Introduction
Hepatocellular carcinoma (HCC) is one of the most
common malignancies worldwide, with a high mortality rate and
ominous prognosis, and is a serious public health threat (1,2). The
high rates of recurrence and metastasis after hepatectomy are major
causes of death in patients with HCC. However, the exact mechanisms
of recurrence and metastasis of HCC are still poorly understood
(3). Therefore, identifying novel
molecular markers in HCC and clarifying the molecular mechanisms
responsible for the recurrence and metastasis may be helpful for
the treatment of HCC and to improve the survival and prognosis of
HCC patients.
Semaphorin 3A (SEMA3A), a secretory protein, is a
founding member of the semaphorin family and has a Sema domain, a
PSI domain and an immunoglobulin-like domain (4). SEMA3A functions in autocrine and
paracrine manners (5). Numerous
studies have found that SEMA3A plays an important role in the
development and in related disorders of the nervous, immune,
cardiovascular and skeletal systems (6–8).
Recent studies also found that SEMA3A regulates the biological
behavior of tumor cells directly or affected them indirectly, by
influencing the tumor microenvironment (9,10).
SEMA3A is overexpressed in glioblastoma, pancreatic, lung and
prostate cancer (11–14). Its increased expression is
positively correlated with advanced malignant grade, cell
migration, metastatic potential and poor prognosis (12,15).
Furthermore, SEMA3A directly decreases focal adhesions, induces
cancer cell invasion in vitro, and modulates
tumor-associated macrophages (15,16).
However, the role of SEMA3A in HCC is not well established.
In the present study, we reported that SEMA3A serves
as a novel hallmark for HCC. We demonstrated that SEMA3A
significantly promotes HCC progression. Notably, our findings
indicate that SEMA3A promotes the proliferation and invasion of HCC
by upregulating the expression of gelsolin-like capping protein
(CapG), galectin-3, enolase 2 and epithelial cell adhesion molecule
(EpCAM). Ultimately, these findings contribute to our current
understanding of HCC progression.
Materials and methods
Patient and tissue specimens
Specimens of HCC and corresponding adjacent
non-tumorous liver tissues (ANLTs) were obtained from 80 HCC
patients undergoing curative hepatic resection at the Department of
Hepatic Surgery, Zhongshan Hospital of Fudan University from
January 2010 to December 2010. The diagnosis for each patient was
confirmed histopathologically. These tissue specimens were freshly
collected for real-time quantitative reverse-transcription
polymerase chain reaction (RT-qPCR) and western blot detection.
Prior informed consent was obtained, and the study protocol was
approved by the Ethics Committee of Zhongshan Hospital.
RNA isolation and RT-qPCR
Total RNA was extracted from cell lines and frozen
tissue specimens using the TRIzol reagent (Invitrogen, Carlsbad,
CA, USA). Complementary DNA synthesis was performed using the High
Capacity cDNA Reverse Transcription kit (Applied Biosystems, Foster
City, CA, USA) according to the manufacturer's instructions. The
mRNA expression in HCC cell lines, 80 cases of HCC tissues and
corresponding ANLTs was assessed by RT-qPCR using the Power
SYBR-Green PCR Master Mix kit on an ABI 7900HT Fast Real-Time PCR
instrument (both from Applied Biosystems). The primers used were as
follows: SEMA3A forward, 5′-GCAAACTATCAGAATGGGAAGA-3′ and reverse,
5′-GCTCCAACATACAGCCTACTCC-3′; GAPDH forward,
5′-AGGTCGGAGTCAACGGATTTG-3′ and reverse,
5′-GTGATGGCATGGACTGTGGT-3′. The relative mRNA expression level of
SEMA3A was normalized to GAPDH. The results were analyzed using the
2−ΔCt method, and the formula was: ΔCt =
CtSEMA3A - CtGAPDH. All of the experiments
were performed in triplicate.
Immunohistochemistry
Formalin-fixed paraffin-embedded tissue sections
were stained for SEMA3A using the avidin-biotin-peroxidase complex
system (Gene Tech, Shanghai, China) according to the manufacturer's
instructions. Briefly, after rehydration and microwave antigen
retrieval, rabbit anti-human polyclonal antibodies against SEMA3A
(1:200 dilution; Abcam, Cambridge, MA, USA) were applied to the
slides, and incubated at 4°C overnight. Then, the slides were
incubated with secondary antibody (Gene Tech) at 37°C for half an
hour. Staining was performed with diaminobenzidine (DAB), and
Mayer's hematoxylin was used for counterstaining. Negative control
slides were probed with bovine serum albumin (BSA) under the same
experimental conditions.
Western blotting
Total cell lysates were generated, and total protein
was extracted and separated using 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), followed by
transfer onto polyvinylidene fluoride (PVDF) membranes (Millipore,
Bedford, MA, USA). The membranes were washed and blocked, and then
incubated with rabbit anti-human polyclonal antibody against SEMA3A
(1:800 dilution), rabbit anti-human polyclonal antibody against
CapG (1:1,000), rabbit anti-human polyclonal antibody against
galectin-3 (1:1,000), rabbit anti-human polyclonal antibody against
enolase 2 (1:800), or rabbit anti-human polyclonal antibody against
EpCAM (1:1,000), and then with horseradish peroxidase
(HRP)-conjugated goat anti-rabbit IgG secondary antibody (1:3,000)
(all from Abcam). GAPDH was detected using rabbit anti-human
monoclonal antibody against GAPDH (1:2,000; Abcam) as a loading
control. All antibody bindings were detected by an enhanced
chemiluminescence (ECL) assay (Millipore). The bands were analyzed
using the Gel-Pro Analyzer 32 software.
Cell lines and cell culture
Five human HCC cell lines (PLC/PRF/5, HepG2,
MHCC97-L, MHCC97-H and HCCLM3) with different invasive and
metastatic potentials were used in the present study. PLC/PRF/5 and
HepG2 cell lines (human HCC cell lines with low metastatic
potential) were purchased from the Chinese Academy of Sciences
(Shanghai, China). HCC cell lines with stepwise metastatic
potential (MHCC97-L, MHCC97-H and HCCLM3, which have the same
genetic background, but different lung metastatic potentials) were
established at our institute (17,18).
These cell lines were routinely maintained and cultured in
Dulbecco's modified Eagle's medium (DMEM), supplemented with 10%
(v/v) fetal bovine serum (FBS) (both from Gibco, Carlsbad, CA, USA)
in a humidified incubator containing 5% CO2 at 37°C.
Cell transfection and clone
selection
The pSV40-GFP-SEMA3A and pGCSIL-GFP-shRNA-SEMA3A
lentiviral vectors were purchased from the Shanghai GeneChem Co.
(Shanghai, China), and the target shRNA sequence was as follows:
5′-TGTTTACAATGGTGGGAAA-3′. The pSV40-GFP-SEMA3A lentivirus was
transfected into PLC/PRF/5 cells with a low metastatic potential,
and the pGCSIL-GFP-shRNA-SEMA3A lentivirus was transfected into the
HCC cells with a high metastatic potential (HCCLM3). The pSV40-GFP
and pGCSIL-GFP lentiviral vectors were used as negative controls.
Then, stably transfected clones were validated by RT-qPCR and
western blotting for SEMA3A.
Cell proliferation, wound-healing and
Matrigel invasion assays
For the cell proliferation assay, cells (20,000
cells/ml) were dispensed in 100 µl aliquots into a 96-well plate.
At the indicated time-points, 10 µl of Cell Counting Kit-8 (CCK-8)
solution (Dojindo, Kumamoto, Japan) was added to each well, and the
plate was incubated for an additional 2 h. The absorbance at 450 nm
was assessed to determine the number of viable cells in each
well.
For the wound-healing assay, cells were seeded into
3.5 cm dishes and cultured for 1–2 days. When the cells were 100%
confluent and formed a monolayer, the medium was changed to serum
starve the cells. After serum starvation, the cell monolayer was
wounded with a plastic pipette tip. The remaining cells were
carefully washed with culture medium to remove the cell debris, and
cultured in an incubator at 37°C for an additional 48 h. At the
indicated time-points, migrating cells at the wound front were
photographed using an inverted microscope (Olympus, Tokyo, Japan).
The percentage of the wound closure at each time-point compared
with time zero was assessed using Image-Pro Plus v6.2 software.
Matrigel invasion assays were performed using
24-well Transwell plates (8-µm pore size; Corning, Corning, NY,
USA). First, ~5×104 cells were suspended in 200 µl of
DMEM with 1% (v/v) FBS, and were placed into the upper chamber of
the insert pre-coated with Matrigel (BD Biosciences, Franklin
Lakes, NJ, USA), and 500 µl of DMEM with 10% (v/v) FBS was added to
the lower chamber. After a 48-h incubation, the Matrigel and the
cells remaining in the upper chamber were removed with cotton
swabs. Then, the cells that passed through and adhered onto the
lower surface of the membrane were fixed in 4% paraformaldehyde for
15 min, and stained with crystal violet. The cells were counted and
photographed in at least 5 random microscopic fields
(magnification, ×200). All of the experiments were performed in
triplicate.
Proteome profiler antibody array
assay
The Proteome Profiler™ Human XL Oncology Array
(ARY026) was purchased from the R&D Systems, Inc. (Minneapolis,
MN, USA). Proteome profiler antibody microarray analysis was
performed according to the protocol provided. Briefly, relative
expression levels of human cancer-related proteins in samples
(total proteins extracted from PLC/PRF/5-SEMA3A or
PLC/PRF/5-control cell lysates) were determined using the following
procedure: each array membrane was placed in a separate well of the
4-well Multi-Dish with 2 ml of array buffer 6, and incubated for 1
h on a rocking platform shaker; then, the array buffer 6 in each
well of the 4-well Multi-Dish was replaced with prepared samples
(0.5 ml total protein + 0.5 ml array buffer 4 + 0.5 ml array buffer
6), and the array membrane was incubated overnight at 4°C. The
following day, each membrane was washed 3 times with 1X washing
buffer, and then incubated in 1.5 ml of array buffer 4/6 with 30 µl
of a detection antibody cocktail for 1 h. Subsequently, each
membrane was washed 3 times again, and then incubated in 2 ml of
diluted streptavidin-HRP for 30 min. The membranes were washed an
additional 3 times, and then evenly covered with 1 ml of the
prepared Chemi Reagent Mix and incubated for 1 min. Ultimately, the
array membranes were exposed in the Tanon-5200 Chemiluminescent
Imaging System (Tanon, Shanghai, China), and analyzed with the
Gel-Pro Analyzer 32 software. The fluorescence intensity of each
spot on the array membranes was quantified, and the mean value was
calculated.
HCC metastatic mouse model
Approximately 5×106 cells of the
different groups (HCCLM3-shRNA-SEMA3A, HCCLM3-control,
PLC/PRF/5-SEMA3A and PLC/PRF/5-control) were suspended in 100 µl of
DMEM, and then subcutaneously injected into the upper left flank
regions of nude mice (BALB/c-nu/nu, male, 4 weeks old; 3 in each
group). After ~4 weeks, when the subcutaneous tumor size reached ~1
cm in diameter, the subcutaneous tumor tissues were removed, minced
into small pieces of equal volume (~2×2×2 mm3), and then
implanted into the livers of other nude mice (BALB/c-nu/nu, male, 4
weeks old; 6 in each group). All of the mice were monitored on
alternate days and sacrificed 6 weeks later. All of the livers of
the mice were removed, and the volumes of liver tumors were
calculated as follows: V (cm3) = 3ab2/4 (a
and b, respectively represent the largest and smallest tumor
diameters measured by vernier caliper) (19). All of the lungs were also removed,
fixed in 4% paraformaldehyde for 2–3 days, embedded in paraffin,
serially sectioned and stained with hematoxylin and eosin. The
total number of lung metastases was examined and counted under a
microscope, and the lung metastatic lesions were classified into 4
grades (grade I, ≤20; grade II, 20–50 tumor cells; grade III,
50–100; and grade IV, >100 tumor cells) on the basis of the
number of tumor cells present at the maximal section for each
metastatic lesion as previously described (20). All of the mice used in the present
study, were purchased from the Chinese Academy of Sciences, and
housed under specific pathogen-free (SPF) conditions. All of the
mice received humane care and were manipulated according to the
protocols (approved by the Medical Experimental Animal Care
Commission) and the criteria outlined in the ‘Guide for the Care
and Use of Laboratory Animals’ (prepared by the National Academy of
Sciences and published by the National Institutes of Health, NIH
publication 86–23 revised 1985).
Statistical analysis
All of the data were analyzed using IBM SPSS
Statistics for Windows (version 19.0.0). Values are expressed as
the mean ± SD. Quantitative data were compared between groups using
Student's t-test. Categorical data were analyzed using the
Chi-square or Fisher's exact tests. All of the statistical analyses
used a two-sided test, with P<0.05 as statistically
significant.
Results
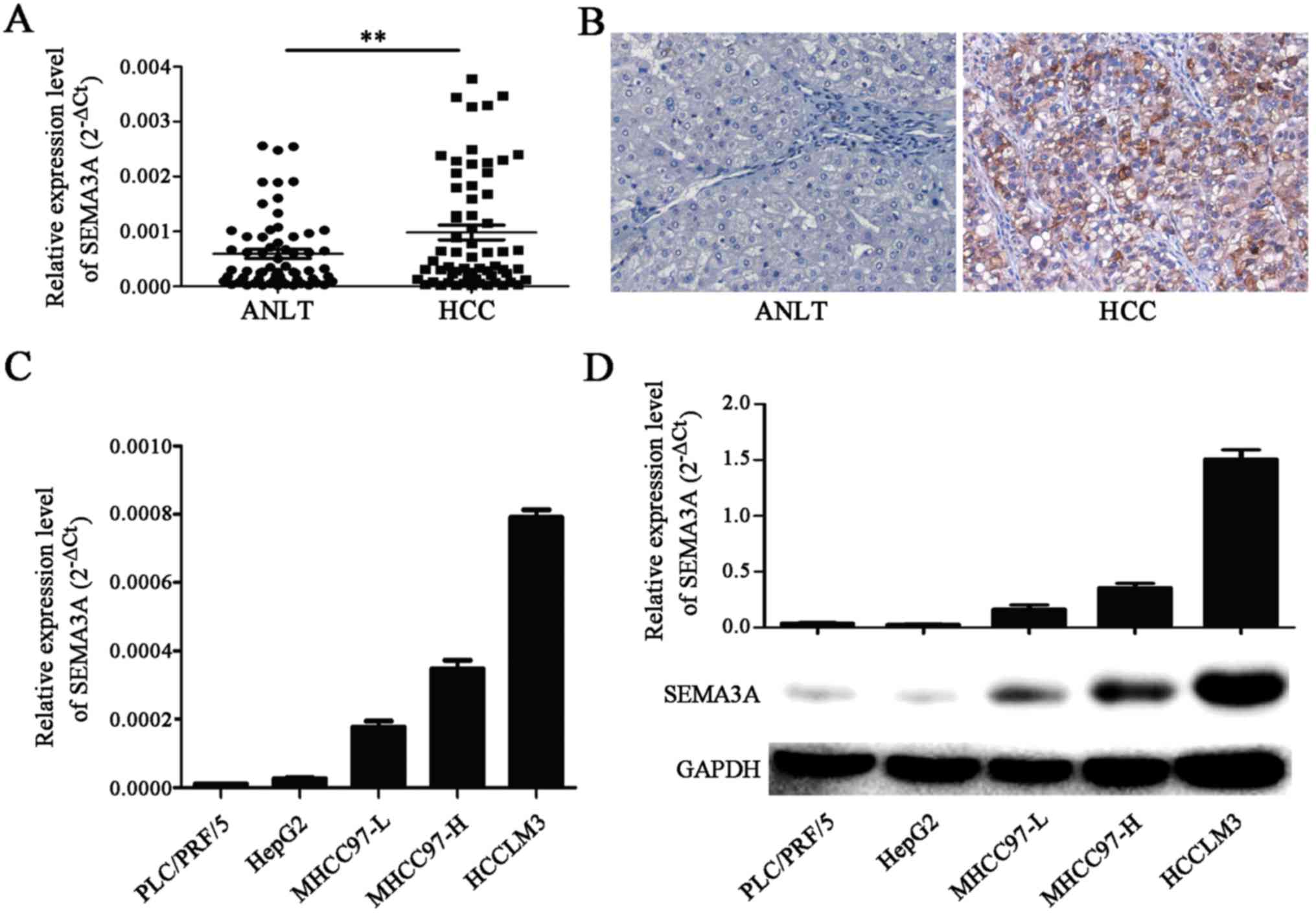
SEMA3A is upregulated in human HCC
tissues
Both the mRNA and protein expression levels of
SEMA3A were detected by RT-qPCR and immunohistochemistry in 80
cases of HCC and the corresponding ANLTs. RT-qPCR revealed that
SEMA3A was significantly overexpressed in the HCC tissues when
compared to the ANLTs (P<0.01) (Fig.
1A). The results were confirmed by immunohistochemical analyses
on the aforementioned 80 samples (Fig.
1B). These data demonstrated that SEMA3A is highly expressed in
HCC.
The expression level of SEMA3A is
associated with the invasive and metastatic potential of HCC cell
lines
The expression levels of SEMA3A in 5 established HCC
cell lines were assessed by RT-qPCR and western blotting. The
results revealed that both the mRNA and protein expression levels
of SEMA3A were significantly higher in the highly invasive and
metastatic HCC cell lines (MHCC97-L, MHC97-H and HCCLM3) than in
the HCC cell lines with low invasive and metastatic potential
(PLC/PRF/5 and HepG2; P<0.05) (Fig.
1C and D). These findings indicate that the expression of
SEMA3A is positively correlated with the invasive and metastatic
potential of HCC cells.
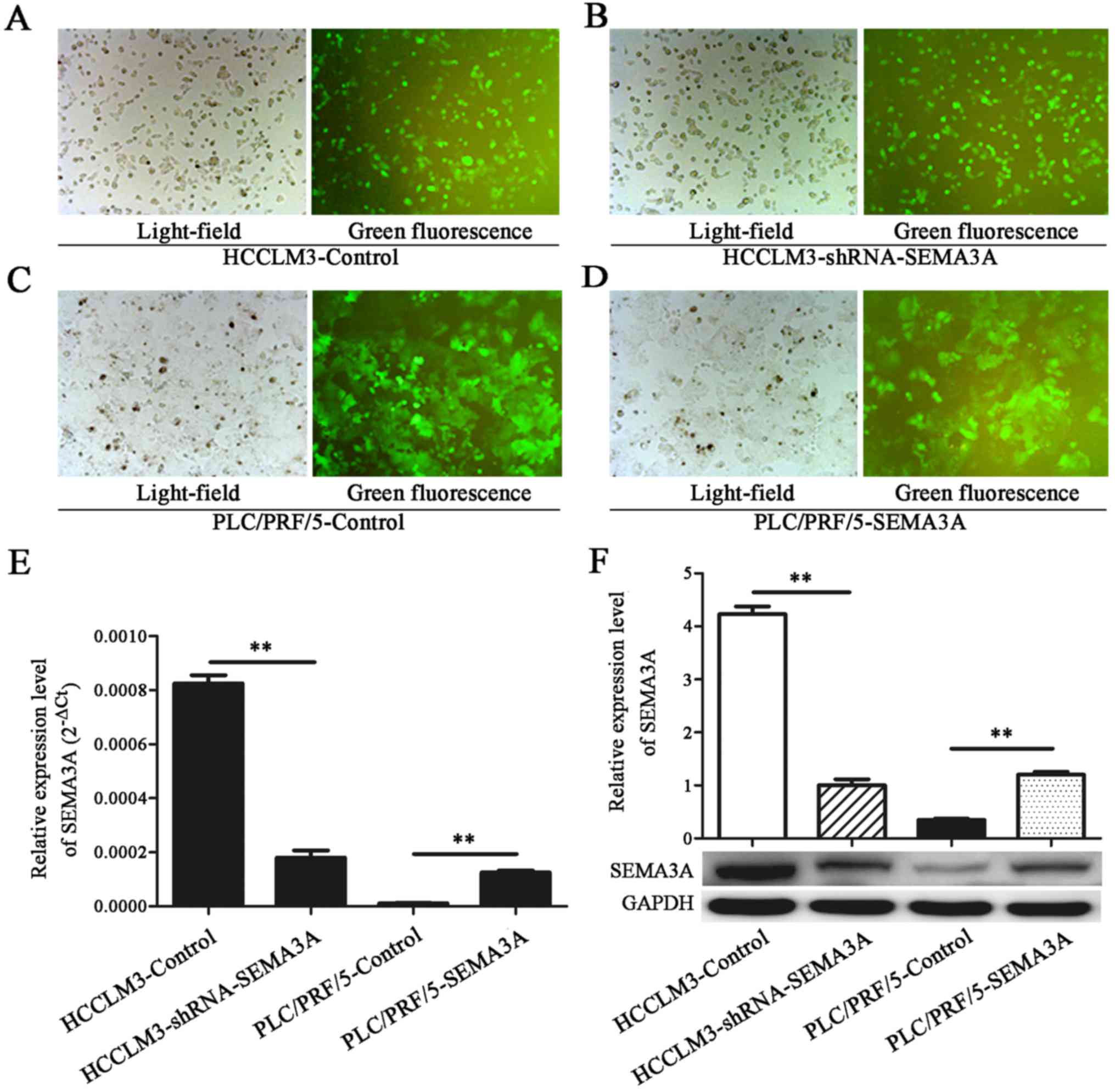
SEMA3A promotes proliferation,
migration and invasion of HCC cells in vitro
To further explore the biological functions of
SEMA3A in HCC, we used the lentiviral transfection approach to
stably upregulate or downregulate the expression of SEMA3A in the
transfected HCC cell lines (Fig.
2A-D). SEMA3A expression was ascertained by RT-qPCR and western
blotting (Fig. 2E and F).
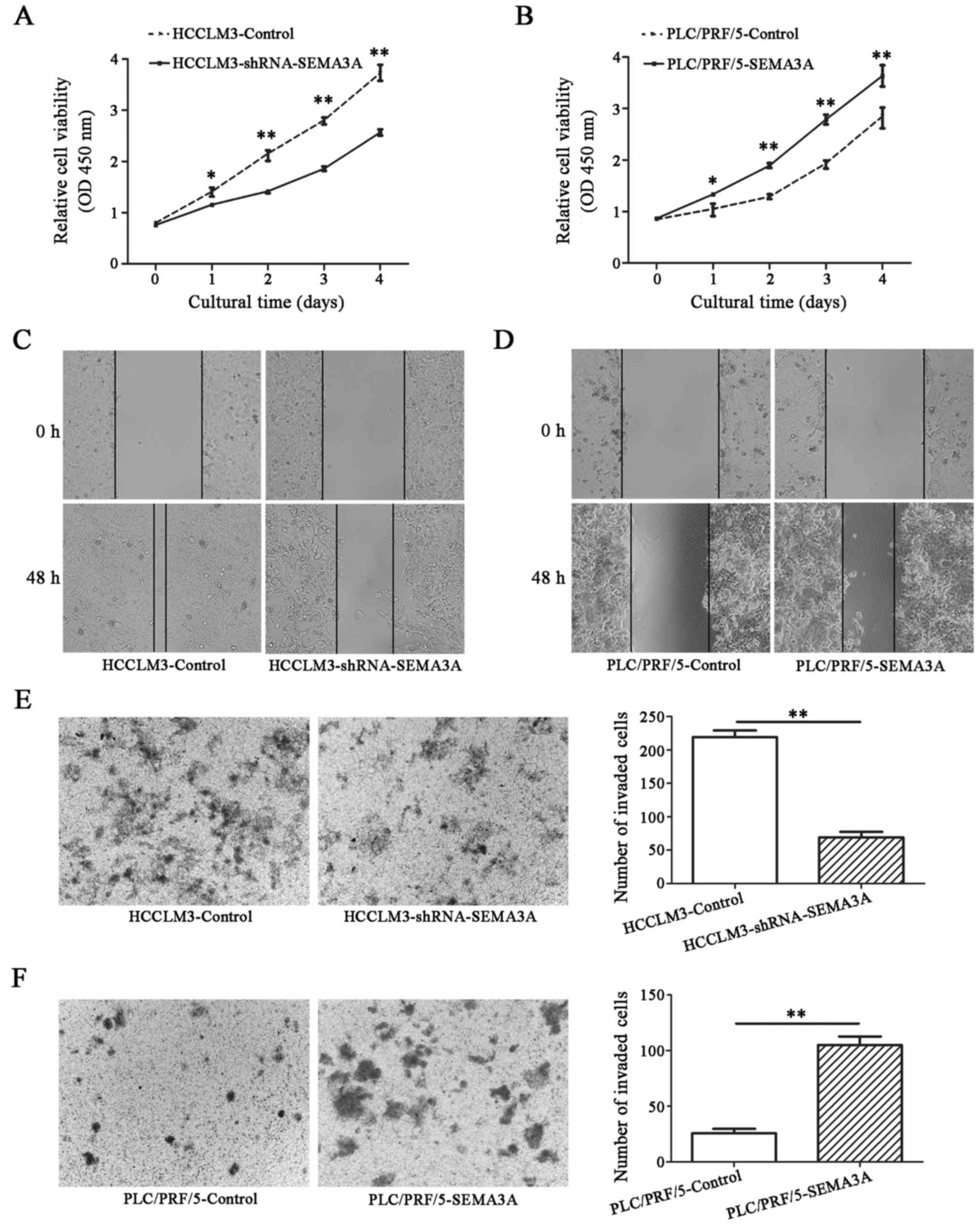
Downregulation of SEMA3A by shRNA in HCCLM3 cells significantly
suppressed cell proliferation after 24 h (P<0.05) (Fig. 3A). In contrast, cell proliferation
of PLC/PRF/5-SEMA3A cells was significantly higher than that of the
PLC/PRF/5-control cells (P<0.05) (Fig. 3B).
Furthermore, the migration and invasion abilities of
the HCCLM3-shRNA-SEMA3A and PLC/PRF/5-SEMA3A cells, and the
corresponding control cell lines were investigated using
wound-healing and invasion assays. In the wound-healing assays,
microscopic examination at 48 h post-wounding revealed a
significant delay in the wound closure rate of the
HCCLM3-shRNA-SEMA3A cells compared with the HCCLM3-control cells
(29.4±3.1 vs. 91.5±4.5%, respectively; P<0.01) (Fig. 3C), whereas PLC/PRF/5-SEMA3A cells
had a significant increase in the wound closure rate compared with
the control cell line (43.1±3.8 vs. 19.9±2.9%, respectively;
P<0.01) (Fig. 3D). In addition,
invasion assays revealed that the number of invasive
HCCLM3-shRNA-SEMA3A cells was significantly less than that of the
invasive HCCLM3-control cells (69.0±8.3 vs. 219.3±9.8,
respectively; P<0.01) (Fig. 3E)
while PLC/PRF/5-SEMA3A cells were significantly more invasive than
the PLC/PRF/5-control cells (105.0±7.6 vs. 25.7±4.1, respectively;
P<0.01) (Fig. 3F).
SEMA3A upregulates the expression of
CapG, galectin-3, enolase 2 and EpCAM in HCC cells
To explore which signaling pathways contributed to
HCC proliferation and invasion induced by SEMA3A, PLC/PRF/5-SEMA3A
and PLC/PRF/5-control cells were investigated for the activation
status of multiple pathways, which was determined by Proteome
Profiler Human XL Oncology Array assays. Notably, we found that
overexpression of SEMA3A in PLC/PRF/5 cells caused a significant
increase in the expression levels of CapG, galectin-3, enolase 2
and EpCAM (P<0.05) (Fig. 4A and
B). Furthermore, western blot analyses confirmed that SEMA3A
upregulated the expression of CapG, galectin-3, enolase 2 and EpCAM
in the PLC/PRF/5 cells, and the downregulation of SEMA3A by shRNA
in HCCLM3 cells caused a distinct decrease in those cancer-related
proteins (Fig. 4C).
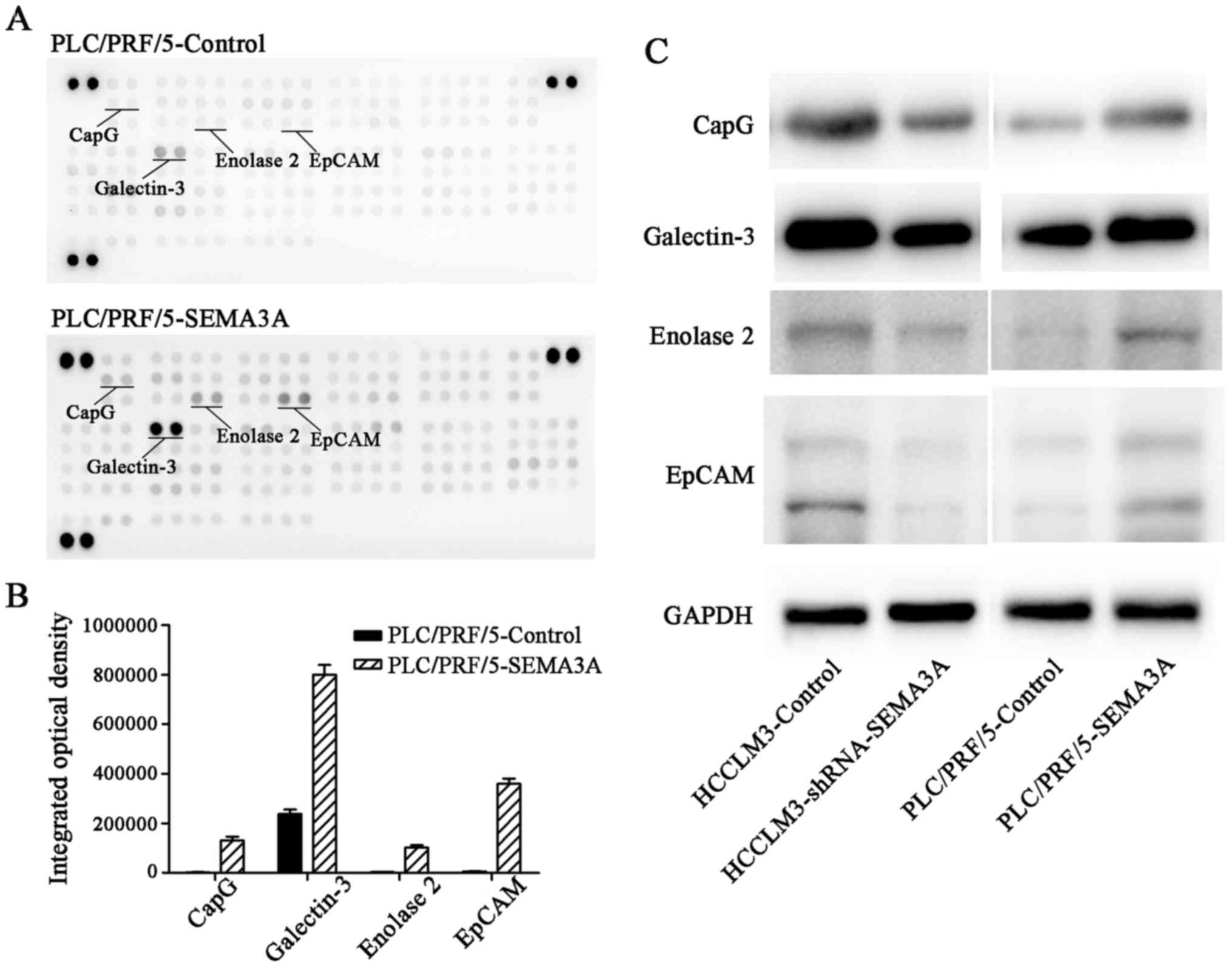 | Figure 4.SEMA3A increases CapG, galectin-3,
enolase 2 and EpCAM in HCC. (A and B) Proteome profiler antibody
array assay revealed that CapG, galectin-3, enolase 2 and EpCAM
were significantly overexpressed in the PLC/PRF/5-SEMA3A cells
(P<0.05). (C) Western blotting ascertained that the expression
levels of CapG, galectin-3, enolase 2 and EpCAM were markedly
decreased in the HCCLM3-shRNA-SEMA3A cells, but increased in the
PLC/PRF/5-SEMA3A cells. SEMA3A, semaphorin 3A; HCC, hepatocellular
carcinoma; CapG, gelsolin-like capping protein; EpCAM, epithelial
cell adhesion molecule. |
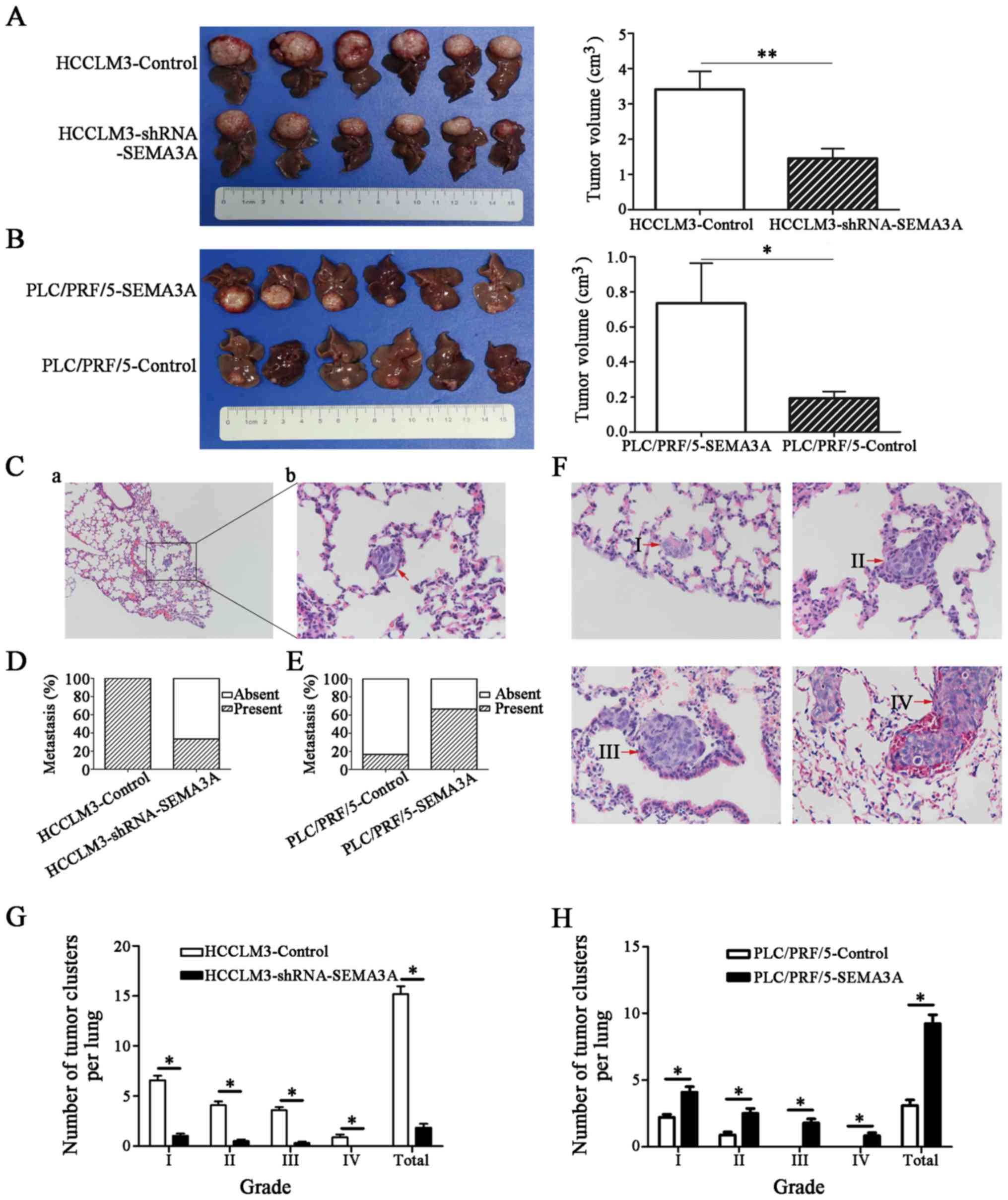
SEMA3A promotes the growth and
metastasis of HCC in vivo
To validate the observations obtained from the in
vitro studies, we used a metastatic HCC model in mice to
clarify the effects of SEMA3A on tumor growth and metastasis of HCC
in vivo. After orthotopic transplantation of subcutaneous
tumor tissues derived from the HCC cell lines into the livers of
nude mice, all of the groups successfully formed liver tumors. We
found that the liver localized tumor size in the
HCCLM3-shRNA-SEMA3A group was markedly smaller than that in the
HCCLM3-control group (1.45±0.28 vs. 3.41±0.51 cm3,
respectively; P<0.01) (Fig. 5A).
Similarly, the liver localized tumor size in the PLC/PRF/5-SEMA3A
group was 0.75±0.19 cm3, significantly larger than that
in the PLC/PRF/5-control group (0.21±0.03 cm3;
P<0.05) (Fig. 5B). Pulmonary
metastasis occurred in 100% (6/6) of the HCCLM3-control nude mice
(Fig. 5C), much higher than was
observed in the HCCLM3-shRNA-SEMA3A mice (2 out of 6, 33%)
(Fig. 5D). Furthermore, the number
of pulmonary metastatic nodules of each grade was also greater in
the HCCLM3-control mice (P<0.05) (Fig. 5F and G). In addition, only one of
the PLC/PRF/5-control mice had pulmonary metastasis (1 out of 6,
17%), a distinctly lower rate than that observed in the
PLC/PRF/5-SEMA3A group (4 out of 6, 67%) (Fig. 5E). Moreover, the number of pulmonary
metastatic nodules of each grade was also obviously smaller in the
PLC/PRF/5-control group (P<0.05) (Fig. 5H). Collectively, these data
demonstrate an important role for SEMA3A in HCC progression and
metastasis.
Discussion
Hepatocellular carcinoma (HCC) is one of the most
lethal cancers, and few effective therapeutic methods are currently
available (21). Understanding the
molecular mechanisms for HCC progression provides strategies for
the intervention and treatment of HCC and improves the survival and
prognosis of HCC patients (22). In
the present study, we found that the expression of SEMA3A was
elevated in HCC tissues compared with the corresponding adjacent
non-tumorous liver tissues (ANLTs). These findings agree with the
results derived from other solid tumors, including pancreatic,
lung, and prostate cancers (12,13,15).
Therefore, clarifying the effect and mechanism of SEMA3A on HCC
provides the basis for further research and is helpful for
developing a novel, effective and practical diagnostic and
therapeutic method for HCC.
Recent studies reported that increased expression of
SEMA3A is positively related to advanced malignant grade, cell
migration and metastatic potential (12,15).
However, it remains unclear whether SEMA3A contributes to the
progression of HCC. In the present study, we confirmed for the
first time that both SEMA3A mRNA and protein are significantly
overexpressed in HCC tissues compared with the corresponding ANLTs,
which indicates that SEMA3A may play an important role in HCC
progression. Moreover, we demonstrated the expression of SEMA3A in
HCC cell lines and found that SEMA3A expression was positively
correlated with the invasive and metastatic potential of HCC cells.
Our data established for the first time that SEMA3A is upregulated
in HCC and may play a critical role in the invasion and metastasis
of HCC.
To gain insight on the effect of SEMA3A on the
biological behavior of HCC cells, we transfected the corresponding
lentivirus into HCC cells to knock down the SEMA3A expression in
HCCLM3 cells and overexpress SEMA3A in PLC/PRF/5 cells. We found
that upregulation of SEMA3A resulted in a marked promotion of cell
proliferation, migration and invasion of PLC/PRF/5 cells in
vitro, but depletion of SEMA3A led to obvious inhibition of
cell proliferation, migration and invasion in HCCLM3 cells. These
findings confirmed that SEMA3A promoted the proliferation and
invasion of HCC cells.
Current studies ascertained that SEMA3A, as a
secreted protein, interacts with neuropilins and plexins, adjusting
the expression of cancer-related proteins, to regulate cell shape,
adhesion, locomotion and invasion (23–26).
Our results indicated that the overexpression of SEMA3A upregulated
the expression of CapG, galectin-3, enolase 2 and EpCAM in
PLC/PRF/5 cells. Conversely, SEMA3A depletion attenuated these
proteins in HCCLM3 cells. Numerous studies have confirmed that
CapG, galectin-3, enolase 2 and EpCAM can induce tumor progression
(27–30). These results revealed that SEMA3A
promotes the proliferation and invasion of HCC cells by
facilitating CapG, galectin-3, enolase 2 and EpCAM expression.
The role of SEMA3A in the modulation of HCC cell
proliferation and invasion was further confirmed by in vivo
assays, in which we demonstrated that the overexpression of SEMA3A
was associated with an increased volume of orthotopic tumors.
Conversely, knockdown of SEMA3A inhibited orthotopic tumor growth.
Moreover, the number and size of pulmonary metastatic lesions in
the SEMA3A-overexpressing group were all significantly increased
compared with those in the control group, and depletion of SEMA3A
suppressed pulmonary metastasis in an HCC metastatic mouse model,
suggesting a critical role for SEMA3A in the regulation of the
metastasis of HCC.
In conclusion, the present study, reveals for the
first time that SEMA3A is overexpressed in HCC. Furthermore, we
demonstrated the critical role of SEMA3A in promoting HCC growth,
invasion, and metastasis by increasing the expression of CapG,
galectin-3, enolase 2 and EpCAM. Collectively, these features of
SEMA3A make it a potential therapeutic target for HCC.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81472218, 81272724,
81402376 and 81301821).
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ou DP, Yang LY, Huang GW, Tao YM, Ding X
and Chang ZG: Clinical analysis of the risk factors for recurrence
of HCC and its relationship with HBV. World J Gastroenterol.
11:2061–2066. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kolodkin AL, Matthes DJ, O'Connor TP,
Patel NH, Admon A, Bentley D and Goodman CS: Fasciclin IV:
Sequence, expression, and function during growth cone guidance in
the grasshopper embryo. Neuron. 9:831–845. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kolodkin AL, Matthes DJ and Goodman CS:
The semaphorin genes encode a family of transmembrane and secreted
growth cone guidance molecules. Cell. 75:1389–1399. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yazdani U and Terman JR: The semaphorins.
Genome Biol. 7:2112006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hung RJ, Yazdani U, Yoon J, Wu H, Yang T,
Gupta N, Huang Z, van Berkel WJ and Terman JR: Mical links
semaphorins to F-actin disassembly. Nature. 463:823–827. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ito T, Kagoshima M, Sasaki Y, Li C, Udaka
N, Kitsukawa T, Fujisawa H, Taniguchi M, Yagi T, Kitamura H, et al:
Repulsive axon guidance molecule Sema3A inhibits branching
morphogenesis of fetal mouse lung. Mech Dev. 97:35–45. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Maione F, Capano S, Regano D, Zentilin L,
Giacca M, Casanovas O, Bussolino F, Serini G and Giraudo E:
Semaphorin 3a overcomes cancer hypoxia and metastatic dissemination
induced by antiangiogenic treatment in mice. J Clin Invest.
122:1832–1848. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nasarre P, Gemmill RM and Drabkin HA: The
emerging role of class-3 semaphorins and their neuropilin receptors
in oncology. Onco Targets Ther. 7:1663–1687. 2014.PubMed/NCBI
|
|
11
|
Bagci T, Wu JK, Pfannl R, Ilag LL and Jay
DG: Autocrine semaphorin 3a signaling promotes glioblastoma
dispersal. Oncogene. 28:3537–3550. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Müller MW, Giese NA, Swiercz JM, Ceyhan
GO, Esposito I, Hinz U, Büchler P, Giese T, Büchler MW, Offermanns
S, et al: Association of axon guidance factor semaphorin 3a with
poor outcome in pancreatic cancer. Int J Cancer. 121:2421–2433.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou H, Wu A, Fu W, Lv Z and Zhang Z:
Significance of semaphorin-3A and MMP-14 protein expression in
non-small cell lung cancer. Oncol Lett. 7:1395–1400.
2014.PubMed/NCBI
|
|
14
|
Blanc V, Nariculam J, Munson P, Freeman A,
Klocker H, Masters J and Williamson M: A role for class 3
semaphorins in prostate cancer. Prostate. 71:649–658. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Herman JG and Meadows GG: Increased class
3 semaphorin expression modulates the invasive and adhesive
properties of prostate cancer cells. Int J Oncol. 30:1231–1238.
2007.PubMed/NCBI
|
|
16
|
Rivera LB and Bergers G: Location,
location, location: Macrophage positioning within tumors determines
pro- or antitumor activity. Cancer Cell. 24:687–689. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li Y, Tian B, Yang J, Zhao L, Wu X, Ye SL,
Liu YK and Tang ZY: Stepwise metastatic human hepatocellular
carcinoma cell model system with multiple metastatic potentials
established through consecutive in vivo selection and studies on
metastatic characteristics. J Cancer Res Clin Oncol. 130:460–468.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ye QH, Qin LX, Forgues M, He P, Kim JW,
Peng AC, Simon R, Li Y, Robles AI, Chen Y, et al: Predicting
hepatitis B virus-positive metastatic hepatocellular carcinomas
using gene expression profiling and supervised machine learning.
Nat Med. 9:416–423. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun FX, Tang ZY, Lui KD, Ye SL, Xue Q, Gao
DM and Ma ZC: Establishment of a metastatic model of human
hepatocellular carcinoma in nude mice via orthotopic implantation
of histologically intact tissues. Int J Cancer. 66:239–243. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dai Z, Zhou SL, Zhou ZJ, Bai DS, Xu XY, Fu
XT, Chen Q, Zhao YM, Zhu K, Yu L, et al: Capn4 contributes to
tumour growth and metastasis of hepatocellular carcinoma by
activation of the FAK-Src signalling pathways. J Pathol.
234:316–328. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Oliveri RS, Wetterslev J and Gluud C:
Hepatocellular carcinoma. Lancet. 380:470–471. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Villanueva A, Newell P, Chiang DY,
Friedman SL and Llovet JM: Genomics and signaling pathways in
hepatocellular carcinoma. Semin Liver Dis. 27:55–76. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Suzuki K, Kumanogoh A and Kikutani H:
Semaphorins and their receptors in immune cell interactions. Nat
Immunol. 9:17–23. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang Y, He H, Srivastava N, Vikarunnessa
S, Chen YB, Jiang J, Cowan CW and Zhang X: Plexins are
GTPase-activating proteins for Rap and are activated by induced
dimerization. Sci Signal. 5:ra62012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lepelletier Y, Smaniotto S, Hadj-Slimane
R, Villa-Verde DM, Nogueira AC, Dardenne M, Hermine O and Savino W:
Control of human thymocyte migration by
Neuropilin-1/Semaphorin-3A-mediated interactions. Proc Natl Acad
Sci USA. 104:pp. 5545–5550. 2007; View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Garcia F, Lepelletier Y, Smaniotto S,
Hadj-Slimane R, Dardenne M, Hermine O and Savino W: Inhibitory
effect of semaphorin-3A, a known axon guidance molecule, in the
human thymocyte migration induced by CXCL12. J Leukoc Biol.
91:7–13. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Westbrook JA, Cairns DA, Peng J, Speirs V,
Hanby AM, Holen I, Wood SL, Ottewell PD, Marshall H, Banks RE, et
al: CAPG and GIPC1: Breast cancer biomarkers for bone metastasis
development and treatment. J Natl Cancer Inst. 108:pii: djv3602016.
View Article : Google Scholar
|
|
28
|
Wang L and Guo XL: Molecular regulation of
galectin-3 expression and therapeutic implication in cancer
progression. Biomed Pharmacother. 78:165–171. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tang D, Wang M, Sui A, Wang Y, Yang R,
Wang Z, Zhao Y, Jiao W and Shen Y: Prospective validation of
quantitative NSE mRNA in pleural fluid of non-small cell lung
cancer patients. Med Oncol. 30:6992013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gires O and Bauerle PA: EpCAM as a target
in cancer therapy. JJ Clin Oncol. 28:e239–e242. 2010. View Article : Google Scholar
|