Introduction
Rhabdomyosarcoma (RMS) is the most common malignant
soft tissue sarcoma, and is histologically classified into two
major subtypes; embryonal rhabdomyosarcoma (ERMS) and alveolar
rhabdomyosarcoma (ARMS). Although ERMS prognosis has improved via
multidisciplinary treatment (1),
prognosis for ARMS is still poor. In particular, the subgroup of
ARMS which expresses the PAX3-FOXO1 chimera gene derived
from the t(2;13) translocation, especially shows invasive and
metastatic behavior (2,3) resulting in extremely poor prognosis
(4). Therefore, it is necessary to
elucidate the pathogenesis of metastasis in ARMS to develop novel
drugs and treatment methods.
PAX3 is one of the transcriptional factors that play
a significant role in the appropriate development during the
prenatal period. PAX3 regulates the expression of MET during limb
muscle development, and is required for the proper migration of
myogenic precursor to the limbs (5).
Hepatocyte growth factor (HGF) and its receptor
(MET) stimulate the proliferation or migration of various cancer
cell lines (6). Additionally, MET
promotes motility of premyogenic cells (7), and is involved in tumorigenic
processes including proliferation, metastasis and invasion in
malignant tumors such as hepatocellular carcinoma, colon cancer,
lung cancer, bone sarcoma, or ovarian cancer (8,9). In
ARMS, PAX3-FOXO1 as well as PAX3 upregulates MET (10). In mice with RMS, in which HGF is
excessively expressed and there is a deficiency of Ink4a/Arf, RMS
tumors develop at high rates (11)
and the invasiveness of RMS cell lines is enhanced upon forced
expression of MET (12). We have
already confirmed that downregulation of PAX3-FOXO1 caused
reduction in levels of MET, resulting in reduced cell motility upon
HGF stimulation (13). Therefore,
we speculate that HGF and MET are deeply involved in tumorigenesis
and invasiveness of ARMS. Previous studies have reported the
inhibition of metastasis in RMS cell lines by small interfering RNA
(siRNA), drugs or by the constitutive activation of MET (14–16).
However, the changes in the downstream processes of RMS metastasis
are unknown.
In this study, we compared the responses of ERMS and
ARMS cell lines to HGF stimulation and investigated the signal
transduction pathways downstream of MET. Our study suggests that
ERK2 signaling, downstream of MET is a therapeutic target to
suppress the invasive and metastatic properties of ARMS.
Materials and methods
Cell culture and reagents
Human ERMS cell lines RD, CTR subclone 11 (CT11),
and KP-RMS-KH (KH) (17), and human
ARMS cell lines SJ-Rh30 (Rh30), SCMC-RM2 (RM2) (18), and SJ-Rh28 (Rh28) were used. HepG2
cells were used as the MET expressing control cell line. Cells were
maintained in RPMI-1640 containing 10% fetal calf serum (FCS) at
37°C in a 5% CO2 incubator. Recombinant human HGF was
purchased from PeproTech Inc (Rocky Hill, NJ, USA).
Quantification real-time PCR
Total RNA was extracted from the cell lines using
QIAamp RNeasy Protect Mini kit (Qiagen, Valencia, CA, USA).
PAX3-FOXO1 mRNA and MET mRNA were amplified by the polymerase chain
reaction (Takara Bio Inc., Siga, Japan) from the cDNA template
which was prepared by using Superscript™ First-Strand Synthesis
System for RT-PCR (Invitrogen, Carlsbad, CA, USA), and real-time
RT-PCR was carried out in the 7300 Real-time PCR System (Applied
Biosystems, Foster City, CA, USA) sing SYBR-Green 1 (Takara Bio
Inc.). In order to quantify the target mRNAs level,
glucose-6-phosphate dehydrogenase (GAPDH) gene was used as an
internal control. The cycle threshold values (Cq) for target mRNAs
and GAPDH for each sample were calculated. A normalized target
value was then derived by subtracting the amount of target mRNAs by
that of GAPDH (∆Cq) and calculated by the ∆∆Cq method (19).
siRNA
Silencer select siRNA for ERK1 (#4390624 S11137;
Ambion, Austin, TX, USA), ERK2 (#4390624 S11141; Ambion) or
negative control #1 (#4390843; Ambion) was transfected in Rh30
using Lipofectamine RNAiMAX (Invitrogen) according to the
manufacturer's recommendation. The concentration of siRNA was 0.5
nM, and the final concentration of lipofectamine RNAiMAX was
0.1%.
Western blot analysis
Cells were seeded on 6-well plates at the
concentration of 2×105 cells/well in 2 ml growth medium
(RPMI-1640 with 10% FCS). After overnight incubation, the medium
was replaced with serum-free medium for starvation. Incubations
were carried out for 30 min in the presence or absence of various
inhibitors, followed by HGF addition and culture for 24 h. Whole
cell lysates were separated by SDS-polyacrylamide gel
electrophoresis. Analysis was performed using various antibodies as
previously described (13).
Cell growth analysis
Cells were seeded on 6-well plates at the
concentration of 2×105 cells/35 mm-well in 2 ml of
growth medium (RPMI-1640 with 10% FCS). After culturing overnight,
the medium was replaced with fresh RPMI and serum-starved (0 h).
Then, the cells were cultured without serum, in the presence of 10%
FCS or HGF. The cells were harvested by trypsin processing after
24, 72 and 120 h and counted by Coulter counter.
Cell cycle analysis
Cells were harvested after being cultured for 24 h
in the presence of nocodasol in the culture bottle. Then, these
were seeded on a 100-mm dish and cultured overnight. The medium was
changed to the serum free medium, and the cells were cultured for
another 24 h without serum, in the presence of 10% FCS or HGF 75
ng/ml. These were harvested after trypsinization and washed once
with PBS. After releasing the cells in the propidium iodide
solution (50 mg/ml) to which RNase had been added and incubation
for 30 min, analysis was made by flow cytometry.
Wound-healing assay
Cells were seeded on 12-well plates and cultured for
24 h. These cell layers were then scratched with a pipette tip and
washed with RPMI (0 h). After incubating in serum-free medium, or
with rapamycin (100 ng/ml), PD98059 (20 µM), and U0126 (20 µM) for
30 min, HGF (75 ng/ml) was added and the cells were then cultured
for 24 h. Images of each well were captured at 0 and 24 h, and the
distance between edges of the wound region was measured.
Migration assay
An insert (pore size 8 micrometer; Nunc A/S,
Roskilde, Denmark) was placed on the wells of 12-well plates, and
cells were seeded in the upper chamber of the insert
(3×104 cells/insert). Various inhibitors were added to
the upper chamber of the insert and treatments were carried out for
30 min. HGF (75 ng/ml) was then added to the lower chamber of the
insert, and cells were cultured for 24 h. The cells that migrated
were fixed and stained with Diff-Quik (Sysmex, Hyogo, Japan) and
counted under the microscope.
Results
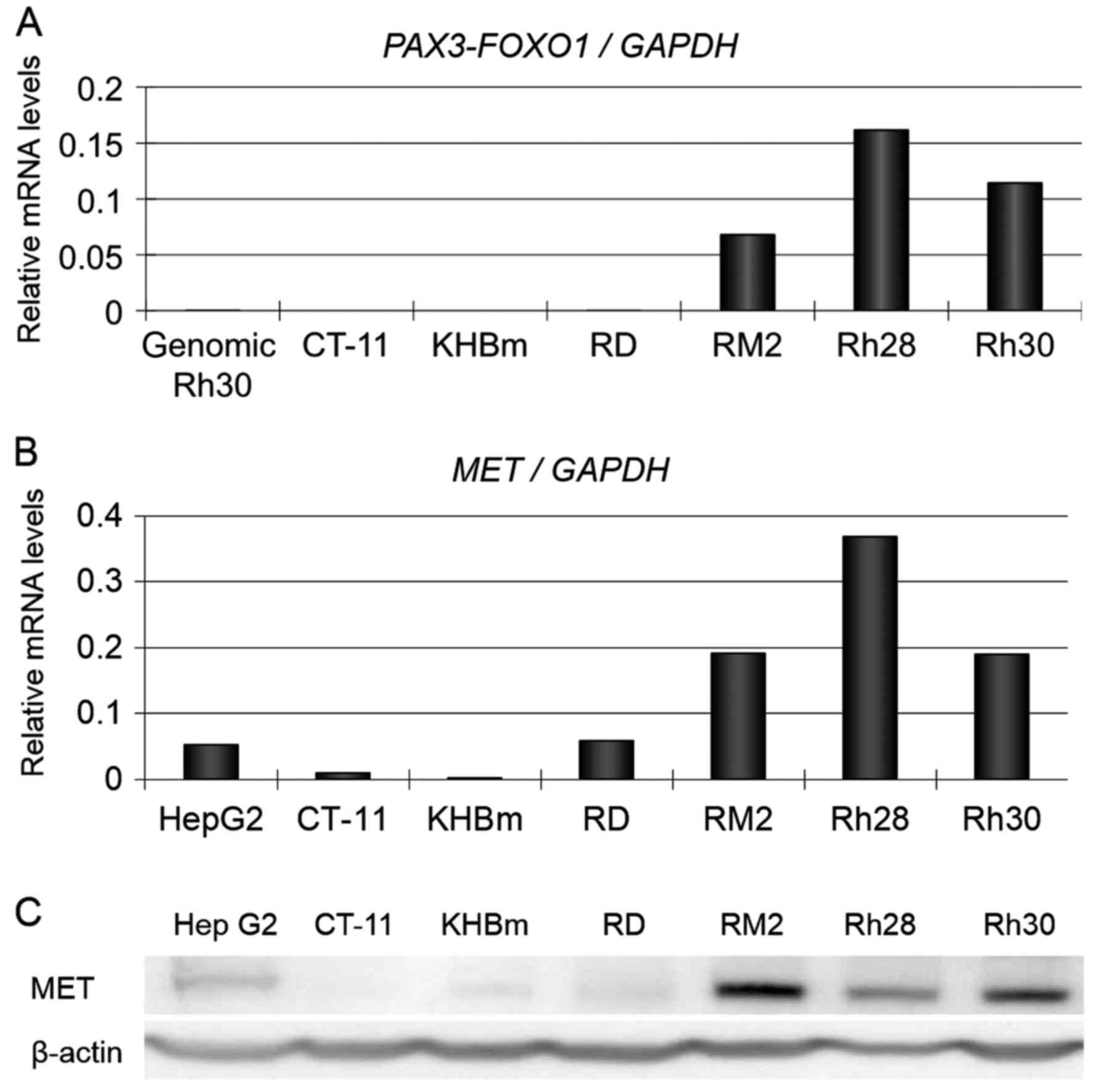
MET is overexpressed in parallel with
PAX3-FOXO1 expression in ARMS cells
PAX3-FOXO1 and MET mRNA levels were
examined in both ERMS and ARMS cells. PAX3-FOXO1 expression
was detected only in the ARMS cell lines (Fig. 1A) whereas MET mRNA levels in
the ARMS cell lines (Rh28, Rh30, and RM2) were apparently higher
than those in ERMS cells (RD, CT-11, and KH-BM) (Fig. 1B). By western blotting, we confirmed
that MET expression in the three ARMS cell lines was higher than in
the three ERMS cell lines that did not express PAX3-FOXO1
fusion gene (Fig. 1C).
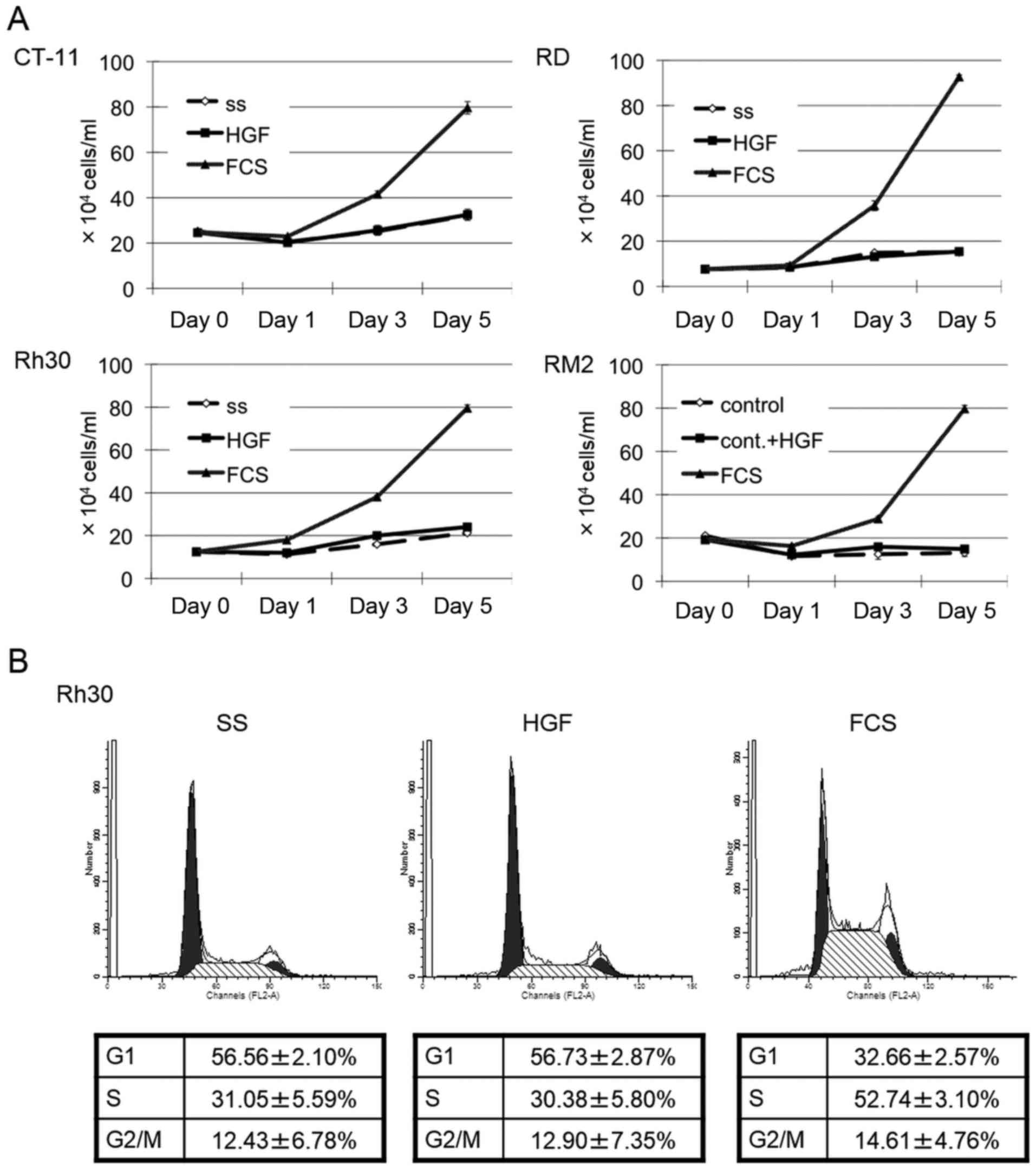
HGF stimulation does not influence
proliferation of alveolar type Rh30 cells
We investigated the effect of HGF stimulation on
ERMS and ARMS cell proliferation under serum starved conditions.
Upon stimulation with HGF, there was no significant growth in the
ERMS cells, RD and CT-11, or in the ARMS cells, Rh30 and RM2,
whereas all four cell lines proliferated in the presence of 10% FCS
(Fig. 2A). We then examined the
influence of HGF on cell cycle progression in ARMS Rh30 cells. When
ARMS Rh30 cells were stimulated with HGF, no significant changes in
cell cycle phase distribution were observed, whereas addition of
10% FCS apparently stimulated a decrease of cells in the G1 phase
and increased the cells in the S phase indicating G1-S cell cycle
progression (Fig. 2B).
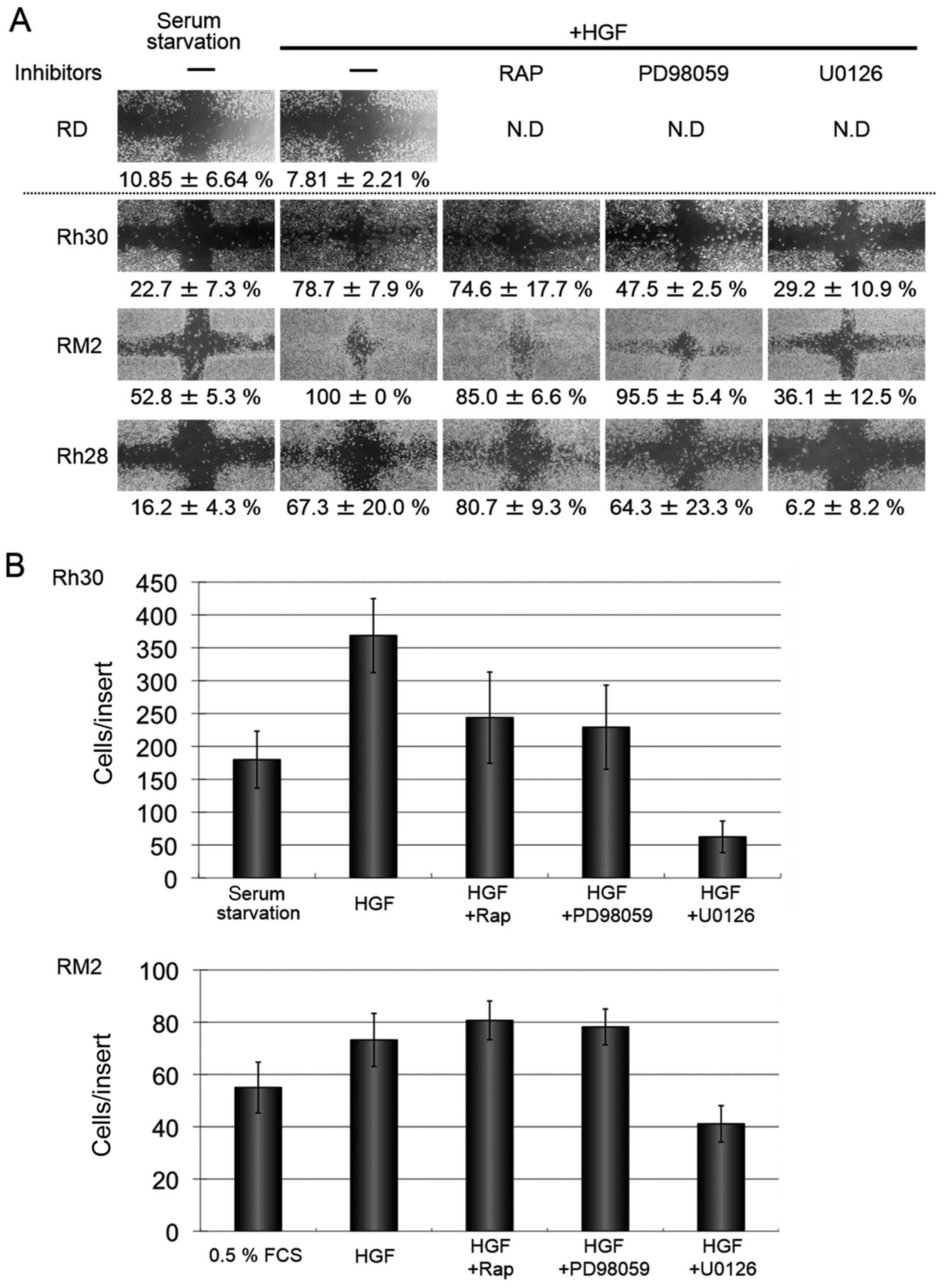
U0126 inhibits HGF-stimulated motility
of ARMS cell lines
The effect of HGF on the motility of the ARMS cell
lines, Rh30, RM2, and Rh28 was then examined by a wound-healing
assay. In 24 h after HGF stimulation, the percentage of the
distance between the wound edges in comparison to unstimulated
control cells were calculated to be 78.7±7.9, 100±0, and
67.3±20.0%, respectively. On the other hand, wound healing was not
observed in the ERMS cell line, RD (Fig. 3A). We then studied the effects of
several specific inhibitors against the potent signaling downstream
of MET on the HGF-stimulated motility. Motility of the three ARMS
cells upon HGF stimulation was not obviously inhibited by the mTOR
inhibitor, rapamycin (100 ng/ml) (74.6±17.7, 85.0±6.6 and
80.7±9.3%, respectively). When the MEK inhibitor, PD98059 (20 µM)
was used, motility was inhibited partially (47.5±2.5, 95.5±5.4 and
64.3±23.3%, respectively). However, another MEK inhibitor, U0126 at
the concentration of 20 µM could remarkably inhibit the motility of
the three ARMS cell lines, Rh30, RM2, and Rh28 (29.2±10.9,
36.1±12.5 and 6.2±8.2%, respectively) (Fig. 3A).
Further, we confirmed this phenomenon by a migration
assay. The number of migrating cells upon HGF stimulation was
increased in ARMS cell lines, Rh30 and RM2 (369±56 cells and 73±10
cells, respectively) compared with unstimulated controls (179±43
cells and 55±9 cells, respectively). HGF-stimulated migration was
not completely inhibited by rapamycin (100 ng/ml) (244±69 cells and
81±7 cells, respectively) or by PD98059 (20 µM) (229±64 cells and
78±1 cells, respectively). However, it was markedly inhibited by
U0126 at 20 µM (Fig. 3B) (63±24
cells and 41±7 cells, respectively).
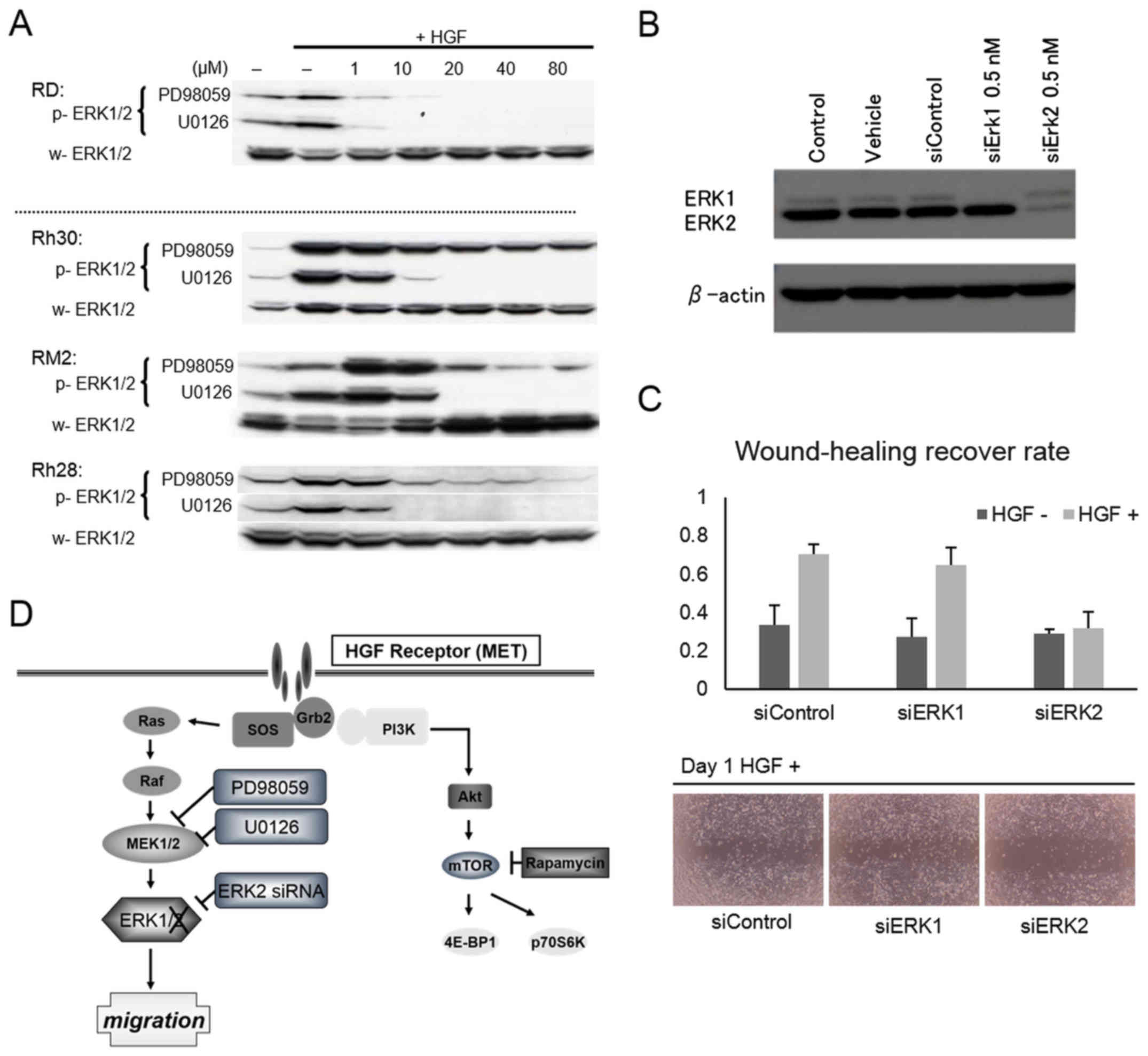
ERK1/2 in RMS cells was completely
dephosphorylated by U0126
In order to investigate differences in the
inhibitory effects of MEK inhibitors PD98059 and U0126 on the
motility of ARMS cells, ERK1/2 phosphorylation status was compared
in the ERMS cell line RD and the ARMS cell lines Rh30, RM2, and
Rh28. In all three ARMS cell lines even 20 µM of PD98059 did not
completely inhibit ERK2 phosphorylation, whereas the same
concentration of PD98059 completely blocked ERK2 phosphorylation
(Fig. 4A) in ERMS RD cells. On the
other hand, 20 µM of U0126 completely blocked ERK2 phosphorylation
in ERMS as well as ARMS cells (Fig.
4A).
HGF-stimulated motility of Rh30
stimulation is inhibited by ERK2 siRNA
In order to confirm the role of ERK2 in promoting
HGF-stimulated mobility of ARMS cells, we knocked down ERK1 or ERK2
expression using specific siRNA in the ARMS Rh30 cells. ERK1 and
ERK2 protein levels were sufficiently suppressed by siRNA
transfection (Fig. 4B). When the
wounding assay was performed using the siRNA transfected cells, the
percentages of wound-healing recovery for HGF-stimulated cells with
the control siRNA, ERK1 siRNA, and ERK2 siRNA were calculated to be
70.7±4.98, 65.0±9.17, and 32.1±8.01%, respectively. This indicated
that HGF-stimulated motility of Rh30 cells was not inhibited by the
knockdown of ERK1, but ERK2.
Discussion
Clinically, ARMS is well known to be more aggressive
than ERMS, probably due to the presence of the PAX3-FOXO1 fusion
protein (20). It has been reported
that MET is upregulated in ARMS cell lines that express PAX3-FOXO1
(10). Recently it was reported
that MET is regulated by the muscle-specific miR1/206 (21).
First, we confirmed that MET is more strongly
expressed in the PAX3-FOXO1-positive human ARMS cell lines Rh30,
RM2, and Rh28 than in the human ERMS cell lines RD, CT-11, and KH
(Fig. 1). We then studied the
biological significance of MET expression in ARMS cells. Unlike
observations in other cancers (8,9,22), HGF
stimulation did not promote RMS cell proliferation (Fig. 2). Since HGF and MET contribute to
motility in colon cancer, osteosarcoma, and small cell lung cancer
cell lines (23), we studied the
effect of HGF on motility in RMS cell lines using the wound-healing
assay and migration assays. Our findings, that HGF stimulated
motility in the three ARMS cell lines, but not in the ERMS RD cells
(Fig. 3), suggests that HGF/MET
signaling regulates the metastatic and invasive features of ARMS.
Ras-Raf-ERK1/2 and PI3K-AKT-mTOR pathways are well known to be
downstream of HGF/MET signaling (24,25).
We confirmed that HGF could phosphorylate ERK1/2 that is downstream
of MET, in all three ARMS cell lines suggesting that these pathways
are active in ARMS.
In order to determine the pathway responsible for
motility of ARMS cells, we used several inhibitors including the
mTOR inhibitor, rapamycin (26),
the ERK1 inhibitor, PD98059, and the ERK1/2 inhibitor, U0126. The
inhibitory effects of these compounds on the HGF-stimulated
motility of Rh30, Rh28, and RM2 cells, were studied by both the
wound-healing and migration assay. Our finding, that the motility
of all three cell lines was not inhibited by either rapamycin or by
PD98059, but was completely inhibited by U0126 (Fig. 3), indicates that ERK1/2 signaling
regulates motility of ARMS cells.
We next examined the HGF-stimulated phosphorylation
status of ERK1/2 upon treatment with PD98059 or U0126 at the same
concentrations used in the wounding and migration assays. At a
concentration of 20 µM, the ERK1 inhibitor PD98059 could not
completely dephosphorylate ERK2 (remaining lower band in western
blotting in Fig. 4A) in all three
ARMS cell lines. However, U0126, an inhibitor of both ERK1 and
ERK2, completely dephosphorylated ERK1 and ERK2 at a concentration
of 20 µM or higher (Fig. 4A).
According to all the above results, we hypothesized that ERK2
phosphorylation was responsible for the HGF-stimulated motility in
ARMS cells.
We then knocked down ERK2 expression by using a
specific siRNA against ERK2. The inhibitory effect on the mobility
of ARMS Rh30 cells was studied using an ERK2 siRNA designed with
confirmed efficacy (Fig. 4B).
Knockdown of ERK2 inhibited the wound-healing induced by HGF at a
concentration which did not stimulate cell growth in Rh30 cells
(Fig. 4C). Inhibition of
wound-healing by ERK2 knockdown indicates that ERK2 is a key
mediator for the motility of ARMS Rh30 cells. The isoform
specificity of ERK1 and ERK2 has not been widely addressed. Indeed
several studies have shown redundant functions for ERK1 and ERK2
(27,28), but it has been recently reported
that ERK2, but not ERK1, mediates cell motility (29,30),
which is consistent with our observations (Fig. 4B and C).
Our findings are summarized in Fig. 4D. PAX3-FOXO1 upregulates the HGF
receptor MET, in ARMS cells. HGF stimulates motility of ARMS cells
mainly through the ERK1/2 pathway. ERK2 phosphorylation
particularly appears to be the most critical event in ERK1/2
signaling for cell motility in the more aggressive ARMS cells.
HGF/MET has been reported to be involved in
tumorigenicity in various cell types (6). The possibility of modifying tumor
metastasis by inducing angiogenesis has also been suggested
(31,32). Previous studies have reported that
the levels of HGF in serum, amplification of MET, or MET
overexpression were clinically associated with poor prognosis for
some cancers (33,34). Even in RMS, overexpression of MET as
well as expression of PAX3-FOXO1 are closely associated with
malignancy, progression, and invasion (13,35,36).
In addition, HGF promotes the metastatic behavior of
MET- positive RMS cell lines to the bone marrow or lymph nodes and
contribute to their resistance to Radio-chemotherapy (37). The existence of metastasis is an
important poor prognostic factor in patients with PAX3-FOXO1
positive ARMS (38). Furthermore,
the frequency of PAX3-FOXO1 positive cells in metastatic ARMS
tumors is higher than that in the primary ARMS tumors in
preclinical mouse models (39).
These reports and our results collectively suggest that the poor
outcome of ARMS results from metastasis induced by high expression
of PAX3-FOXO1 and MET.
In this study, our results suggest that the
MET-ERK1/2 pathway, especially the phosphorylation of ERK2 plays a
major role in cell motility of ARMS. Selective inhibitors against
this pathway, and particularly against ERK2 phosphorylation, might
be a novel anticancer agent to regulate the invasive and metastatic
characteristics of ARMS.
Acknowledgements
This work was supported by JSPS KAKENHI grant number
25253095.
References
|
1
|
Kikuchi K, Rubin BP and Keller C:
Developmental origins of fusion-negative rhabdomyosarcomas. Curr
Top Dev Biol. 96:33–56. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kikuchi K, Hettmer S, Aslam MI, Michalek
JE, Laub W, Wilky BA, Loeb DM, Rubin BP, Wagers AJ and Keller C:
Cell-cycle dependent expression of a translocation-mediated fusion
oncogene mediates checkpoint adaptation in rhabdomyosarcoma. PLoS
Genet. 10:e10041072014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kikuchi K, Taniguchi E, Chen HI, Svalina
MN, Abraham J, Huang ET, Nishijo K, Davis S, Louden C, Zarzabal LA,
et al: Rb1 loss modifies but does not initiate alveolar
rhabdomyosarcoma. Skelet Muscle. 3:272013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sorensen PH, Lynch JC, Qualman SJ,
Tirabosco R, Lim JF, Maurer HM, Bridge JA, Crist WM, Triche TJ and
Barr FG: PAX3-FKHR and PAX7-FKHR gene fusions are prognostic
indicators in alveolar rhabdomyosarcoma: A report from the
children's oncology group. J Clin Oncol. 20:2672–2679. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Epstein JA, Shapiro DN, Cheng J, Lam PY
and Maas RL: Pax3 modulates expression of the c-Met receptor during
limb muscle development. Proc Natl Acad Sci USA. 93:4213–4218.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Christensen JG, Burrows J and Salgia R:
c-Met as a target for human cancer and characterization of
inhibitors for therapeutic intervention. Cancer Lett. 225:1–26.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Takayama H, La Rochelle WJ, Anver M,
Bockman DE and Merlino G: Scatter factor/hepatocyte growth factor
as a regulator of skeletal muscle and neural crest development.
Proc Natl Acad Sci USA. 93:5866–5871. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Coltella N, Manara MC, Cerisano V,
Trusolino L, Di Renzo MF, Scotlandi K and Ferracini R: Role of the
MET/HGF receptor in proliferation and invasive behavior of
osteosarcoma. FASEB J. 17:1162–1164. 2003.PubMed/NCBI
|
|
9
|
Koon EC, Ma PC, Salgia R, Welch WR,
Christensen JG, Berkowitz RS and Mok SC: Effect of a
c-Met-specific, ATP-competitive small-molecule inhibitor SU11274 on
human ovarian carcinoma cell growth, motility, and invasion. Int J
Gynecol Cancer. 18:976–984. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ginsberg JP, Davis RJ, Bennicelli JL,
Nauta LE and Barr FG: Up-regulation of MET but not neural cell
adhesion molecule expression by the PAX3-FKHR fusion protein in
alveolar rhabdomyosarcoma. Cancer Res. 58:3542–3546.
1998.PubMed/NCBI
|
|
11
|
Sharp R, Recio JA, Jhappan C, Otsuka T,
Liu S, Yu Y, Liu W, Anver M, Navid F, Helman LJ, et al: Synergism
between INK4a/ARF inactivation and aberrant HGF/SF signaling in
rhabdomyosarcomagenesis. Nat Med. 8:1276–1280. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ferracini R, Olivero M, Di Renzo MF,
Martano M, De Giovanni C, Nanni P, Basso G, Scotlandi K, Lollini PL
and Comoglio PM: Retrogenic expression of the MET proto-oncogene
correlates with the invasive phenotype of human rhabdomyosarcomas.
Oncogene. 12:1697–1705. 1996.PubMed/NCBI
|
|
13
|
Kikuchi K, Tsuchiya K, Otabe O, Gotoh T,
Tamura S, Katsumi Y, Yagyu S, Tsubai-Shimizu S, Miyachi M, Iehara
T, et al: Effects of PAX3-FKHR on malignant phenotypes in alveolar
rhabdomyosarcoma. Biochem Biophys Res Commun. 365:568–574. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lukasiewicz E, Miekus K, Kijowski J,
Drabik G, Wilusz M, Bobis-Wozowicz S and Majka M: Inhibition of
rhabdomyosarcoma's metastatic behavior through downregulation of
MET receptor signaling. Folia Histochem Cytobiol. 47:485–489.
2009.PubMed/NCBI
|
|
15
|
Miekus K, Lukasiewicz E, Jarocha D, Sekula
M, Drabik G and Majka M: The decreased metastatic potential of
rhabdomyosarcoma cells obtained through MET receptor downregulation
and the induction of differentiation. Cell Death Dis. 4:e4592013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Skrzypek K, Kusienicka A, Szewczyk B,
Adamus T, Lukasiewicz E, Miekus K and Majka M: Constitutive
activation of MET signaling impairs myogenic differentiation of
rhabdomyosarcoma and promotes its development and progression.
Oncotarget. 6:31378–31398. 2015.PubMed/NCBI
|
|
17
|
Tsuchiya K, Hosoi H, Misawa-Furihata A,
Houghton PJ and Sugimoto T: Insulin-like growth factor-I has
different effects on myogenin induction and cell cycle progression
in human alveolar and embryonal rhabdomyosarcoma cells. Int J
Oncol. 31:41–47. 2007.PubMed/NCBI
|
|
18
|
Hayashi Y, Sugimoto T, Horii Y, Hosoi H,
Inazawa J, Kemshead JT, Inaba T, Hanada R, Yamamoto K, Gown AM, et
al: Characterization of an embryonal rhabdomyosarcoma cell line
showing amplification and over-expression of the N-myc oncogene.
Int J Cancer. 45:705–711. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Taulli R, Scuoppo C, Bersani F, Accornero
P, Forni PE, Miretti S, Grinza A, Allegra P, Schmitt-Ney M,
Crepaldi T, et al: Validation of met as a therapeutic target in
alveolar and embryonal rhabdomyosarcoma. Cancer Res. 66:4742–4749.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yan D, Dong XE, Chen X, Wang L, Lu C, Wang
J, Qu J and Tu L: MicroRNA-1/206 targets c-Met and inhibits
rhabdomyosarcoma development. J Biol Chem. 284:29596–29604. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Marampon F, Bossi G, Ciccarelli C, Di
Rocco A, Sacchi A, Pestell RG and Zani BM: MEK/ERK inhibitor U0126
affects in vitro and in vivo growth of embryonal rhabdomyosarcoma.
Mol Cancer Ther. 8:543–551. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ma PC, Tretiakova MS, Nallasura V,
Jagadeeswaran R, Husain AN and Salgia R: Downstream signalling and
specific inhibition of c-MET/HGF pathway in small cell lung cancer:
Implications for tumour invasion. Br J Cancer. 97:368–377. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Paumelle R, Tulasne D, Kherrouche Z, Plaza
S, Leroy C, Reveneau S, Vandenbunder B and Fafeur V: Hepatocyte
growth factor/scatter factor activates the ETS1 transcription
factor by a RAS-RAF-MEK-ERK signaling pathway. Oncogene.
21:2309–2319. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Okano J, Shiota G, Matsumoto K, Yasui S,
Kurimasa A, Hisatome I, Steinberg P and Murawaki Y: Hepatocyte
growth factor exerts a proliferative effect on oval cells through
the PI3K/AKT signaling pathway. Biochem Biophys Res Commun.
309:298–304. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hosoi H, Dilling MB, Shikata T, Liu LN,
Shu L, Ashmun RA, Germain GS, Abraham RT and Houghton PJ: Rapamycin
causes poorly reversible inhibition of mTOR and induces
p53-independent apoptosis in human rhabdomyosarcoma cells. Cancer
Res. 59:886–894. 1999.PubMed/NCBI
|
|
27
|
Srinivasan R, Zabuawala T, Huang H, Zhang
J, Gulati P, Fernandez S, Karlo JC, Landreth GE, Leone G and
Ostrowski MC: Erk1 and Erk2 regulate endothelial cell proliferation
and migration during mouse embryonic angiogenesis. PLoS One.
4:e82832009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Voisin L, Saba-El-Leil MK, Julien C,
Frémin C and Meloche S: Genetic demonstration of a redundant role
of extracellular signal-regulated kinase 1 (ERK1) and ERK2
mitogen-activated protein kinases in promoting fibroblast
proliferation. Mol Cell Biol. 30:2918–2932. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
von Thun A, Birtwistle M, Kalna G,
Grindlay J, Strachan D, Kolch W, von Kriegsheim A and Norman JC:
ERK2 drives tumour cell migration in three-dimensional
microenvironments by suppressing expression of Rab17 and liprin-β2.
J Cell Sci. 125:1465–1477. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Radtke S, Milanovic M, Rossé C, De Rycker
M, Lachmann S, Hibbert A, Kermorgant S and Parker PJ: ERK2 but not
ERK1 mediates HGF-induced motility in non-small cell lung carcinoma
cell lines. J Cell Sci. 126:2381–2391. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sengupta S, Sellers LA, Li RC, Gherardi E,
Zhao G, Watson N, Sasisekharan R and Fan TP: Targeting of
mitogen-activated protein kinases and phosphatidylinositol 3 kinase
inhibits hepatocyte growth factor/scatter factor-induced
angiogenesis. Circulation. 107:2955–2961. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ren Y, Cao B, Law S, Xie Y, Lee PY, Cheung
L, Chen Y, Huang X, Chan HM, Zhao P, et al: Hepatocyte growth
factor promotes cancer cell migration and angiogenic factors
expression: a prognostic marker of human esophageal squamous cell
carcinomas. Clin Cancer Res. 11:6190–6197. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee J, Seo JW, Jun HJ, Ki CS, Park SH,
Park YS, Lim HY, Choi MG, Bae JM, Sohn TS, et al: Impact of MET
amplification on gastric cancer: Possible roles as a novel
prognostic marker and a potential therapeutic target. Oncol Rep.
25:1517–1524. 2011.PubMed/NCBI
|
|
34
|
Aune G, Lian AM, Tingulstad S, Torp SH,
Forsmo S, Reseland JE, Stunes AK and Syversen U: Increased
circulating hepatocyte growth factor (HGF): A marker of epithelial
ovarian cancer and an indicator of poor prognosis. Gynecol Oncol.
121:402–406. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen Y, Takita J, Mizuguchi M, Tanaka K,
Ida K, Koh K, Igarashi T, Hanada R, Tanaka Y, Park MJ, et al:
Mutation and expression analyses of the MET and CDKN2A genes in
rhabdomyosarcoma with emphasis on MET overexpression. Genes
Chromosomes Cancer. 46:348–358. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Rees H, Williamson D, Papanastasiou A,
Jina N, Nabarro S, Shipley J and Anderson J: The MET receptor
tyrosine kinase contributes to invasive tumour growth in
rhabdomyosarcomas. Growth Factors. 24:197–208. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jankowski K, Kucia M, Wysoczynski M, Reca
R, Zhao D, Trzyna E, Trent J, Peiper S, Zembala M, Ratajczak J, et
al: Both hepatocyte growth factor (HGF) and stromal-derived
factor-1 regulate the metastatic behavior of human rhabdomyosarcoma
cells, but only HGF enhances their resistance to radiochemotherapy.
Cancer Res. 63:7926–7935. 2003.PubMed/NCBI
|
|
38
|
Breneman JC, Lyden E, Pappo AS, Link MP,
Anderson JR, Parham DM, Qualman SJ, Wharam MD, Donaldson SS, Maurer
HM, et al: Prognostic factors and clinical outcomes in children and
adolescents with metastatic rhabdomyosarcoma - a report from the
Intergroup Rhabdomyosarcoma Study IV. J Clin Oncol. 21:78–84. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Nishijo K, Chen QR, Zhang L, McCleish AT,
Rodriguez A, Cho MJ, Prajapati SI, Gelfond JA, Chisholm GB,
Michalek JE, et al: Credentialing a preclinical mouse model of
alveolar rhabdomyosarcoma. Cancer Res. 69:2902–2911. 2009.
View Article : Google Scholar : PubMed/NCBI
|