Introduction
The endocannabinoid system, which includes the
G-protein-coupled cannabinoid receptors 1 and 2 (CB1 and CB2) and
their endogenous ligands (endocannabinoids), participates in a
variety of physiological and pathological conditions, including
inflammation and cancer (1–3). CB1 is abundantly expressed in the
central nervous system, while CB2 is predominantly expressed by
immune cells. Endogenous and exogenous cannabinoids exert
anti-inflammatory and antitumor effects (1,4–9).
Glioma, the most common type of brain tumor,
originates from differentiated glial cells of the central nervous
system. Glial fibrillary acidic protein (GFAP) is a marker of glial
differentiation (10). Low-grade
malignancies have moderate expression of the protein, while
high-grade malignancies have weak GFAP expression (11). Glioblastoma, a poorly differentiated
type of glioma, is highly malignant and exhibits aggressive
invasive growth. Existing treatments, including surgery,
radiotherapy and chemotherapy, are largely unable to cure
glioblastoma, which leads to death in most cases (12,13).
Therefore, new therapeutic approaches for treating glioblastoma are
urgently needed. Recently, many synthetic agonists for cannabinoid
receptors, which act on the endocannabinoid system, have been
developed. Of these synthetic agonists, WIN 55,212-2 and KM-233
mimic the effects of endogenous cannabinoids and decrease cell
viability in both glioma and glioblastoma (14,15).
Various microenvironmental stresses modulate cell
signaling in physiological and pathological conditions. Hypoxia, a
type of stress that is fundamental in the microenvironment of a
variety of solid tumors (16), has
been implicated in the persistence and development of malignancies.
Hypoxic stimuli prevent apoptosis and promote tumor invasion,
metastasis and angiogenesis by regulating a variety of genes,
including vascular endothelial growth factor (VEGF) and
hypoxia-inducible factor 1α (17).
Hypoxic regions are frequently found in glioblastoma (18,19)
and extensive hypoxia is associated with a worse prognosis
(16).
In the present study, we examined the effects of
hypoxia on the endocannabinoid system in U-87 MG human malignant
glioblastoma cells. We found that hypoxia suppressed the expression
of the genes encoding GFAP and cannabinoid receptors; increased the
expression of the gene encoding cyclooxygenase (COX)-2, the enzyme
that metabolizes endocannabinoids (20,21),
and attenuated cannabinoid receptor agonist (WIN 55,212-2)-induced
cell death. Our findings provide insight into novel mechanisms by
which hypoxia inhibits the endocannabinoid system and promotes the
malignant phenotype of glioblastoma.
Materials and methods
Chemicals
WIN 55,212-2 and Dulbecco's modified Eagle's medium
(DMEM) were obtained from Wako Pure Chemical Industries (Osaka,
Japan). Fetal bovine serum (FBS) was obtained from Invitrogen
(Carlsbad, CA, USA).
Preparation of primary rat glial
cells
All experiments were performed in accordance with
the guidelines set by the Animal Welfare Committee of Kanazawa
University. Primary glial cells were prepared from newborn
Sprague-Dawley rats as previously described (22–24).
Briefly, following isoflurane anesthesia, rats were decapitated and
cells were mechanically dissociated from the hippocampi and plated
onto culture dishes (35 mm).
Cell culture
U-87 MG human malignant glioblastoma cells were
provided by Dr Nakata (Kanazawa University, Kanazawa, Japan). U-87
MG and primary rat glial cells were maintained in DMEM containing
10% FBS at 37°C in 5% CO2. Then, cells were divided into
2 groups. The cells in the normoxia group were incubated in 5%
CO2 and 20% O2, while the cells in the
hypoxia group were maintained in 5% CO2 and 1.5%
O2.
Reverse transcription polymerase chain
reaction (RT-PCR)
To evaluate the expression of human CB1
(CNR1), rat CB1 (Cnr1), human CB2 (CNR2), rat
CB2 (Cnr2), human COX-1 (PTGS1), human COX-2
(PTGS2), human VEGF (VEGFA) and human GFAP
(GFAP) mRNA in the cells, RT-PCR was performed as follows.
Briefly, RNA was extracted from the cells and cDNA was generated
using the reverse transcriptase ReverTra Ace® (Toyobo,
Tokyo, Japan). We performed PCR-based, subtype-specific gene
amplification with LA Taq (Takara, Tokyo, Japan) using the primers
specified in Table I.
 | Table I.Nucleotide sequences of the PCR
primers used for amplification of the genes encoding human CB1
(CNR1; NM_001160226), rat CB1 (Cnr1; NM_012784),
human CB2 (CNR2; NM_001841), rat CB2 (Cnr2;
NM_001164142), human COX-1 (PTGS1; NM_000962), human COX-2
(PTGS2; NM_000963), human VEGF (VEGFA; NM_001025366),
human GFAP (GFAP; NM_001131019), human β-actin (ACTB;
NM_001101) and rat β-actin (Actb; NM_031144). |
Table I.
Nucleotide sequences of the PCR
primers used for amplification of the genes encoding human CB1
(CNR1; NM_001160226), rat CB1 (Cnr1; NM_012784),
human CB2 (CNR2; NM_001841), rat CB2 (Cnr2;
NM_001164142), human COX-1 (PTGS1; NM_000962), human COX-2
(PTGS2; NM_000963), human VEGF (VEGFA; NM_001025366),
human GFAP (GFAP; NM_001131019), human β-actin (ACTB;
NM_001101) and rat β-actin (Actb; NM_031144).
| Gene | Forward
(5′→3′) | Reverse
(5′→3′) | Products (bp) |
|---|
| Human CB1
(CNR1; NM_001160226) | CAG GCC TTC CTA CCA
CTT CAT | ACC CCA CCC AGT TTG
AAC AGA | 141 |
| Rat CB1
(Cnr1; NM_012784) | CAG GCC TTC CTA CCA
CTT CAT | ACC CCA CCC AGT TTG
AAC AGA | 141 |
| Human CB2
(CNR2; NM_001841) | AAG CCC TCA TAC CTG
TTC AT | ACA GAG GCT GTG AAG
GTC AT | 164 |
| Rat CB2
(Cnr2; NM_001164142) | AAG CCC TCG TAC CTG
TTC AT | ACA GAG GCC GTG AAG
GTC AT | 164 |
| Human COX-1
(PTGS1; NM_000962) | AAT GCC ACC TTC ATC
CGA GA | TGG GTG AAG TGT TGT
GCA AAG | 311 |
| Human COX-2
(PTGS2; NM_000963) | CAG CAA ATC CTT GCT
GTT CC | GTG AAG TGC TGG GCA
AAG AAT | 526 |
| Human VEGF
(VEGFA; NM_001025366) | CCT TGC CTT GCT GCT
CTA CCT | GAT GTC CAC CAG GGT
CTC GAT | 151 |
| Human GFAP
(GFAP; NM_001131019) | GAT GAT GGA GCT CAA
TGA CCG | CAG CCT CAG GTT GGT
TTC ATC | 274 |
| Human β-actin
(ACTB; NM_001101) | ATG GTG GGC ATG GGT
CAG AAG | CTG GGG TGT TGA AGG
TCT CAA | 262 |
| Rat β-actin
(Actb; NM_031144) | ATG GTG GGT ATG GGT
CAG AAG | CTG GGG TGT TGA AGG
TCT CAA | 262 |
Cell proliferation and viability
assays
Cell proliferation and viability were analyzed using
the Cell Counting Kit-8 (CCK-8; Wako) as previously described
(25). U-87 MG cells were seeded in
96-well plates at a density of 1×102 cells/well. After a
24-h incubation period, the cells were divided into 2 groups. The
cells in the normoxia group were incubated in 5% CO2 and
20% O2, while the cells in the hypoxia group were
maintained in 5% CO2 and 1.5% O2. After 3
days of continuous treatment, cells were allowed to recover in 5%
CO2 and 20% O2 for ~30 min. Then, the cells
were incubated with WST-8 (10 µl of WST-8 in 100 µl of medium) for
3 h at 37°C. The absorbance of the colored formazan product,
generated by mitochondrial dehydrogenases in metabolically active
cells, was recorded at 450 nm. Cell proliferation and viability
were expressed as a ratio of the absorbance in treated wells
relative to that in untreated control wells.
Cannabinoid receptor agonist
treatment
Cells were treated with the cannabinoid receptor
agonist WIN 55,212-2 in 5% CO2 and 20% O2
following a prior 3-day exposure to normoxia or hypoxia. After 2
days of agonist treatment, cell viability was analyzed using CCK-8
(Wako), as described above.
Statistical analysis
Data are presented as the mean ± standard error of
the mean (SEM) from at least 3 independent experiments. Statistical
analysis was performed using Student's unpaired t-test or
Kruskal-Wallis non-parametric analysis of variance (ANOVA) followed
by a Bonferroni post hoc test, and results were considered
statistically significant at P<0.05.
Results
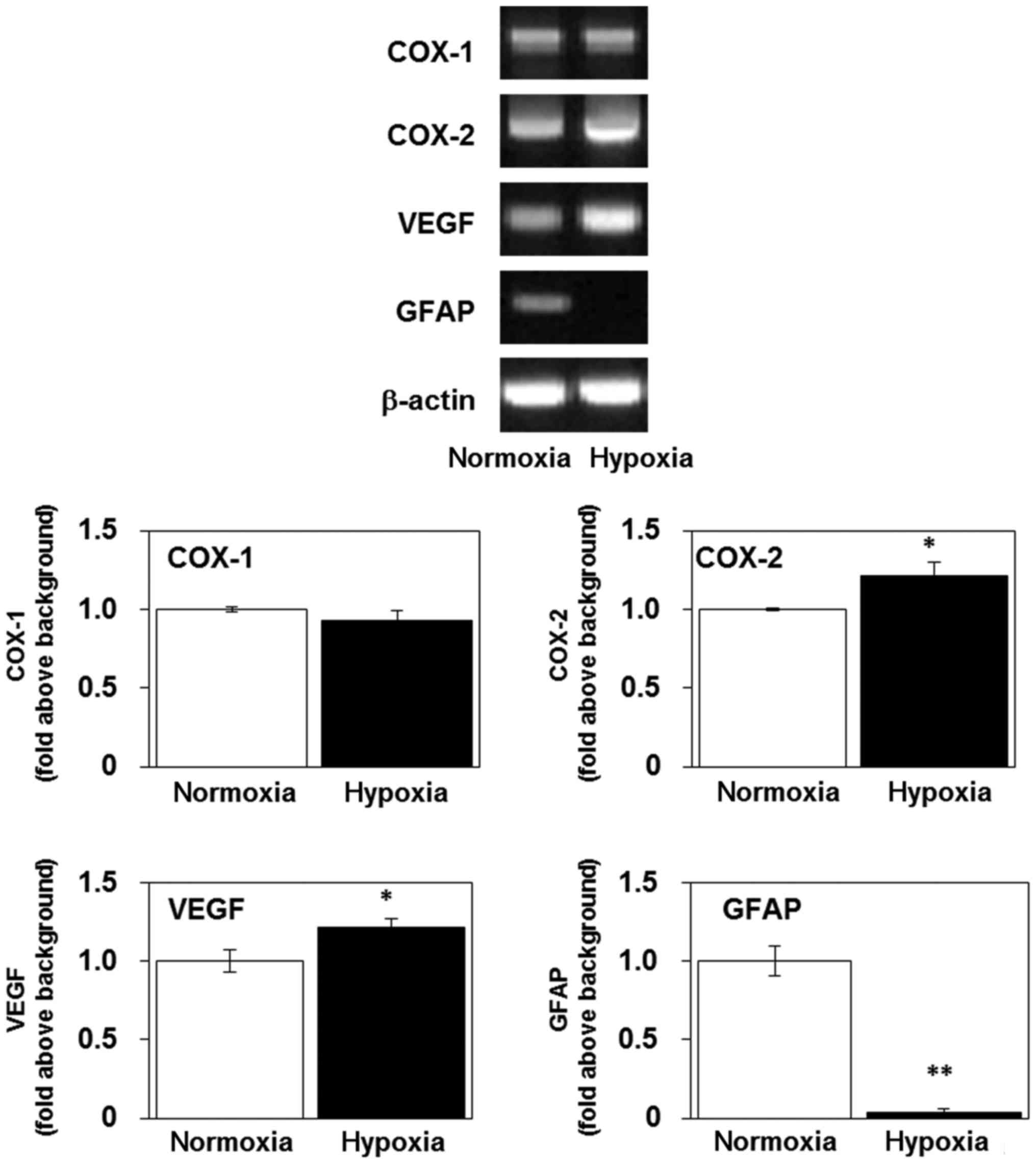
Hypoxia increases the levels of mRNA
encoding COX-2 and VEGF, and decreases the level of mRNA encoding
GFAP, in U-87 MG cells
We examined the effects of continuous exposure (3
days) to hypoxia on the expression of the genes for COX-1, COX-2,
VEGF and GFAP. Hypoxic exposure increased the levels of the mRNA
encoding COX-2 and VEGF, but not COX-1, in the U-87 MG cells
(Fig. 1), indicating that hypoxia
activated inflammatory and angiogenic responses (26,27).
Notably, GFAP mRNA expression decreased over 72 h of hypoxia
(Fig. 1), indicating that the cells
had begun to dedifferentiate. Inflammation, angiogenesis and
dedifferentiation are predictors of poor antitumor treatment
responses and have previously been reported to potently activate
tumor progression and invasion (28,29).
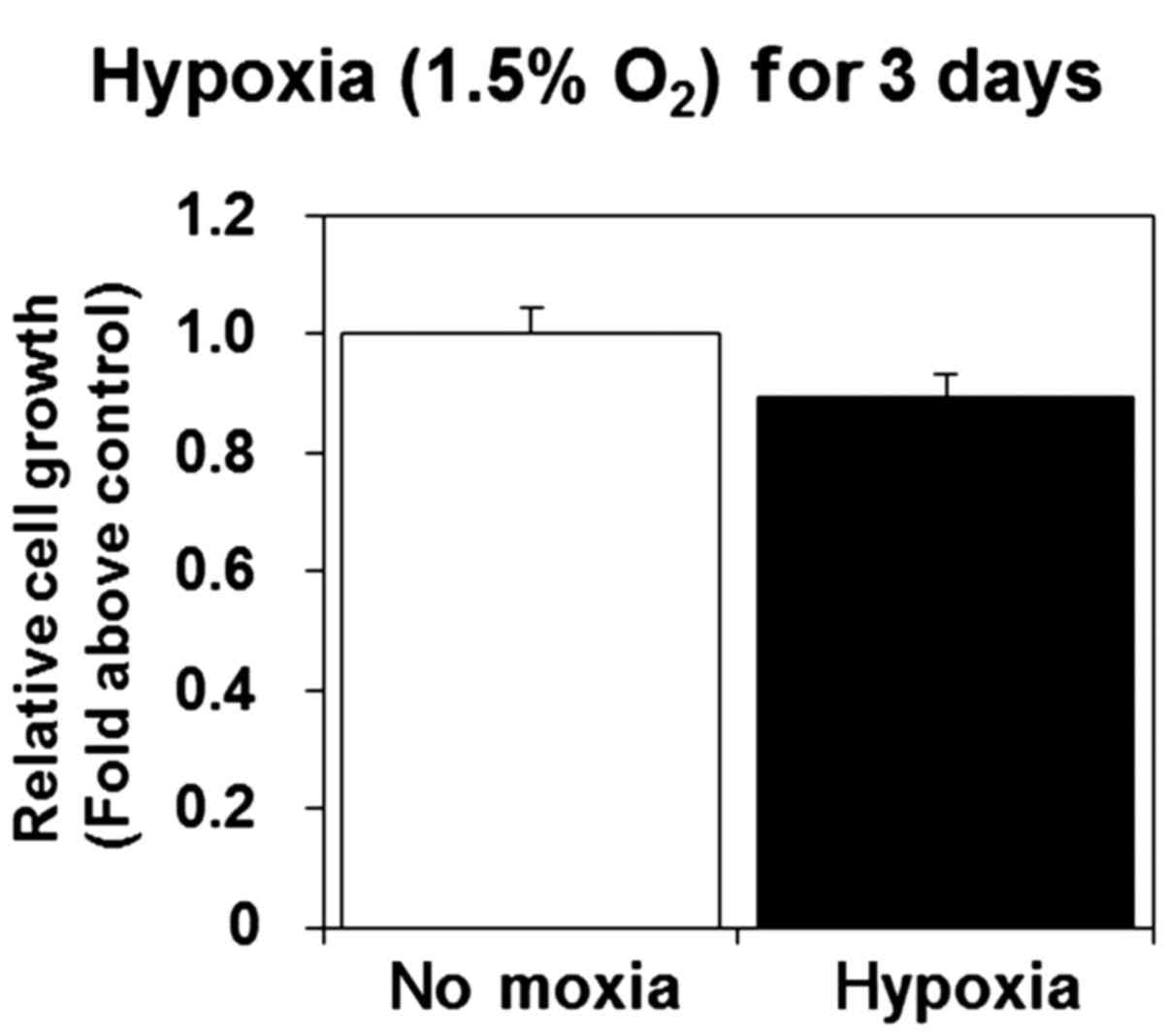
Hypoxia does not affect cell viability
and induces lamellipodia formation in U-87 MG cells
We previously showed that a 3-day exposure to
hypoxia inhibited cell proliferation and viability in mouse
fibroblast cells (30). In the
present study, we investigated the effect of sustained exposure to
hypoxia on U-87 MG cell proliferation and viability. We found that
proliferation and viability were not significantly altered by
hypoxia (Fig. 2), indicating that
U-87 MG cells, unlike mouse fibroblast cells, are tolerant to
hypoxia.
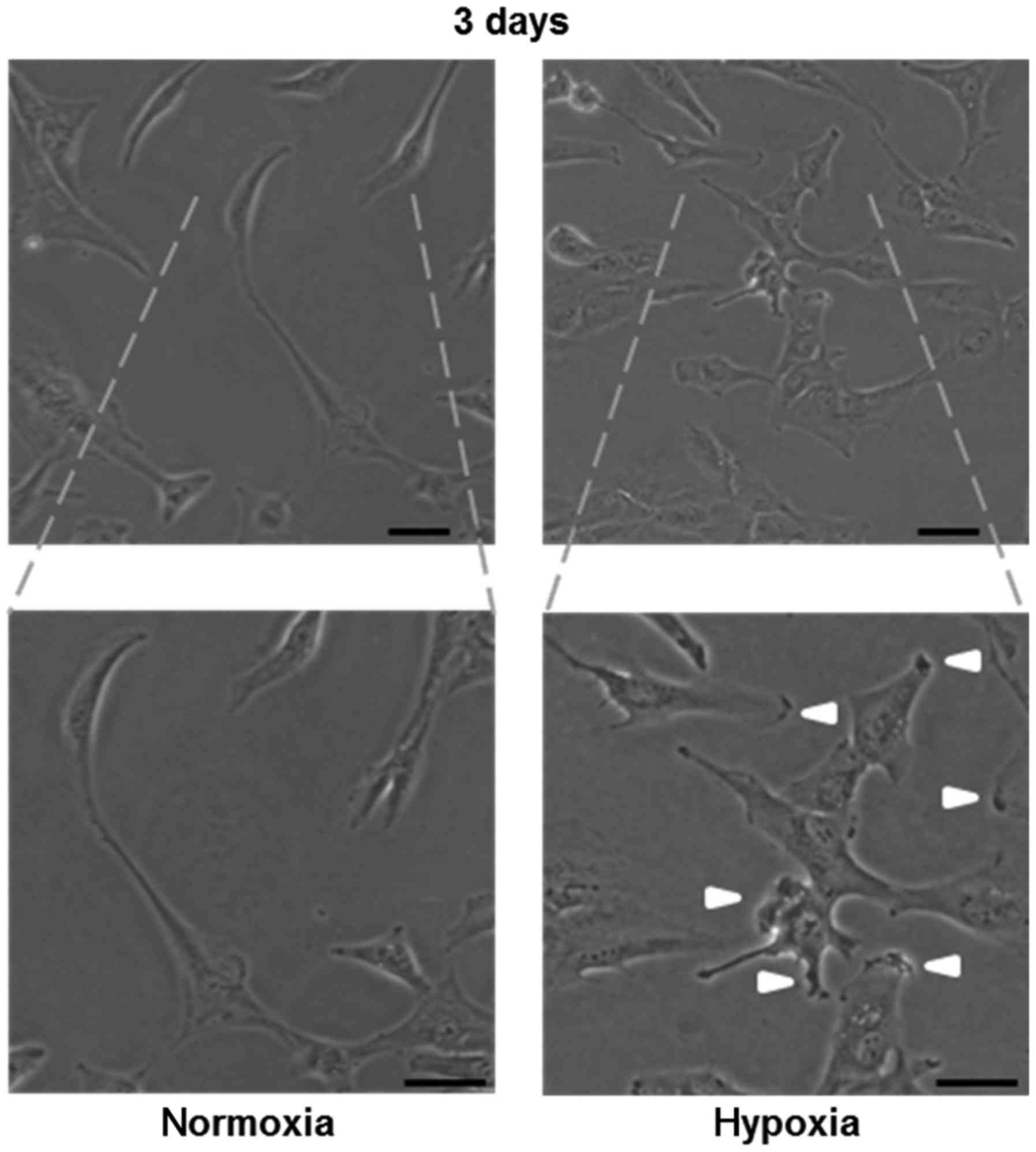
Next, we examined the morphology of U-87 MG cells
after sustained exposure to hypoxia. We found that lamellipodia
formation occurred at the cell edges (Fig. 3), which is known to facilitate cell
migration (31).
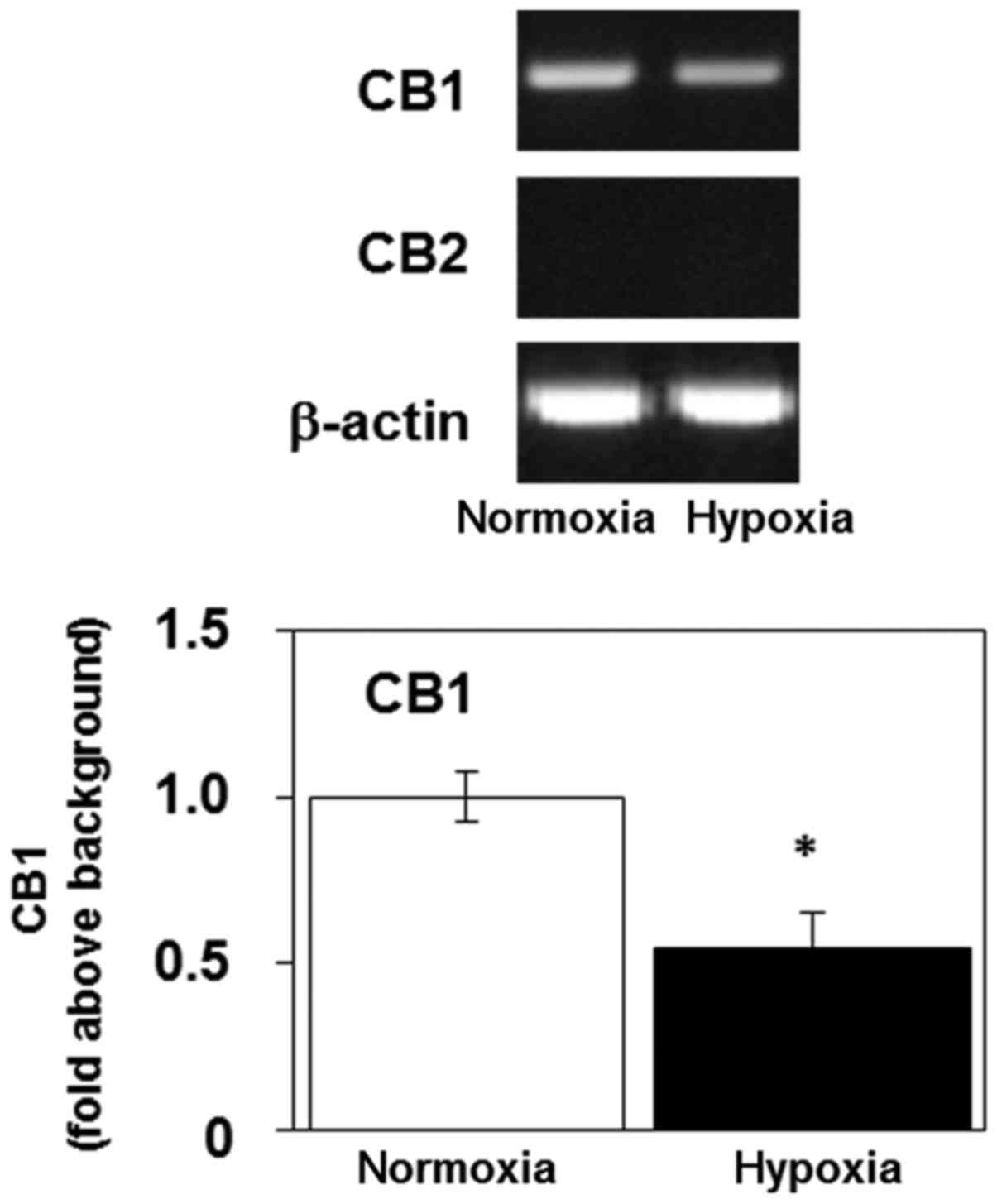
Hypoxia decreases the level of mRNA
encoding CB1 in U-87 MG cells
Next, we examined the expression of CB1
(CNR1) and CB2 (CNR2) mRNA in U-87 MG cells using
RT-PCR. The cannabinoid receptors facilitate the transduction of
extracellular signals to the cytoplasm (2). The gene product encoding CB1 was
detectable in U-87 MG cells, but the level of mRNA encoding CB2 was
below the limit of detection (Fig.
4). Sustained 3-day exposure of U-87 MG cells to hypoxia
decreased their expression of mRNA encoding CB1 (Fig. 4). These results indicate that
hypoxia may regulate the cannabinoid system by modulating the
expression of CB1.
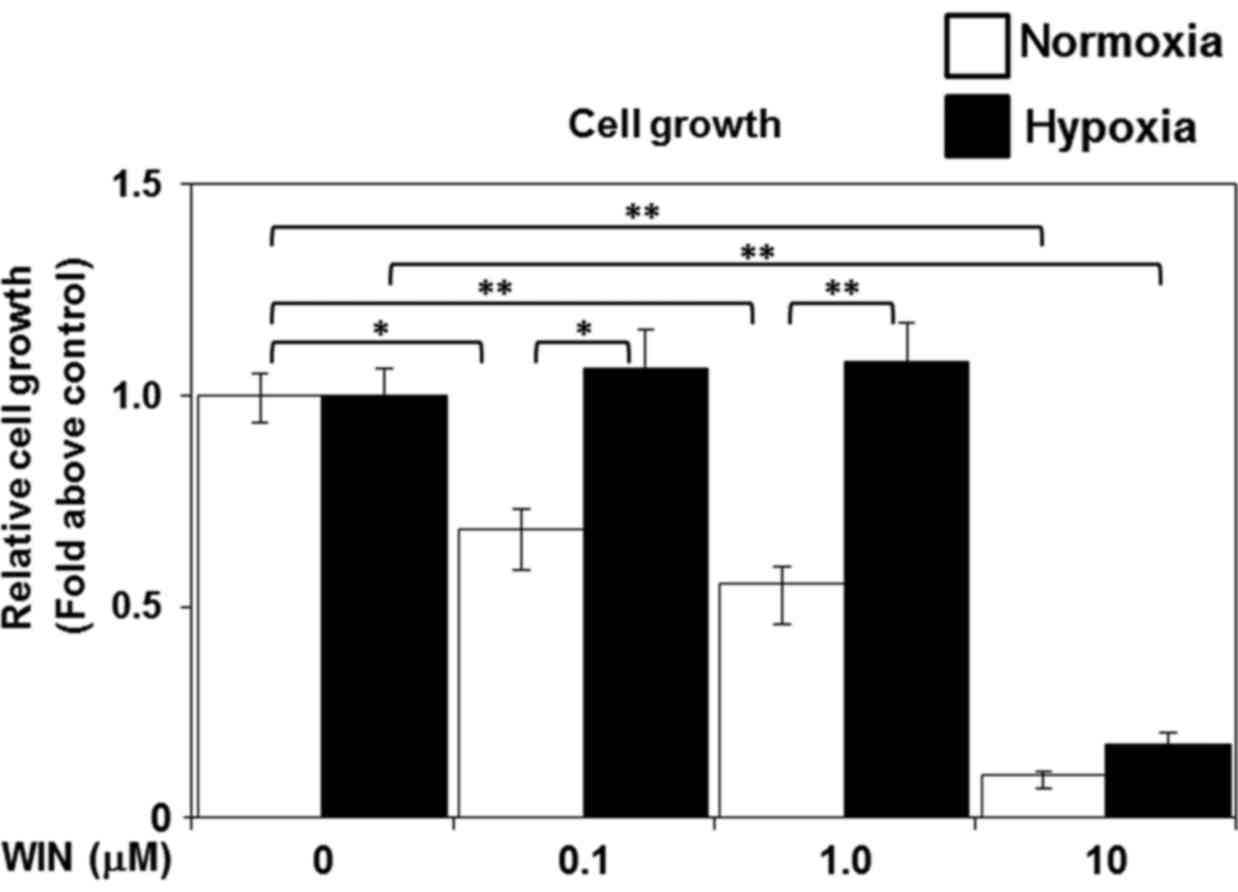
Hypoxia attenuates cannabinoid
receptor agonist-induced cell death in U-87 MG cells
Recently, cannabinoid receptor agonists have been
shown to mimic the effects of endogenous cannabinoids and cause
cell death in glioblastoma (15).
Given the hypoxia-induced downregulation of cannabinoid receptor
expression that we observed, we next examined the effects of
sustained hypoxia on cannabinoid receptor agonist-induced cell
death. We treated the cells with WIN 55,212-2 (0.1, 1.0, or 10 µM)
after an initial 3-day exposure to hypoxia (1.5% O2) or
normoxia (20% O2). We analyzed cell proliferation and
viability 2 days after treatment with WIN 55,212-2. WIN 55,212-2
treatment significantly reduced the viability of the cells under
normoxic conditions in a dose-dependent manner, compared to the
viability of the untreated normoxic controls (Fig. 5). However, the low doses of WIN
55,212-2 (0.1 or 1.0 µM) failed to induce cell death in the hypoxia
group (Fig. 5). These results
suggest that hypoxia confers tolerance to cannabinoid receptor
agonist-induced cellular toxicity in glioblastoma.
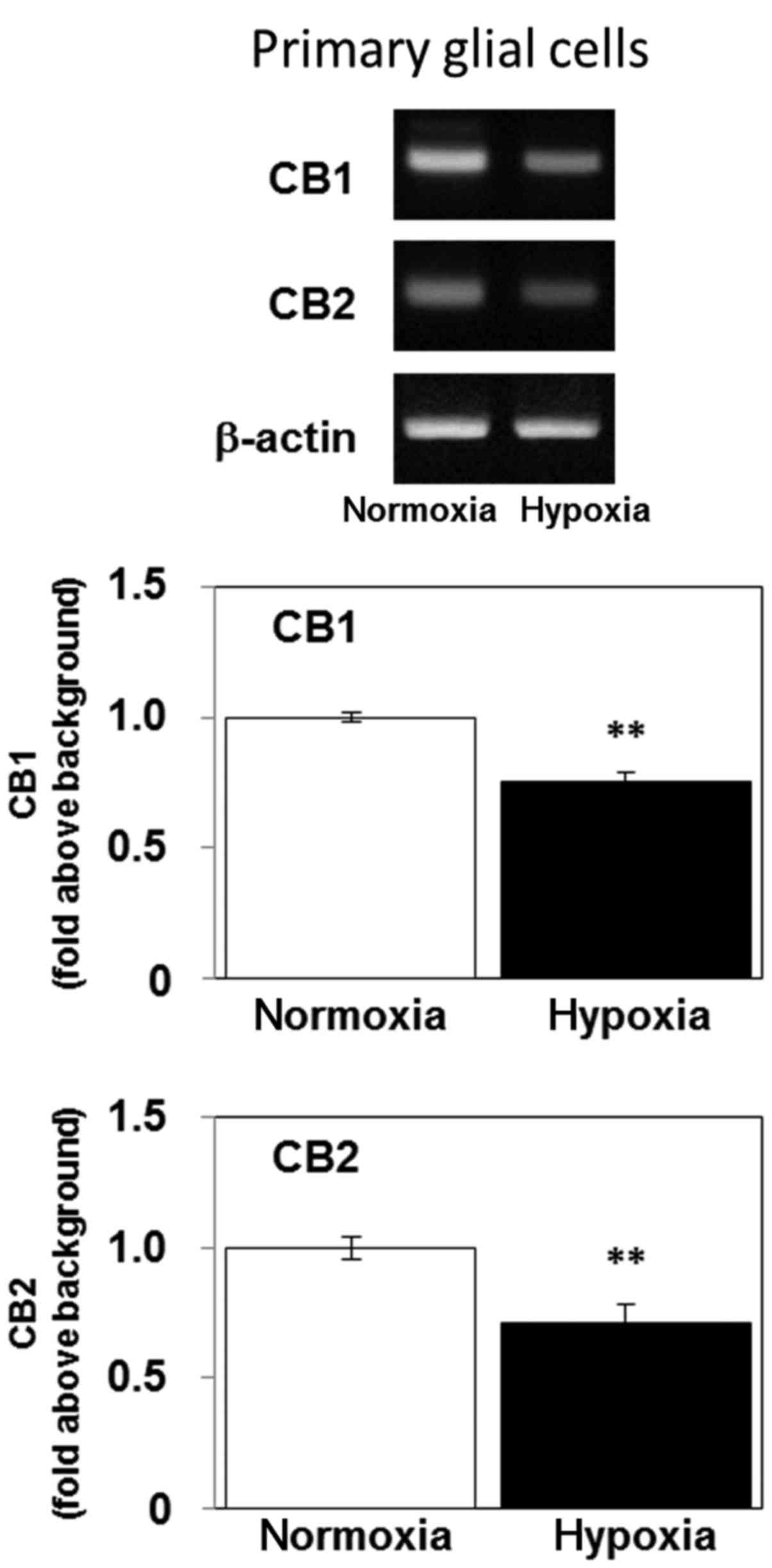
Hypoxia decreases the level of mRNA
encoding CB1 and CB2 in primary rat glial cells
We next examined the effects of hypoxia on the
expression levels of CB1 (Cnr1) and CB2 (Cnr2) mRNA
in primary rat glial cells. Continuous 5-day exposure of primary
glial cells to hypoxia decreased their expression of mRNA encoding
both CB1 and CB2 (Fig. 6). These
results indicate that hypoxia may regulate the expression levels of
cannabinoid receptors in normal glial cells as well as glioblastoma
cells.
Discussion
Several lines of evidence show that cannabinoids
influence a variety of physiological and pathological conditions
(2,8,32).
Notably, cannabinoids can inhibit the growth of tumors, including
glioblastoma, and have been proposed as potential antitumor agents
(7–9,14,33,34).
In contrast, hypoxia is a common feature of solid tumors, and has
been implicated in the persistence and development of malignancies
(16). Hypoxic regions are
frequently found in glioblastoma and hypoxia promotes a more
malignant phenotype (16). However,
the effects of hypoxia on the endocannabinoid system are not fully
understood. In the present study, we presented novel findings that
hypoxia downregulates mRNA encoding CB1, upregulates mRNA encoding
COX-2, and prevented cannabinoid receptor agonist-induced cell
death in U-87 MG cells. Our results suggest that hypoxia may
promote the progression of glioblastoma by inhibiting the
endocannabinoid system.
We found that the cannabinoid receptor agonist WIN
55,212-2-induced glioblastoma cell death in a dose-dependent
manner. We also observed preferential expression of mRNA encoding
CB1, in the absence of mRNA encoding CB2, in U-87 MG cells. We and
other investigators have previously found that cannabinoid receptor
agonists (WIN 55,212-2 and KM-233) suppress cell growth via CB1, as
antagonists for CB1, but not CB2, reversed cannabinoid receptor
agonist efficacy in U-87 MG cells (7,14). We
showed that hypoxia decreased the expression of mRNA encoding CB1
and inhibited WIN 55,212-2-induced cell death in U-87 MG cells,
indicating a CB1-dependent mechanism of action. These results
suggest that the activation of cannabinoid receptors in
glioblastoma is essential for inhibition of tumor progression.
Endocannabinoids are expressed in the brain (35), thus, it is possible that they
suppress developing brain tumors via activation of cannabinoid
receptors. Our present results suggest that hypoxia may counteract
the beneficial antitumor effects of endocannabinoids by
downregulating cannabinoid receptor expression in the brain.
Notably, several studies have indicated that
activation of CB2 plays a more dominant role in the inhibition of
glioma and glioblastoma progression than CB1 (36,37).
Although CB1 is predominantly expressed in the U-87 MG glioblastoma
cells that we used in the present study, many glioma and
glioblastoma cells express functional CB2 (38). To date, the effects of hypoxia on
the expression of CB2 in glioma and glioblastoma cells have not
been revealed. In the present study, we showed that hypoxia
inhibited the expression of CB2 in normal primary glial cells.
Further experiments are necessary to reveal whether hypoxia alters
the expression of CB2 in glioma and glioblastoma cells.
Hypoxia increased the expression levels of mRNA
encoding VEGF and COX-2, which are associated with the activation
of angiogenic and inflammatory responses, in U-87 MG cells.
Angiogenesis is necessary for tumor progression and inflammation
induces tumor cell survival, growth and migration (39). COX-2 is known to metabolize
endocannabinoids (20,21). Thus, upregulation of COX-2 may
inhibit endocannabinoid signaling by decreasing the concentration
of endocannabinoids. Anandamide and 2-arachidonoylglycerol are
major endocannabinoids that are produced, metabolized and kept at
constant concentrations in the brain (2,35).
However, hypoxia upregulates the expression of COX-2, which may
decrease the concentration of endocannabinoids in hypoxic regions
of glioblastoma, resulting in promotion of tumor growth. Thus,
hypoxia may facilitate the progression of glioblastoma by
inhibiting the endocannabinoid system in two ways: i)
downregulation of cannabinoid receptors; and ii) upregulation of
the endocannabinoid-metabolizing enzyme COX-2.
Notably, we detected lower GFAP mRNA
expression after 72 h of hypoxia, indicating dedifferentiation of
astrocytes. Moreover, 72 h exposure to hypoxia significantly failed
to induce cell death in the U-87 MG cells. Poorly differentiated
and undifferentiated carcinomas are characterized by highly
malignant tumors exhibiting aggressive invasive growth (40). We previously showed that 72 h
exposure to hypoxia inducedcell death in mouse fibroblast cells
(30). Thus, these results suggest
that U-87 MG cells, unlike mouse fibroblast cells, may have
increased hypoxia tolerance.
Hypoxia concomitantly downregulates the expression
of mRNA encoding GFAP and CB1, but the relationship between
dedifferentiation and endocannabinoid signaling is unknown. Further
experiments are necessary to reveal the relationship between
them.
Lamellipodia are formed by actin assembly at the
edge of a cell in the direction of migration. Phosphoinositide
3-kinase (PI3K) activation is known to induce lamellipodia
formation (31). Hypoxia activates
PI3K (41) and induces lamellipodia
formation, as shown Fig. 3, which
may facilitate tumor invasion.
Hypoxia inhibited the expression of mRNA encoding
CB1 in primary glial cells, as well as in U-87 MG cells, indicating
that hypoxia-induced downregulation of CB1 is common to normal
glial and malignant glioma cells. Although hypoxia-induced
inhibition of the endocannabinoid system is thought to promote
cancer progression, its role in normal glial cells has not been
revealed. Further experiments are necessary to elucidate the
effects of endocannabinoid inhibition mediated by hypoxia in
healthy glial cells.
In conclusion, the present study demonstrated that
hypoxia downregulates CB1 receptors, upregulates COX-2 and prevents
cannabinoid receptor agonist-induced cell death in U-87 MG cells
in vitro. These results suggest that hypoxia may promote
brain tumor progression by inhibiting the endocannabinoid
system.
Acknowledgements
The present study was supported by Grants-in-Aid for
Science and Culture (nos. 23500466, 25282021, 26430013, 26650173,
15KT0003, 16H005513 and 16K13013) from the Ministry of Education,
Culture, Sports, Science and Technology of Japan. This manuscript
has been edited and corrected by an experienced proofreader who is
a native speaker of English and who is under the direct supervision
of Honyaku Center Inc. (Tokyo, Japan).
Glossary
Abbreviations
Abbreviations:
|
CB
|
cannabinoid receptor
|
|
GFAP
|
glial fibrillary acidic protein
|
|
VEGF
|
vascular endothelial growth factor
|
|
COX-2
|
cyclooxygenase-2
|
References
|
1
|
Gui H, Tong Q, Qu W, Mao CM and Dai SM:
The endocannabinoid system and its therapeutic implications in
rheumatoid arthritis. Int Immunopharmacol. 26:86–91. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kano M, Ohno-Shosaku T, Hashimotodani Y,
Uchigashima M and Watanabe M: Endocannabinoid-mediated control of
synaptic transmission. Physiol Rev. 89:309–380. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ligresti A, De Petrocellis L and Di Marzo
V: From phytocannabinoids to cannabinoid receptors and
endocannabinoids: Pleiotropic physiological and pathological roles
through complex pharmacology. Physiol Rev. 96:1593–1659. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cabral GA and Griffin-Thomas L:
Cannabinoids as therapeutic agents for ablating neuroinflammatory
disease. Endocr Metab Immune Disord Drug Targets. 8:159–172. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cabral GA, Rogers TJ and Lichtman AH:
Turning over a new leaf: Cannabinoid and endocannabinoid modulation
of immune function. J Neuroimmune Pharmacol. 10:193–203. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Centonze D, Finazzi-Agrò A, Bernardi G and
Maccarrone M: The endocannabinoid system in targeting inflammatory
neurodegenerative diseases. Trends Pharmacol Sci. 28:180–187. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Echigo R, Sugimoto N, Yachie A and
Ohno-Shosaku T: Cannabinoids inhibit peptidoglycan-induced
phosphorylation of NF-κB and cell growth in U87MG human malignant
glioma cells. Oncol Rep. 28:1176–1180. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guindon J and Hohmann AG: The
endocannabinoid system and cancer: Therapeutic implication. Br J
Pharmacol. 163:1447–1463. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Salazar M, Carracedo A, Salanueva IJ,
Hernández-Tiedra S, Lorente M, Egia A, Vázquez P, Blázquez C,
Torres S, García S, et al: Cannabinoid action induces
autophagy-mediated cell death through stimulation of ER stress in
human glioma cells. J Clin Invest. 119:1359–1372. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Singh R, Singh B and Malhotra SK: A new
‘marker’ protein for astrocytes. Biosci Rep. 6:73–80. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dell'albani P, Rodolico M, Pellitteri R,
Tricarichi E, Torrisi SA, D'Antoni S, Zappia M, Albanese V,
Caltabiano R, Platania N, et al: Differential patterns of NOTCH1-4
receptor expression are markers of glioma cell differentiation.
Neuro Oncol. 16:204–216. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Burnet NG, Lynch AG, Jefferies SJ, Price
SJ, Jones PH, Antoun NM, Xuereb JH and Pohl U: High grade glioma:
Imaging combined with pathological grade defines management and
predicts prognosis. Radiother Oncol. 85:371–378. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Daumas-Duport C, Scheithauer B, O'Fallon J
and Kelly P: Grading of astrocytomas. A simple and reproducible
method. Cancer. 62:2152–2165. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gurley SN, Abidi AH, Allison P, Guan P,
Duntsch C, Robertson JH, Kosanke SD, Keir ST, Bigner DD, Elberger
AJ, et al: Mechanism of anti-glioma activity and in vivo efficacy
of the cannabinoid ligand KM-233. J Neurooncol. 110:163–177. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ortega A, Rangel-López E, Hidalgo-Miranda
A, Morales A, Ruiz-García E, Meneses-García A, Herrera-Gómez A,
Aguilar-Ponce JL, González-Herrera IG, Guevara-Salazar P, et al: On
the effects of CP 55–940 and other cannabinoid receptor agonists in
C6 and U373 cell lines. Toxicol In Vitro. 29:1941–1951. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang L, Lin C, Wang L, Guo H and Wang X:
Hypoxia and hypoxia-inducible factors in glioblastoma multiforme
progression and therapeutic implications. Exp Cell Res.
318:2417–2426. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Horiuchi A, Imai T, Shimizu M, Oka K, Wang
C, Nikaido T and Konishi I: Hypoxia-induced changes in the
expression of VEGF, HIF-1 alpha and cell cycle-related molecules in
ovarian cancer cells. Anticancer Res. 22:2697–2702. 2002.PubMed/NCBI
|
|
18
|
Bao B, Azmi AS, Ali S, Ahmad A, Li Y,
Banerjee S, Kong D and Sarkar FH: The biological kinship of hypoxia
with CSC and EMT and their relationship with deregulated expression
of miRNAs and tumor aggressiveness. Biochim Biophys Acta.
1826:272–296. 2012.PubMed/NCBI
|
|
19
|
Kawai N, Maeda Y, Kudomi N, Miyake K,
Okada M, Yamamoto Y, Nishiyama Y and Tamiya T: Correlation of
biological aggressiveness assessed by 11C-methionine PET
and hypoxic burden assessed by 18F-fluoromisonidazole
PET in newly diagnosed glioblastoma. Eur J Nucl Med Mol Imaging.
38:441–450. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Alhouayek M and Muccioli GG: COX-2-derived
endocannabinoid metabolites as novel inflammatory mediators. Trends
Pharmacol Sci. 35:284–292. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hermanson DJ, Gamble-George JC, Marnett LJ
and Patel S: Substrate-selective COX-2 inhibition as a novel
strategy for therapeutic endocannabinoid augmentation. Trends
Pharmacol Sci. 35:358–367. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
McCarthy KD and de Vellis J: Preparation
of separate astroglial and oligodendroglial cell cultures from rat
cerebral tissue. J Cell Biol. 85:890–902. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ohno-Shosaku T, Maejima T and Kano M:
Endogenous cannabinoids mediate retrograde signals from depolarized
postsynaptic neurons to presynaptic terminals. Neuron. 29:729–738.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sugimoto N, Leu H, Inoue N, Shimizu M,
Toma T, Kuroda M, Saito T, Wada T and Yachie A: The critical role
of lipopolysaccharide in the upregulation of aquaporin 4 in glial
cells treated with Shiga toxin. J Biomed Sci. 22:782015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Leu H, Sugimoto N, Shimizu M, Toma T, Wada
T, Ohta K and Yachie A: Tumor necrosis factor-α modifies the
effects of Shiga toxin on glial cells. Int Immunopharmacol.
38:139–143. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mongiardi MP: Angiogenesis and hypoxia in
glioblastoma: A focus on cancer stem cells. CNS Neurol Disord Drug
Targets. 11:878–883. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang S, Liu Z, Wang L and Zhang X:
NF-kappaB signaling pathway, inflammation and colorectal cancer.
Cell Mol Immunol. 6:327–334. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Friedmann-Morvinski D: Glioblastoma
heterogeneity and cancer cell plasticity. Crit Rev Oncog.
19:327–336. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sowers JL, Johnson KM, Conrad C, Patterson
JT and Sowers LC: The role of inflammation in brain cancer. Adv Exp
Med Biol. 816:75–105. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sugimoto N, Shido O, Matsuzaki K, Katakura
M, Hitomi Y, Tanaka M, Sawaki T, Fujita Y, Kawanami T, Masaki Y, et
al: Long-term heat exposure prevents hypoxia-induced apoptosis in
mouse fibroblast cells. Cell Biochem Biophys. 70:301–307. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sugimoto N, Takuwa N, Yoshioka K and
Takuwa Y: Rho-dependent, Rho kinase-independent inhibitory
regulation of Rac and cell migration by LPA1 receptor in
Gi-inactivated CHO cells. Exp Cell Res. 312:1899–1908. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Klein TW: Cannabinoid-based drugs as
anti-inflammatory therapeutics. Nat Rev Immunol. 5:400–411. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Guzmán M: Cannabinoids: Potential
anticancer agents. Nat Rev Cancer. 3:745–755. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sarfaraz S, Adhami VM, Syed DN, Afaq F and
Mukhtar H: Cannabinoids for cancer treatment: Progress and promise.
Cancer Res. 68:339–342. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zoerner AA, Gutzki FM, Batkai S, May M,
Rakers C, Engeli S, Jordan J and Tsikas D: Quantification of
endocannabinoids in biological systems by chromatography and mass
spectrometry: A comprehensive review from an analytical and
biological perspective. Biochim Biophys Acta. 1811:706–723. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Blázquez C, Salazar M, Carracedo A,
Lorente M, Egia A, González-Feria L, Haro A, Velasco G and Guzmán
M: Cannabinoids inhibit glioma cell invasion by down-regulating
matrix metalloproteinase-2 expression. Cancer Res. 68:1945–1952.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rocha FC, Dos Santos Júnior JG, Stefano SC
and da Silveira DX: Systematic review of the literature on clinical
and experimental trials on the antitumor effects of cannabinoids in
gliomas. J Neurooncol. 116:11–24. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
De Jesús ML, Hostalot C, Garibi JM, Sallés
J, Meana JJ and Callado LF: Opposite changes in cannabinoid CB1 and
CB2 receptor expression in human gliomas. Neurochem Int.
56:829–833. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ono M: Molecular links between tumor
angiogenesis and inflammation: Inflammatory stimuli of macrophages
and cancer cells as targets for therapeutic strategy. Cancer Sci.
99:1501–1506. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dai C and Holland EC: Astrocyte
differentiation states and glioma formation. Cancer J. 9:72–81.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Muz B, de la Puente P, Azab F and Azab AK:
The role of hypoxia in cancer progression, angiogenesis,
metastasis, and resistance to therapy. Hypoxia. 3:83–92. 2015.
View Article : Google Scholar : PubMed/NCBI
|