Introduction
Gliomas are the most common primary central nervous
system tumors and account for approximately 80% of malignant brain
tumors (1). Despite recent
improvements in both diagnostic methods and therapeutic
interventions, including surgical resection combined with
postoperative radiation and chemotherapy, overall survival has not
markedly improved for glioma patients (2), especially for high-grade gliomas
(grades III and IV). Therefore, it is necessary to obtain a better
understanding of the molecular mechanisms involved in
carcinogenesis and progression to develop novel and effective
therapeutic strategies for this malignancy.
MicroRNAs (miRNAs) are a class of small non-coding,
regulatory RNA molecules that mediate the expression levels of
specific proteins (3). Through
modulating the protein levels of their target genes, miRNAs have
been implicated in various biological processes, including cell
proliferation, cell cycle progression, apoptosis, migration and
differentiation (4). Moreover,
mounting evidence suggests that the deregulation of miRNA
expression plays a pivotal role in the initiation and progression
of numerous cancers (5–7), including gliomas (8–10). For
example, miR-217 inhibited cell proliferation and invasion by
targeting Runx2 in human glioma (8). In addition, miR-184 inhibited cell
proliferation and invasion and specifically targeted TNFAIP2 in
glioma (9). miR-195 inhibited the
proliferation of human glioma cells by directly targeting cyclin D1
and cyclin E1 (10). Consequently,
the use of miRNAs in clinical anticancer therapy is both promising
and expected (5–10).
miR-543 is a member of a miRNA cluster located in
the imprinted DLK1-DIO3 region on human chromosome 14 (11). As a member of the miRNA family,
miR-543 is aberrantly expressed in several human cancers, such as
hepatocellular carcinoma and colorectal cancer (12,13),
demonstrating the role of miR-543 in tumorigenesis. However, the
role of miR-543 in glioma remains unknown. In the present study,
the expression level of miR-543 in glioma cell lines and tissues
was investigated. A series of in vitro and in vivo
experiments were performed to elucidate the function of miR-543 in
glioma. In addition, for further exploration, a label-free
quantitative proteomic approach was applied in the present study to
determine the landscape of differentially expressed proteins
associated with miR-543-mediated carcinogenesis in glioma.
Materials and methods
Human specimens
The human glioma and normal brain specimens from
patients were collected at the Department of Neurosurgery, the
Second Affiliated Hospital of Soochow University. All glioma
specimens were confirmed by pathological diagnosis and classified
according to the World Health Organization (WHO) criteria. The
study comprised of 6 WHO I, 22 WHO II, 16 WHO III and 18 WHO IV
glioma samples. Ten normal brain tissues were obtained from
patients with cerebral injury. Specimens were immediately
snap-frozen in liquid nitrogen after surgical removal and stored at
−80°C before use. Informed consent was obtained from all patients,
and the study was approved by the Ethics Committee of the Second
Affiliated Hospital of Soochow University.
Cell culture
Human glioma cell lines (U87, U251, U343, A172, T98G
and LN229) were purchased from the Cell Bank of Chinese Academy of
Science. Primary normal human astrocytes (NHA) were kindly provided
by Professor Ming Li (Department of Neurosurgery, The Second
Affiliated Hospital of Soochow University, Suzhou, China). The
glioma cells were maintained in Dulbeccos modified Eagles medium
(DMEM) supplemented with 10% fetal bovine serum (FBS) and
antibiotics (100 U/ml penicillin and 100 U/ml streptomycin; Gibco,
Grand Island, NY, USA). Cells were grown in a 37°C incubator with
5% CO2. Primary human astrocytes were maintained in
astrocyte media (Sciencell Research Laboratories, Carlsbad, CA,
USA) containing 10% FBS, 1% astrocyte growth supplement and 1%
penicillin/streptomycin.
RNA extraction and real-time
quantitative PCR (qRT- PCR)
Total RNA was extracted from cell lines and frozen
tissues using TRIzol reagent (Invitrogen, Carlsbad, CA, USA).
Complementary DNA (cDNA) was synthesized using an miRNA reverse
transcription kit (Shanghai GenePharma Co., Ltd., Shanghai, China)
according to the manufacturers instructions. Then, the expression
levels of miR-543 were quantified using a QuantiNova SYBR-Green PCR
kit (Qiagen, Hilden, Germany) with the ABI 7900HT Fast system
(Applied Biosystems, Foster City, CA, USA). Primer sequences of
miR-543 were CAGTGCTAAAACATTCGCGG (forward) and
TATGGTTGTTCACGACTCCTTCAC (reverse). The U6 gene was used as an
internal reference. The primer sequences were synthesized by
Shanghai GenePharma. The reaction was performed for 40 cycles under
following conditions: denaturation at 95°C for 3 min, 95°C for 10
sec and 55°C for 30 sec. The relative expression ratio of miR-543
was calculated using the 2−ΔΔCT method.
Cell transfection
U87 and U251 cells were transfected with the miR-543
mimic and the corresponding negative control (miR-NC), which were
obtained from Shanghai GenePharma. The sequences were as follows:
miR-543 mimic, 5-UGGCG GAGAACUGAUAAGGGUCCUUAUCAGUUCUCCGUCC AUU-3;
negative control (miR-NC), 5-UUCUCCGAACGUG
UCACGUTTACGUGACACGUUCGGAGAATT-3.
miR-543 mimic or miR-NC was transfected into the
glioma cells when the cells reached 60–70% confluence using
Lipofectamine 3000 (Invitrogen) according to the manufacturers
instructions. Transfection efficiencies were determined in every
experiment at 48 h after transfection by qRT-PCR.
Cell proliferation assay
The number of viable cells was assayed using a Cell
Counting kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan) per
the manufacturers protocol. The cells were seeded into 96-well
plates at a density of 2×103 cells/well. Then, 10 µl of
the CCK-8 solution was added to each well every day for 6 days, and
the plates were incubated for 1 h at 37°C. The optical density (OD)
at 450 nm was measured using a microplate reader (Bio-Rad
Laboratories, Hercules, CA, USA).
Colony formation assay
The colony formation rate was measured using the
plate colony formation assay. Then, 1,000 cells of each group were
added to each well of the 6-well plates and cultured for 10 days in
DMEM medium containing 10% FBS. Then, the cells were washed with
phosphate-buffered saline (PBS) and fixed using a crystal violet
cell colony staining kit (Genemed, Shanghai, China). Images of the
colonies were taken and the colonies were counted under a
microscope (Olympus, Tokyo, Japan).
Cell cycle assay
For cell cycle analysis, cells in each group were
fixed with cold 70% ethanol at 4°C for 12 h. The cells were washed
twice with PBS and then stained with propidium iodide (PI) with
RNase in PBS. The treated cells were then evaluated by flow
cytometry (Beckman Coulter, Inc., Brea, CA, USA).
Cell apoptosis assay
Cellular apoptosis levels were detected using an
Annexin V-PE apoptosis detection kit PE (BD Biosciences, San Diego,
CA, USA) by flow cytometer (Beckman Coulter). Cells in each group
were collected and resuspended in 400 µl of 1X binding buffer
containing 50 µl Annexin V-PE and 7-AAD (7-amino-actinomycin D) at
4°C in the dark for 15 min.
Wound healing assay
The wound healing assay was conducted to detect cell
migration. Cells were seeded in 6-well plates and cultured to 80%
confluence. A 10-µl pipette tip was applied to generate a linear
wound. Then, cells were cultured for 24 h and observed under a
microscope (Olympus). The widths of wounds were determined at 0 and
24 h.
Cell invasion assay
For the cell invasion assay, Transwell chambers
(Corning, Inc., Corning, NY, USA) were covered with Matrigel (BD
Bioscience), and 3×104 cells suspended in 150 µl of
serum-free medium were added to the upper chambers. The lower
chambers were filled with 600 µl of DMEM with 10% FBS. After 24 h
of incubation, the cells on the upper surface were removed, and the
cells on the lower surface were stained with 0.1% crystal violet
for 15 min. The stained cells were counted in 10 randomly selected
fields under a microscope (magnification, ×400; Olympus).
Tumor xenograft growth assay
All procedures and experiments involving animals in
this study were approved by the Institutional Animal Care and Use
Committee and the Local Ethics Board. Female immunodeficient nude
mice of 4–5 weeks of age were purchased from Shanghai SLAC
Laboratory Animal Co., Ltd. (Shanghai, China) and maintained under
specific pathogen-free (SPF) conditions. U87 and U251 cells and
glioma cells (2×106) harboring miR-543 or miR-NC were
separately injected subcutaneously (s.c.) into the flanks of nude
mice (n=5). Growth curves and tumor volumes were measured every
other day. The tumor volume was determined according to the
following formula: TV (mm3) = 1/2 × width2 ×
length. On the 35th day after the injection, the mice were
sacrificed and the tumors were removed, weighed and
photographed.
Protein extraction
U87 cells were transfected with miR-543 mimic or
miR-NC. Prepared samples were first frozen to a dry powder with a
vacuum freeze drier. The freeze-dried powder was dissolved in 200
µl of TEAB (tetraethylammonium bromide) dissolution buffer and 800
µl of cold acetone containing 10 mM DTT (dithiothreitol). Then, the
resuspended powder was incubated for ~2 h. After centrifugation at
13,000 r/min for 20 min at 15°C, the precipitate was collected and
mixed with 800 µl of cold acetone containing a solution of 10 mM
DTT for 1 h at 56°C to break the protein disulfide bonds. The
sample was centrifuged at 13,000 r/min for 20 min at 15°C. The
precipitate was collected, dried and stored at −80°C for later
use.
Protein quantification and
digestion
Total protein concentration was measured using the
Bradford method (14). For each
sample, 100 µg of protein was dissolved in 500 µl of dissolution
buffer and then diluted with 500 µl of 50 mM
NH4HCO3. After reduction and alkylation, 2 µg
of trypsin was added and then incubated overnight at 37°C for
protein digestion. After protein digestion, an equal volume of 0.1%
FA (formic acid) was added for acidification. Peptides were
purified thrice using a Strata-X C18 pillar, washed with 0.1% FA +
5% ACN (acetonitrile) twice, and eluted with 1 ml of 0.1% FA + 80%
ACN. Eluted peptides were dried with a vacuum concentration
meter.
LC-MS/MS analysis
LC-MS/MS analysis was performed on an AB SCIEX
NanoLC-MS/MS system (Triple TOF 5600 plus; AB Sciex, Framingham,
MA, USA). Samples were chromatographed using a 120-min gradient
from 2 to 35% (buffer A 0.1% (v/v) FA, 5% (v/v) ACN, buffer B 0.1%
(v/v) FA, 95% (v/v) ACN] after direct injection onto a 20-cm
PicoFrit emitter (New Objective, Inc., Woburn, MA, USA) packed to
20 cm with a Magic C18 AQ 3-µm 200-Å stationary phase. MS1 spectra
were collected in the range from 360 to 1,460 m/z for 250 ms. The
40 most intense precursors with a charge state of 2–5 were selected
for fragmentation, and MS2 spectra were collected in the range
50–2,000 m/z for 100 ms. Precursor ions were excluded from
reselection for 15 sec.
Data analysis
The original MS/MS file data were submitted to
ProteinPilot Software v4.5 for data analysis. For protein
identification, the Paragon algorithm (15), which was integrated into
ProteinPilot, was compared against the uniprot homo database for
database searching. The parameters were set as follows: the
instrument was TripleTOF 5600 plus, cysteine modified with
iodoacetamide; biological modifications were selected as ID focus
and trypsin was selected for protein digestion. For false discovery
rate (FDR) calculation, an automatic decoy database search strategy
was employed to estimate FDR using the PSPEP (Proteomics System
Performance Evaluation Pipeline Software, integrated in the
ProteinPilot Software). Only unique peptides with global FDR values
of fit <1% were considered for further analysis. Skyline v2.5
software was employed for peptide and protein quantification. For
differentially expressed protein (DEP) determination, fold-changes
were calculated as the average comparison pairs among biological
replicates. Proteins with a fold-change >2 or <0.5 were
considered to be significantly differentially expressed.
Bioinformatics and annotations
To determine the biological and functional
properties of the identified proteins, the identified protein
sequences were mapped with Gene Ontology terms. A homology search
was first performed for all the identified sequences with a
localized NCBI BLASTP program against the NCBInr Homo database. The
e-value was set to <1e-5, and the best hit for each query
sequence was considered for Gene Ontology (GO) term matching. The
GO term matching was performed with Blast2GO v4.5 pipeline
(16). In addition, an enrichment
analysis of Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways
was performed. To identify candidate biomarkers, we employed a
hypergeometric test to perform pathway enrichment. A P<0.05 was
set as the threshold value.
Statistical analysis
Data from at least three independent experiments are
presented as the mean ± standard deviation (SD). Statistical
analyses were performed using Statistical Package for the Social
Sciences (SPSS) software version 19.0 (SPSS, Inc., Chicago, IL,
USA). The differences between the groups were analyzed using the
Students t-test or the one-way analysis of variance (ANOVA). A
P<0.05 was considered to be statistically significant.
Results
miR-543 expression was downregulated
in glioma cell lines and human tissues
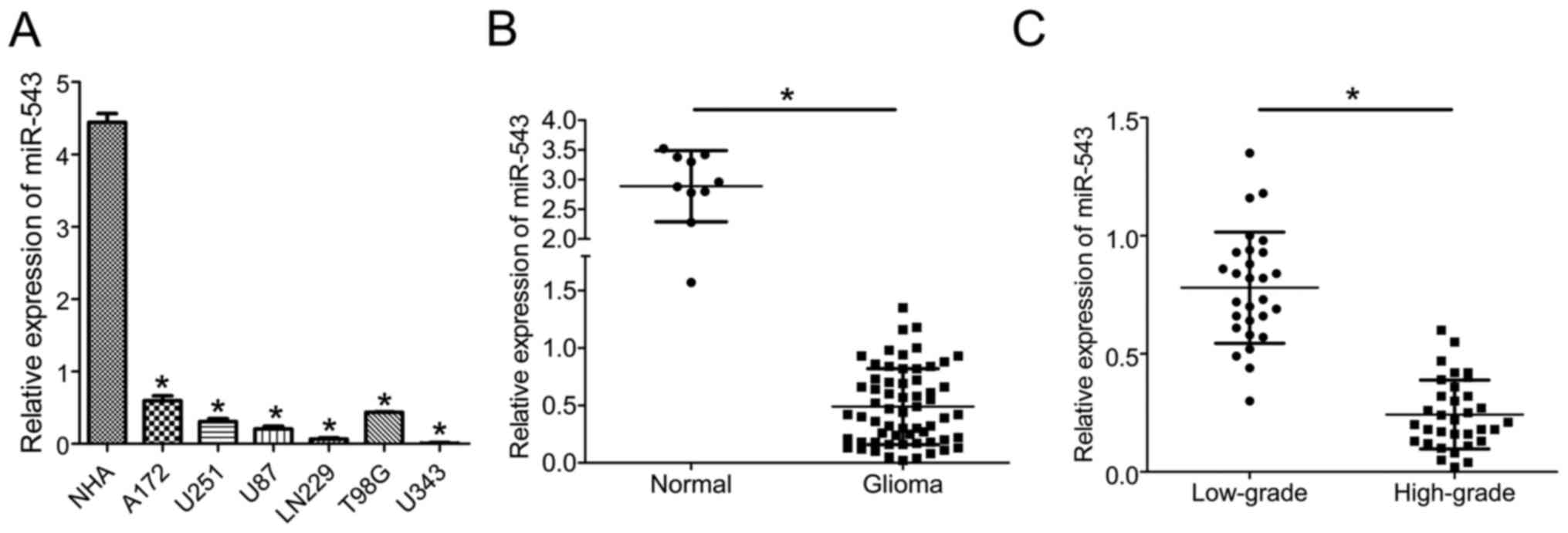
To explore the potential role of miR-543 in glioma
progression, the expression levels of miR-543 in six glioma cell
lines and normal human astrocytes (NHA) were first analyzed by
quantitative real-time PCR (qRT-PCR). The expression level of
miR-543 was significantly reduced in all of the six analyzed glioma
cell lines compared with NHA (P<0.01; Fig. 1A). Furthermore, miR-543 expression
was further investigated in 62 glioma tissues and 10 normal brain
tissues. As shown in Fig. 1B, the
expression of miR-543 was markedly reduced in glioma samples
compared with normal brain tissues (P<0.01). In addition,
miR-543 expression in high-grade (grades III and IV) gliomas was
significantly decreased compared with low-grade (grades I and II)
gliomas (P<0.01; Fig. 1C). Taken
together, these findings indicate that miR-543 might be a hallmark
of glioma and associated with the progression of glioma.
miR-543 inhibited the growth of glioma
cells in vitro
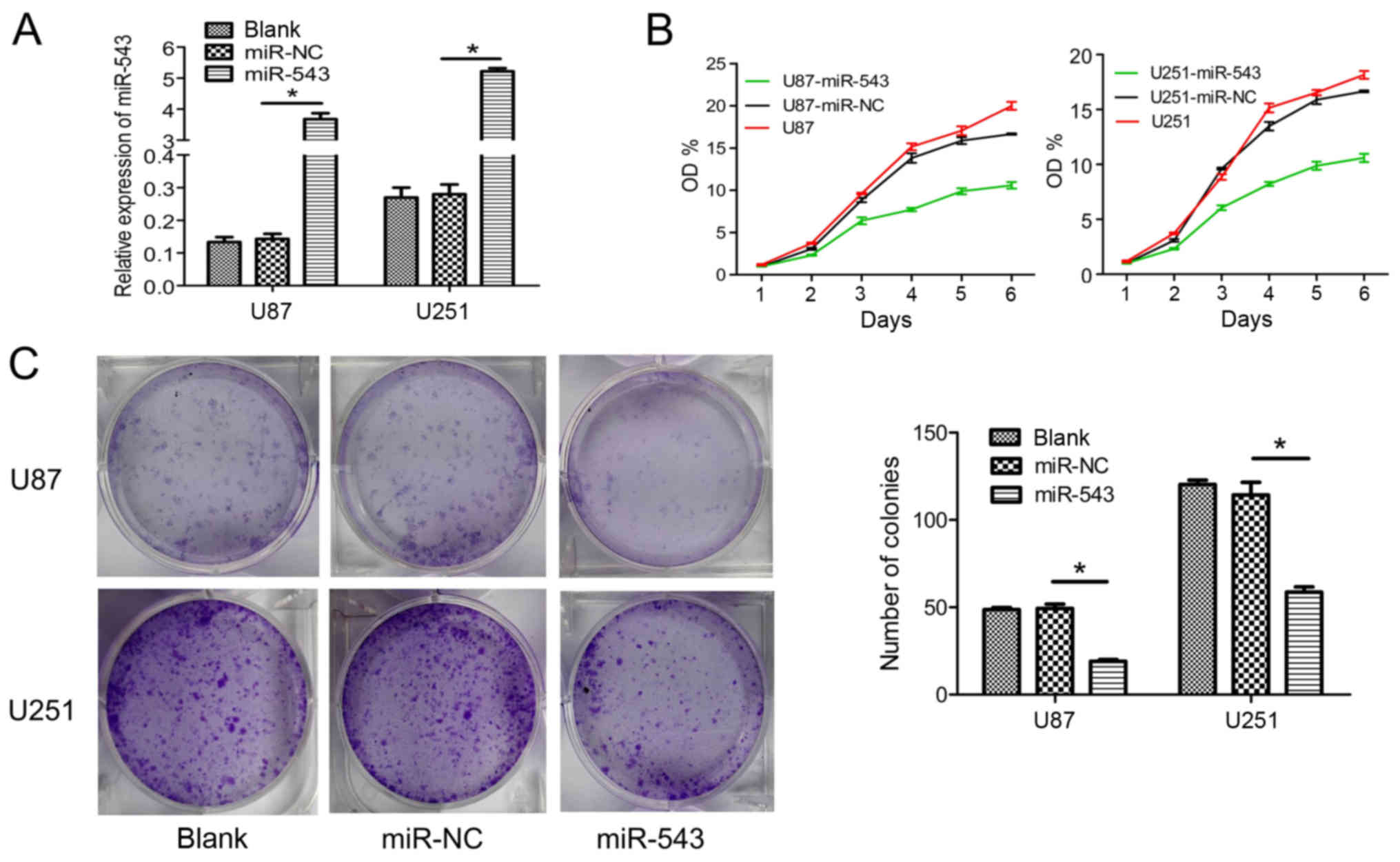
To further examine the role of miR-543 in glioma
cells, U87 and U251 cells were transfected with miR-543 mimic or
miR-NC. For subsequent assays, U87-miR-543 and U251-miR-543
represent the glioma cells transfected with miR-543 mimic. The
negative controls U87-miR-NC and U251-miR-NC represent the glioma
cells transfected with miR-NC. U87 and U251 cells were used as the
blank control. Increased expression of U87-miR-543 and U251-miR-543
were confirmed using qRT-PCR (Fig.
2A). Then, glioma cell growth was detected in vitro.
CCK-8 assays showed that the overexpression of miR-543
significantly inhibited cell proliferation in U87 and U251 cells
compared with control groups (Fig.
2B). Furthermore, the overexpression of miR-543 markedly
inhibited colony formation in glioma cells (Fig. 2C). These results suggested that
miR-543 contributed to the reduced proliferation of glioma
cells.
miR-543 induces apoptosis and inhibits
the cell cycle
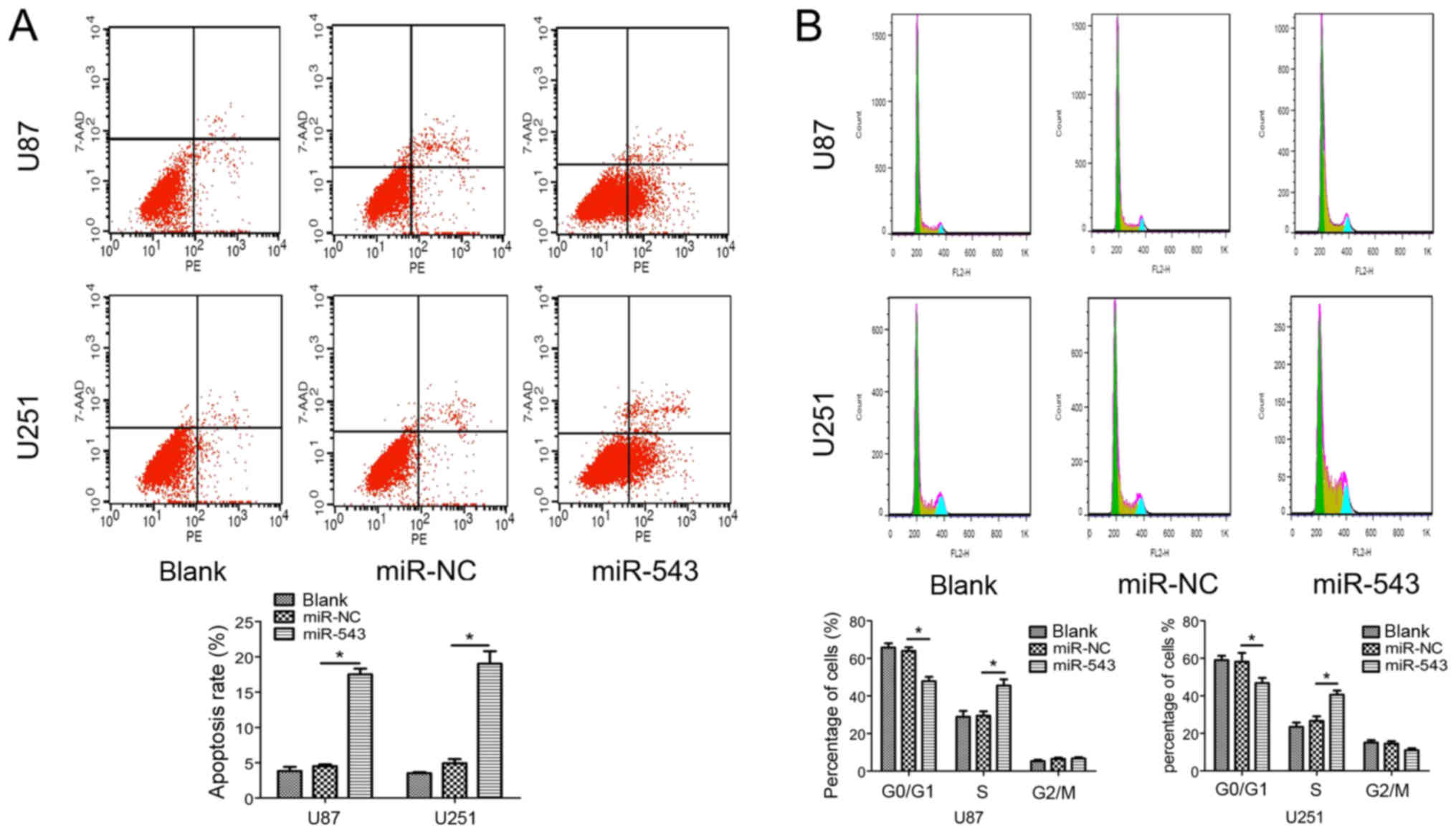
Furthermore, flow cytometry was used to examine the
effects of miR-543 on cell apoptosis and cell cycle distribution.
The results showed that the proportion of apoptotic cells
transfected with miR-543 was significantly increased compared with
control cells (Fig. 3A). In
addition, as shown in Fig. 3B,
miR-543 overexpression increased the percentage of cells in the S
phase (P<0.01). The result showed that miR-543 overexpression in
U87 and U251 cells significantly induced cell cycle arrest at the S
phase. Collectively, these findings indicate that miR-543 acts as a
tumor suppressor in the growth of glioma, likely via modulating
cell cycle progression.
miR-543 regulates the migration and
invasion of glioma cells in vitro
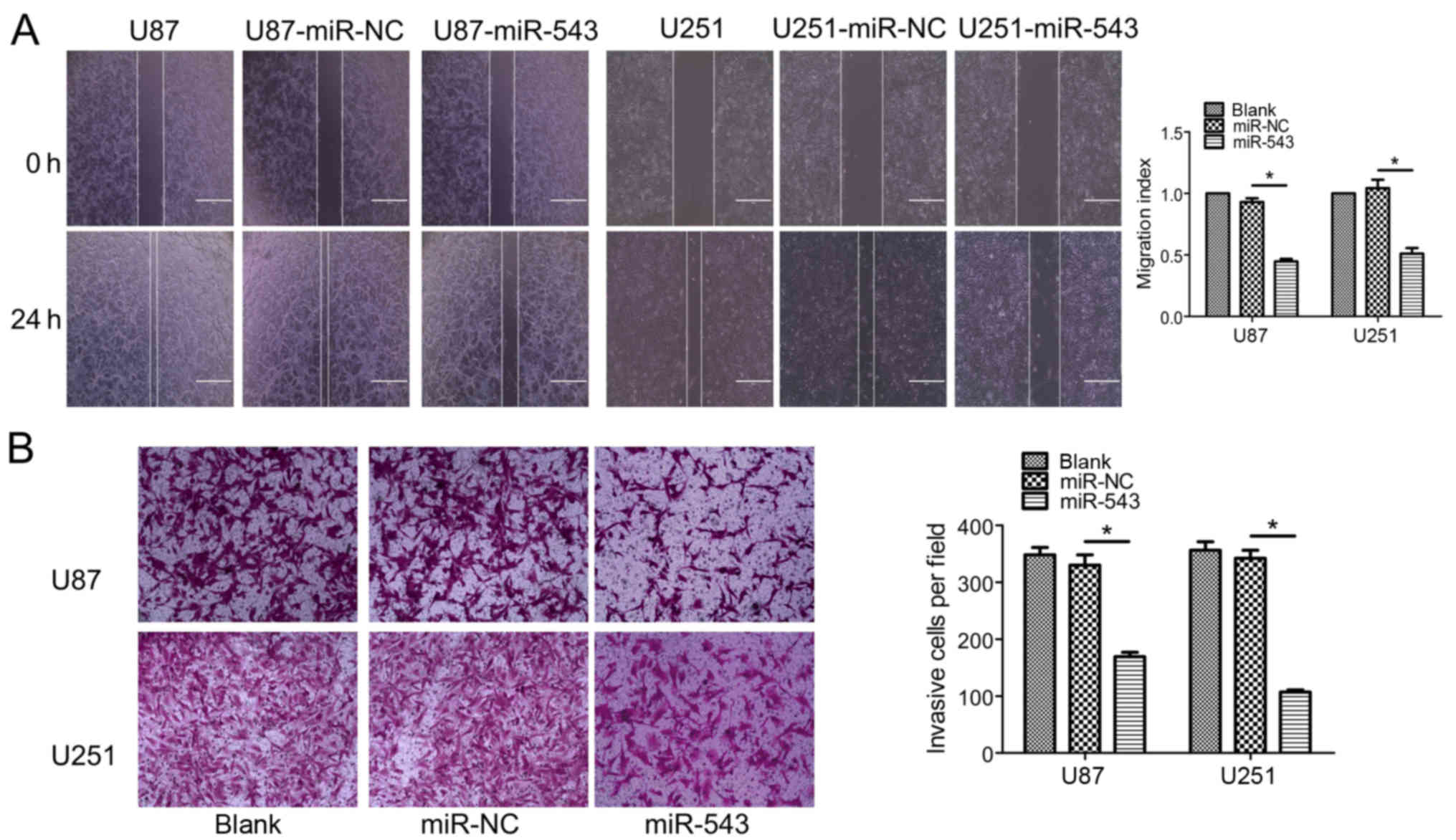
To further clarify the effect of miR-543 on the
migration and invasion ability of U87 and U251 cells, wound healing
and Transwell invasion assays were performed. As shown in Fig. 4A, a significant difference between
the migration of miR-543 overexpression cells and control cells was
noted (P<0.01). The Transwell invasion assay showed that the
overexpression of miR-543 significantly decreased the number of
glioma cells capable of invasion (Fig.
4B; P<0.01). These results suggest that miR-543 inhibits the
migration and invasion of glioma cells in vitro.
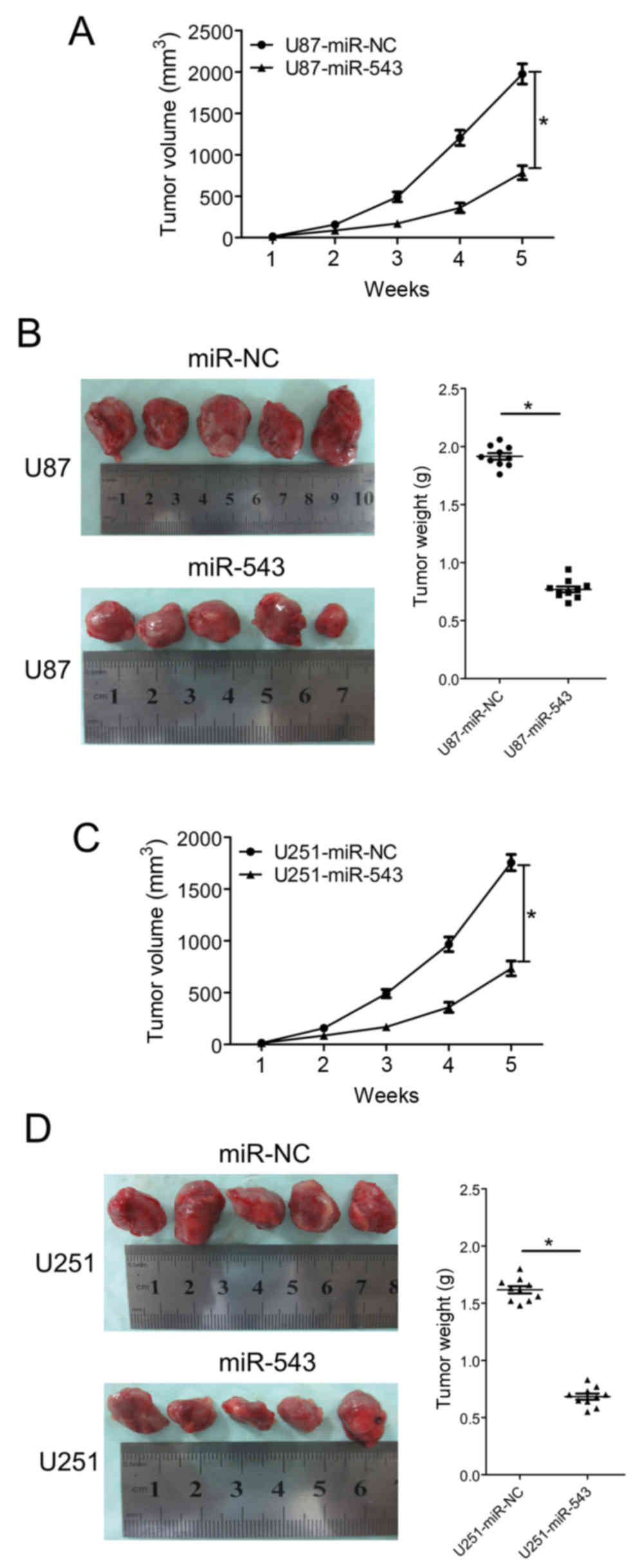
miR-543 suppresses the growth of
glioma xenografts
The in vitro study indicated that miR-543
inhibits glioma cell proliferation and invasion; therefore, the
growth inhibitory effect of miR-543 in vivo was evaluated
using a subcutaneously xenografted glioma model. U87 and U251 cells
were chosen for the in vivo study. The glioma cells stably
expressing miR-543 or miR-NC were subcutaneously inoculated into
nude mice. Tumor volumes were estimated in the subcutaneous model
every week after injection. Five weeks after inoculation, the mice
were sacrificed and tumor tissues were extracted. Tumors formed by
miR-543 overexpressing cells were visibly smaller than those formed
by control cells (Fig. 5B and D).
The average volume and weight of tumors formed by miR-543
overexpressing cells were significantly decreased compared with
control cells (both P<0.01; Fig.
5). The results demonstrated that miR-543 could suppress the
growth of glioma xenografts.
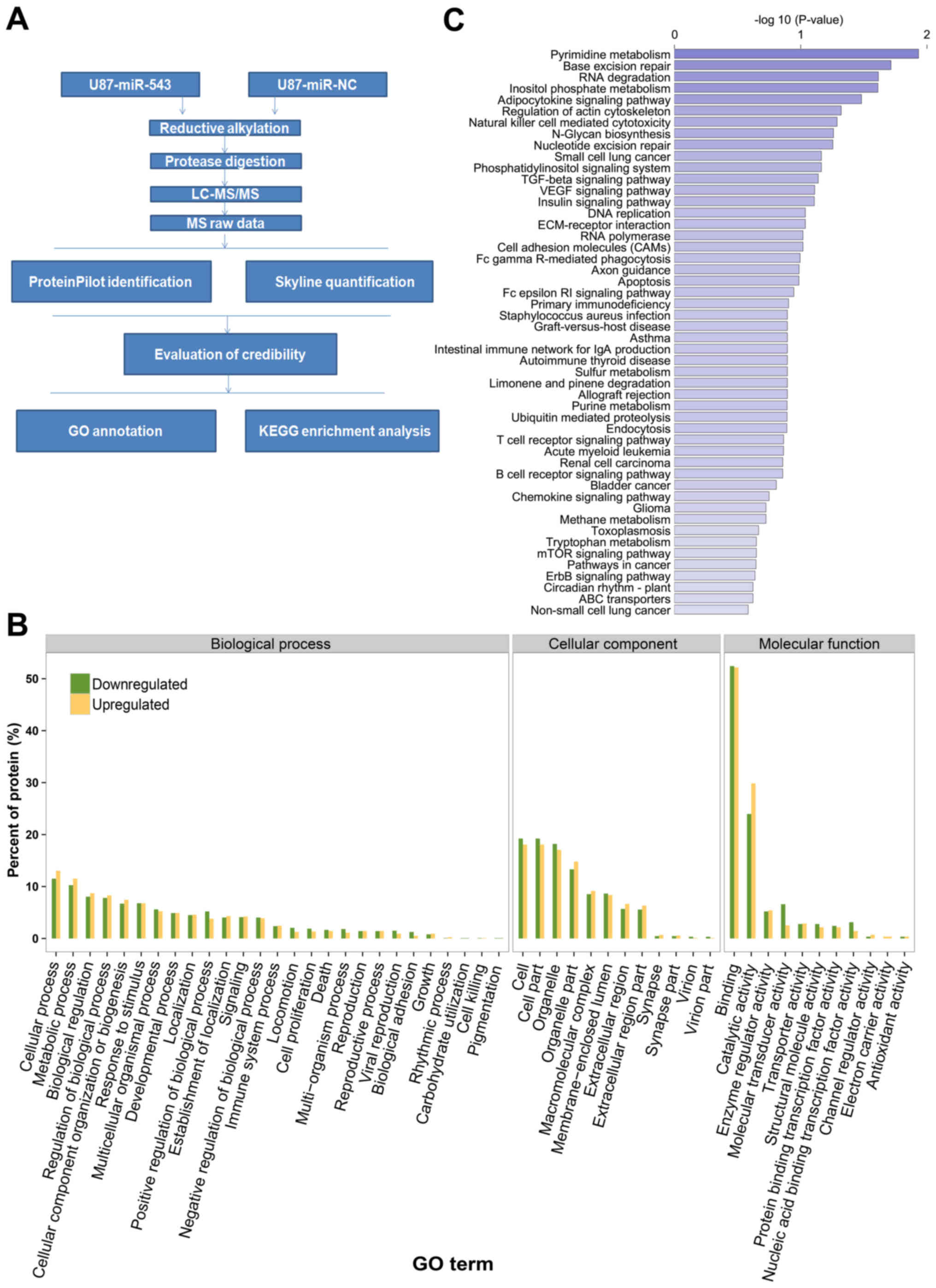
Identification of miR-543-regulated
proteins using a quantitative proteomic approach
Given that miR-543 was downregulated in U87 cells,
we hypothesized that the upregulation of miR-543 alters the
expression of its target genes. A label-free quantitative
proteomics approach was used to identify dysregulated proteins in
U87 glioma cells with or without miR-543 overexpression. The
workflow in the quantitative proteomics approach is presented in
Fig. 6A. A total of 2,446 proteins
were quantified according to the criteria described in Materials
and methods. Among all the quantified proteins, 339 proteins were
differentially expressed after miR-543 overexpression. Of these,
165 were upregulated and 174 were downregulated. The identified
differentially expressed proteins included CKAP5, SYNJ2, POMP,
CSN5, NOL7 and GOLM1. Table I
presents the previously noted 20 upregulated and downregulated
proteins.
 | Table I.Proteins differentially expressed
after miR-543 was overexpressed in U87 glioma cells. |
Table I.
Proteins differentially expressed
after miR-543 was overexpressed in U87 glioma cells.
| Protein | miR-543:miR-NC |
|---|
| Upregulated
(20) |
|
|
Q5LJA5 | 34.14326014 |
|
A0A024RDE8 | 26.47960363 |
|
GSLG1 | 20.15439479 |
|
B7Z410 | 19.53041636 |
|
ARK72 | 16.30160722 |
|
AN32B | 10.8127484 |
|
B4DEX5 | 10.1040579 |
|
CKAP5 | 10.04365302 |
|
Q6IAX1 | 8.927064395 |
|
A0A024QZN8 | 7.504923236 |
|
F8W8R3 | 7.432445004 |
|
SYNJ2 | 7.16088658 |
|
POMP | 6.727857068 |
|
IF4B | 6.688933707 |
|
A0A0C4DFT3 | 6.599762499 |
|
EXOS3 | 6.591721953 |
|
B7ZM73 | 5.992988916 |
|
SSRD | 5.988039064 |
|
Q53HS1 | 5.814460789 |
|
MPPA | 5.634894759 |
| Downregulated
(20) |
|
|
H32 | 0.036087 |
|
B3KVB3 | 0.057414 |
|
CUL4A | 0.05955 |
|
GOLM1 | 0.062021 |
|
NUP53 | 0.074681 |
|
D6R905 | 0.07473 |
|
Q5U091 | 0.079087 |
|
Q53G69 | 0.090318 |
|
A0A024R7M6 | 0.092072 |
|
RUSD2 | 0.096349 |
|
OVCA2 | 0.103314 |
|
RBM15 | 0.121837 |
|
C9K0W8 | 0.133651 |
|
A0A024R7D5 | 0.145933 |
|
ARI3C | 0.1481 |
|
NOL7 | 0.154437 |
|
A8K4H7 | 0.163771 |
|
CE051 | 0.174426 |
|
B4DKM0 | 0.176571 |
|
PP2AA | 0.178847 |
Bioinformatic analysis of
miR-543-regulated proteins
Based on the results of the proteomic analysis, the
miR-543-regulated proteins were classified by their GO cellular
components, biological process and molecular function to achieve an
overview of the GO distribution (Fig.
6B). GO cellular component classification results revealed that
many miR-543-regulated proteins were assigned to cells, cell parts,
organelle and organelle parts. GO biological process analysis
revealed that the miR-543-regulated proteins were involved in
diverse biological processes, such as cellular process, metabolic
process, biological regulation, regulation of biological process
and cellular component organization or biogenesis. In the GO
molecular function category, binding, catalytic activity, enzyme
regulator activity and molecular transducer activity were noted as
the main classifications. As a whole, this bioinformatic
information indicated that miR-543 may act as an important
regulator of different cellular functions across extensive
biological processes.
To gain insights into the function of miR-543, the
enrichment analysis of KEGG pathways among miR-543 regulated
proteins was performed. Fig. 6C
showed the pathways associated with miR-543 regulated proteins with
the bioinformatic analysis. Moreover, according to the P-values
obtained with hypergeometric tests, we identified pathways markedly
enriched and likely involved in miR-543-mediated tumorigenesis. The
enriched pathways included pyrimidine metabolism, base excision
repair, RNA degradation, inositol phosphate metabolism,
adipocytokine signaling pathway and regulation of the actin
cytoskeleton. These results may indicate the involvement of miR-543
in intricate physiological and pathological systems.
Discussion
It was believed that the exploration of the
molecular mechanisms involved in tumor initiation and progression
affords the possibility to identify molecules that may serve as new
targets for the effective therapy of glioma. There is accumulating
evidence demonstrating that miRNAs play crucial roles in
tumorigenic processes, such as cell proliferation, apoptosis,
angiogenesis and invasion (17–19).
miRNAs have been associated with tumor suppressor and oncogenic
activities in glioma (20–22). Therefore, it has been proposed that
miRNAs may act as potential targets for the gene treatment of
glioma.
Some studies have demonstrated the association
between miR-543 and several human cancers. For instance, Yu et
al (12) demonstrated that
miR-543 was dramatically overexpressed in hepatocellular carcinoma
(HCC), and the overexpression of miR-543 promoted the proliferative
and invasive potential of the HCC cell line HepG2, and miR-543
acted as an oncogene by targeting PAQR3 in HCC. Moreover, miR-543
promoted gastric cancer cell proliferation by targeting SIRT1
(23). On the contrary, Fan et
al (13) reported that miR-543
expression was significantly downregulated in tumors from patients
with colorectal cancer (CRC), and miR-543 inhibited the growth and
metastasis of CRC cells in vitro and in vivo by
targeting KRAS, MTA1 and HMGA2. Another similar study found that
miR-543 suppressed endometrial cancer oncogenicity via targeting
FAK and TWIST1 expression (24).
The different roles of miR-543 in different tumors suggested that
miR-543 may exert different functions in different cell types. To
the best of our knowledge, the role of miR-543 in glioma has not
been reported. In the present study, the expression level of
miR-543 was extensively downregulated in glioma cell lines and
tissues. Furthermore, the expression level of miR-543 was
negatively associated with high-grade glioma. A functional study
demonstrated that miR-543 in glioma cells induced apoptosis and
inhibited cell growth, the cell cycle, migration and invasion. In
addition, the in vivo study showed that miR-543 suppressed
the tumorigenicity of glioma cells. According to all these results,
miR-543 may function as a tumor suppressor in glioma and may serve
as a potential biomarker for glioma.
miRNAs generally demonstrate their biological
activities by regulating their target genes through mRNA
degradation and translational repression. In the latter mechanism,
the mRNA of the target remains unchanged. Unlike most studies using
microarray and RNA-Seq to measure dysregulated mRNAs, we used a
proteomic approach to reveal protein changes after miR-543
overexpression (25,26). Studies have demonstrated that one
miRNA can regulate the levels of hundreds of proteins (4); thus, it is valuable and efficient to
conduct a high-throughput protein discovery for further
exploration. Proteomic profiling is the most frequently used
approaches for high-throughput protein detection, and proteomic
pathway analyses are emerging as a pivotal method for comprehending
molecular networks (27,28). Although numerous studies of miRNA
function in tumors have been reported, little information is
available on the landscape of differentially expressed proteins
associated with miRNAs that mediate the progression of tumors. In
the present study, we investigated for the first time the proteomic
profile of U87-miR-543 and the corresponding negative control
U87-miR-NC. The application of the proteomic profile to identify
proteins should be a prerequisite before further exploration to
elucidate the key molecules and molecular pathways involved in
miR-543-mediated tumorigenesis. In the present study, 339 proteins
were identified as dysregulated after miR-543 overexpression. Among
these, 165 were upregulated and 174 were downregulated. Among the
identified differentially expressed proteins, some molecules may be
involved in miR-543-mediated in tumorigenesis. For example, CKAP5
(cytoskeleton-associated protein 5), also known as colonic hepatic
tumor overexpressed gene (ch-TOG), played a critical role in
centrosome clustering in cancer cells (29) and is essential for tumor cell
survival in non-small cell lung cancer (NSCLC) and head and neck
squamous cell carcinoma (HNSCC) (30). POMP, the proteasome maturation
protein, was targeted by miR-101 to impair proteasome assembly and
activity, resulting in the accumulation of p53 and cyclin-dependent
kinase inhibitors, cell cycle arrest and apoptosis (31). As a member of the COP9 signalosome
complex, CSN5 (also known as Jab1) is overexpressed in numerous
human cancers and affects multiple pathways associated with cell
proliferation and apoptosis (32).
Moreover, multiple pathways are significantly enriched and likely
involved in miR-543-mediated tumorigenesis. For instance, members
of the inositol phosphate metabolism pathway regulate cell
proliferation, migration and phosphatidylinositol-3-kinase
(PI3K)/Akt signaling and are frequently dysregulated in cancer
(33). Consequently, the present
study provided deep viewpoint into the key molecules and molecular
pathways involved for further exploration.
Despite the exciting potential as therapeutic
targets in human cancers, there are some scientific challenges. The
major challenge to miRNA-based therapeutics is an effective mode of
delivery of miRNA-targeting agents. Limitations that may be
overcome by delivery include, but are not limited to, poor in
vivo stability, inappropriate biodistribution, disruption and
saturation of endogenous RNA machinery, and untoward side effects
(34). However, miRNA-based
therapies, either restoring or repressing miRNAs expression and
activity, hold great promise.
In conclusion, this study first found that miR-543
expression was downregulated in glioma and that its expression was
negatively associated with advanced tumor stage, and miR-543 may
functioned as a novel tumor suppressor in glioma. Proteomic
analysis shows the landscape of differentially expressed proteins
associated with miR-543, providing comprehensive and deep insight
into the potential molecular mechanisms. Taken together, our data
indicate that miR-543 may be a future therapeutic target in glioma
therapy.
Acknowledgements
The present study was supported by the Foundation
Program of Suzhou Science and Technology Project (SYSD2014092), the
Research and Innovation Project for College Graduates of Jiangsu
Province (KYLX_1263), and the Foundation of Young Member of the
Second Affiliated Hospital of Soochow University (SDFEYQN1409).
References
|
1
|
Ohgaki H: Epidemiology of brain tumors.
Methods Mol Biol. 472:323–342. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Huse JT, Holland E and DeAngelis LM:
Glioblastoma: Molecular analysis and clinical implications. Annu
Rev Med. 64:59–70. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Orang Valinezhad A, Safaralizadeh R and
Kazemzadeh-Bavili M: Mechanisms of miRNA-mediated gene regulation
from common downregulation to mRNA-specific upregulation. Int J
Genomics. 2014:9706072014.PubMed/NCBI
|
|
4
|
Wei Y, Schober A and Weber C: Pathogenic
arterial remodeling: The good and bad of microRNAs. Am J Physiol
Heart Circ Physiol. 304:H1050–H1059. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang S, Hao J, Xie F, Hu X, Liu C, Tong
J, Zhou J, Wu J and Shao C: Downregulation of miR-132 by promoter
methylation contributes to pancreatic cancer development.
Carcinogenesis. 32:1183–1189. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang W, Zhou X and Wei M: MicroRNA-144
suppresses osteosarcoma growth and metastasis by targeting ROCK1
and ROCK2. Oncotarget. 6:10297–10308. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Weiner-Gorzel K, Dempsey E, Milewska M,
McGoldrick A, Toh V, Walsh A, Lindsay S, Gubbins L, Cannon A,
Sharpe D, et al: Overexpression of the microRNA miR-433 promotes
resistance to paclitaxel through the induction of cellular
senescence in ovarian cancer cells. Cancer Med. 4:745–758. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhu Y, Zhao H, Feng L and Xu S:
MicroRNA-217 inhibits cell proliferation and invasion by targeting
Runx2 in human glioma. Am J Transl Res. 8:1482–1491.
2016.PubMed/NCBI
|
|
9
|
Cheng Z, Wang HZ, Li X, Wu Z, Han Y, Li Y,
Chen G, Xie X, Huang Y, Du Z, et al: MicroRNA-184 inhibits cell
proliferation and invasion, and specifically targets TNFAIP2 in
Glioma. J Exp Clin Cancer Res. 34:272015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang H, Lu Y, Luo L, Li W, Liang C, He H
and Ba Y: MicroRNA-195 inhibits the proliferation of human glioma
cells by directly targeting cyclin D1 and cyclin E1. PLoS One.
8:e549322013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Haga CL and Phinney DG: MicroRNAs in the
imprinted DLK1-DIO3 region repress the epithelial-to-mesenchymal
transition by targeting the TWIST1 protein signaling network. J
Biol Chem. 287:42695–42707. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu L, Zhou L, Cheng Y, Sun L, Fan J, Liang
J, Guo M, Liu N and Zhu L: MicroRNA-543 acts as an oncogene by
targeting PAQR3 in hepatocellular carcinoma. Am J Cancer Res.
4:897–906. 2014.PubMed/NCBI
|
|
13
|
Fan C, Lin Y, Mao Y, Huang Z, Liu AY, Ma
H, Yu D, Maitikabili A, Xiao H, Zhang C, et al: MicroRNA-543
suppresses colorectal cancer growth and metastasis by targeting
KRAS, MTA1 and HMGA2. Oncotarget. 7:21825–21839. 2016.PubMed/NCBI
|
|
14
|
Bradford MM: A rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal Biochem. 72:248–254.
1976. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shilov IV, Seymour SL, Patel AA, Loboda A,
Tang WH, Keating SP, Hunter CL, Nuwaysir LM and Schaeffer DA: The
Paragon Algorithm, a next generation search engine that uses
sequence temperature values and feature probabilities to identify
peptides from tandem mass spectra. Mol Cell Proteomics.
6:1638–1655. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Conesa A and Götz S: Blast2GO: A
comprehensive suite for functional analysis in plant genomics. Int
J Plant Genomics. 2008:6198322008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu WG, Shang YL, Cong XR, Bian X and Yuan
Z: MicroRNA-135b promotes proliferation, invasion and migration of
osteosarcoma cells by degrading myocardin. Int J Oncol.
45:2024–2032. 2014.PubMed/NCBI
|
|
18
|
Yu SH, Zhang CL, Dong FS and Zhang YM:
miR-99a suppresses the metastasis of human non-small cell lung
cancer cells by targeting AKT1 signaling pathway. J Cell Biochem.
116:268–276. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tsai MM, Wang CS, Tsai CY, Chen CY, Chi
HC, Tseng YH, Chung PJ, Lin YH, Chung IH, Chen CY, et al:
MicroRNA-196a/−196b promote cell metastasis via negative regulation
of radixin in human gastric cancer. Cancer Lett. 351:222–231. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang QQ, Xu H, Huang MB, Ma LM, Huang QJ,
Yao Q, Zhou H and Qu LH: MicroRNA-195 plays a tumor-suppressor role
in human glioblastoma cells by targeting signaling pathways
involved in cellular proliferation and invasion. Neuro Oncol.
14:278–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gao X and Jin W: The emerging role of
tumor-suppressive microRNA-218 in targeting glioblastoma stemness.
Cancer Lett. 353:25–31. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Che S, Sun T, Wang J, Jiao Y, Wang C, Meng
Q, Qi W and Yan Z: miR-30 overexpression promotes glioma stem cells
by regulating Jak/STAT3 signaling pathway. Tumour Biol.
36:6805–6811. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li J, Dong G, Wang B, Gao W and Yang Q:
miR-543 promotes gastric cancer cell proliferation by targeting
SIRT1. Biochem Biophys Res Commun. 469:15–21. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bing L, Hong C, Li-Xin S and Wei G:
MicroRNA-543 suppresses endometrial cancer oncogenicity via
targeting FAK and TWIST1 expression. Arch Gynecol Obstet.
290:533–541. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang W, Luo J, Sheng W, Xue J, Li M, Ji J,
Liu P, Zhang X, Cao J and Zhang S: Proteomic profiling of
radiation-induced skin fibrosis in rats: Targeting the
ubiquitin-proteasome system. Int J Radiat Oncol Biol Phys.
95:751–760. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang S, Wang W, Gu Q, Xue J, Cao H, Tang
Y, Xu X, Cao J, Zhou J, Wu J, et al: Protein and miRNA profiling of
radiation-induced skin injury in rats: The protective role of
peroxiredoxin-6 against ionizing radiation. Free Radic Biol Med.
69:96–107. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yan W and Chen SS: Mass spectrometry-based
quantitative proteomic profiling. Brief Funct Genomics Proteomics.
4:27–38. 2005. View Article : Google Scholar
|
|
28
|
Hanash S: Disease proteomics. Nature.
422:226–232. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fielding AB, Lim S, Montgomery K, Dobreva
I and Dedhar S: A critical role of integrin-linked kinase, ch-TOG
and TACC3 in centrosome clustering in cancer cells. Oncogene.
30:521–534. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Martens-de Kemp SR, Nagel R, Stigter-van
Walsum M, van der Meulen IH, van Beusechem VW, Braakhuis BJ and
Brakenhoff RH: Functional genetic screens identify genes essential
for tumor cell survival in head and neck and lung cancer. Clin
Cancer Res. 19:1994–2003. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang X, Schulz R, Edmunds S, Krüger E,
Markert E, Gaedcke J, Cormet-Boyaka E, Ghadimi M, Beissbarth T,
Levine AJ, et al: MicroRNA-101 suppresses tumor cell proliferation
by acting as an endogenous proteasome inhibitor via targeting the
proteasome assembly factor POMP. Mol Cell. 59:243–257. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sang MM, Du WQ, Zhang RY, Zheng JN and Pei
DS: Suppression of CSN5 promotes the apoptosis of gastric cancer
cells through regulating p53-related apoptotic pathways. Bioorg Med
Chem Lett. 25:2897–2901. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tan J, Yu CY, Wang ZH, Chen HY, Guan J,
Chen YX and Fang JY: Genetic variants in the inositol phosphate
metabolism pathway and risk of different types of cancer. Sci Rep.
5:84732015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang Y, Wang Z and Gemeinhart RA:
Progress in microRNA delivery. J Control Release. 172:962–974.
2013. View Article : Google Scholar : PubMed/NCBI
|