Introduction
Chemotherapy is one of the major modalities of
cancer treatment; however, the effect of chemotherapy is often
diminished by drug resistance (1).
The occurrence of chemoresistance is a serious problem during the
treatment of local and disseminated disease (2). Based on tumor response to the initial
therapy, cancer resistance can be seperated into two categories,
intrinsic or acquired (1,3). Intrinsic drug resistance concerns
patients who express pre-existing, resistance-mediating factors
before receiving chemotherapy. Acquired drug resistance is related
to patients who were initially sensitive to the chemotherapy but
who developed resistance during treatment (1,3).
Numerous advanced techniques have increased our ability to identify
novel genes and signaling pathways that are related to tumor
responsiveness to a particular drug treatment (1). Various mechanisms of overcoming drug
resistance have been developed to treat cancer (4).
Drug resistance is caused by a variety of factors,
such as cancer stem cells, drug efflux, drug inactivation,
alterations in drug targets, DNA damage repair, and deregulated
apoptosis (1). Among these factors,
drug efflux-mediated resistance due to overexpression of ABC
transporters is the most frequent occurrence (4). ABC transport molecules, which are
expressed on the cellular membrane and on the membranes of small
vesicles, have important physiologic functions and affect the
transport of chemotherapeutic reagents in humans (4,5).
Currently, there are 48 transporters in the ABC family, and 13 of
the ABC transporters are related to tumor drug resistance,
including P-glycoprotein (P-gp, MDR1/ABCB1), multidrug resistance
proteins (MRPs/ABCCs) and breast cancer resistance protein
(BCRP/ABCG2) (4,5). P-gp (MDR1) normally protects several
organs from toxic compounds, inhibiting them from entering the
cytosol and discharging them to the cell exterior (5). However, in patients with tumors, P-gp
(MDR1) ejects chemotherapeutic agents out of the cells, decreasing
their efficacy. P-gp (MDR1) is overexpressed in cancer cells and is
one of the main obstacles to successful chemotherapy treatments for
cancer (5).
Apigenin (4′,5,7-trihydroxyflavone) is a flavone
found in various plants. Apigenin present in common fruits and
vegetables, such as apples, grapes, cherries, celery, parsley,
Chinese cabbage, bell peppers, wine and tea. Similar to most
flavones, apigenin has anti-inflammatory (6), antioxidant (7), anti-telomerase (8) and antidepressant (9) properties. The most important
characteristic of apigenin is its anticancer activity. Previous
studies indicate that apigenin induces a decreased risk of certain
cancers, particularly breast, digestive tract, skin, prostate,
lung, and ovary cancers and certain hematological malignancies
(10–15). In in vivo models, apigenin
inhibited prostate cancer progression in a transgenic
adenocarcinoma of the mouse prostate (TRAMP) model (16). Apigenin induced attenuation of tumor
growth in U937 xenografts (17).
Apigenin strongly inhibited tumor growth in nude mice (18). In in vitro models, apigenin
inhibited cell growth and induced apoptosis in cancer cell lines
including breast (19), lung
(20), colon (21,22),
prostate (23), leukemia (24), and pancreatic (25) cells. These reports propose that
apigenin could be used as a chemopreventive and/or chemotherapeutic
agent for cancer.
STAT3 is a transcription factor that modulates
development and physiology, and it is abnormally expressed in
pathological situations such as cancer (26). Upon ligand binding, STAT3 is
activated, resulting in dimerization, translocation to the nucleus,
binding to DNA response elements and the induction of transcription
of genes implicated in cell survival and proliferation. Cancer
cells expressing constitutively activated STAT3 are more resistant
to apoptosis and chemotherapy (26).
In this study, we investigated whether apigenin
overcomes drug resistance and the mechanism of action. For this
purpose, we tested the effects of apigenin on proliferation and
apoptosis of MCF-7 cells and adriamycin-resistant MCF-7/ADR cells.
We analyzed whether apigenin recovers cells from adriamycin
resistance, resulting in downregulation of P-gp (MDR1) expression.
We also investigated whether apigenin inhibits the STAT3 signaling
pathway, leading to the suppression of breast cancer development
and drug resistance. We report here that apigenin overcomes drug
resistance, thus it may help in cancer treatment.
Materials and methods
Compounds
Apigenin (4′,5,7-trihydroxyflavone), HIF-1α
(hypoxia-inducible factor 1-α) inhibitor (EF-24),
7-aminoactinomycin D (7-AAD), rhodamine 123 and nicardipine were
purchased from Sigma Chemical Co. (St. Louis, MO, USA). These
compounds were dissolved in dimethyl sulfoxide (DMSO) or ethanol,
and the final concentration of DMSO or ethanol in the controls and
in each sample did not exceed 0.1%. The STAT3 inhibitor (S3I-201)
was purchased from Calbiochem (San Diego, CA, USA). JAK (Janus
kinase) inhibitor I was obtained from Santa Cruz Biotechnology,
Inc. (Santa Cruz, CA, USA). Annexin V, Alexa Fluor® 488
Conjugate was purchased from Thermo Fisher Scientific Korea (Seoul,
Korea). An EZ-western chemiluminescent detection kit was obtained
from Daeillab Service Co. (Seoul, Korea).
Cell culture
MCF-7 (ATCC, American Type Culture Collection,
Manassas, VA, USA) and MCF-7/ADR (a gift from Professor Hwa Jeong
Lee, Ewha Womans University, Seoul, Korea) cells were cultured in
Dulbecco's modified Eagle's medium (DMEM) and RPMI-1640 medium,
respectively, containing 50 U/ml penicillin, 50 mg/ml streptomycin
and 10% fetal bovine serum (FBS; Welgene, Daegu, Korea) at 37°C in
an atmosphere of 5% CO2.
Antibodies
Primary antibodies directed against cleaved
caspase-8, poly(ADP-ribose) polymerase (PARP) and P-gp (MDR1) were
obtained from Cell Signaling Technology, Inc. (Danvers, MA, USA).
Primary antibodies directed against STAT3 and phospho-STAT3
(Tyr705) were purchased from Upstate-Millipore (Billerica, MA,
USA). The anti-tubulin antibody came from Sigma Chemical Co.
Horseradish peroxidase (HRP)-conjugated secondary antibodies (mouse
and rabbit) were obtained from Calbiochem, and anti-goat secondary
antibody was from Jackson ImmunoResearch (West Grove, PA, USA).
MTT assay
MCF-7 and MCF-7/ADR cells were plated in 96-well
culture plates at a density of 3×103 cells/well and
incubated for 24 h at 37°C. Then, they were treated with adriamycin
(0–20 µg/ml), apigenin (0–100 µM), STAT3 inhibitor (0–500 µM), JAK
inhibitor I (0–10 µM), or HIF-1α inhibitor (0–100 µM) for 48 or 72
h. After incubation, MTT reagents (0.5 mg/ml) were inserted to each
well, and the plates were incubated in a humidified incubator at
37°C for 2 h. At the end of the incubation period, the medium was
removed, the resulting formazan was dissolved in DMSO, and the
optical density was determined at 570 nm using an ELISA plate
reader.
Clonogenic survival assay
For the colony formation assay, MCF-7 and MCF-7/ADR
cells were plated in 6-well culture plates at a density of
5×102 cells/well. After 24 h, the cells were treated
with different concentrations of apigenin (0–80 µM) or vehicle and
maintained for 10 days at 37°C. Medium was changed every 3 days.
Finally, the plates were stained with hematoxylin, and colony
numbers were counted.
Cell cycle analyses by flow
cytometry
MCF-7 and MCF-7/ADR cells treated with apigenin
(0–80 µM) were harvested with 0.25% trypsin and washed once with
phosphate-buffered saline (PBS). After the cells were centrifuged,
they were fixed in cold 95% ethanol with 0.5% Tween-20 and stored
at −20°C for at least 30 min. The cells were incubated in 50 µg/ml
of propidium iodide (PI) (including 1% sodium citrate and 50 µg/ml
RNase A) at room temperature in the dark for 30 min. The analysis
of apoptotic cells was performed with a FACScan flow cytometer
(Becton-Dickinson, Mountain View, CA, USA), and the data were
analyzed using CellQuest software.
Annexin V/7AAD apoptosis assay
Apoptotic cell death was measured by an Annexin
V-FITC and 7-AAD assay. Cells were stained with Annexin V-FITC and
7-AAD for 30 min at room temperature in the dark. The apoptotic
cell population was analyzed with a FACSCalibur™ flow cytometer by
measuring the signal in the FL-1 and FL-3 channels.
Western blot analysis
Harvested cells were lysed in modified RIPA buffer
containing 150 mM NaCl, 1% NP-40, 0.5% deoxycholate, 0.1% SDS, 50
mM Tris (pH 8.0), 1 mM EDTA, 1 mM phenylmethylsulfonyl fluoride
(PMSF), 1 mM NaF, 1 mM Na3VO4 and a protease
inhibitor mixture]. The lysates were cleared by centrifugation at
13,000 rpm for 15 min at 4°C. The supernatants were collected and
stored at −70°C until use. The protein concentration was quantified
using a Bio-Rad Bradford protein assay (Bio-Rad, Hercules, CA,
USA). Total proteins (30 µg) were electrophoresed using 6–15%
reducing SDS-polyacrylamide gels and transferred to nitrocellulose
membranes. After blocking with 0.1% Tween-20 in PBS (PBST)
containing 1% skim milk and 1% BSA for 1 h, the membranes were
incubated overnight at 4°C with the indicated primary antibodies.
After washing in 1X PBST for 15 min (3 times × 5 min), the
membranes were incubated with diluted enzyme-linked secondary
antibodies. After washing in 1X PBST for 1 h (4 times × 15 min),
the protein bands were detected using the EZ-western
chemiluminescent detection kit and visualized by exposing the
membranes to X-ray film.
Immunocytochemistry
MCF-7 and MCF-7/ADR cells (2×104
cells/well) were plated in 8-well chamber slides, incubated
overnight at 37°C and treated with apigenin (80 µM) for 24 h. The
cells were fixed with 4% paraformaldehyde for 30 min and treated
with 3% hydrogen peroxide (H2O2) in methanol
for 20 min. The cells were washed with PBS, blocked with 5% BSA in
PBS for 1 h and incubated with an anti-STAT3 primary antibody
(1:100 dilution) for 24 h at 4°C. After the cells were rinsed with
PBS, they were incubated with anti-rabbit biotin-conjugated
secondary antibody for 1 h at room temperature. Finally, the cells
were treated with Vectastain ABC reagent (Vector Laboratories, Inc.
Burlingame, CA, USA) for 30 min at 4°C and stained with
diaminobenzidine tetra chloride (DAB) and hematoxylin. The cells
were mounted with mounting medium and subsequently analyzed by
microscopy.
RNA extraction and reverse
transcription-polymerase chain reaction (RT-PCR)
RNA was isolated from cells using easy-blue RNA
extraction kit (iNtRON Biotech, Sungnam, Korea) according to the
manufacturer's instructions. Isolated RNA content was measured
using the NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies
Inc., Wilmington, DE, USA). Total cellular RNA (2 µg) from each
sample was reverse transcribed using cDNA synthesis kit (Takara,
Otsu, Shiga, Japan). PCR was conducted in a 20-µl reaction mixture
consisting of DNA template, 10 pM of each gene-specific primer, 10X
Taq buffer, 2.5 mM dNTP mixture, and 1 U of Taq DNA polymerase
(Takara). PCR was performed using the specific primer and primer
sequences are shown in Table I.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Type | Primer name |
| Sequences |
|---|
| Human | MDR1 | Forward: | 5′-CCC ATC ATT GCA
ATA GCA GG-3′ |
|
|
| Reverse: | 5′-GTT CAA ACT TCT
GCT CCT GA-3′ |
| Human | MRP1 | Forward: | 5′-GCC GAA GGA GAG
ATC ATC-3′ |
|
|
| Reverse: | 5′-AAC CCG AAA ACA
AAA CAG G-3′ |
| Human | MRP3 | Forward: | 5′-GTG GGG ATC AGA
CAG AGA T-3′ |
|
|
| Reverse: | 5′-TAT CGG CAT CAC
TGT AAA CA-3′ |
| Human | MRP5 | Forward: | 5′-CAG CCA GTC CTC
ACA TCA-3′ |
|
|
| Reverse: | 5′-GAA GCC CTC TTG
TCT TTT TT-3′ |
| Human | BCRP | Forward: | 5′-TGA CGG TGA GAG
AAA ACT TAC-3′ |
|
|
| Reverse: | 5′-TGC CAC TTT ATC
CAG ACC T-3′ |
| Human | GAPDH | Forward: | 5′-CGT CTT CAC CAC
CAT GGA GA-3′ |
|
|
| Reverse: | 5′-CGG CCA TCA CGC
CAC AGT TT-3′ |
Measurement of VEGF and MMP-9 secreted
from MCF-7 and MCF-7/ADR cells by ELISA
The levels of VEGF (vascular endothelial growth
factor) and MMP-9 (matrix metalloproteinases-9) were determined by
sandwich ELISA using the BD Pharmingen human ELISA set (Pharmingen,
San Diego, CA, USA). Briefly, plates were coated with capture
antibody in ELISA coating buffer (Sigma) and incubated overnight at
4°C. Plates were washed with PBS-Tween-20 (0.05%) and subsequently
blocked (10% FBS in PBS) for 1 h at 20°C. Serial dilutions of
standard antigen or sample in dilution buffer (10% FBS in PBS) were
added to the plates and plates were incubated for 2 h at 20°C.
After washing, biotin-conjugated anti-mouse IgE and
streptavidin-conjugated horseradish peroxidase (SAv-HRP) were added
to the plates and plates were incubated for 1 h at 20°C. Finally,
tetramethylbenzidine (TMB) substrate solution was added to the
plates and after 15 min incubation in the dark, 2 N
H2SO4 solution was added to stop the
reaction. Optical densities were measured at 450 nm on an automated
ELISA reader (Versa Max, Molecular Devices, Sunnyvale, CA,
USA).
Rhodamine 123 efflux assay
MCF-7 and MCF-7/ADR cells were treated with apigenin
(0–80 µM) or nicardipine (positive control, 12.5 µM) for 24 h and
incubated for another 1 h with 1 µg/ml of rhodamine 123.
Accumulation of rhodamine 123 in the cells was analyzed by flow
cytometry.
Statistical analysis
All experiment results were expressed as the means ±
standard deviations (SD) of at least three separate tests.
Student's t-test was used for single variable comparisons, and a
P-value <0.05 was considered statistically significant.
Statistical analyses were performed using PRISM software (GraphPad
Software Inc., La Jolla, CA, USA).
Results
Apigenin suppresses the growth of
MCF-7/ADR cells
Before investigating the effect of apigenin on the
growth of adria- mycin-resistant cells, we determined whether
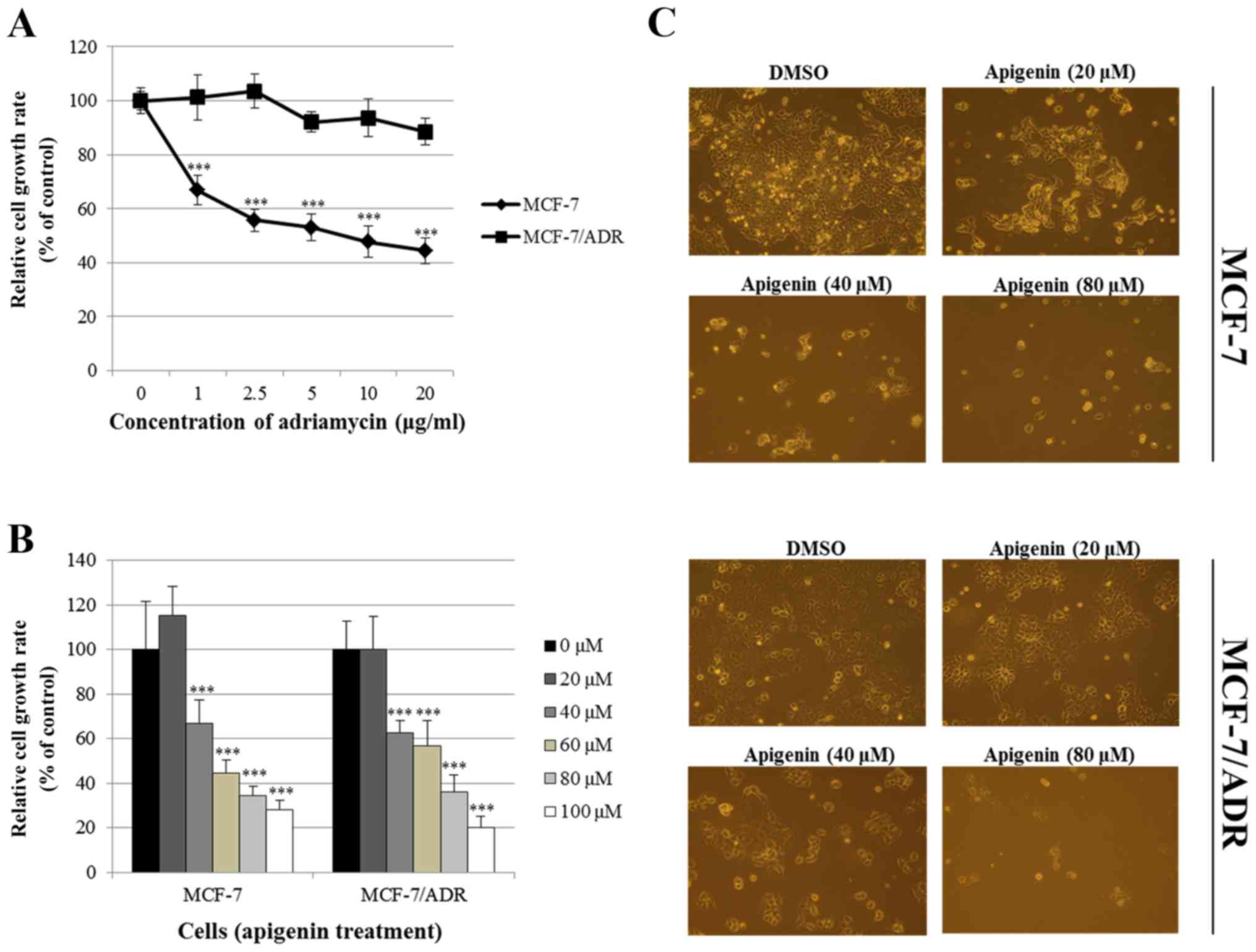
MCF-7/ADR cells display resistance to adriamycin. Fig. 1A demonstrates that adriamycin
decreased the growth rate of MCF-7 cells, while it did not affect
the growth rate of MCF-7/ADR cells, confirming that the MCF-7/ADR
cell line is adriamycin-resistant. Fig.
1B indicates that apigenin significantly suppressed the
proliferation of both MCF-7 and MCF-7/ADR cells in a dose-dependent
manner after 72 h of treatment. This growth inhibition induced by
apigenin was confirmed by microscopic observation (Fig. 1C). Apigenin also induced
morphological changes in these cells; the shape of the cells became
round and shrunken. The changes were common both to MCF-7 and
MCF-7/ADR (Fig. 1C). These results
indicate that apigenin suppresses the growth of
adriamycin-resistant cells.
Apigenin inhibits clonogenic survival
of MCF-7/ADR cells
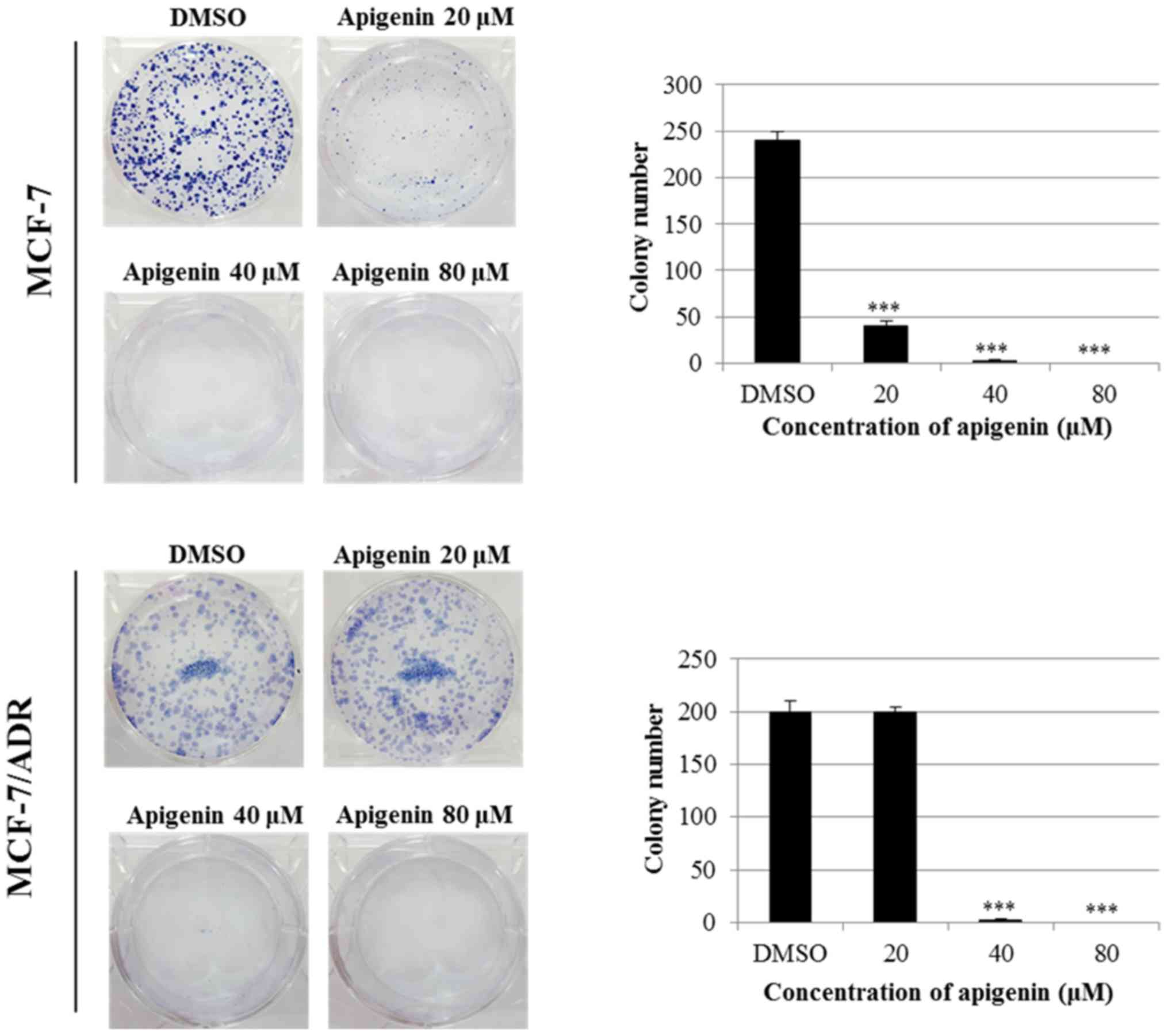
Next, we investigated the effect of apigenin on the
clonogenic survival of MCF-7/ADR cells using a colony formation
assay. As shown in Fig. 2, apigenin
strongly inhibited colony formation in both MCF-7 and MCF-7/ADR
cells in a dose-dependent manner. The effect of apigenin was
stronger in MCF-7 cells than in MCF-7/ADR cells, as evidenced by
the relative absence of colony formation in MCF-7 cells and the
presence of colonies in MCF-7/ADR cells with 20 µM treatment. These
results indicate that MCF-7/ADR cells are drug-resistant and that
apigenin overcomes this resistance.
Apigenin induces apoptosis in
MCF-7/ADR cells
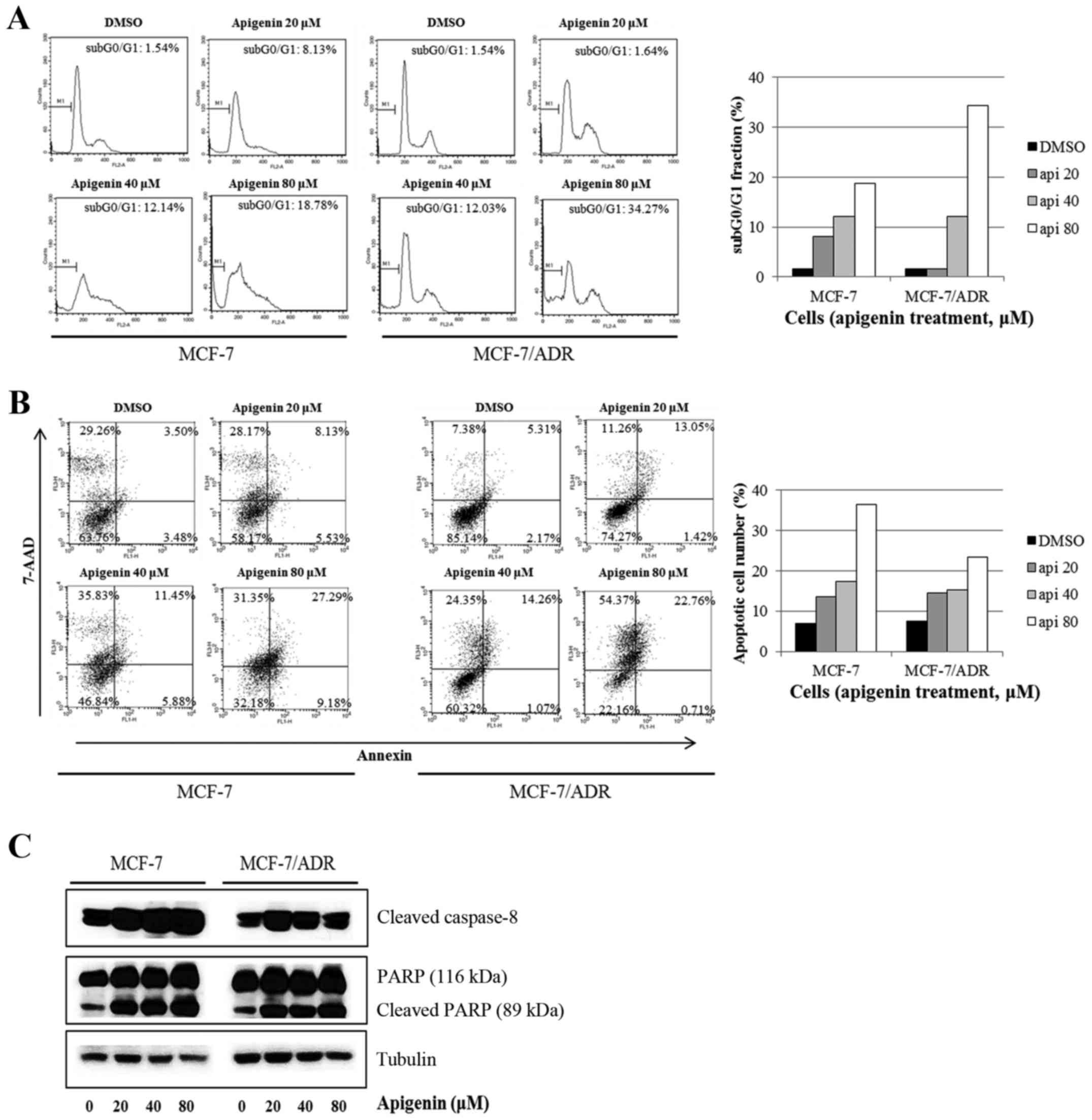
To study whether apigenin blocks cell proliferation
by promoting changes in cell cycle progression, the effect of
apigenin on the cell cycle profile was assessed in MCF-7 and
MCF-7/ADR cells. For this purpose, cells were treated with apigenin
(0–80 µM) for 72 h and then analyzed for cell cycle stage by flow
cytometry. The results demonstrated that apigenin induced an
increase in the sub-G0/G1 apoptotic
population in both cells (Fig. 3A).
In the Annexin V assays, we found that apigenin increased the
number of apoptotic cells in both cell lines (Fig. 3B). Moreover, we observed that
apigenin induces not only apoptosis but necrosis. Consistent with
these results, we also found that apigenin altered the apoptotic
protein machineries; apigenin upregulated the levels of cleaved
caspase-8 and PARP in both cell lines (Fig. 3C). Therefore, our data indicate that
apigenin induces apoptosis in chemotherapy-resistant breast cancer
cells.
Apigenin downregulates MDR1 expression
in MCF-7/ADR cells
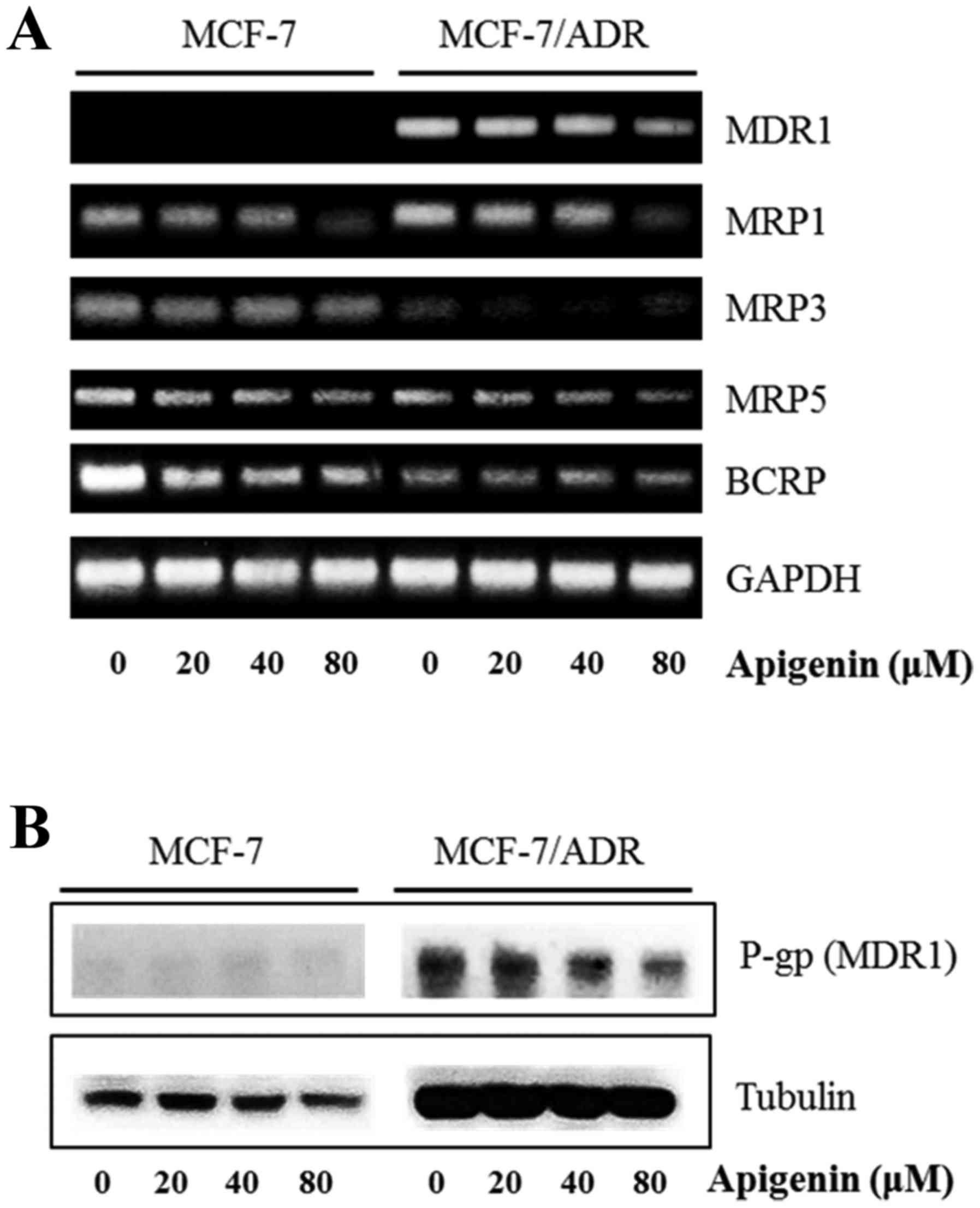
Because P-gp (MDR1), multidrug resistance-associated
proteins (MRP), and breast cancer resistance protein (BCRP), are
important contributors to drug resistance, we checked whether
apigenin regulates their expression. RT-PCR demonstrated that
apigenin reduced the mRNA expression of MDR1, MRP1, MRP3, MRP5 and
BCRP (Fig. 4). Western blot
analysis indicated that P-gp (MDR1) was not induced in MCF-7 cells
in the presence or absence of apigenin, while apigenin decreased
the protein expression of P-gp (MDR1) at 80 µM in MCF-7/ADR cells
(Fig. 4). These results indicate
that apigenin inhibits drug resistance by reducing the expression
of drug resistance proteins in MCF-7/ADR cells.
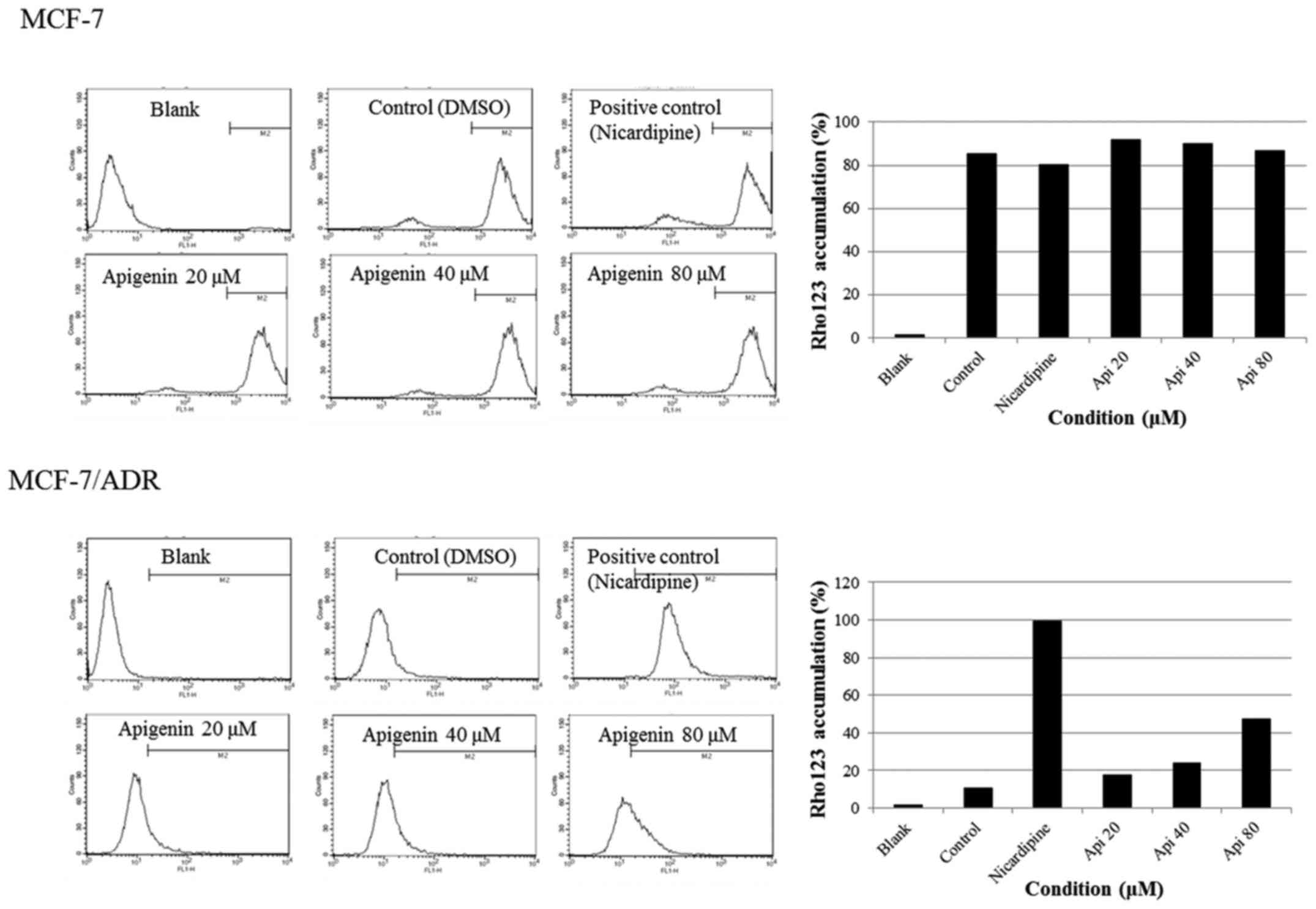
Apigenin recovers accumulation of
rhodamine 123 in MCF-7/ADR cells
Because drug resistance is associated with drug
efflux by the cells, we performed a rhodamine 123 assay to
investigate whether apigenin recovers drug accumulation in
MCF-7/ADR cells. As expected, rhodamine 123 accumulation was high
in the MCF-7 cell line, which is a drug-sensitive cell line
(Fig. 5). In contrast, rhodamine
123 was ejected from the drug-resistant MCF-7/ADR cells, but
apigenin recovered rhodamine 123 accumulation (Fig. 5). These results indicate that
apigenin inhibits drug resistance by suppressing drug efflux in
MCF-7/ADR cells.
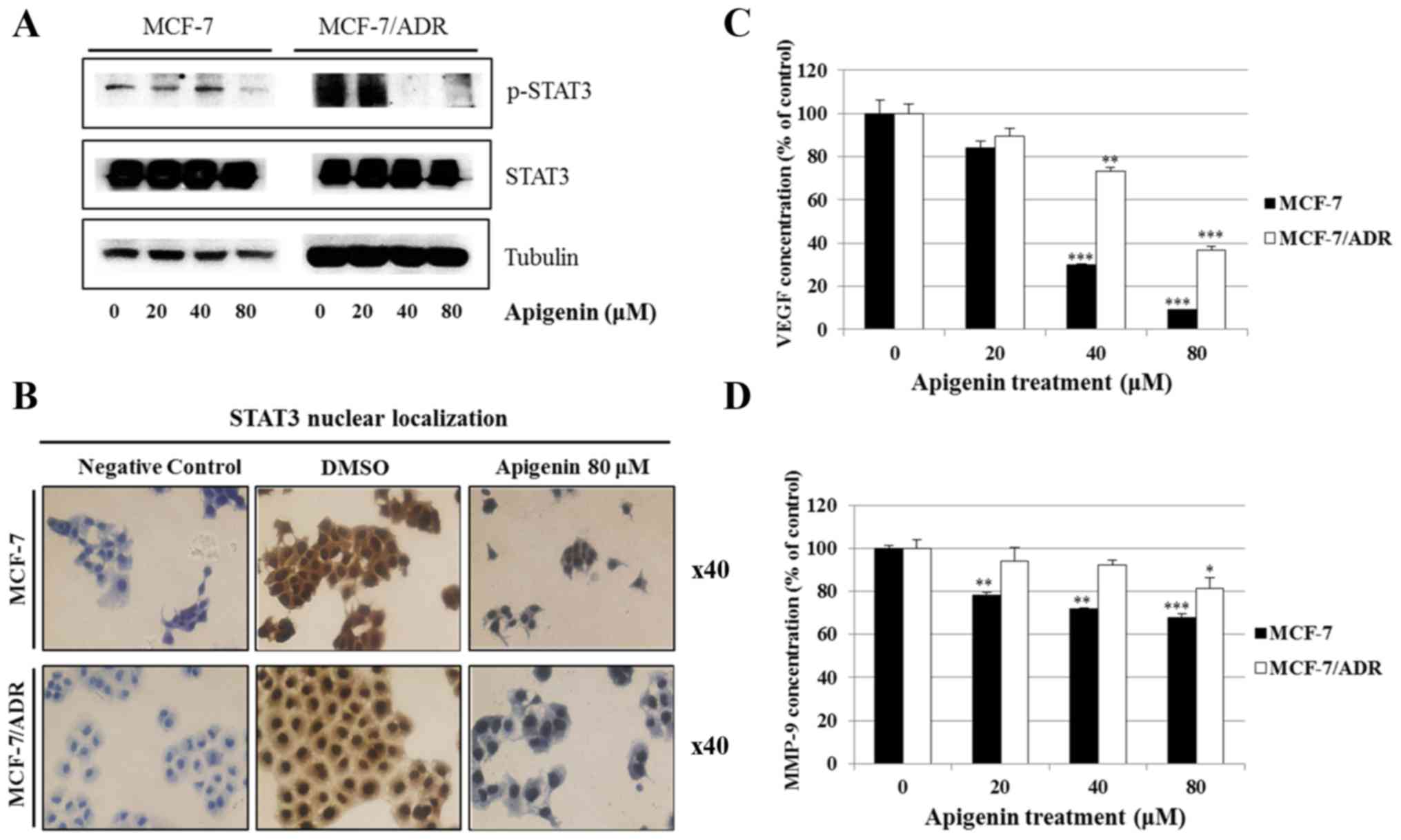
Apigenin inhibits the STAT3 signaling
pathway, resulting in suppression of drug resistance
Since STAT3 is related to oncogenic signaling, and
it is known that STAT3 deactivation reverses chemotherapeutic
resistance (27), we investigated
whether apigenin inhibits the STAT3 signaling pathway in MCF-7/ADR
cells. We found that apigenin reduced the expression of p-STAT3 in
both MCF-7 and MCF-7/ADR cells (Fig.
6A). Immunocytochemical study demonstrated that apigenin
decreased the nuclear localization of STAT3 in both MCF-7 and
MCF-7/ADR cells (Fig. 6B). ELISA
demonstrated that apigenin decreased the production of
intracellular VEGF and MMP-9, which are STAT3 target genes in both
MCF-7 and MCF-7/ADR cells (Fig. 6C and
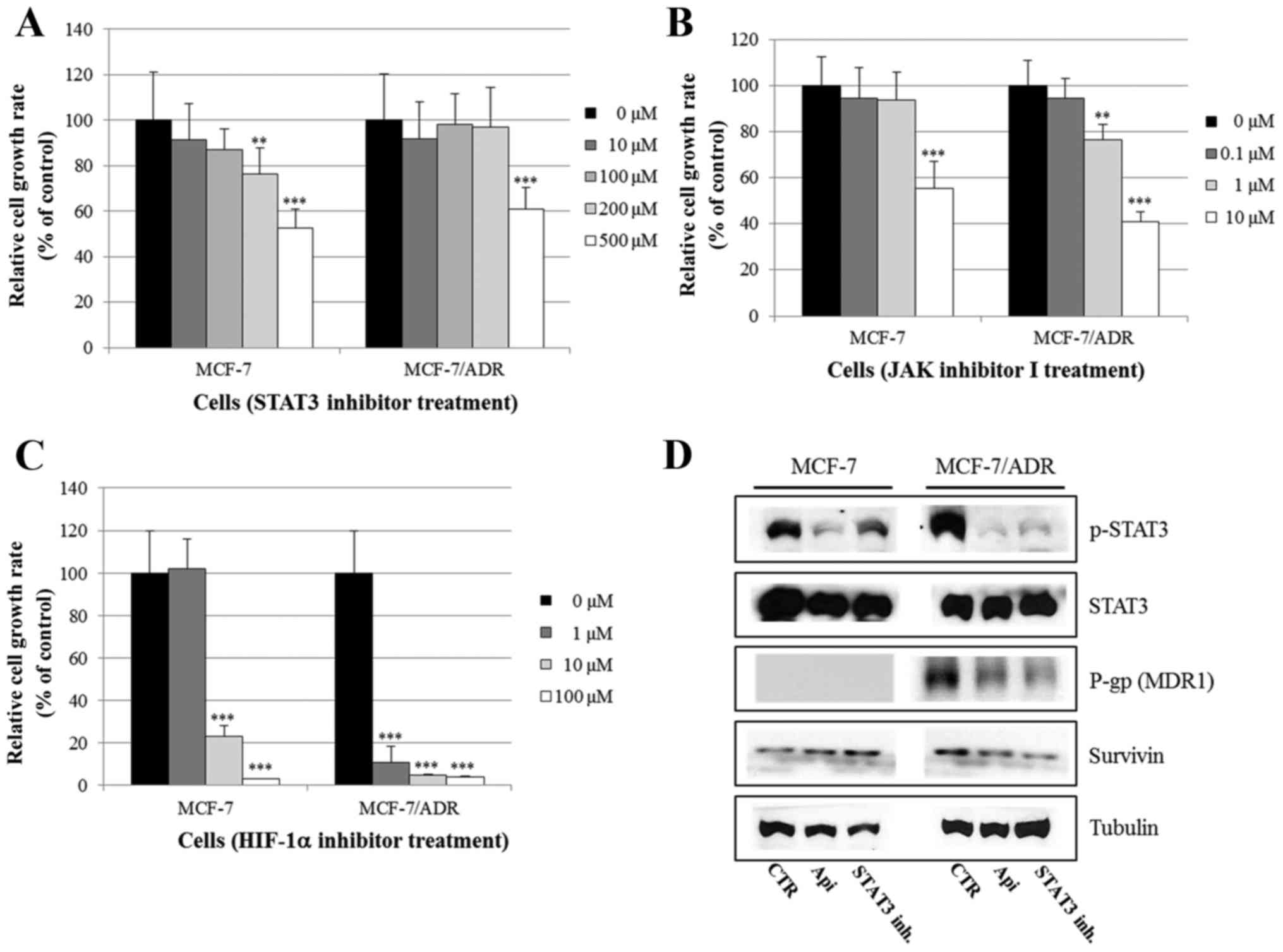
D). Moreover, the STAT3 inhibitor S3I-201, the JAK inhibitor I,
and the HIF-1α inhibitor EF-24 induced cell growth inhibition in
both MCF-7 and MCF-7/ADR cells, as shown in Fig. 7A-C. Fig.
7D demonstrates that both apigenin (80 µM) and the STAT3
inhibitor S3I-201 reduced the expression of p-STAT3, P-gp (MDR1)
and survivin (a STAT3 target gene) in MCF-7/ADR cells. These
results indicate that apigenin suppresses drug resistance by
inhibiting the STAT3 signaling pathway in MCF-7/ADR cells.
Therefore, STAT3 may be a promising therapeutic target for
overcoming drug resistance in breast cancer.
Discussion
Multidrug resistance (MDR) is a serious problem that
leads to therapeutic failure in various cancers, and overcoming MDR
is an important endeavor that is necessary to increase the overall
survival of cancer patients. Natural plant products, such as
phytoestrogens, are objects of intense research for their potential
to reverse MDR, which would result in successful chemotherapy
treatments. In the present study, the mechanism by which apigenin
overcomes drug resistance was investigated in breast cancer. The
aim of the study was to determine whether apigenin might be a
useful compound for reversing drug resistance. For this purpose, we
used the adriamycin-resistant MCF-7/ADR cells.
Apigenin significantly inhibited the growth of both
MCF-7 and MCF-7/ADR cells. The clonogenic survival assay revealed
that apigenin decreased anchorage-dependent colony formation of
both cell lines in a dose-dependent manner. This growth inhibition
was associated with an increase in the
sub-G0/G1 apoptotic population in MCF-7 and
MCF-7/ADR cells. Apigenin increased the number of apoptotic cells
in a dose-dependent manner, as assessed by Annexin V assays.
Moreover, apigenin induced apoptosis via a caspase-dependent
apoptosis pathway, as shown by the cleavage of caspase-8 and PARP.
We did not detect caspase-3, which is important in the apoptosis
pathway, because MCF-7 cells do not contain caspase-3 due to a
genomic deletion (28). Our results
indicate that apigenin contains strong apoptotic properties.
Activation of apoptotic caspases induces inactivation or activation
of substrates, leading to a signaling cascade, and the initiation
of this signaling pathway permits the controlled destruction of
cellular components (29).
Apigenin-induced apoptosis seems to be mediated by the activation
of caspase-8.
Apigenin downregulated the mRNA expression of MDR1
and MRPs, as well as the protein expression of P-gp (MDR1).
Apigenin also inhibited drug efflux from the MCF-7/ADR
adriamycin-resistant cells in a dose-dependent manner, as revealed
by the rhodamine 123 efflux assay. These results indicate that
apigenin overcomes drug resistance. In agreement with our data, it
was reported that apigenin inhibits anti-estrogen-resistant breast
cancer cell growth through estrogen receptor-α-dependent and
independent mechanisms (30). It
was also reported that apigenin inhibited not only P-gp (MDR1) but
also BCRP by increasing cellular uptake of adriamycin in
multidrug-resistant cells (31).
P-gp is a transmembrane glycoprotein that diminishes intracellular
drug concentrations by pumping drugs out of the cells (27). Regulation of P-gp expression is
important to avoid drug resistance; overexpression of P-gp induces
resistance to chemotherapeutic reagents. Apigenin appears to
inhibit P-gp-mediated drug resistance in MCF-7/ADR cells.
Interestingly, apigenin reduced the expression of
p-STAT3, suggesting that it negatively regulates the STAT3 pathway
in MCF-7/ADR cells. Apigenin inhibited nuclear localization of
STAT3 in MCF-7/ADR cells, as revealed by immunocytochemistry.
Apigenin inhibited the production of VEGF and MMP-9, which are
STAT3 target genes, in MCF-7/ADR cells, as revealed by the ELISA
assay. The STAT3 inhibitor S3I-201, JAK inhibitor I and the HIF-1α
inhibitor EF-24 decreased the growth of both MCF-7 and MCF-7/ADR
cells. These results clearly indicate that apigenin induces
growth-suppressive activity and overcomes drug resistance by
inhibiting the STAT3 signaling pathway. STAT3 is a transcription
factor that regulates target gene expression in response to
cytokines or growth factors and plays a key role in many cellular
processes. STAT3 traditionally operates as a cancer promoter,
however, it upregulates cancer suppressor genes as revealed
recently (32,33). STAT3 promotes the increase of cell
growth, angiogenesis, invasion and metastasis and inhibits
apoptosis (34–36). Overexpression of STAT3 and p-STAT3
is related to poor prognosis in melanoma cancer (34–36).
Several upstream kinases (Jak and Src) are responsible for the
constitutive STAT3 phosphorylation (37,38).
It is known that resveratrol inhibits STAT3 signaling inducing
cancer cells containing p-STAT3 (39). STAT3 (40) and HIF-1 (41) can bind to VEGF promoter which
contains various transcription factor binding sites. The
interaction of STAT3 with HIF-1 induces VEGF transcriptional
activation (42). Constitutive
activation of STAT3 is responsible for the resistance to
chemotherapy-induced apoptosis in some tumors (27). Moreover, it was reported that
multidrug resistance was associated with STAT3 mRNA
overexpression in cisplatin-resistant lung cancer cells (27). STAT3 is activated highly in MDR
malignancies, and inhibition of STAT3 activity might reverse
chemoresistance (27). Constitutive
activation of the STAT3 pathway could be an early indicator of drug
resistance. Therefore, apigenin seems to reverse drug resistance by
inhibiting STAT3 signaling, which indicates that apigenin could be
a useful natural therapy that overcomes drug resistance.
It should be noted that apigenin acts as a
suppressor of MUC1 expression in MCF-7 brest cancer cells.
Moreover, it was proved that this loss of expression is associated
with the loss of viability of MCF-7 cells (43). Apigenin could be a promising therapy
for the treatment and prevention of breast cancer.
Acknowledgements
This study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
(NRF), funded by the Ministry of Science, ICT and Future Planning
(2015R1C1A2A01051539). This study was also supported by a grant
funded by the Traditional Korean Medicine R&D Project of the
Ministry of Health and Welfare (HI12C1889 and HI13C0530).
References
|
1
|
Holohan C, Van Schaeybroeck S, Longley DB
and Johnston PG: Cancer drug resistance: An evolving paradigm. Nat
Rev Cancer. 13:714–726. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Luqmani YA: Mechanisms of drug resistance
in cancer chemotherapy. Med Princ Pract. 14:(Suppl 1). 35–48. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zahreddine H and Borden KL: Mechanisms and
insights into drug resistance in cancer. Front Pharmacol. 4:282013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xue X and Liang XJ: Overcoming drug
efflux-based multidrug resistance in cancer with nanotechnology.
Chin J Cancer. 31:100–109. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Amin ML: P-glycoprotein inhibition for
optimal drug delivery. Drug Target Insights. 7:27–34. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee JH, Zhou HY, Cho SY, Kim YS, Lee YS
and Jeong CS: Anti-inflammatory mechanisms of apigenin: inhibition
of cyclooxygenase-2 expression, adhesion of monocytes to human
umbilical vein endothelial cells, and expression of cellular
adhesion molecules. Arch Pharm Res. 30:1318–1327. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Romanová D, Vachálková A, Cipák L, Ovesná
Z and Rauko P: Study of antioxidant effect of apigenin, luteolin
and quercetin by DNA protective method. Neoplasma. 48:104–107.
2001.PubMed/NCBI
|
|
8
|
Moon DO, Kim MO, Choi YH, Lee HG, Kim ND
and Kim GY: Gossypol suppresses telomerase activity in human
leukemia cells via regulating hTERT. FEBS Lett. 582:3367–3373.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nakazawa T, Yasuda T, Ueda J and Ohsawa K:
Antidepressant-like effects of apigenin and
2,4,5-trimethoxycinnamic acid from Perilla frutescens in the forced
swimming test. Biol Pharm Bull. 26:474–480. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shukla S and Gupta S: Apigenin: A
promising molecule for cancer prevention. Pharm Res. 27:962–978.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Heederik D, Kromhout H, Burema J,
Biersteker K and Kromhout D: Occupational exposure and 25-year
incidence rate of non-specific lung disease: The Zutphen Study. Int
J Epidemiol. 19:945–952. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Knekt P, Järvinen R, Seppänen R,
Hellövaara M, Teppo L, Pukkala E and Aromaa A: Dietary flavonoids
and the risk of lung cancer and other malignant neoplasms. Am J
Epidemiol. 146:223–230. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hertog MG, Feskens EJ, Hollman PC, Katan
MB and Kromhout D: Dietary flavonoids and cancer risk in the
Zutphen Elderly Study. Nutr Cancer. 22:175–184. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rossi M, Negri E, Lagiou P, Talamini R,
Dal Maso L, Montella M, Franceschi S and La Vecchia C: Flavonoids
and ovarian cancer risk: A case-control study in Italy. Int J
Cancer. 123:895–898. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bosetti C, Spertini L, Parpinel M,
Gnagnarella P, Lagiou P, Negri E, Franceschi S, Montella M,
Peterson J, Dwyer J, et al: Flavonoids and breast cancer risk in
Italy. Cancer Epidemiol Biomarkers Prev. 14:805–808. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shukla S, Bhaskaran N, Babcook MA, Fu P,
Maclennan GT and Gupta S: Apigenin inhibits prostate cancer
progression in TRAMP mice via targeting PI3K/Akt/FoxO pathway.
Carcinogenesis. 35:452–460. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Budhraja A, Gao N, Zhang Z, Son YO, Cheng
S, Wang X, Ding S, Hitron A, Chen G, Luo J, et al: Apigenin induces
apoptosis in human leukemia cells and exhibits anti-leukemic
activity in vivo. Mol Cancer Ther. 11:132–142. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu LZ, Fang J, Zhou Q, Hu X, Shi X and
Jiang BH: Apigenin inhibits expression of vascular endothelial
growth factor and angiogenesis in human lung cancer cells:
Implication of chemoprevention of lung cancer. Mol Pharmacol.
68:635–643. 2005.PubMed/NCBI
|
|
19
|
Seo HS, Ju JH, Jang K and Shin I:
Induction of apoptotic cell death by phytoestrogens by
up-regulating the levels of phospho-p53 and p21 in normal and
malignant estrogen receptor α-negative breast cells. Nutr Res.
31:139–146. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu HF, Chie YJ, Yang MS, Lee CS, Fu JJ,
Yang JS, Tan TW, Wu SH, Ma YS, Ip SW, et al: Apigenin induces
caspase-dependent apoptosis in human lung cancer A549 cells through
Bax- and Bcl-2-triggered mitochondrial pathway. Int J Oncol.
36:1477–1484. 2010.PubMed/NCBI
|
|
21
|
Wang W, Heideman L, Chung CS, Pelling JC,
Koehler KJ and Birt DF: Cell-cycle arrest at G2/M and growth
inhibition by apigenin in human colon carcinoma cell lines. Mol
Carcinog. 28:102–110. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Turktekin M, Konac E, Onen HI, Alp E,
Yilmaz A and Menevse S: Evaluation of the effects of the flavonoid
apigenin on apoptotic pathway gene expression on the colon cancer
cell line (HT29). J Med Food. 14:1107–1117. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gupta S, Afaq F and Mukhtar H: Involvement
of nuclear factor-kappa B, Bax and Bcl-2 in induction of cell cycle
arrest and apoptosis by apigenin in human prostate carcinoma cells.
Oncogene. 21:3727–3738. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ruela-de-Sousa RR, Fuhler GM, Blom N,
Ferreira CV, Aoyama H and Peppelenbosch MP: Cytotoxicity of
apigenin on leukemia cell lines: Implications for prevention and
therapy. Cell Death Dis. 1:e192010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ujiki MB, Ding XZ, Salabat MR, Bentrem DJ,
Golkar L, Milam B, Talamonti MS, Bell RH Jr, Iwamura T and Adrian
TE: Apigenin inhibits pancreatic cancer cell proliferation through
G2/M cell cycle arrest. Mol Cancer. 5:762006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Siveen KS, Sikka S, Surana R, Dai X, Zhang
J, Kumar AP, Tan BK, Sethi G and Bishayee A: Targeting the STAT3
signaling pathway in cancer: Role of synthetic and natural
inhibitors. Biochim Biophys Acta. 1845:136–154. 2014.PubMed/NCBI
|
|
27
|
Zhang X, Xiao W, Wang L, Tian Z and Zhang
J: Deactivation of signal transducer and activator of transcription
3 reverses chemotherapeutics resistance of leukemia cells via
down-regulating P-gp. PLoS One. 6:e209652011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jänicke RU: MCF-7 breast carcinoma cells
do not express caspase-3. Breast Cancer Res Treat. 117:219–221.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
McIlwain DR, Berger T and Mak TW: Caspase
functions in cell death and disease. Cold Spring Harb Perspect
Biol. 5:a0086562013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Long X, Fan M, Bigsby RM and Nephew KP:
Apigenin inhibits antiestrogen-resistant breast cancer cell growth
through estrogen receptor-alpha-dependent and estrogen
receptor-alpha-independent mechanisms. Mol Cancer Ther.
7:2096–2108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Saeed M, Kadioglu O, Khalid H, Sugimoto Y
and Efferth T: Activity of the dietary flavonoid, apigenin, against
multidrug-resistant tumor cells as determined by pharmacogenomics
and molecular docking. J Nutr Biochem. 26:44–56. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
de la Iglesia N, Konopka G, Puram SV, Chan
JA, Bachoo RM, You MJ, Levy DE, Depinho RA and Bonni A:
Identification of a PTEN-regulated STAT3 brain tumor suppressor
pathway. Genes Dev. 22:449–462. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lewis HD, Winter A, Murphy TF, Tripathi S,
Pandey VN and Barton BE: STAT3 inhibition in prostate and
pancreatic cancer lines by STAT3 binding sequence oligonucleotides:
Differential activity between 5′ and 3′ ends. Mol Cancer Ther.
7:1543–1550. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kortylewski M, Jove R and Yu H: Targeting
STAT3 affects melanoma on multiple fronts. Cancer Metastasis Rev.
24:315–327. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Niu G, Bowman T, Huang M, Shivers S,
Reintgen D, Daud A, Chang A, Kraker A, Jove R and Yu H: Roles of
activated Src and Stat3 signaling in melanoma tumor cell growth.
Oncogene. 21:7001–7010. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xie TX, Huang FJ, Aldape KD, Kang SH, Liu
M, Gershenwald JE, Xie K, Sawaya R and Huang S: Activation of stat3
in human melanoma promotes brain metastasis. Cancer Res.
66:3188–3196. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sellers LA, Feniuk W, Humphrey PP and
Lauder H: Activated G protein-coupled receptor induces tyrosine
phosphorylation of STAT3 and agonist-selective serine
phosphorylation via sustained stimulation of mitogen-activated
protein kinase. Resultant effects on cell proliferation. J Biol
Chem. 274:16423–16430. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang Y, Turkson J, Carter-Su C, Smithgall
T, Levitzki A, Kraker A, Krolewski JJ, Medveczky P and Jove R:
Activation of Stat3 in v-Src-transformed fibroblasts requires
cooperation of Jak1 kinase activity. J Biol Chem. 275:24935–24944.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kotha A, Sekharam M, Cilenti L, Siddiquee
K, Khaled A, Zervos AS, Carter B, Turkson J and Jove R: Resveratrol
inhibits Src and Stat3 signaling and induces the apoptosis of
malignant cells containing activated Stat3 protein. Mol Cancer
Ther. 5:621–629. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Niu G, Wright KL, Huang M, Song L, Haura
E, Turkson J, Zhang S, Wang T, Sinibaldi D, Coppola D, et al:
Constitutive Stat3 activity up-regulates VEGF expression and tumor
angiogenesis. Oncogene. 21:2000–2008. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Forsythe JA, Jiang BH, Iyer NV, Agani F,
Leung SW, Koos RD and Semenza GL: Activation of vascular
endothelial growth factor gene transcription by hypoxia-inducible
factor 1. Mol Cell Biol. 16:4604–4613. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jung JE, Lee HG, Cho IH, Chung DH, Yoon
SH, Yang YM, Lee JW, Choi S, Park JW, Ye SK, et al: STAT3 is a
potential modulator of HIF-1-mediated VEGF expression in human
renal carcinoma cells. FASEB J. 19:1296–1298. 2005.PubMed/NCBI
|
|
43
|
Zhou Y, Rajabi H and Kufe D: Mucin 1
C-terminal subunit oncoprotein is a target for small-molecule
inhibitors. Mol Pharmacol. 79:886–893. 2011. View Article : Google Scholar : PubMed/NCBI
|