Introduction
Prostate cancer is the second leading cause of
cancer mortality in men in the US. Androgen deprivation therapy
(ADT) which aims at decreasing androgen levels and/or blocking
androgen binding with an androgen receptor (AR) remains the
first-line treatment of advanced prostate cancer (1). Although, this approach works out
satisfactorily in the early use, prostate cancer may eventually
become uncontrollable, termed castration-resistant prostate cancer
(CRPC). Since 2004, docetaxel-based chemotherapy has been
established as the first-line therapy in patients with CRPC
(2). Unfortunately, nearly half of
the patients are initially docetaxel-insensitive, and the patients
that are initially sensitive to docetaxel may acquire resistance to
docetaxel within 1 year (3). In
recent years, a number of new drugs have obtained FDA approval and
have been used as the second-line therapy in CRPC, such as
cabazitaxel, enzalutamide and abiraterone. However, after
second-line treatment, resistance may still occur inevitably and
the overall survival is only prolonged for <5 months (4). Therefore, a deeper understanding of
the mechanisms of acquired docetaxel-resistance is extremely
valuable.
Docetaxel belongs to taxane chemotherapeutic drugs.
It can inhibit the mobility of microtubules via binding
microtubules and preventing their depolymerization, consequently
causing stagnation of mitosis and cell apoptosis (5). This mechanism was once thought to be
the main mechanism of taxanes treatment for prostate cancer, until
recently some research teams proved that taxanes also suppressed
the AR signaling pathways. Taxanes block AR nuclear translocation,
thus, inhibiting the expression of AR regulation genes (6). The discovery of this new mechanism
indicates that taxanes and ADT drugs may have a synergistic effect,
since both therapies inhibit the AR signaling pathway by targeting
different compositions of the AR axis, for example, while
abiraterone inhibits the generation of androgen, docetaxel can
block nuclear import of AR. However, a new clinical study reported
that the pharmacological activity of docetaxel was lower than
expected in the patients who were under abiraterone application
previously. Meanwhile, those who were under abiraterone application
and acquired resistance did not respond to docetaxel treatment
(7). This result suggests that the
two classes of drugs in addition to sharing a common
pharmacological mechanism (acting on the AR signaling pathway), may
also share common resistance mechanisms. Recently, various
laboratory studies and clinical studies have also suggested that
there is a cross-resistance phenomenon between docetaxel and
second-line ADT drugs (7–11).
Calpain is comprised of a family of relatively
conservative intracellular cysteine proteases. Currently, a total
of 14 calpain family members have been identified. Most of calpain
activity is due to µ-calpain and m-calpain, which are heterodimers
comprised of a large 80 kDa catalytic subunit (CAPN1 and CAPN2,
respectively) and a common 28 kDa regulatory subunit (CAPNS1)
(12). In cancer cells, the
expression of calpain is usually increased, since it can be used
for cleaving the cytoskeleton in order to promote migration and
invasion. In schwannoma and meningeoma, µ-calpain is overexpressed
(13). In clear cell renal cell
carcinoma, the expression of CAPN1 mRNA is increased (14). In colorectal cancer, increased
expression of m-calpain has also been observed (15). However, discordant results have been
presented for prostate cancer (16,17).
Recently, Liu et al indicated that long-term ADT induction
in hormone-naïve prostate cancer (HNPC) cell line LNCaP had the
ability to develop it into a more invasive CRPC cell subline, and
calpain in this CRPC cell subline was hyperactivated. In addition,
using calpeptin (a calpain inhibitor) suppressed the migration and
invasion of this subline (18),
reminding us that calpain plays an important role in the process of
HNPC developing into CRPC. In another study, Jorfi et al
demonstrated that when calpeptin was used in combination with
docetaxel in prostate cancer cells, calpeptin increased the
concentration of docetaxel by means of decreasing microvesicle
release (19). Since calpain
promotes ADT resistance and there is generally a cross-resistance
phenomenon between docetaxel and ADT drugs, we explored whether
calpain also plays a promoting role in acquired
docetaxel-resistance.
Androgen receptor splicing variants (AR-Vs) which
have been widely discussed lately, lack the ligand-binding domain
(LBD). Furthermore, they can perform constitutive transcriptional
activity in the absence of androgen and drive prostate cancer
development in the direction of castration-resistance. In addition,
these AR-Vs are generally overexpressed in CRPC compared with HNPC
(20,21). AR-V7 (also known as AR3) is one of
the major AR-Vs. CRPC patients with a high expression of AR-V7 have
a relatively shorter overall survival (22). Other recent studies strongly
confirmed that AR-V7 played an important role in the resistance of
abiraterone and enzalutamide (23,24).
Although, the promoting role of AR-V7 concerning ADT-treatment
resistance has been widely recognized, whether AR-V7 also plays a
crucial role in the resistance of docetaxel remains unclear.
After 2 in vitro docetaxel-resistant prostate
cancer cell sublines were successfully established, we focused on
the resistance mechanisms of docetaxel in respect to calpain and
AR-V7. Subsequently we studied the biological functions by
targeting calpain and the influence on the transcriptional activity
of the AR signaling pathway by targeting importin-β, in order to
find new biomarkers for CRPC treatment from a new perspective as
well as appropriate methods to overcome docetaxel-resistance in
CRPC.
Materials and methods
Cell lines and chemicals
Human CRPC cell lines, PC3 and Rv1, human HNPC cell
line, LNCaP, human benign prostatic hyperplasia cell line, BPH-1,
were obtained from the American Type Culture Collection (ATCC;
Manassas, VA, USA) and maintained in RPMI-1640 medium (Gibco,
Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS)
and 1% penicillin/streptomycin. Docetaxel-resistant clones, PC3-DR
and RV1-DR, were selected by culturing cells with docetaxel in a
dose-escalation manner at a 48-h exposure. The initial culture was
at a concentration of 2.5 nM docetaxel for PC3 and 1 nM for Rv1.
After sensitive clones were no longer present, surviving PC3 and
Rv1 cells repopulated the flask and the concentration of docetaxel
was increased to 5, 10 and 20 nM for PC3, and 2.5, 5 and 10 nM for
Rv1. The process of acquired drug resistance took 4 months, and the
docetaxel-resistant cell sublines were named PC3-DR and Rv1-DR. In
parallel, parental PC3 and Rv1 cells were exposed to dimethyl
sulfoxide (DMSO) (vehicle solution) in the same dose-escalation
manner. For incubation experiments, docetaxel and calpeptin were
purchased from Selleckchem (Houston, TX, USA) and importazole was
purchased from Merck Millipore (Billerica, MA, USA).
Western blot analysis
Western blot analysis was carried out as previously
described (25,26). Briefly, cell proteins were isolated
using RIPA buffer (Beyotime Institute of Biotechnology, Jiangsu,
China) supplemented with a protease inhibitor mixture stock
solution (Roche Molecular Biochemicals, Mannheim, Germany) and
phenylmethanesulfonyl fluoride (PMSF). Equivalent amounts of
protein were resolved by 10% SDS polyacrylamide gels and
transferred to polyvinylidene fluoride (PVDF) membranes, which
then, were blocked with 5% (w/v) nonfat dried-milk and incubated
with the indicated primary antibody in Tris-buffered saline
overnight at 4°C. Primary antibodies were FAK antibody (Epitomics,
Burlingame, CA, USA), CAPNS1 antibody (ABclonal, Cambridge, MA,
USA), AR-V7 monoclonal antibody (A&G Pharmaceutical, Columbia,
MD, USA) (AR-V7 monoclonal antibody recognizes AR-V7, but does not
cross-react with intact AR or other AR-Vs), AR antibody (N-20)
(Santa Cruz Biotechnology, Santa Cruz, CA, USA) [AR antibody (N-20)
recognizes AR-FL, AR-V7 and other non-specific AR-Vs], PSA antibody
(Ruiying Biology, Jiangsu, China), lamin B1 antibody, α-tubulin
antibody (both from Abcam, Cambridge, UK) and β-actin (Santa Cruz
Biotechnology). The blots then were stained with the appropriate
horseradish peroxidase-conjugated secondary antibodies, goat
anti-rabbit (ProSci, Inc., Poway, CA, USA) and goat anti-mouse
(Santa Cruz Biotechnology). Bands were visualized with enhanced
chemiluminescence (Thermo Fisher Scientific Inc., Waltham, MA, USA)
using the ChemiDoc XRS+ system (Bio-Rad, Hercules, CA, USA).
Cell viability and proliferation
assay
To verify the effects on cell viability, we used the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay (27). Cells were seeded at a
density of 1×104 in 96-well culture dishes and 24 h
later the medium was removed and replaced with new medium alone
(control) or with medium containing the drug. After 72 h, the cells
were incubated with 0.1 µg/µl of aqueous MTT solution for 4 h at
37°C. Afterwards, formazan complexes were solubilized by the
addition of 150 µl of DMSO, and the color absorbance was assessed
on a microplate spectrophotometer (Molecular Dynamics) at 450 nm
(test wavelength) and 620 nm (reference wavelength). The percentage
of surviving cells was estimated by dividing the A450-A620 nm of
the treated cells by the A450-A620 nm of the control cells.
Similarly, cell proliferation was assessed at 24, 48, 72 and 96 h
under different treatments at a density of 3×103
cells/well by MTT method as previously described.
Migration assay
The migratory ability of the PC3-DR and Rv1-DR cells
under docetaxel, calpeptin, docetaxel combined with calpeptin and
DMSO treatment was assessed using a Corning Transwell assay
according to the manufacturer's protocol. A total of
1×105 cells in 200 µl of serum-free medium containing
drugs were seeded into the upper chamber of the system. The lower
chambers were filled with 0.75 ml of 20% FBS complete medium. After
24 h of incubation, the cells in the upper chamber were removed
with a cotton swab, and the transmigrated cells were fixed in
methanol and stained with crystal violet. The stained cells were
counted by photographing 5 fields/membrane.
Cell cycle assay
After 24 h of starvation and synchronization in
serum-free medium, the cells were cultured in complete media
containing different drugs for 2 days (3×105
cells/well), and then harvested and washed twice with cold PBS.
Subsequently, the cells were fixed in ice-cold 75% ethanol at 4°C
overnight and incubated with 50 µg/ml of propidium iodide and 50
µg/ml of RNase A at 37°C for 30 min in the dark. The percentage of
cells in the G0/G1, S and G2/M phases was assessed by FACS (BD
Biosciences, Franklin Lakes, NJ, USA). Data were analyzed with the
ModFit LT software.
Cell apoptosis assay
Cells were cultured in complete media containing
different drugs for 2 or 3 days (3×105 cells/well).
Adherent and detached cells were pooled, washed and labeled with
Annexin V-FITC and propidium iodide (PI) using the Annexin
V-FITC/PI apoptosis detection kit (KeyGen Biotech, Jiangsu, China)
according to the manufacturer's instructions. The percentage of
cells displaying Annexin V-FITC/PI staining were measured by FACS
(BD Biosciences). Data were analyzed using the ModFit LT
software.
Statistical analysis
Statistical comparisons of at least 3 independent
measurements were performed using the unpaired Student's t-test
with a 95% confidence interval. For all statistical analyses,
results of p≤0.05 were considered statistically significant. Data
are provided as the mean ± SD.
Results
Establishment and identification of 2
in vitro docetaxel-resistant prostate cancer cell sublines
We used CRPC cell lines Rv1 and PC3 as the parental
cell lines. The docetaxel-resistant CRPC cell sublines were
established by continuous exposure to gradually increasing
concentrations of docetaxel. The PC3 and Rv1 docetaxel-resistant
cell sublines (named PC3-DR and Rv1-DR) were established with
abilities of stable growth and proliferation at a final
concentration of 20 and 10 nM docetaxel, respectively. The
identification was carried out using MTT assay and a FACS cell
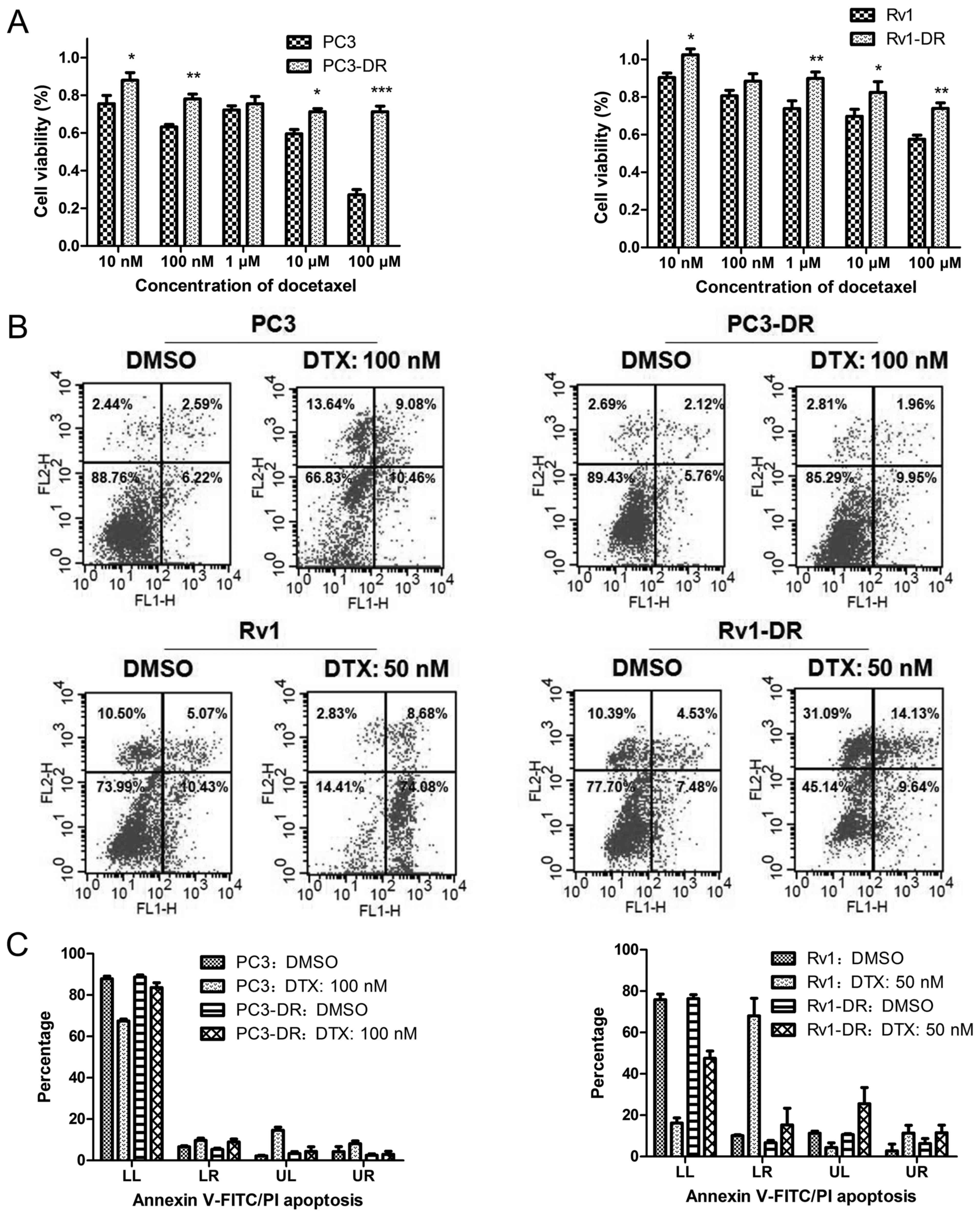
apoptosis assay. Compared with their parental cells both
docetaxel-resistant cell sublines had a higher survival rate
(Fig. 1A), and a lower apoptosis
rate (Fig. 1B and C) when exposed
to docetaxel.
Activation and overexpression of
calpain is induced by docetaxel treatment
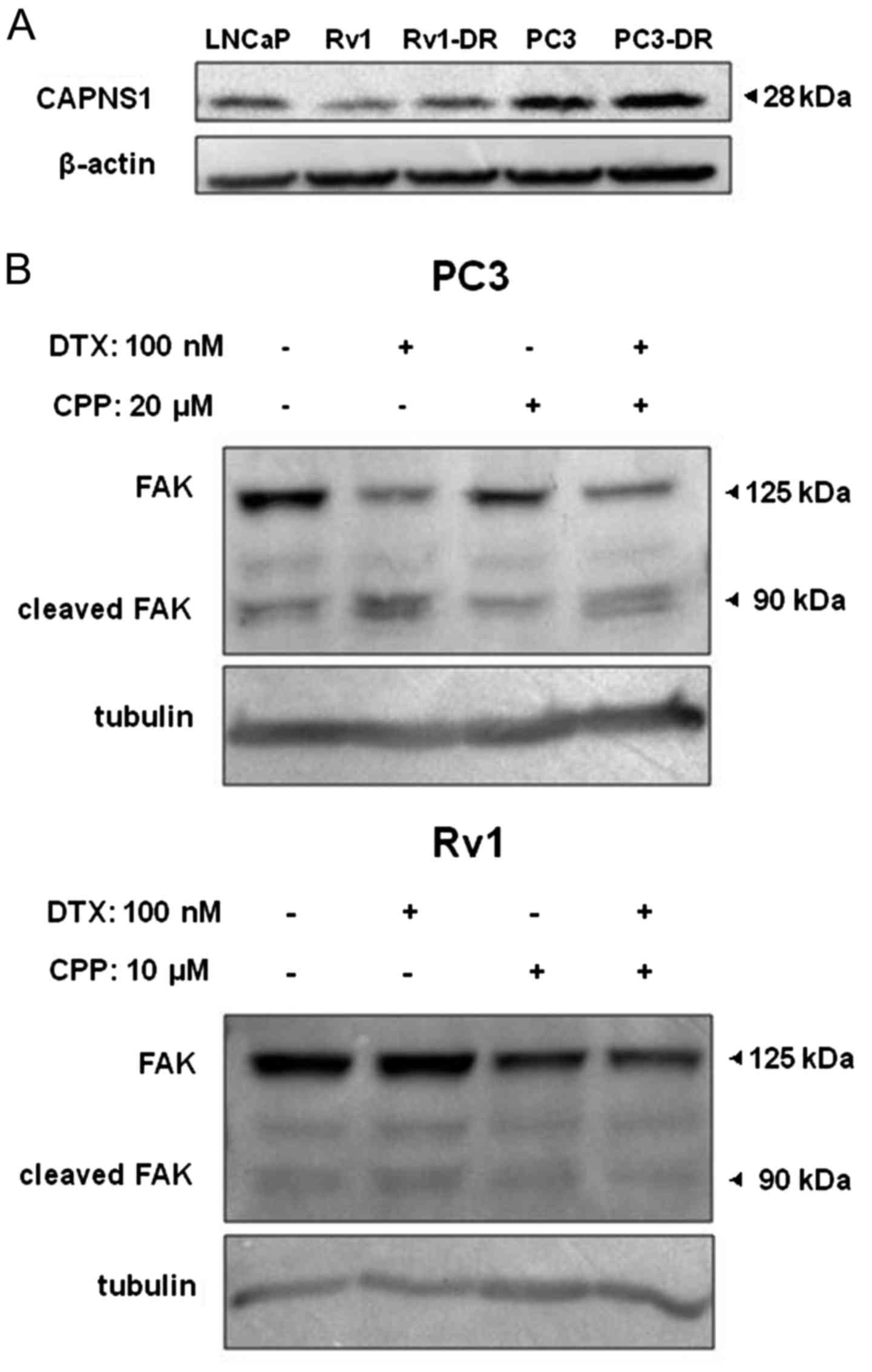
To assess the expression of calpain in
docetaxel-resistant CRPC cell sublines and parental cell lines, we
performed western blot analysis. As shown in Fig. 2A, high expression of calpain S1 was
clearly observed in Rv1-DR cells compared with Rv1 cells, but no
specific difference in the expression levels of calpain was
observed between the PC3 and PC3-DR cells. Furthermore, to assess
the activity of calpain, we chose focal adhesion kinase (FAK) as
the marker. FAK (125 kDa) can be specifically cleaved into 90 kDa
fragments by calpain (28). As
shown in Fig. 2B, the bands at 90
kDa were upregulated under docetaxel treatment (100 nM) in both Rv1
and PC3 cells; while with calpeptin (a calpain inhibitor) treatment
(10 µM for Rv1 and 20 µM for PC3 cells), the 90 kDa bands remained
steady, and with calpeptin + docetaxel treatment, the 90 kDa bands
were ‘rescued’ from upregulation.
Effects of calpain inhibitor combined
with docetaxel on biological functions in PC3-DR and Rv1-DR
cells
To assess docetaxel susceptibility in
docetaxel-resistant cell sublines we used an MTT assay and found
that docetaxel susceptibility was significantly increased with
exposure to calpeptin (20 µM for PC3-DR and 10 µM for Rv1-DR) in
both PC3-DR (Fig. 3A) and Rv1-DR
(data not shown) cells. To compare the efficiency between calpeptin
combined with docetaxel and docetaxel alone in terms of cell
proliferation, migration, cell cycle and cell apoptosis, we used
MTT and Transwell migration assays, and FACS cell cycle and
apoptosis assays, respectively. The results revealed that calpeptin
combined with docetaxel had a distinct advantage over docetaxel
alone in terms of inhibition of cell proliferation and cell
migration in both PC3-DR (Fig. 3B and
C) and Rv1-DR (data not shown) cells. G2/M phase arrest was
more clearly observed after calpeptin combined with docetaxel
treatment for 2 days than docetaxel alone in PC3-DR cells (Fig. 3D), but not in Rv1-DR cells (Fig. 3E). Early stage cell apoptosis was
more obvious after calpeptin combined with docetaxel treatment for
2 days than docetaxel alone in Rv1-DR cells (Fig. 3G), but not in PC3-DR cells (Fig. 3F). However, when we prolonged the
exposure time to 3 days, early- and late-stage cell apoptosis were
both more obvious under calpeptin combined with docetaxel treatment
than docetaxel alone in PC3-DR cells (Fig. 3H).
Changes in the expression of AR during
docetaxel resistance
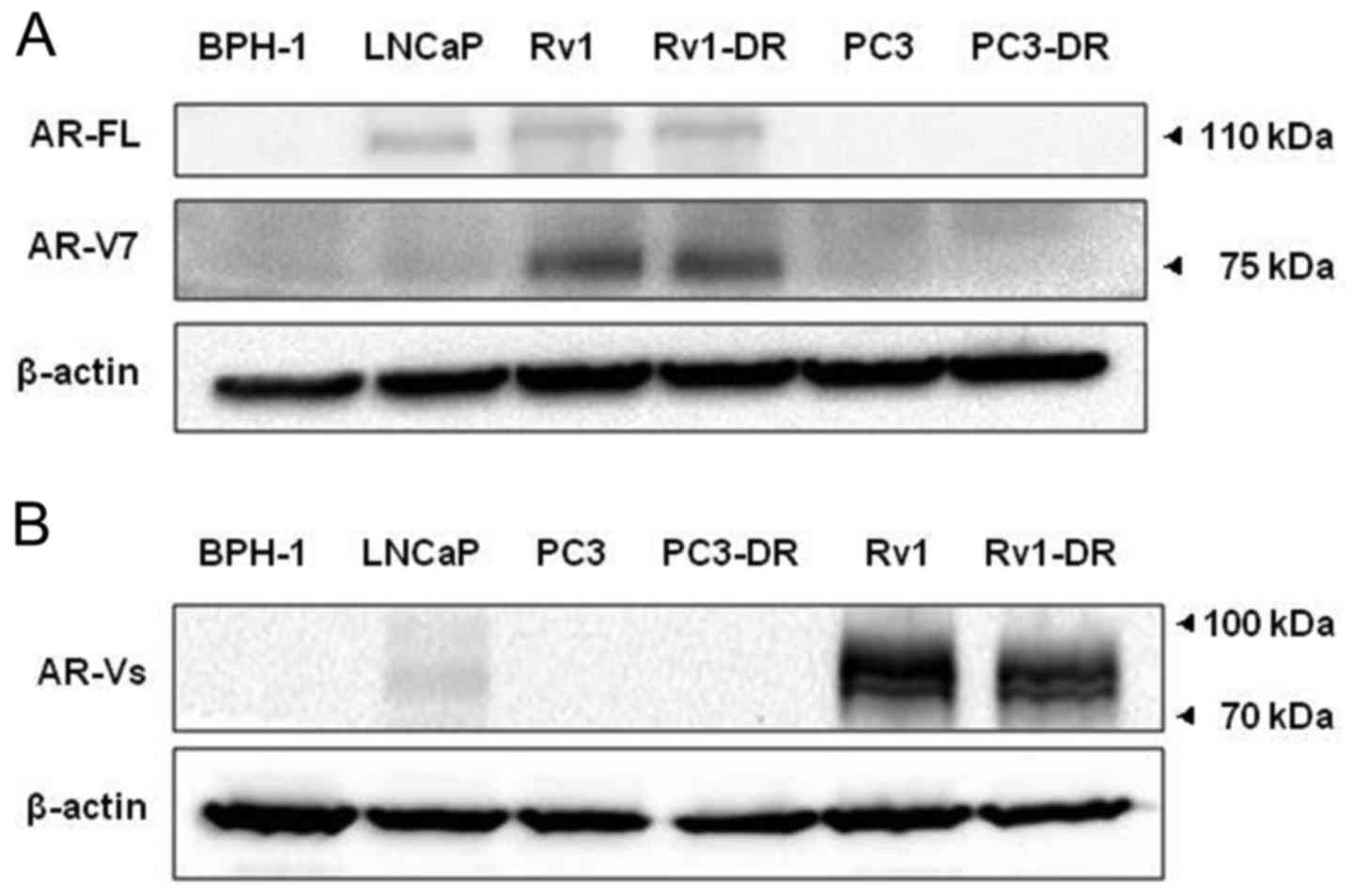
To assess the changes in the expression of AR-FL,
AR-Vs and AR-V7 in Rv1-DR and its parental cell line Rv1, we used
prostate cell line BPH-1, LNCaP, PC3 and PC3-DR as controls.
Compared with LNCaP and Rv1, we found that the expression of AR-V7
remained at a high level in the Rv1-DR cells (Fig. 4A), while the expression of AR-FL
remained very low expression (Fig.
4A), and other non-specific AR-Vs were significantly
downregulated (Fig. 4B).
Effects of importin-β inhibitor on AR
nuclear import and transcriptional activity
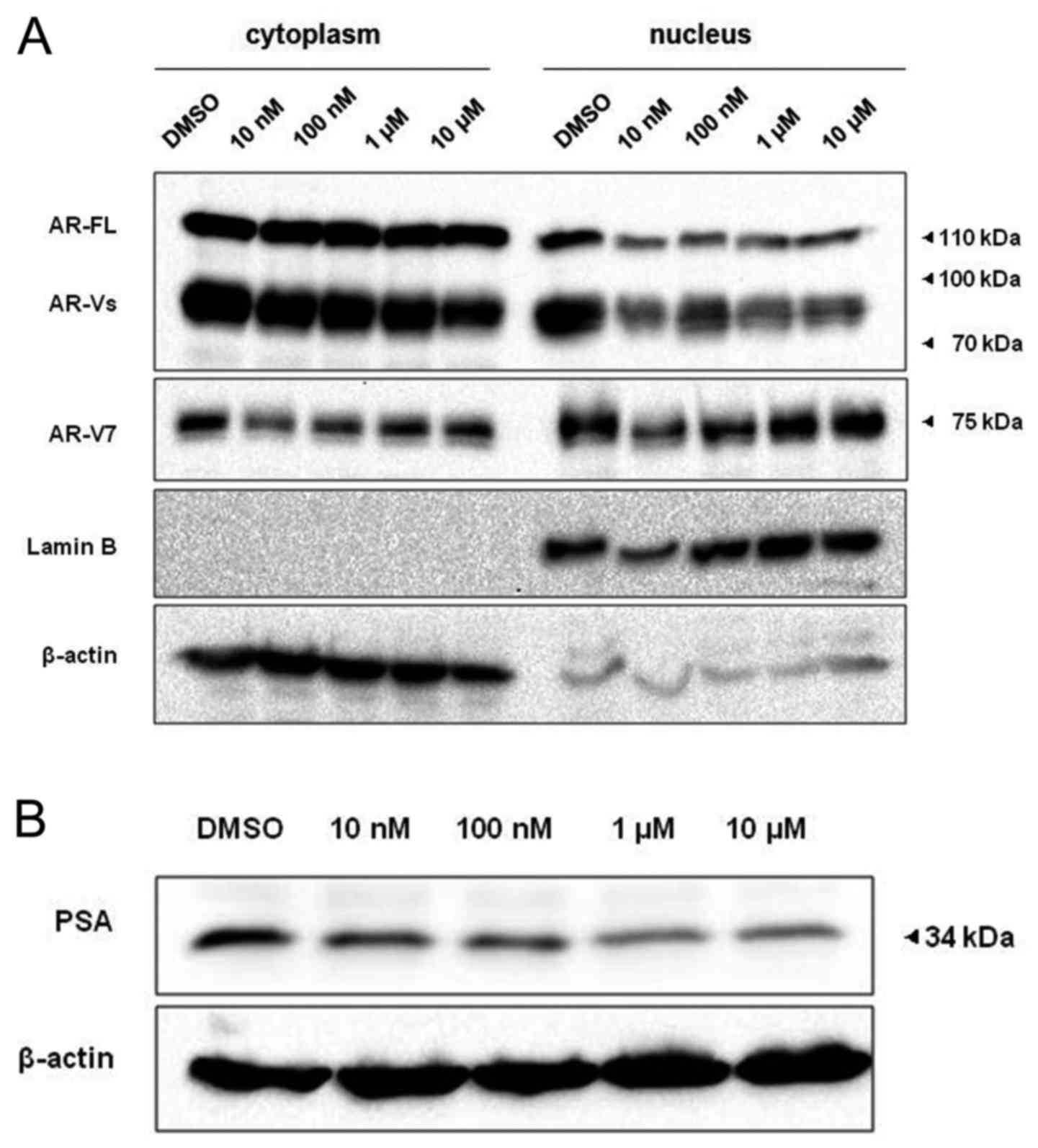
To explore the effect of importin-β inhibitor on AR
nuclear import and transcriptional activity, we used a specific
importin-β inhibitor (importazole). After a 48-h treatment with
gradually increasing concentrations of importazole, nuclear and
cytoplasmic protein were isolated using a Nuclear and Cytoplasmic
Protein Extraction kit (P0027; Beyotime Institute of Biotechnology)
for assessing AR and whole cell protein was isolated for assessing
the transcriptional activity (measured with PSA). The results
revealed that after using importazole AR-FL and AR-Vs in the
nucleus were significantly downregulated at a concentration as low
as 10 nM (Fig. 5A) and the
expression of PSA decreased slightly with the increase of drug
concentration (Fig. 5B); while
AR-V7 in the nucleus was not significantly altered. (Fig. 5A).
Discussion
As docetaxel chemotherapy remains a mainstay in the
treatment of CRPC, it is imperative to derive a better
understanding of the mechanisms underlying the inherent and
acquired docetaxel-resistance, both of which are commonly observed
clinically. In the present study we established 2 in vitro
docetaxel-resistant prostate cancer cell sublines (PC3-DR and
Rv1-DR) by continuously exposing these sublines to gradually
increasing concentrations of docetaxel as described in previous
studies (29), and identified a
docetaxel-resistant character using MTT and FACS cell apoptosis
assays.
Previous studies showed that calpain activity played
an important role in tumor biology functions. Calpain can promote
cell invasion and migration by cleaving the cytoskeleton-related
protein FAK (30). However, the
role calpain plays in cell survival and apoptosis is still unclear.
Calpain can cleave wild-type p53 and induce suppression of
p53-dependent apoptosis restrained (31). Calpain is also capable of cleaving
anti-apoptotic molecules of the Bcl-2 family leading to the
promotion of apoptosis (32,33).
This uncertainty may be due to the cellular localization of calpain
when it is activated. Recently, it was reported that
calpain-dependent proteolysis of the AR into AR-Vs may be
androgen-independent mechanism (34–36).
Furthermore, calpain can cleave FHL2 and remove the AR and AR-Vs
inhibited effect of FHL2, suggesting that AR and AR-Vs are easily
imported to the nucleus as transcription factors which drive CRPC
progression (37). The promoting
role calpain plays in HNPC developing into CRPC has been described,
however, the impact of calpain in the process of CRPC developing
into docetaxel-resistant CRPC has not been reported. In the present
study we assessed the common regulatory subunit CAPNS1 of µ-calpain
and m-calpain in 2 docetaxel-resistant CRPC cell sublines and their
parental cell lines, and found that CAPNS1 was upregulated in
Rv1-DR in comparison to Rv1. In the detection of calpain activity,
we chose FAK as the marker, and found that calpain was activated
with docetaxel treatment in PC3 and Rv1 cells. Since short-term
docetaxel treatment can activate calpain, and long-term docetaxel
induction can upregulate calpain expression, can we assume that
calpain is a key factor that promotes CRPC resistance to docetaxel?
Or is this just an accompanying phenomenon along with other cell
physiological functions due to environmental changes? In order to
identify this hypothesis, we studied the effects of calpain
inhibitor combined with docetaxel on biological functions in PC3-DR
and Rv1-DR cells subsequently. We found that calpain inhibition
sensitized PC3-DR and Rv1-DR cells to docetaxel, and calpeptin
combined with docetaxel exhibited a distinct advantage over
docetaxel alone in terms of inhibition of cell proliferation,
inhibition of cell migration, G2/M phase arrest and induction of
cell apoptosis. These results indicate that calpain plays a
promoting role in acquired docetaxel-resistance of PC3 and Rv1.
Notably, Rv1-DR cells did not exhibit obvious G2/M
arrest in calpeptin combined with docetaxel therapy, but exhibited
distinct cell apoptosis in combination therapy. These results
indicated that Rv1-DR may possess a different mechanism prior to
G2/M arrest inducing cell apoptosis. This mechanism may be the
inhibition of the AR-signaling pathway since Rv1-DR cells are very
rich in AR (6,38). In addition, we found that PC3-DR
cells exhibited an obvious G2/M arrest at a lower concentration of
40 nM of docetaxel and 20 µM of calpeptin treatment, but when we
increased the concentration of docetaxel to 100 nM, we still did
not observe distinct cell apoptosis. Subsequently, we prolonged the
exposure time from 2 to 3 days, and observed both early- and
late-stage cell apoptosis after calpeptin was combined with
docetaxel treatment, suggesting that the cell apoptosis induced by
the combintion of calpeptin with docetaxel is more time-dependent
than concentration-dependent in PC3-DR.
However, it should be noted that, the present study
only examined the common regulatory subunit CAPNS1 expression of
the heterodimers µ-calpain and m-calpain and further exploration is
warranted, whether to expand the molecular mechanism more
accurately or to develop highly selective targeting drugs. We used
the degradation of only one substrate to determine calpain
activity, however further validation of calpain activity with other
substrates may be more persuasive. A deeper elucidation of the
underlying mechanism in the combination of calpeptin with docetaxel
therapy and a in vivo animal experiment evaluating the
efficacy and safety of the therapy are both important topics for
further study.
To date, a variety of mechanisms involved in
docetaxel-resistance have been described, but the mechanisms
described in literature are mostly common mechanisms which are
found in many tumor cells, such as structural and functional
changes of microtubules, increasing drug efflux and alterations in
apoptosis-related proteins (39),
while mechanisms specific to prostate cancer are poorly understood.
AR-V7 exists in benign and malignant prostatic tissue, but is
mostly enriched in metastatic disease (21,22).
The recurrence rate of HNPC after radical surgery has a close and
positive correlation to the expression level of AR-V7 in prostatic
tissue (21). Moreover, in CRPC
bone metastases, overall survival exhibits a strong negative
correlation to the expression level of AR-V7 (22). These results indicate that AR-V7
often marks a poor prognosis of malignant prostate cancer.
Additionally, previous studies have shown that AR-V7 plays an
important role in ADT resistance (23,40),
particularly Antonarakis et al demonstrated with strong
evidence that the expression of AR-V7 in circulating tumor cells
had a negative correlation to abiraterone and enzalutamide
treatment outcomes (24). AR-V7 has
been widely accepted to be involved in the resistance of ADT,
meanwhile, various laboratory studies and clinical studies suggest
that there is a cross-resistance phenomenon between docetaxel and
second-line ADT drugs (7,9–11),
however it is poorly understood whether AR-V7 also confers
docetaxel-resistance to prostate cancer cells. Recently, Martin
et al observed that a small molecule inhibitor targeting the
N-terminal of AR-Vs sensitized prostate cancer cells to docetaxel
(41). In addition, Steinestel
et al discovered that docetaxel-treated patients had a
higher expression of AR-V7 in circulating tumor cells (42). Taking into the consideration the
aforementioned facts, we assessed the changes in the expression of
AR-FL, AR-Vs and AR-V7 in Rv1-DR and its parental cell line Rv1,
and found that the expression of AR-V7 remained at a high level in
the docetaxel-resistant cell subline, while full-length the
expression of AR remained very low and other non-specific AR-Vs
were significantly downregulated. This is a possible consequence of
the participation of the constitutively and transcriptionally
active AR-V7 in the acquired resistance to docetaxel and overcoming
drug toxicity in Rv1 cells.
The human AR is an 110 kDa, 919 amino acid protein
composed of 4 domains: i) the amino terminal activation domain
(NTD); ii) the DNA-binding domain (DBD); iii) the hinge region; and
iv) the carboxyl ligand-binding domain (LBD) (43). The NTD includes the majority of the
AR and determines androgen specific transcriptional activity
(44), while the nucleus
localization is a crucial step in the process of the AR signaling
pathway (45). AR-V7 lacks the
hinge region and LBD, thus making AR-V7 very different from AR-FL
(21). Canonical AR nuclear
localization signal (NLS) is located in the hinge region that AR-V7
lacks and Chan et al demonstrated that mysterious exon
3-encoded DBD of AR-V7 joined with NLS to reconstitute a bipartite
AR NLS which enhanced AR-V7 nuclear localization (46). In prostate cancer cells, AR-FL is
transported into the nucleus mediated by microtubules and actin.
However, docetaxel which is a microtubule stabilizing agent, can
block AR-FL and inhibit the AR signaling pathway by microtubule
stabilization. Nevertheless, due to the lack of a hinge structure
which binds to microtubules, AR-V7 may have a
microtubule-independent way of translocating to the nucleus, hence
AR-V7 nuclear translocation is not regulated by docetaxel (6,38).
Kaku et al assessed the nuclear import
abilities of each domain of AR and their mechanisms were related to
Ran and importin-α/β. They discovered that NTD and DBD showed
ligand-independent nuclear import ability, and importin-β knockdown
strongly blocked DBD import (47).
Importazole is a small molecule inhibitor of the transport receptor
importin-β (48). A former study
suggested that importazole blocked the nuclear import of AR-V7 as
was determined by FRAP assay, however, docetaxel was only capable
of blocking nuclear import of AR-FL, but not AR-V7 (49). Moreover, the concentration they used
to treat the cells was as much as 50 µM and under such a high
concentration it is hard to guarantee that the results of the
experiment are still caused by the normal pharmacological effects
of importazole. Furthermore it is hard to achieve such a
concentration in human prostate cells. In the present study we used
gradient increasing doses of importazole to treat Rv1-DR cells for
48 h. The nucleus and cytoplasm proteins were isolated to assess
the distribution of AR-FL, AR-Vs and AR-V7, in order to study the
effect of importazole on their nuclear import. To illustrate that
the isolation of nuclear and cytoplasmic protein was successful, we
chose β-actin as a cytoplasmic marker and lamin B as a nuclear
marker. We found that gradient increasing doses of importazole
could not block the nuclear import of AR-V7. Furthermore, we
studied the effect of importazole on AR transcriptional activity,
and observed that PSA was only slightly downregulated with the
increase of drug concentration. Although, importazole could block
the nuclear import of AR-FL and non-specific AR-Vs significantly at
a concentration as low as 10 nM, the AR transcriptional activity
was slightly inhibited. These results indicated that the expression
of AR-FL which remains at a low level and non-specific AR-Vs which
are significantly downregulated are not the key factors of the AR
signaling pathway in Rv1-DR. Moreover, AR-V7 which remains highly
expressed in Rv1-DR and is not regulated by importazole may be the
key factor of the AR signaling pathway and confer
docetaxel-resistance to Rv1-DR cells.
We have to point out that only using PSA expression
to illustrate the AR transcriptional activity is not convincing
enough, we could also use other AR target gene products or
luciferase assay to assess AR transcriptional activity. We
suggested that importazole can block the nuclear import of AR-FL
and AR-Vs. A Co-IP assay study on the interaction between
importin-β and AR-FL as well as AR-Vs may provide us with more
powerful evidence. Additionally, the effect of importazole on the
biological functions of prostate cancer has not been reported yet.
These issues are all urgent and warrant elucidation in the
future.
In the present study we demonstrated that
overexpression and activation of calpain confers chemoresistance to
CRPC cells. Targeting calpain sensitized PC3-DR and Rv1-DR cells to
docetaxel. The calpain-induced insensitivity to docetaxel was
partially reversed by calpeptin (a calpain inhibitor). These
results concerning the biological functions collectively highlight
an important role for calpain in the modulation of
docetaxel-resistance of prostate cancer cells. Moreover, we
suggested that AR-V7 may play a promoting role in CRPC-acquired
docetaxel-resistance. An importin-β inhibitor was only capable of
slightly decreasing the transcriptional activity of the AR
signaling pathway via blocking nuclear import of AR-FL and some
non-specific AR-Vs, instead of AR-V7.
In conclusion, we provided evidence that calpain and
AR-V7 play crucial roles in mediating the chemoresistance of CRPC
cells to docetaxel. In addition, the expression status of calpain
and AR-V7 could potentially be used as biomarkers to aid treatment
selection and sequencing in prostate cancer. Furthermore, targeting
calpain and AR-V7 may thus have potential benefits in improving the
effectiveness of chemotherapy and overcoming docetaxel-resistance
in prostate cancer.
Acknowledgements
This research was supported by the National Natural
Science Foundation of China (Grant no. 30872924, 81072095 and
81372760), Hubei Province Scientific and Technical International
Joint Project (Grant no. 2015BHE007) to Professor Xiaoping Zhang.
The authors also thank Professor Shaoyong Chen (Beth Israel
Deaconess Medical Center and Harvard Medical School, Boston, USA)
for providing the Rv1 cell line.
References
|
1
|
Harris WP, Mostaghel EA, Nelson PS and
Montgomery B: Androgen deprivation therapy: Progress in
understanding mechanisms of resistance and optimizing androgen
depletion. Nat Clin Pract Urol. 6:76–85. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tannock IF, de Wit R, Berry WR, Horti J,
Pluzanska A, Chi KN, Oudard S, Théodore C, James ND, Turesson I, et
al: TAX 327 Investigators: Docetaxel plus prednisone or
mitoxantrone plus prednisone for advanced prostate cancer. N Engl J
Med. 351:1502–1512. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Loriot Y and Fizazi K: Taxanes: Still a
major weapon in the armamentarium against prostate cancer. Eur
Urol. 63:983–985. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Heidenreich A, Bastian PJ, Bellmunt J,
Bolla M, Joniau S, van der Kwast T, Mason M, Matveev V, Wiegel T,
Zattoni F, et al: European Association of Urology: EAU guidelines
on prostate cancer. Part II: Treatment of advanced, relapsing, and
castration-resistant prostate cancer. Eur Urol. 65:467–479. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jordan MA and Wilson L: Microtubules as a
target for anticancer drugs. Nat Rev Cancer. 4:253–265. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhu ML, Horbinski CM, Garzotto M, Qian DZ,
Beer TM and Kyprianou N: Tubulin-targeting chemotherapy impairs
androgen receptor activity in prostate cancer. Cancer Res.
70:7992–8002. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mezynski J, Pezaro C, Bianchini D, Zivi A,
Sandhu S, Thompson E, Hunt J, Sheridan E, Baikady B, Sarvadikar A,
et al: Antitumour activity of docetaxel following treatment with
the CYP17A1 inhibitor abiraterone: Clinical evidence for
cross-resistance? Ann Oncol. 23:2943–2947. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schweizer MT, Zhou XC, Wang H, Bassi S,
Carducci MA, Eisenberger MA and Antonarakis ES: The influence of
prior abiraterone treatment on the clinical activity of docetaxel
in men with metastatic castration-resistant prostate cancer. Eur
Urol. 66:646–652. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
van Soest RJ, van Royen ME, de Morrée ES,
Moll JM, Teubel W, Wiemer EA, Mathijssen RH, de Wit R and van
Weerden WM: Cross-resistance between taxanes and new hormonal
agents abiraterone and enzalutamide may affect drug sequence
choices in metastatic castration-resistant prostate cancer. Eur J
Cancer. 49:3821–3830. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nadal R, Zhang Z, Rahman H, Schweizer MT,
Denmeade SR, Paller CJ, Carducci MA, Eisenberger MA and Antonarakis
ES: Clinical activity of enzalutamide in docetaxel-naïve and
docetaxel-pretreated patients with metastatic castration-resistant
prostate cancer. Prostate. 74:1560–1568. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cheng HH, Gulati R, Azad A, Nadal R,
Twardowski P, Vaishampayan UN, Agarwal N, Heath EI, Pal SK, Rehman
HT, et al: Activity of enzalutamide in men with metastatic
castration-resistant prostate cancer is affected by prior treatment
with abiraterone and/or docetaxel. Prostate Cancer Prostatic Dis.
18:122–127. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Goll DE, Thompson VF, Li H, Wei W and Cong
J: The calpain system. Physiol Rev. 83:731–801. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kimura Y, Koga H, Araki N, Mugita N,
Fujita N, Takeshima H, Nishi T, Yamashima T, Saido TC, Yamasaki T,
et al: The involvement of calpain-dependent proteolysis of the
tumor suppressor NF2 (merlin) in schwannomas and meningiomas. Nat
Med. 4:915–922. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Braun C, Engel M, Seifert M, Theisinger B,
Seitz G, Zang KD and Welter C: Expression of calpain I messenger
RNA in human renal cell carcinoma: Correlation with lymph node
metastasis and histological type. Int J Cancer. 84:6–9. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lakshmikuttyamma A, Selvakumar P, Kanthan
R, Kanthan SC and Sharma RK: Overexpression of m-calpain in human
colorectal adenocarcinomas. Cancer Epidemiol Biomarkers Prev.
13:1604–1609. 2004.PubMed/NCBI
|
|
16
|
Mamoune A, Luo JH, Lauffenburger DA and
Wells A: Calpain-2 as a target for limiting prostate cancer
invasion. Cancer Res. 63:4632–4640. 2003.PubMed/NCBI
|
|
17
|
Rios-Doria J, Day KC, Kuefer R, Rashid MG,
Chinnaiyan AM, Rubin MA and Day ML: The role of calpain in the
proteolytic cleavage of E-cadherin in prostate and mammary
epithelial cells. J Biol Chem. 278:1372–1379. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu T, Mendes DE and Berkman CE: Prolonged
androgen deprivation leads to overexpression of calpain 2:
Implications for prostate cancer progression. Int J Oncol.
44:467–472. 2014.PubMed/NCBI
|
|
19
|
Jorfi S, Ansa-Addo EA, Kholia S, Stratton
D, Valley S, Lange S and Inal J: Sci Rep. 5:130062015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dehm SM, Schmidt LJ, Heemers HV, Vessella
RL and Tindall DJ: Splicing of a novel androgen receptor exon
generates a constitutively active androgen receptor that mediates
prostate cancer therapy resistance. Cancer Res. 68:5469–5477. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Guo Z, Yang X, Sun F, Jiang R, Linn DE,
Chen H, Chen H, Kong X, Melamed J, Tepper CG, et al: A novel
androgen receptor splice variant is up-regulated during prostate
cancer progression and promotes androgen depletion-resistant
growth. Cancer Res. 69:2305–2313. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hörnberg E, Ylitalo EB, Crnalic S, Antti
H, Stattin P, Widmark A, Bergh A and Wikström P: Expression of
androgen receptor splice variants in prostate cancer bone
metastases is associated with castration-resistance and short
survival. PLoS One. 6:e190592011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li Y, Chan SC, Brand LJ, Hwang TH,
Silverstein KA and Dehm SM: Androgen receptor splice variants
mediate enzalutamide resistance in castration-resistant prostate
cancer cell lines. Cancer Res. 73:483–489. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Antonarakis ES, Lu C, Wang H, Luber B,
Nakazawa M, Roeser JC, Chen Y, Mohammad TA, Chen Y, Fedor HL, et
al: AR-V7 and resistance to enzalutamide and abiraterone in
prostate cancer. N Engl J Med. 371:1028–1038. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang X, Jin TG, Yang H, DeWolf WC,
Khosravi-Far R and Olumi AF: Persistent c-FLIP(L) expression is
necessary and sufficient to maintain resistance to tumor necrosis
factor-related apoptosis-inducing ligand-mediated apoptosis in
prostate cancer. Cancer Res. 64:7086–7091. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu S, Tao Z, Hai B, Liang H, Shi Y, Wang
T, Song W, Chen Y, OuYang J, Chen J, et al: miR-424(322) reverses
chemoresistance via T-cell immune response activation by blocking
the PD-L1 immune checkpoint. Nat Commun. 7:114062016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu S, Wang T, Song W, Jiang T, Zhang F,
Yin Y, Jiang SW, Wu K, Yu Z, Wang C, et al: The inhibitory effects
of AR/miR-190a/YB-1 negative feedback loop on prostate cancer and
underlying mechanism. Sci Rep. 5:135282015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cooray P, Yuan Y, Schoenwaelder SM,
Mitchell CA, Salem HH and Jackson SP: Focal adhesion kinase
(pp125FAK) cleavage and regulation by calpain. Biochem
J. 318:41–47. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Domingo-Domenech J, Vidal SJ,
Rodriguez-Bravo V, Castillo-Martin M, Quinn SA, Rodriguez-Barrueco
R, Bonal DM, Charytonowicz E, Gladoun N, de la Iglesia-Vicente J,
et al: Suppression of acquired docetaxel resistance in prostate
cancer through depletion of Notch- and Hedgehog-dependent
tumor-initiating cells. Cancer Cell. 22:373–388. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chan KT, Bennin DA and Huttenlocher A:
Regulation of adhesion dynamics by calpain-mediated proteolysis of
focal adhesion kinase (FAK). J Biol Chem. 285:11418–11426. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kubbutat MH and Vousden KH: Proteolytic
cleavage of human p53 by calpain: A potential regulator of protein
stability. Mol Cell Biol. 17:460–468. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wood DE, Thomas A, Devi LA, Berman Y,
Beavis RC, Reed JC and Newcomb EW: Bax cleavage is mediated by
calpain during drug-induced apoptosis. Oncogene. 17:1069–1078.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gao G and Dou QP: N-terminal cleavage of
bax by calpain generates a potent proapoptotic 18-kDa fragment that
promotes bcl-2-independent cytochrome c release and apoptotic cell
death. J Cell Biochem. 80:53–72. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen H, Libertini SJ, Wang Y, Kung HJ,
Ghosh P and Mudryj M: ERK regulates calpain 2-induced androgen
receptor proteolysis in CWR22 relapsed prostate tumor cell lines. J
Biol Chem. 285:2368–2374. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pelley RP, Chinnakannu K, Murthy S,
Strickland FM, Menon M, Dou QP, Barrack ER and Reddy GP:
Calmodulin-androgen receptor (AR) interaction: Calcium-dependent,
calpain-mediated breakdown of AR in LNCaP prostate cancer cells.
Cancer Res. 66:11754–11762. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Libertini SJ, Tepper CG, Rodriguez V,
Asmuth DM, Kung HJ and Mudryj M: Evidence for calpain-mediated
androgen receptor cleavage as a mechanism for androgen
independence. Cancer Res. 67:9001–9005. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
McGrath MJ, Binge LC, Sriratana A, Wang H,
Robinson PA, Pook D, Fedele CG, Brown S, Dyson JM, Cottle DL, et
al: Regulation of the transcriptional coactivator FHL2 licenses
activation of the androgen receptor in castrate-resistant prostate
cancer. Cancer Res. 73:5066–5079. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Darshan MS, Loftus MS, Thadani-Mulero M,
Levy BP, Escuin D, Zhou XK, Gjyrezi A, Chanel-Vos C, Shen R, Tagawa
ST, et al: Taxane-induced blockade to nuclear accumulation of the
androgen receptor predicts clinical responses in metastatic
prostate cancer. Cancer Res. 71:6019–6029. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Antonarakis ES and Armstrong AJ: Evolving
standards in the treatment of docetaxel-refractory
castration-resistant prostate cancer. Prostate Cancer Prostatic
Dis. 14:192–205. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mostaghel EA, Marck BT, Plymate SR,
Vessella RL, Balk S, Matsumoto AM, Nelson PS and Montgomery RB:
Resistance to CYP17A1 inhibition with abiraterone in
castration-resistant prostate cancer: Induction of steroidogenesis
and androgen receptor splice variants. Clin Cancer Res.
17:5913–5925. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Martin SK, Banuelos CA, Sadar MD and
Kyprianou N: N-terminal targeting of androgen receptor variant
enhances response of castration resistant prostate cancer to taxane
chemotherapy. Mol Oncol. 9:628–639. 2014. View Article : Google Scholar
|
|
42
|
Steinestel J, Luedeke M, Arndt A,
Schnoeller TJ, Lennerz JK, Wurm C, Maier C, Cronauer MV, Steinestel
K and Schrader AJ: Detecting predictive androgen receptor
modifications in circulating prostate cancer cells. Oncotarget. Apr
23–2015.(Epub ahead of print).
|
|
43
|
Yong EL, Ghadessy F, Wang Q, Mifsud A and
Ng SC: Androgen receptor transactivation domain and control of
spermatogenesis. Rev Reprod. 3:141–144. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Jenster G, van der Korput HA, Trapman J
and Brinkmann AO: Identification of two transcription activation
units in the N-terminal domain of the human androgen receptor. J
Biol Chem. 270:7341–7346. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gelmann EP: Molecular biology of the
androgen receptor. J Clin Oncol. 20:3001–3015. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chan SC, Li Y and Dehm SM: Androgen
receptor splice variants activate androgen receptor target genes
and support aberrant prostate cancer cell growth independent of
canonical androgen receptor nuclear localization signal. J Biol
Chem. 287:19736–19749. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kaku N, Matsuda K, Tsujimura A and Kawata
M: Characterization of nuclear import of the domain-specific
androgen receptor in association with the importin alpha/beta and
Ran-guanosine 5′-triphosphate systems. Endocrinology.
149:3960–3969. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Soderholm JF, Bird SL, Kalab P,
Sampathkumar Y, Hasegawa K, Uehara-Bingen M, Weis K and Heald R:
Importazole, a small molecule inhibitor of the transport receptor
importin-β. ACS Chem Biol. 6:700–708. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang G, Liu X, Li J, Ledet E, Alvarez X,
Qi Y, Fu X, Sartor O, Dong Y and Zhang H: Androgen receptor splice
variants circumvent AR blockade by microtubule-targeting agents.
Oncotarget. 6:23358–23371. 2015. View Article : Google Scholar : PubMed/NCBI
|