Introduction
Malignant melanoma is an aggressive form of skin
cancer that develops from the pigment-producing cells, melanocytes.
It has become more and more common worldwide (1). It is also estimated that 76,380 new
cases of melanoma were diagnosed in 2016 (2). Only early-staged melanomas can be
successfully treated, mainly through surgical excision. BRAF
mutations occur in 50% of melanoma and among the BRAF mutations
observed in melanoma, over 90% are at codon 600 (3). Although, drugs targeting BRAF
mutations, such as vemurafenib and dabrafenib, have been developed,
clinical results with molecular targeted and also recently
developed immunotherapeutic agents have not had satisfactory
results. Consequently, a deeper understanding of the molecular
biology of melanoma has been recognized as critical for
progressively identifying therapeutic targets and novel treatment
approaches (4).
Apoptosis is a process of programmed cell death that
occurs in multicellular organisms. Defective apoptotic processes
have been implicated in a wide variety of diseases, including
cancer. Drugs or treatment strategies that can restore the
apoptotic signaling pathways towards normality have the potential
to eliminate cancer cells. Thus, the mechanisms underlying
apoptosis are crucial and help in the understanding of the
pathogenesis of dysregulated apoptosis (5).
One of the best understood activation mechanisms of
apoptosis is the intrinsic pathway, which is regulated by BCL-2
family proteins (6). The BCL-2
family is composed of three functionally distinct groups:
anti-apoptotic proteins (BCL-2, BCL-xL, MCL1, BCL-w and BCL2A1),
multi-domain pro-apoptotic proteins (BAX and BAK) and BH3-only
pro-apoptotic proteins (BIM, BID, PUMA, BAD and NOXA) (6). These three groups are functionally
associated with each other; in other words, the anti-apoptotic
proteins prevent apoptosis by neutralizing the BH3-only
pro-apoptotic proteins that can directly activate pro-apoptotic
proteins BAX and BAK, and thereby restrain their pro-apoptotic
activity (7). In addition, BAK is
inhibited predominantly by BCL-xL, MCL1 and BCL2A1, whereas BAX can
be inhibited by all of the pro-survival proteins (8,9). Both
BAX and BAK have pore-forming activities that mediate the release
of apoptotic molecules, such as Smac and cytochrome c, from
the mitochondria into the cytosol (10), which triggers caspase-mediated
apoptosis (11).
Coptidis Rhizoma, called ‘Oren’ in Japanese,
is a herbal medicine used for the treatment of damp-heat syndromes
(12). Its major active component,
berberine, has drawn extensive attention due to the anticancer
effects, which are related to the effects of these traditional
herbs in the treatment of diseases by removing damp-heat and
purging fire and counteracting toxicity (13). Coptidis Rhizoma water
extracts (CR) has been reported to exhibit anticancer activity in
human pancreatic and liver cancer and leukemia (14,15).
Yet, the underlying mechanisms have not been well understood.
In the present study, we examined the effect of CR
on human melanoma cells. We showed that CR triggered apoptosis in
human melanoma cells through the mitochondrial apoptotic pathway.
The anti-apoptotic BCL-2 family proteins, BCL2A1, MCL1 and BCL-w,
played an important role in activating the multi-domain
pro-apoptotic proteins BAX and BAK induced by CR.
Materials and methods
Cell culture and herbal medicine
extraction
Human melanoma cell lines, A2058, UACC257, UACC62,
MeWo, SK-Mel-2, M14, Malme3M and mouse fibroblast cells, NIH3T3,
were cultured in RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA)
with 10% fetal bovine serum (FBS; ICN, Biomedicals, Inc., Aurora,
OH, USA), 1 mM L-glutamine (Invitrogen), 100 U/ml penicillin, and
100 µg/ml streptomycin in a humidified atmosphere of 95% air and 5%
CO2 at 37°C.
The crude Coptidis Rhizoma was added to an
appropriate volume of distilled water (v/v, 1:8) and boiled under a
low flame for 50 min. The extract was filtered and then
freeze-dried to obtain dried powder. The voucher sample of CR
(INM_ID: 00000052; University of Toyama, Toyama, Japan) was
preserved at the Cooperative Research Project by the Joint
Usage/Research Center, Institute of Natural Medicine, University of
Toyama. After reconstitution, CR was dissolved in water. Berberine
chloride hydrate was purchased from Wako Pure Chemical Industries,
Ltd. (Osaka, Japan).
Annexin V/7-AAD staining
Apoptotic cell numbers were determined using the
Muse Annexin V and Dead Cell kit (Merck Millipore, Hayward, CA,
USA) according to the manufacturer's instructions. After treatment
with CR (100 µg/ml) for 24 h, all cells were collected and diluted
with phosphate-buffered saline (PBS) containing 1% BSA as a
dilution buffer to a concentration of 5×105 cells/ml.
Then, 100 µl of cell suspension was added to 100 µl Muse Annexin V
and Dead Cell reagent, incubated for 20 min at room temperature,
and analyzed using the Muse Cell Analyzer (Merck Millipore).
Western blotting
Whole cell lysates were prepared as previously
described (16). Primary antibodies
used were BAX (D2E11) (#5023), BAK (D4E4) (#12105), PARP (#9542),
BCL-w (31H4) (#2724), MCL1 (D35A5) (#5453), A1/Bfl-1 (#4647),
BCL-xL (54H6) (#2746), and BCL-2 (50E3) (#2870) purchased from Cell
Signaling Technology, Inc. (Beverly, MA, USA), and the primary
dilution was performed at 1:1,000. α-tubulin (T9026) was purchased
from Sigma-Aldrich Company (St. Louis, MO, USA), and the primary
dilution was performed at 1:2,000.
Measurement of caspase activity
For measurement of the activities of caspase-3 and
−7, the Caspase-Glo® 3/7 assay was carried out according
to the manufacturer's instructions. This kit is based on the
cleavage of the amino acid sequence DEVD of a luminogenic substrate
by caspase-3 and −7, which results in a luminescent signal. Human
melanoma cells in exponential growth were placed at a final
concentration of 5×103 cells/90 µl/well in a
white-walled multiwell plate. The cells were treated with CR (100
µg/ml) or with the vehicle for another 18 h. Then, 100 µl of
Caspase-Glo® 3/7 reagent was added, and the plates were
incubated for another 30 min. Caspase-3 and −7 activities were
recorded using a GloMax-Multi Detection system.
Isolation of mitochondrial and
cytosolic protein
The mitochondrial and cytosolic fractions were
isolated using Mitochondria/Cytosol Fractionation kit (Abcam)
according to the manufacturer's instructions. Briefly, CR-treated
melanoma cells were harvested, and then washed in 10 ml ice-cold
PBS. After removing the supernatant, the cells were re-suspended in
1 ml 1X Cytosol Extraction Buffer Mix containing DTT and protease
inhibitors, incubated for 10 min on ice, homogenized, and
centrifuged at 700 × g for 10 min. The supernatant was centrifuged
again at 13,000 × g for 30 min. The remaining supernatant was the
cytosolic fraction, and the pellet re-suspended with Mitochondrial
Extraction Buffer Mix containing DTT and protease inhibitors was
the mitochondrial fraction. The final result was performed with
western blotting.
Real-time RT-PCR
Total RNAs were prepared using the RNeasy Plus Mini
kit (Qiagen, Hilden, Germany) and subjected to real-time PCR on an
ABI Prism 7300 sequence detection system (Life Technologies
Corporation, Carlsbad, CA, USA). The expression levels of
MCL1, BCL2A1, BCL-w and MITF mRNAs were
normalized to β-actin mRNA. The primers used were: 5′-TCG
TAA GGA CAA AAC GGG AC-3′ (sense) and 5′-CAT TCC TGA TGC CAC CTT
CT-3′ (antisense), for MCL1 mRNA; 5′-CCC GGA TGT GGA TAC CTA
TAA GGA GA-3′ (sense) and 5′-GTC ATC CAG CCA GAT TTA GGT TCA-3′
(antisense), for BCL2A1 mRNA; 5′-TCA ACA AGG AGA TGG AAC
CAC-3′ (sense) and 5′-ATA GAG CTG TGA ACT CCG CC-3′ (antisense),
for BCL-w mRNA; 5′-GTC ATC CAG CCA GAT TTA GGT TCA-3′
(sense) and 5′-CAT TGT TAT GCT GGA AAT GCT AGA A-3′ (antisense),
for MITF mRNA; and 5′-GCA CAG AGC CTC GCC TT-3′ (sense) and
5′-GTT GTC GAC GAC GAG CG-3′ (antisense), for β-actin
mRNA.
Cell viability assay
Cell viability was measured using CellTiter-Glo 2.0
assay (Promega Corporation, Madison, WI, USA) according to the
manufacturer's instructions. Briefly, melanoma cells were seeded
into 96-well plates (5–10×103 cells/90 µl/well) and
incubated overnight. They were further cultured with CR (50 or 100
µg/ml) or berberine (12.5 or 25 µM) for 24, 48 and 72 h, and lysed
with firefly luciferase reagents. The luciferase activity was
measured by GloMax Multi-detection system (Promega).
Intracellular staining and flow
cytometric analysis of BAK activation
A2058 melanoma cells were seeded into 6-well plates
(2×105/2 ml/well) and incubated overnight. They were
further cultured with CR (100 µg/ml) for 24 h. After being
harvested and washed by PBS, the cells were fixed and permeabilized
by adding Fixation/Permeabilization solution (BD Cytofix/Cytoperm™
Fixation/Permeabilization kit; BD Biosciences, Franklin Lakes, NJ,
USA) for 20 min at 4°C (500 µl/tube). After washing twice with
Perm/Wash buffer (BD Biosciences), the cells were incubated with
anti-BAK (Ab-1) Mouse mAb (AM03, Merck) (1:50) for 1 h at room
temperature. After washing with Perm/Wash buffer twice, cells were
incubated with FITC-labeled anti-mouse antibody (F0313, DAKO)
(1:25) for 1 h at room temperature. Finally, the stained cells
(1×104/sample) were analyzed on a BD Accuri C6 flow
cytometer (BD Biosciences). For quantitation and comparison, the
median of the fluorescence intensity values was calculated using
FlowJo software.
Establishment of A2058
BAX−/− and BAK−/− melanoma cell lines
For knocking out BAX or BAK, the target sequences
containing the PAM sequences were inserted into GeneArt CRISPR
nuclease vector with OFP (Life Technologies, Gaithersburg, MD,
USA). The inserted sequences used were: 5′-AAG GAC GAA ACA CCG TCT
GGG CTT CGG CTA CCG TCG TTT TAG AGC TAG AA-3′ for BAK; and 5′-AAG
GAC GAA ACA CCG CCG AGA GGT CTT TTT CCG AGG TTT TAG AGC TAG AA-3′
for BAX.
For the establishment of BAX- or BAK-knockout cells,
A2058 cells were transfected with GeneArt CRISPR nuclease vector
containing BAX or BAK and selected by FACS sorting methods. After
single cell cloning, the targeted sequences were confirmed by
sequencing.
Statistical analysis
Statistical significance was calculated using Excel
software (Microsoft Corporation, Redmond, WA, USA). More than three
means were compared using two-way analysis of variance (ANOVA) with
Bonferroni correction. P<0.05 was considered statistically
significant.
Results
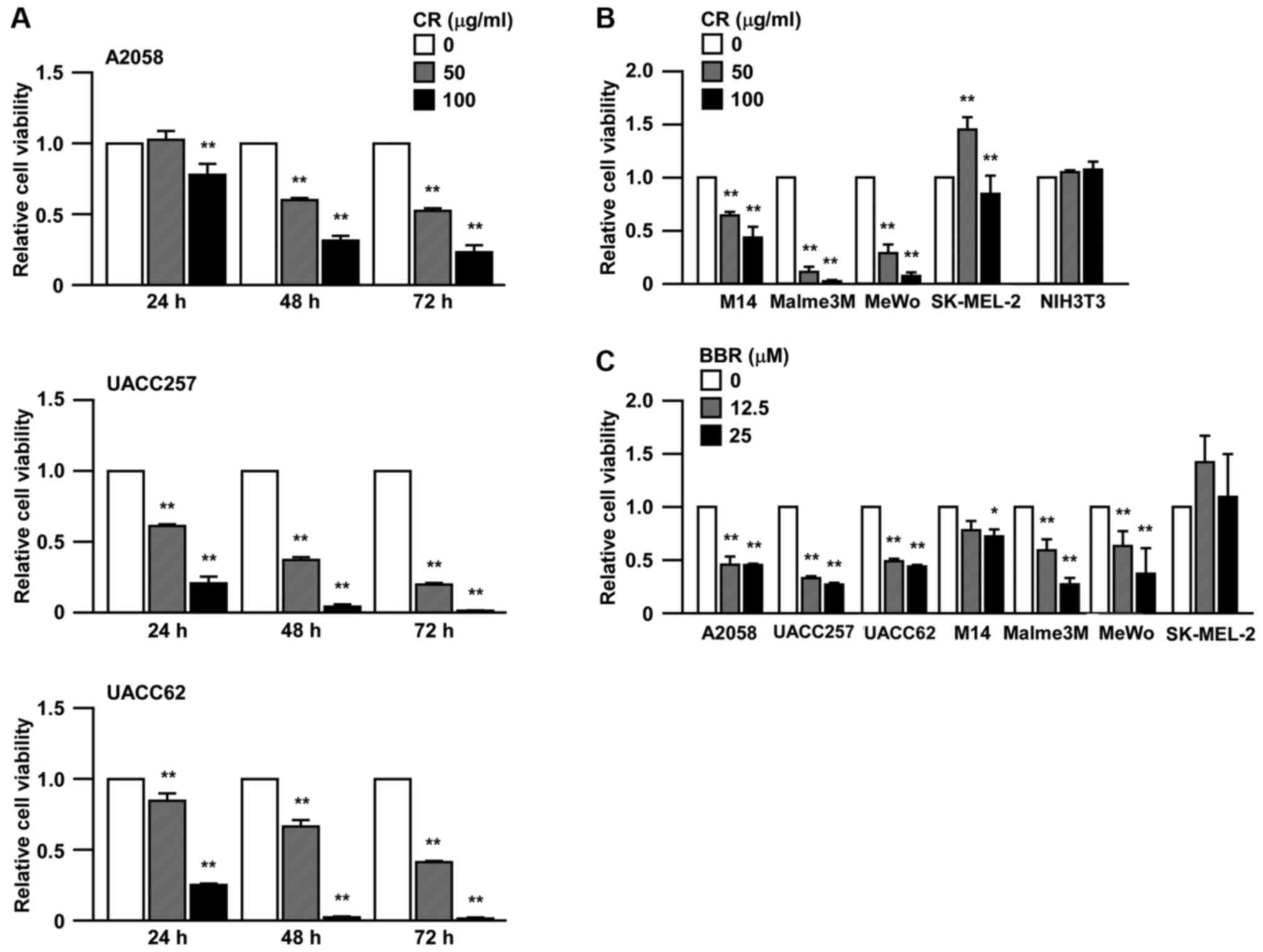
CR inhibits the cell viability in some
melanoma cell lines
We first tested the inhibitory effects on cell
viability following CR treatment in three human melanoma cell lines
with BRAF V600E mutation, A2058, UACC257 and UACC62. After
treatment with CR, the cell viability was significantly reduced in
a dose- and time-dependent manner in all three cell lines (Fig. 1A), similar to other cancers after
treatment of 100 µg/ml CR (17–19).
In addition, we assessed the cell viability in other melanoma cell
lines, M14 (BRAF V600E), Malme3M (BRAF V600E), MeWo (no mutation in
BRAF and NRAS), and SK-MEL-2 (NRAS Q61R), with a different genetic
background (Fig. 1B). Except
SK-MEL-2, the cell viability was strongly inhibited by CR in M14,
Malme3M and MeWo cell lines. The growth inhibition by CR was not
detected in NIH3T3 mouse fibroblast cells. We also used one of the
major components of CR, berberine (BBR), to treat the melanoma
cells (Fig. 1C). BBR also inhibited
the melanoma cell viability except for SK-MEL-2 cells.
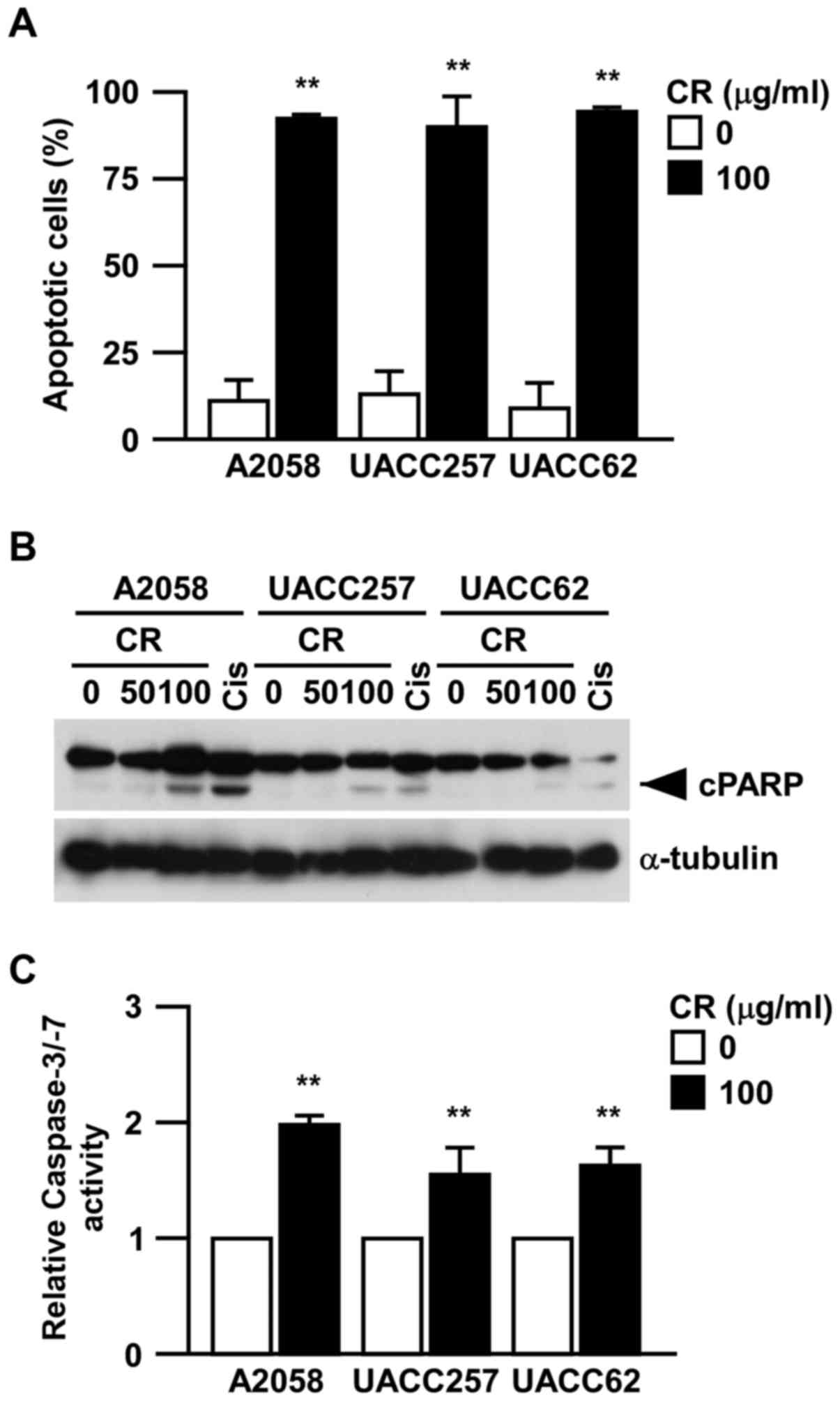
CR induces apoptosis in human melanoma
cells
To investigate whether or not the suppression of
cell viability by CR was attributed to apoptosis, we examined the
apoptotic cells using flow cytometric analysis with Annexin V
staining (Fig. 2A). Strikingly, in
all three human melanoma cell lines, the number of Annexin
V-positive cells was significantly increased by CR. Furthermore, we
also detected the induction of cleavages of PARP by CR in all
melanoma cell lines (Fig. 2B) and
the activation of caspases by CR using luciferase-based methods
(Fig. 2C). These data suggest that
the inhibition of cell viability by CR may be mediated largely
through the induction of caspase-mediated apoptosis.
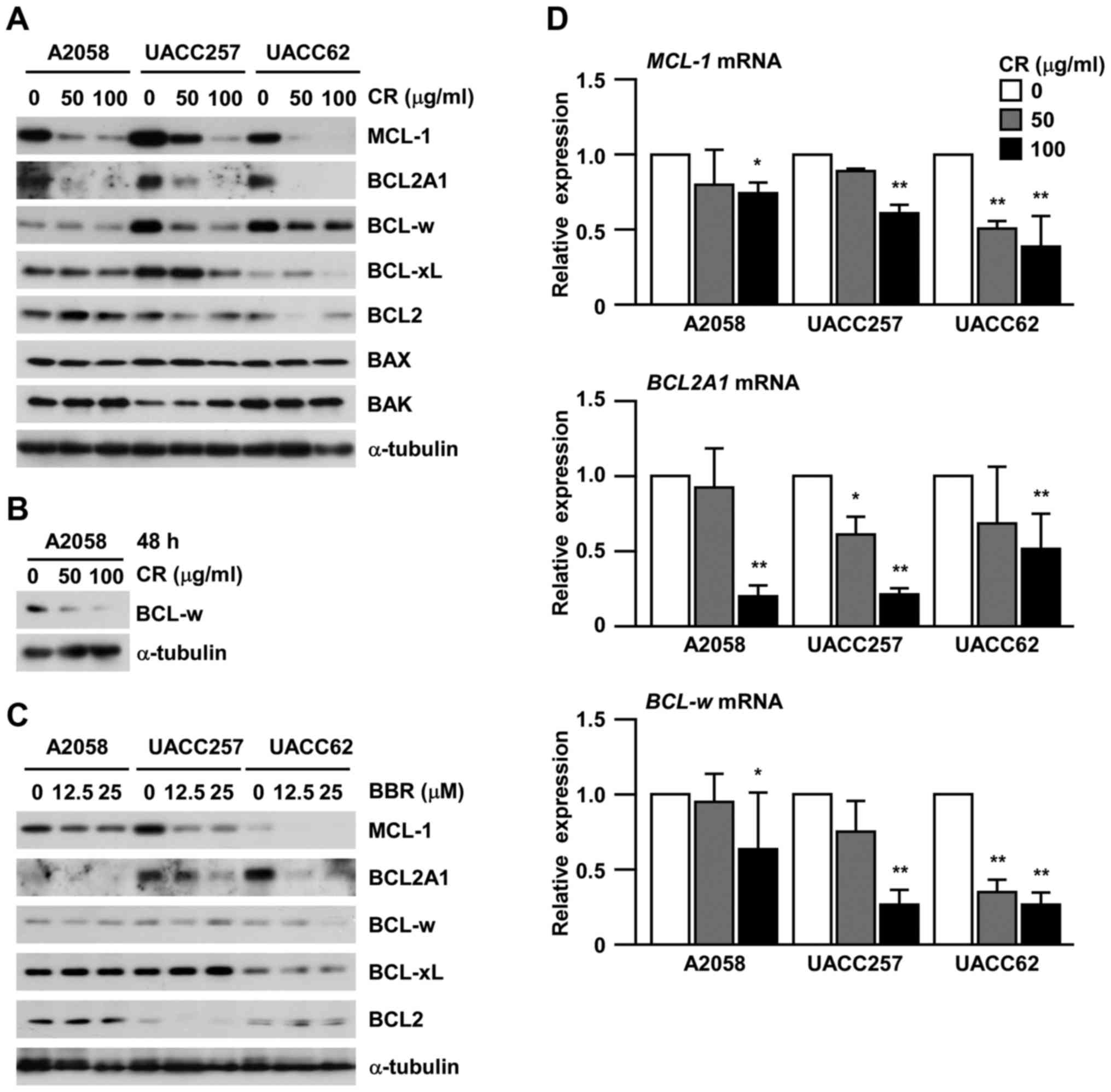
CR induces apoptosis by inhibiting the
anti-apoptotic proteins, MCL1, BCL2A1 and BCL-w in human melanoma
cells
Given that certain crude extracts from natural
medicines induce apoptosis through the intrinsic pathway (20,21),
we next determined the involvement of anti-apoptotic BCL-2 family
members, including BCL2, BCL-xL, BCL-w, MCL1 and BCL2A1, in
CR-induced apoptosis (Fig. 3A).
Although BCL2 and BCL-xL were not affected, the protein levels of
anti-apoptotic proteins, MCL1 and BCL2A1, were consistently
suppressed by CR in all three cell lines in a dose-dependent
manner. In the case of BCL-w, UACC257 and UACC62 cells showed
suppression by CR at 24 h, but A2058 cells did not. When A2058
cells were treated with CR for 48 h, the suppression of BCL-w was
detected (Fig. 3B). In the case of
multi-domain pro-apoptotic proteins of the BCL-2 family, BAX and
BAK, their protein expression levels did not show any significant
change by CR treatment (Fig. 3A).
In addition, we assessed the effects of BBR on anti-apoptotic BCL-2
family members. MCL1 and BCL2A1 were suppressed by BBR, although
BCL-w was not suppressed in the A2058 and UACC257 cells (Fig. 3C). Consistently, we detected the
suppression of mRNA levels of MCL1, BCL2A1 and
BCL-w, in all three cell lines (Fig. 3D). This suggests that CR may induce
apoptosis by suppressing the mRNA expression of MCL1,
BCL2A1 and BCL-w in human melanoma cells.
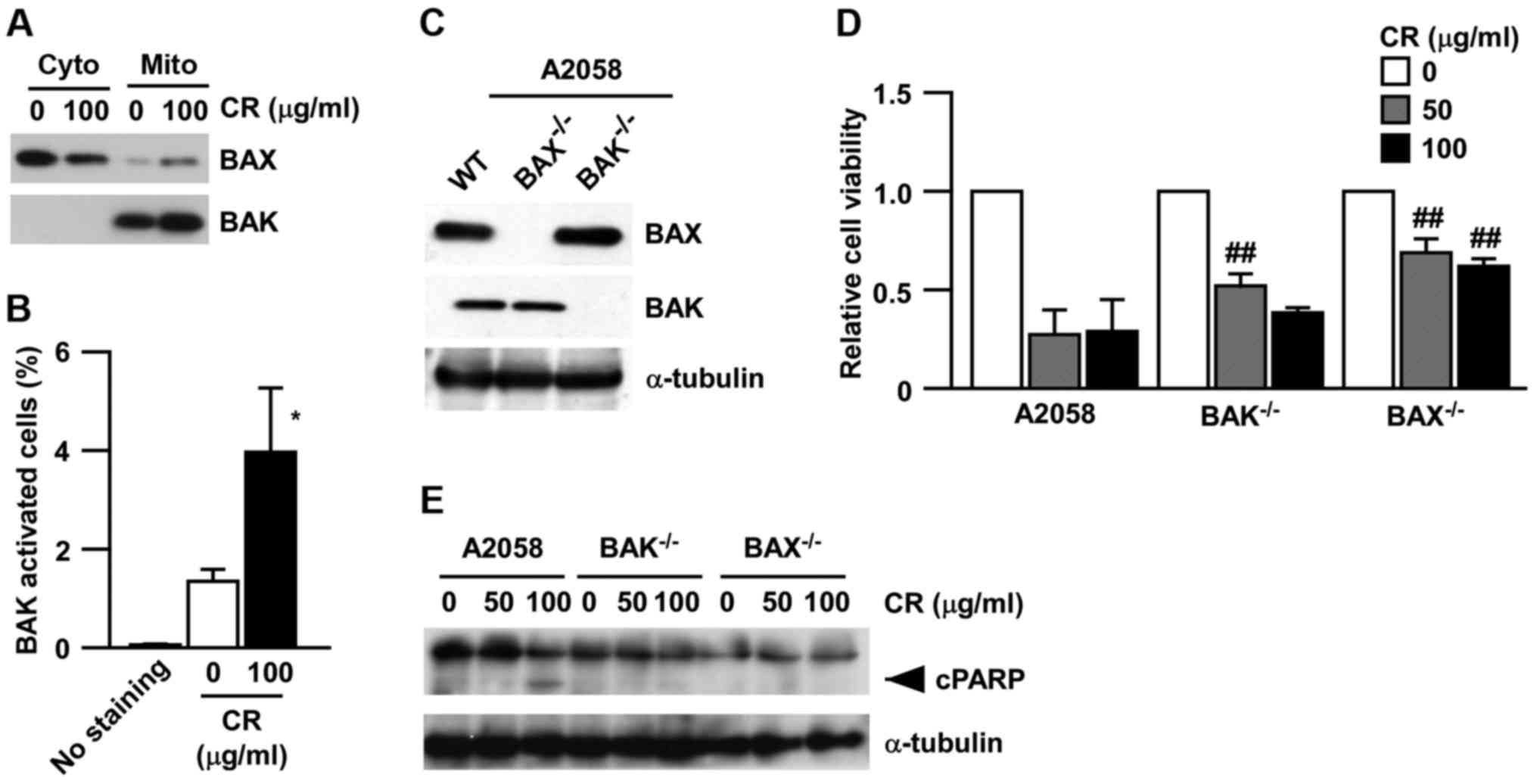
CR-induced apoptosis is mediated
through the activation of BAX/BAK
To investigate the responsible effectors of BCL2
family by CR, we focused on multi-domain pro-apoptotic proteins of
the BCL-2 family, BAX and BAK, both of which were not affected by
CR at their protein levels (Fig.
3A). These two proteins have pore-forming activities that
mediate the release of apoptotic molecules, such as Smac and
cytochrome c (10). BAX has
been known as a cytosolic protein, which translocates to
mitochondria during its activation (22). In contrast, BAK is expressed on the
mitochondrial membrane and shows a conformational change during its
activation, which is recognized by a specific antibody (23). As shown in Fig. 4A, BAX translocation from the cytosol
to mitochondria was induced by CR, although BAK only existed in the
mitochondrial fraction, not in the cytosolic fraction. However, BAK
conformational change was also detected following CR treatment by
intracellular staining using a specific antibody, which
specifically detects the conformational change of BAK (Fig. 4B). These results suggest that CR
activated both BAX and BAK in human melanoma cells. Furthermore, to
check the significance of BAX/BAK, we established A2058 cell lines
with knockout of BAX or BAK (named A2058 BAX−/− and
A2058 BAK−/−, respectively) (Fig. 4C) and treated these established cell
lines with CR. As shown in Fig. 4D,
both A2058 BAK−/− and A2058 BAX−/− increased
their cell viability following treatment with CR. In addition, PARP
cleavage was significantly reduced in both A2058 BAK−/−
and A2058 BAX−/− cell lines, compared with the A2058
parental cells (Fig. 4E). These
results strongly support that both BAX and BAK are required for
CR-induced apoptosis.
Discussion
In the present study, we determined that Coptidis
Rhizoma water extract (CR) suppressed cell viability through the
induction of apoptosis in human melanoma. In addition, the
apoptosis was mediated through the suppression of BCL-w, MCL1 and
BCL2A1, resulting in the activation of BAX/BAK.
CR and its active component berberine (BBR) have
been revealed as having cytotoxicity effects on various types of
cancer such as breast, colon and lung (24–26).
Although BBR is the major anticancer component in CR, some studies
have reported that CR may be more effective than BBR alone
(14,27). In the present study, both BBR and CR
suppressed cell growth in the human melanoma cells (Fig. 1), although their effects on BCL-w
protein were different (Fig. 3).
This implies that the underlying mechanisms of CR may be
complicated and that BBR alone may not be sufficient to understand
the mechanisms of actions of CR. Further investigation is required
for identifying the active component in CR with anticancer
activity; in other words, CR itself or traditional Kampo/Chinese
medicines containing CR may be applicable for melanoma, although
its mechanism of action has not been determined in detail.
We identified that CR suppressed MCL1, BCL2A1 and
BCL-w in human melanoma cells (Fig.
3A). It has been reported that MCL1 is known as an oncogene
which is commonly amplified in various types of cancers, and that
MCL1-amplified cancer is more sensitive to MCL1 suppression
(28). In contrast, BCL2A1 has been
confirmed as a lineage-specific anti-apoptotic melanoma oncogene
with amplification, which confers resistance to BRAF inhibition
(29). In addition, BCL2A1 is also
regulated by the melanoma-specific oncogene, MITF, which is also
amplified specifically in melanoma UACC62 and UACC257 cells
(30). In the present study,
MITF mRNA and protein were also suppressed by CR (data not
shown), supporting the functional significance of the MITF/BCL2A1
axis in CR-induced apoptosis. Various studies indicate that
inhibition of both MCL1 and BCL2A1 expression could lead to ~70%
cell death in melanoma cells, which achieves better anticancer
effects (8,31,32).
In the human melanoma cells used, only SK-MEL-2 was relatively
insensitive to CR (Fig. 1), which
does not have any gene copy number alteration in the MCL1,
BCL2A1 and MITF regions. In addition, the effects of
CR or BBR on NRAS-mutant melanoma cells including SK-MEL-2 cells
remain to be further investigated since only NRAS-mutant melanoma
showed the induction of cell growth at a lower dose of CR and BBR.
Collectively, BCL2A1 and MITF amplification are
specific to melanoma, thus CR may be applicable for other cancer
patients with MCL1 amplification.
Anticancer drugs, such as cisplatin, exhibit high
cytotoxicity even in normal cells. CR does not show any adverse
side-effects, which is supported historically and clinically by its
usage for >2,000 years as a traditional Kampo medicine in Japan
or traditional Chinese medicine in China. However, in addition to
cytotoxicity, many other pharmacological effects of CR have been
reported including anti-inflammation, anti-infection and
anti-hypertension (13). In
addition, CR and BBR prevent cachexia (33), which is characterized by profound
weight loss, anorexia, and weakness. Thus, CR and other traditional
Kampo/Chinese medicines containing CR may be useful as anticancer
drugs or as adjuvant therapeutic agents.
In summary, we demonstrated that CR induced
apoptosis by BAX and BAK activation in human melanoma cells, which
was mediated through the suppression of MCL1, BCL2A1 and BCL-w.
This natural herbal medicine, CR or various formulations containing
CR may constitute an effective and safe treatment for human
melanoma.
Acknowledgements
The present study was supported in part by
Grant-in-Aid 16K18413 for Young Scientists (B) (S.Y.) and by
Grant-in-Aid 15K08902 for Scientific Research (C) (I.S.) from the
Ministry of Education, Culture, Sports, Scienceand Technology
(Japan), and The Uehara Memorial Foundation (S.Y.).
References
|
1
|
Liu J, Fukunaga-Kalabis M, Li L and Herlyn
M: Developmental pathways activated in melanocytes and melanoma.
Arch Biochem Biophys. 563:13–21. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ascierto PA, Kirkwood JM, Grob JJ, Simeone
E, Grimaldi AM, Maio M, Palmieri G, Testori A, Marincola FM and
Mozzillo N: The role of BRAF V600 mutation in melanoma. J Transl
Med. 10:852012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Karachaliou N, Pilotto S, Teixidó C,
Viteri S, González-Cao M, Riso A, Morales-Espinosa D, Molina MA,
Chaib I, Santarpia M, et al: Melanoma: Oncogenic drivers and the
immune system. Ann Transl Med. 3:2652015.PubMed/NCBI
|
|
5
|
Wong RS: Apoptosis in cancer: From
pathogenesis to treatment. J Exp Clin Cancer Res. 30:872011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Juin P, Geneste O, Gautier F, Depil S and
Campone M: Decoding and unlocking the BCL-2 dependency of cancer
cells. Nat Rev Cancer. 13:455–465. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Czabotar PE, Lessene G, Strasser A and
Adams JM: Control of apoptosis by the BCL-2 protein family:
Implications for physiology and therapy. Nat Rev Mol Cell Biol.
15:49–63. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Simmons MJ, Fan G, Zong WX, Degenhardt K,
White E and Gélinas C: Bfl-1/A1 functions, similar to Mcl-1, as a
selective tBid and Bak antagonist. Oncogene. 27:1421–1428. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Willis SN, Chen L, Dewson G, Wei A, Naik
E, Fletcher JI, Adams M and Huang DC: Proapoptotic Bak is
sequestered by Mcl-1 and Bcl-xL, but not Bcl-2, until displaced by
BH3-only proteins. Genes Dev. 19:1294–1305. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hartman ML and Czyz M: Anti-apoptotic
proteins on guard of melanoma cell survival. Cancer Lett.
331:24–34. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fischer U, Jänicke RU and Schulze-Osthoff
K: Many cuts to ruin: A comprehensive update of caspase substrates.
Cell Death Differ. 10:76–100. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang N, Tan HY, Li L, Yuen MF and Feng Y:
Berberine and Coptidis Rhizoma as potential anticancer agents:
Recent updates and future perspectives. J Ethnopharmacol.
176:35–48. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tang J, Feng Y, Tsao S, Wang N, Curtain R
and Wang Y: Berberine and Coptidis Rhizoma as novel antineoplastic
agents: A review of traditional use and biomedical investigations.
J Ethnopharmacol. 126:5–17. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hara A, Iizuka N, Hamamoto Y, Uchimura S,
Miyamoto T, Tsunedomi R, Miyamoto K, Hazama S, Okita K and Oka M:
Molecular dissection of a medicinal herb with anti-tumor activity
by oligonucleotide microarray. Life Sci. 77:991–1002. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lin CC, Ng LT, Hsu FF, Shieh DE and Chiang
LC: Cytotoxic effects of Coptis chinensisEpimedium sagittatum
extracts and their major constituents (berberine, coptisine and
icariin) on hepatoma and leukaemia cell growth. Clin Exp Pharmacol
Physiol. 31:65–69. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sakurai H, Suzuki S, Kawasaki N, Nakano H,
Okazaki T, Chino A, Doi T and Saiki I: Tumor necrosis
factor-alpha-induced IKK phosphorylation of NF-kappaB p65 on serine
536 is mediated through the TRAF2, TRAF5, and TAK1 signaling
pathway. J Biol Chem. 278:36916–36923. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim YJ, Kang SA, Hong MS, Park HJ, Kim MJ,
Park HJ and Kim HK: Coptidis Rhizoma induces apoptosis in human
colorectal cancer cells SNU-C4. Am J Chin Med. 32:873–882. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu J, He C, Zhou K, Wang J and Kang JX:
Coptis extracts enhance the anticancer effect of estrogen receptor
antagonists on human breast cancer cells. Biochem Biophys Res
Commun. 378:174–178. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tan HY, Wang N, Tsao SW, Zhang Z and Feng
Y: Suppression of vascular endothelial growth factor via
inactivation of eukaryotic elongation factor 2 by alkaloids in
Coptidis rhizome in hepatocellular carcinoma. Integr Cancer Ther.
13:425–434. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cao C, Liu B, Zeng C, Lu Y, Chen S, Yang
L, Li B and Li Y and Li Y: A polymethoxyflavone from Laggera
pterodonta induces apoptosis in imatinib-resistant K562R cells via
activation of the intrinsic apoptosis pathway. Cancer Cell Int.
14:1372014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim MJ, Kwon SB, Ham SH, Jeong ES, Choi
YK, Choi KD, Hong JT, Jung SH and Yoon DY: H9 inhibits tumor growth
and induces apoptosis via intrinsic and extrinsic signaling pathway
in human non-small cell lung cancer xenografts. J Microbiol
Biotechnol. 25:648–657. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tikhomirov O and Carpenter G: Bax
activation and translocation to mitochondria mediate EGF-induced
programmed cell death. J Cell Sci. 118:5681–5690. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pichichero E, Cicconi R, Mattei M and
Canini A: Chrysin-induced apoptosis is mediated through p38 and Bax
activation in B16-F1 and A375 melanoma cells. Int J Oncol.
38:473–483. 2011.PubMed/NCBI
|
|
24
|
Kang JX, Liu J, Wang J, He C and Li FP:
The extract of huanglian, a medicinal herb, induces cell growth
arrest and apoptosis by upregulation of interferon-beta and
TNF-alpha in human breast cancer cells. Carcinogenesis.
26:1934–1939. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Peng PL, Hsieh YS, Wang CJ, Hsu JL and
Chou FP: Inhibitory effect of berberine on the invasion of human
lung cancer cells via decreased productions of
urokinase-plasminogen activator and matrix metalloproteinase-2.
Toxicol Appl Pharmacol. 214:8–15. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu K, Yang Q, Mu Y, Zhou L, Liu Y, Zhou Q
and He B: Berberine inhibits the proliferation of colon cancer
cells by inactivating Wnt/β-catenin signaling. Int J Oncol.
41:292–298. 2012.PubMed/NCBI
|
|
27
|
Li XK, Motwani M, Tong W, Bornmann W and
Schwartz GK: Huanglian, A chinese herbal extract, inhibits cell
growth by suppressing the expression of cyclin B1 and inhibiting
CDC2 kinase activity in human cancer cells. Mol Pharmacol.
58:1287–1293. 2000.PubMed/NCBI
|
|
28
|
Mukherjee N, Lu Y, Almeida A, Lambert K,
Shiau CW, Su JC, Luo Y, Fujita M, Robinson WA, Robinson SE, et al:
Use of a MCL-1 inhibitor alone to de-bulk melanoma and in
combination to kill melanoma initiating cells. Oncotarget. Apr
12–2016.(Epub ahead of print). doi: 10.18632/oncotarget.8695.
|
|
29
|
Nessling M, Kern MA, Schadendorf D and
Lichter P: Association of genomic imbalances with resistance to
therapeutic drugs in human melanoma cell lines. Cytogenet Cell
Genet. 87:286–290. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Haq R, Yokoyama S, Hawryluk EB, Jönsson
GB, Frederick DT, McHenry K, Porter D, Tran TN, Love KT, Langer R,
et al: BCL2A1 is a lineage-specific antiapoptotic melanoma oncogene
that confers resistance to BRAF inhibition. Proc Natl Acad Sci USA.
110:4321–4326. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen L, Willis SN, Wei A, Smith BJ,
Fletcher JI, Hinds MG, Colman PM, Day CL, Adams JM and Huang DC:
Differential targeting of prosurvival Bcl-2 proteins by their
BH3-only ligands allows complementary apoptotic function. Mol Cell.
17:393–403. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Senft D, Berking C, Graf SA, Kammerbauer
C, Ruzicka T and Besch R: Selective induction of cell death in
melanoma cell lines through targeting of Mcl-1 and A1. PLoS One.
7:e308212012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Iizuka N, Hazama S, Yoshimura K, Yoshino
S, Tangoku A, Miyamoto K, Okita K and Oka M: Anticachectic effects
of the natural herb Coptidis Rhizoma and berberine on mice bearing
colon 26/clone 20 adenocarcinoma. Int J Cancer. 99:286–291. 2002.
View Article : Google Scholar : PubMed/NCBI
|