusion Research Institute (NFRI) of
Korea funded by the Government.
References
|
1
|
Pietrzyk L, Torres A, Maciejewski R and
Torres K: Obesity and obese-related chronic low-grade inflammation
in promotion of colorectal cancer development. Asian Pac J Cancer
Prev. 16:4161–4168. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Haggar FA and Boushey RP: Colorectal
cancer epidemiology: Incidence, mortality, survival, and risk
factors. Clin Colon Rectal Surg. 22:191–197. 2009. View Article : Google Scholar :
|
|
3
|
Birmingham JM, Busik JV, Hansen-Smith FM
and Fenton JI: Novel mechanism for obesity-induced colon cancer
progression. Carcinogenesis. 30:690–697. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Morrison DS, Parr CL, Lam TH, Ueshima H,
Kim HC, Jee SH, Murakami Y, Giles G, Fang X, Barzi F, et al:
Behavioural and metabolic risk factors for mortality from colon and
rectum cancer: Analysis of data from the Asia-Pacific Cohort
Studies Collaboration. Asian Pac J Cancer Prev. 14:1083–1087. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dy GK, Hobday TJ, Nelson G, Windschitl HE,
O'Connell MJ, Alberts SR, Goldberg RM, Nikcevich DA and Sargent DJ:
Long-term survivors of metastatic colorectal cancer treated with
systemic chemotherapy alone: A north central cancer treatment group
review of 3811 patients, n0144. Clin Colorectal Cancer. 8:88–93.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bussiahn R, Brandenburg R, Gerling T,
Kindel E, Lange H, Lembke N, Weltmann KD, von Woedtke Th and Kocher
T: The hairline plasma: An intermittent negative dc-corona
discharge at atmospheric pressure for plasma medical applications.
Appl Phys Lett. 96:1437012010. View Article : Google Scholar
|
|
7
|
Gadri RB, Roth JR, Montie TC,
Kelly-Wintenberg K, Tsai PPY, Helfritch DJ, Feldman P, Sherman DM
and Karakaya F: Sterilization and plasma processing of room
temperature surfaces with a one atmosphere uniform glow discharge
plasma (OAUGDP). Surf Coat Tech. 131:528–541. 2000. View Article : Google Scholar
|
|
8
|
Kim CH, Kwon S, Bahn JH, Lee K, Jun SI,
Rack PD and Baek SJ: Effects of atmospheric nonthermal plasma on
invasion of colorectal cancer cells. Appl Phys Lett. 96:2437012010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sensenig R, Kalghatgi S, Cerchar E,
Fridman G, Shereshevsky A, Torabi B, Arjunan KP, Podolsky E,
Fridman A, Friedman G, et al: Non-thermal plasma induces apoptosis
in melanoma cells via production of intracellular reactive oxygen
species. Ann Biomed Eng. 39:674–687. 2011. View Article : Google Scholar
|
|
10
|
Cheng X, Sherman J, Murphy W, Ratovitski
E, Canady J and Keidar M: The effect of tuning cold plasma
composition on glioblastoma cell viability. PLoS One. 9:e986522014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Seo K, Ki SH and Shin SM: Methylglyoxal
induces mitochondrial dysfunction and cell death in liver. Toxicol
Res. 30:193–198. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Park J, Bae EK, Lee C, Choi JH, Jung WJ,
Ahn KS and Yoon SS: Establishment and characterization of
bortezomib-resistant U266 cell line: Constitutive activation of
NF-κB-mediated cell signals and/or alterations of
ubiquitylation-related genes reduce bortezomib-induced apoptosis.
BMB Rep. 47:274–279. 2014. View Article : Google Scholar :
|
|
13
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gaut JR and Hendershot LM: The
modification and assembly of proteins in the endoplasmic reticulum.
Curr Opin Cell Biol. 5:589–595. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Faitova J, Krekac D, Hrstka R and Vojtesek
B: Endoplasmic reticulum stress and apoptosis. Cell Mol Biol Lett.
11:488–505. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schröder M and Kaufman RJ: ER stress and
the unfolded protein response. Mutat Res. 569:29–63. 2005.
View Article : Google Scholar
|
|
17
|
Breckenridge DG, Germain M, Mathai JP,
Nguyen M and Shore GC: Regulation of apoptosis by endoplasmic
reticulum pathways. Oncogene. 22:8608–8618. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Haertel B, Straßenburg S, Oehmigen K,
Wende K, von Woedtke T and Lindequist U: Differential influence of
components resulting from atmospheric-pressure plasma on integrin
expression of human HaCaT keratinocytes. BioMed Res Int.
2013:7614512013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim KC, Piao MJ, Madduma Hewage SR, Han X,
Kang KA, Jo JO, Mok YS, Shin JH, Park Y, Yoo SJ, et al: Non-thermal
dielectric-barrier discharge plasma damages human keratinocytes by
inducing oxidative stress. Int J Mol Med. 37:29–38. 2016.
|
|
20
|
Carmichael J, DeGraff WG, Gazdar AF, Minna
JD and Mitchell JB: Evaluation of a tetrazolium-based semiautomated
colorimetric assay: Assessment of chemosensitivity testing. Cancer
Res. 47:936–942. 1987.PubMed/NCBI
|
|
21
|
Fonteriz RI, de la Fuente S, Moreno A,
Lobatón CD, Montero M and Alvarez J: Monitoring mitochondrial
[Ca(2+)] dynamics with rhod-2, ratiometric pericam and aequorin.
Cell Calcium. 48:61–69. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Szegezdi E, Logue SE, Gorman AM and Samali
A: Mediators of endoplasmic reticulum stress-induced apoptosis.
EMBO Rep. 7:880–885. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yan W, Frank CL, Korth MJ, Sopher BL,
Novoa I, Ron D and Katze MG: Control of PERK eIF2α kinase activity
by the endoplasmic reticulum stress-induced molecular chaperone
P58IPK. Proc Natl Acad Sci USA. 99:15920–15925. 2002. View Article : Google Scholar
|
|
24
|
Lee AH, Iwakoshi NN and Glimcher LH: XBP-1
regulates a subset of endoplasmic reticulum resident chaperone
genes in the unfolded protein response. Mol Cell Biol.
23:7448–7459. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yamaguchi H and Wang HG: CHOP is involved
in endoplasmic reticulum stress-induced apoptosis by enhancing DR5
expression in human carcinoma cells. J Biol Chem. 279:45495–45502.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pinton P, Giorgi C, Siviero R, Zecchini E
and Rizzuto R: Calcium and apoptosis: ER-mitochondria
Ca2+ transfer in the control of apoptosis. Oncogene.
27:6407–6418. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Haertel B, von Woedtke T, Weltmann KD and
Lindequist U: Non-thermal atmospheric-pressure plasma possible
application in wound healing. Biomol Ther (Seoul). 22:477–490.
2014. View Article : Google Scholar
|
|
28
|
Moman RM and Najmaldeen H: The
bactericidal efficacy of cold atmospheric plasma technology on some
bacterial strains. Egypt Acad J Biolog Sci. 2:43–47. 2010.
|
|
29
|
Kuo SP, Tarasenko O, Chang J, Popovic S,
Chen CY, Fan HW, Scott A, Lahiani M, Alusta P, Drake JD, et al:
Contribution of a portable air plasma torch to rapid blood
coagulation as a method of preventing bleeding. New J Phys.
11:1150162009. View Article : Google Scholar
|
|
30
|
Hoentsch M, von Woedtke T, Weltmann KD and
Nebe JB: Time-dependent effects of low-temperature
atmospheric-pressure argon plasma on epithelial cell attachment,
viability and tight junction formation in vitro. J Phys D: Appl
Phys. 45:0252062012. View Article : Google Scholar
|
|
31
|
Tuhvatulin AI, Sysolyatina EV, Scheblyakov
DV, Logunov DY, Vasiliev MM, Yurova MA, Danilova MA, Petrov OF,
Naroditsky BS, Morfill GE, et al: Non-thermal plasma causes
p53-dependent apoptosis in human colon carcinoma cells. Acta
Naturae. 4:82–87. 2012.PubMed/NCBI
|
|
32
|
Partecke LI, Evert K, Haugk J, Doering F,
Normann L, Diedrich S, Weiss FU, Evert M, Huebner NO, Guenther C,
et al: Tissue tolerable plasma (TTP) induces apoptosis in
pancreatic cancer cells in vitro and in vivo. BMC Cancer.
12:4732012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee AS: Glucose-regulated proteins in
cancer: Molecular mechanisms and therapeutic potential. Nat Rev
Cancer. 14:263–276. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
van der Kallen CJ, van Greevenbroek MM,
Stehouwer CD and Schalkwijk CG: Endoplasmic reticulum
stress-induced apoptosis in the development of diabetes: Is there a
role for adipose tissue and liver? Apoptosis. 14:1424–1434. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tabas I and Ron D: Integrating the
mechanisms of apoptosis induced by endoplasmic reticulum stress.
Nat Cell Biol. 13:184–190. 2011. View Article : Google Scholar : PubMed/NCBI
|