Introduction
Bladder cancer is one of the most common cancers in
the world, with an estimated 174,000 deaths in 2013 (1). For patients with locally advanced or
metastatic bladder cancer, the standard chemotherapy is
cisplatin-based combination chemotherapy. However, the 5-year
survival rate in patients with stage IV bladder cancer after
treatment is only around 15% (2).
Considering the limited response rate of the regimens and the
toxicity (3), novel therapeutic
agents to replace conventional cytotoxic chemotherapy are
needed.
It has been reported that genetic abnormality of
various genes is observed in patients with bladder cancer.
Recently, it has become clear that the signaling pathways of
fibroblast growth factor receptors (FGFRs), particularly the FGFR3
(4) and FGFR1 (5) pathways, are altered in bladder
cancers. FGFRs (FGFR1-FGFR4) play a key role in the regulation of
proliferation, angiogenesis and apoptosis (6). The binding of FGFs to FGFRs initiates
a phosphorylation cascade, resulting in signaling through various
downstream pathways, including RAS-MAPK, PI3K and STATs (7). Active mutations and/or overexpression
of FGFR3 are common genetic alterations in non-invasive bladder
cancer, but occur at a low frequency in high-grade invasive bladder
cancer (8). FGFR1 is not mutated in
bladder cancer, but overexpression is frequent in bladder cancer
cell lines regardless of tumor grade and stage (5). In addition, it has been reported that
FGFR1 expression is an adverse prognostic risk factor in muscle
invasive bladder cancer after radical cystectomy (9). Recently, a number of clinical trials
of BGJ398, a selective pan FGFR inhibitor, have progressed in
patients with various types of cancers. In a phase 1 clinical trial
using BGJ398 in patients with solid tumors carrying FGFR genetic
alternations, antitumor activity was demonstrated in patients with
FGFR3-mutant bladder cancer (10).
Furthermore, a study of BGJ398 in non-muscle-invasive urothelial
carcinoma of the bladder is currently recruiting participants. FGFR
inhibitor BGJ398 is expected to be a more effective therapeutic
agent for patients with bladder cancer. However, it has been
reported that BGJ398 did not suppress primary tumors, but did
inhibit the development of circulating tumor cells and metastasis
in vivo (11).
Histone deacetylase (HDAC) inhibitors are also
predicted for the treatment of various cancers including bladder
cancer. It has been reported that HDAC inhibitors inhibit cell
growth and induce cell death in bladder cancer cells (12). Furthermore, several studies have
reported that HDAC inhibitors synergize with chemotherapeutic
agents (13–15). Currently, a phase 2 clinical trial
of an HDAC inhibitor for patients with bladder cancer is ongoing.
OBP-801/YM753/spiruchostatin A was identified as a novel HDAC
inhibitor by using a p21 promoter-reporter assay, which showed the
most potent HDAC-inhibitory activity as compared to other HDAC
inhibitors (16).
We showed for the first time that a combined
treatment of FGFR inhibitor BGJ398 and HDAC inhibitor OBP-801
synergistically inhibited cell growth, and induced apoptosis in
human bladder cancer cells. Taken together, we suggest that the
combination is a promising novel therapeutic approach to treat
muscle-invasive bladder cancer.
Materials and methods
Cell culture
Human bladder cancer UM-UC-3 and T24 cells were
purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA) and cultured in RPMI-1640 medium supplemented
with 10% fetal bovine serum (FBS), 2 mM L-glutamine, 100 U/ml
penicillin and 100 µg/ml streptomycin. Cell cultures were incubated
at 37°C in a humidified atmosphere of 5% CO2.
Reagents
BGJ398 was purchased from Selleck Chemicals
(Houston, TX, USA). OBP-801 was supplied by Oncolys BioPharma
(Tokyo, Japan). zVAD-fmk was purchased from R&D Systems
(Minneapolis, MN, USA). BGJ398, OBP-801 and zVAD-fmk were dissolved
in dimethyl sulfoxide (DMSO).
Cell viability assay
The number of viable cells was determined using the
Cell Counting Kit-8 (CCK-8) assay according to the manufacturer's
instructions (Dojindo Laboratories, Kumamoto, Japan). After the
incubation of cells (3×103/well) in 96-well plates for
72 h with the indicated concentrations of BGJ398 or OBP-801, the
kit reagent was added to the medium, and the plates were incubated
for a further 4 h. The absorbance of samples (450 nm) was
determined using a scanning multiwell spectrophotometer (DS Pharma
Biomedical, Co., Ltd., Osaka, Japan).
Combination index
Combination index (CI) values were analyzed using
CalcuSyn software (Biosoft, Great Shelford, UK). CI <1 indicates
synergism greater than the expected additive effect.
Detection of apoptosis
After the incubation of cells
(2×104/well) in 6-well plates for 72 h with the agents,
the cells were harvested. After washing with phosphate-buffered
saline (PBS), the cells were treated with PBS containing 0.1%
Triton X-100 and the nuclei were stained with propidium iodide (PI;
Sigma-Aldrich, St. Louis, MO, USA). The DNA content was measured
using a FACSCalibur (Becton-Dickinson, Franklin Lakes, NJ, USA).
CellQuest software (Becton-Dickinson) was used to analyze the data.
DNA fragmentation was quantified by the percentage of hypodiploid
DNA (sub-G1).
Western blotting
Western blot analysis was carried out as previously
described (17). The following
antibodies were purchased from the indicated sources: rabbit
monoclonal antibodies for anti-Bim (ab32158) (Abcam, Cambridge,
UK), anti-Bcl-xL (#2762), and anti-PARP (#9542) (Cell Signaling
Technology, Beverly, MA, USA); rabbit polyclonal antibodies for
anti-survivin (AF886) (R&D Systems), anti-DR5 (#8074) (Cell
Signaling Technology), anti-DR4 (1139) (ProSci, Inc., Poway, CA,
USA), anti-caspase-3 (#9665), anti-histone H4 (#2935),
anti-acetyl-histone H4 (#9672), anti-Bid (#2002), anti-ERK1/2
(#9102), anti-Shp2 (#3752), anti-phospho ERK1/2 (#9101),
anti-phospho FRS2-α (#3861), anti-phospho Shp2 (#3751) (Cell
Signaling Technology), anti-FRS2-α (SC17841) (Santa Cruz
Biotechnology, Dallas, TX, USA); mouse monoclonal anti-bodies for
anti-caspase-8 (M032-3) and anti-caspase-9 (M054-3) (MBL, Nagoya,
Japan), anti-FLIP (ACX-804-961-0100) (Enzo Life Sciences,
Farmingdale, NY, USA), and anti-GAPDH (5G4) (HyTest, Ltd., Turku,
Finland) were used as primary antibodies. An anti-rabbit
IgG-HRP-conjugated antibody (NA934) (GE Healthcare Life Sciences,
Little Chalfont, UK) and an anti-mouse IgG-HRP-conjugated antibody
(NA931) (GE Healthcare Life Sciences) were used as the secondary
antibodies. The signal was detected using the Chemilumi-One
chemiluminescent kit (Nacalai Tesque, Inc., Kyoto, Japan) or
chemiluminescent HRP substrate (Millipore, Billerica, MA, USA).
Small interfering RNA
transfection
The Bim siRNA and the negative control siRNA were
purchased from Sigma-Aldrich. The Bim siRNA (ACUUACAUCAGAAGGUUGC)
was used for the transfection. At the same time as the
transfection, UM-UC-3 and T24 cells (2×104/well) were
seeded in 6-well plates without antibiotics. The Bim siRNA (5 nM
for UM-UC-3 cells or 10 nM for T24 cells) was transfected into
cells using Lipofectamine RNAiMAX (Invitrogen, Carlsbad, CA, USA)
according to the manufacturer's instructions. Twenty-four hours
after the transfection, the cells were treated with agents for 48 h
and then harvested.
Statistical analysis
Data are expressed as the means ± SD of three
determinations. Statistical analysis was performed using a Students
t-test. Samples were considered significantly different at
P<0.05.
Results
Combined treatment with BGJ398 and
OBP-801 synergistically inhibits cell growth in human bladder
cancer cells
To examine the anti-growth effects of BGJ398 or
OBP-801 alone against human bladder cancer cells, we first assessed
the viable cell number at 72 h after treatment with the indicated
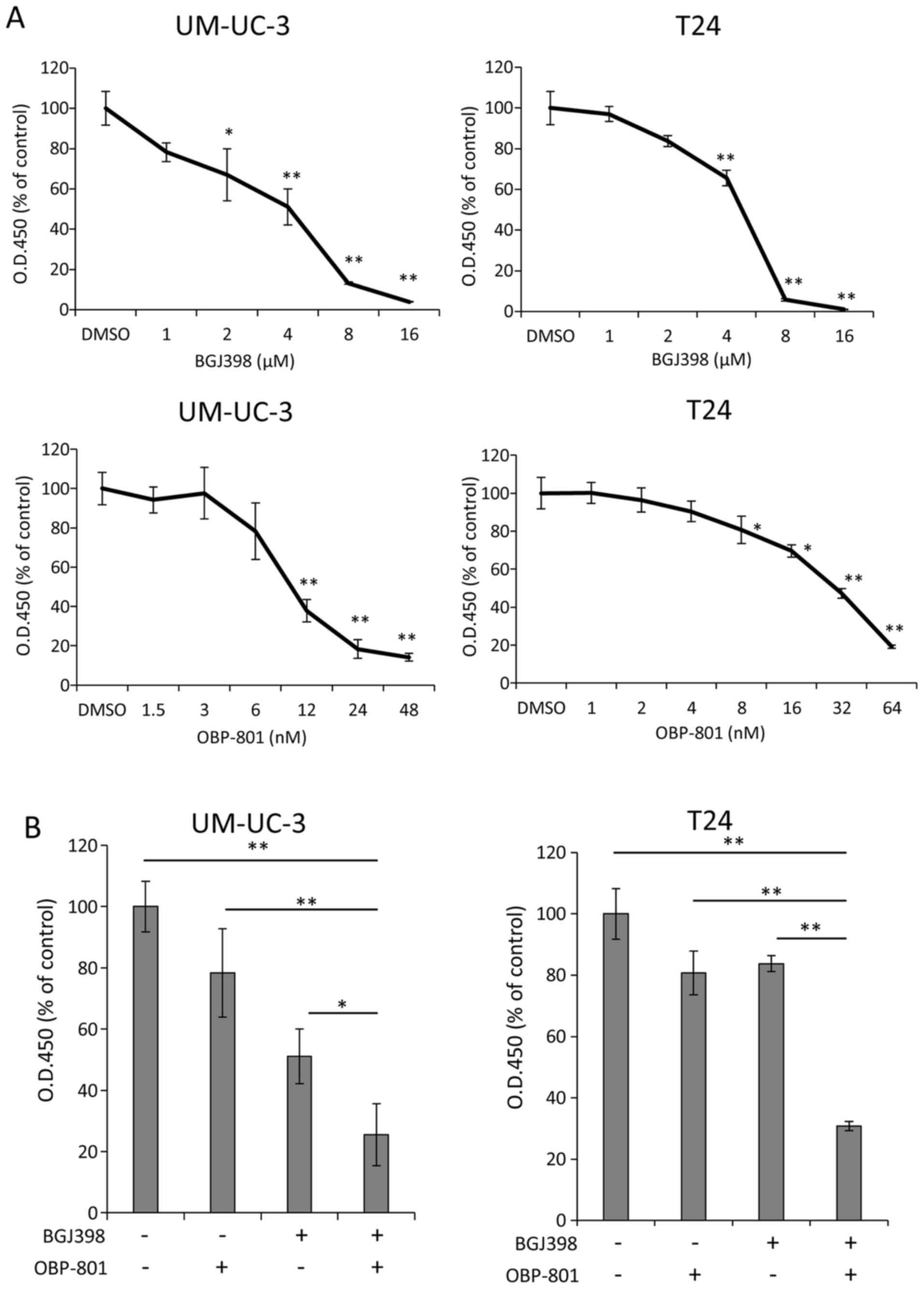
concentrations of the agents. Each agent suppressed the cell growth
of UM-UC-3 and T24 cells in a dose-dependent manner (Fig. 1A). Notably, co-treatment with BGJ398
and OBP-801 markedly inhibited cell growth, compared with the
single treatment of each agent in both cell lines (Fig. 1B). Moreover, the combination index
(CI) values for BGJ398 and OBP-801 were <1.0, indicating a
synergistic effect on the inhibition of cell growth (Table I).
 | Table I.The combination index of BGJ398 and
OBP-801 was calculated in the UM-UC-3 and T24 cells. |
Table I.
The combination index of BGJ398 and
OBP-801 was calculated in the UM-UC-3 and T24 cells.
| BGJ398 (µM) | OBP-801 (nM) | Combination
index |
|---|
| UM-UC-3 | UM-UC-3 |
|
| 2 | 3 | 2.098 |
| 4 | 6 | 0.669 |
| 8 | 12 | 0.216 |
| T24 | T24 |
|
| 1 | 4 | 1.030 |
| 2 | 8 | 0.591 |
| 4 | 16 | 0.259 |
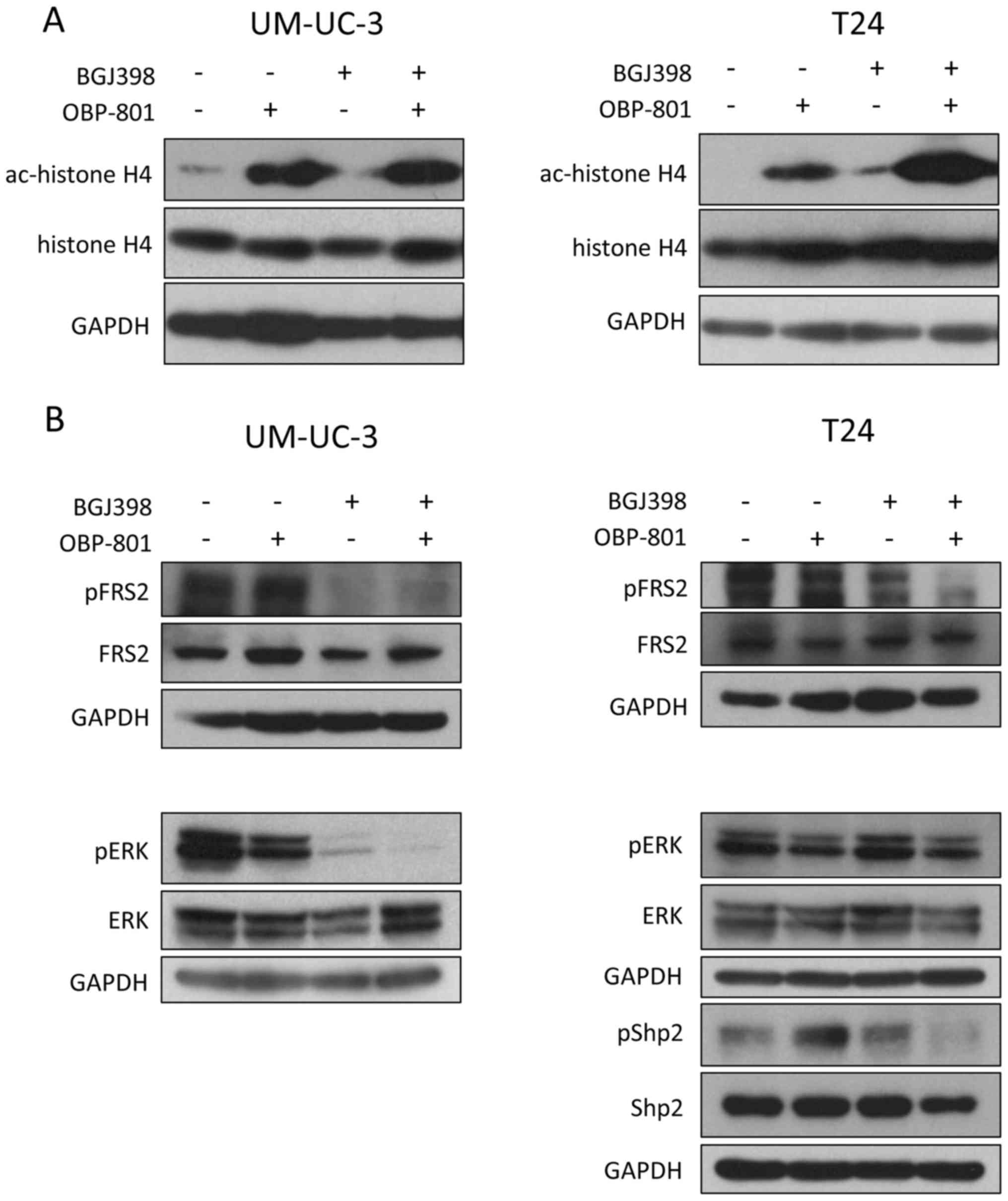
We next examined the effects of the combined
treatment of FGFR inhibitor BGJ398 and HDAC inhibitor OBP-801 on
the FGFR signaling pathways and HDAC-inhibitory activity. OBP-801
alone and the combination increased the acetylation of histone H4
in both UM-UC-3 and T24 cells (Fig.
2A). BGJ398 alone and the combination inhibited the
phosphorylation of FGFR substrate 2 (FRS2), which is a docking
protein linking FGFRs, in UM-UC-3 cells. Furthermore, BGJ398 also
inhibited the phosphorylation of ERK in UM-UC-3 cells (Fig. 2B). On the other hand, BGJ398
slightly inhibited phosphorylation of FRS2, and the co-treatment of
OBP-801 and BGJ398 more clearly inhibited it in T24 cells. However,
the treatment of BGJ398 alone and the combination did not inhibit
the phosphorylation of ERK in T24 cells (Fig. 2B). Therefore, we next examined the
effect of the combination on the phosphorylation of protein
tyrosine phosphatase Shp2. Shp2 is a critical downstream mediator
of FGFR-FRS2 signaling. As shown in Fig. 2B, the combined treatment inhibited
the phosphorylation of Shp2 in T24 cells.
Combined treatment with BGJ398 and
OBP-801 induces caspase-dependent apoptosis in human bladder cancer
cells
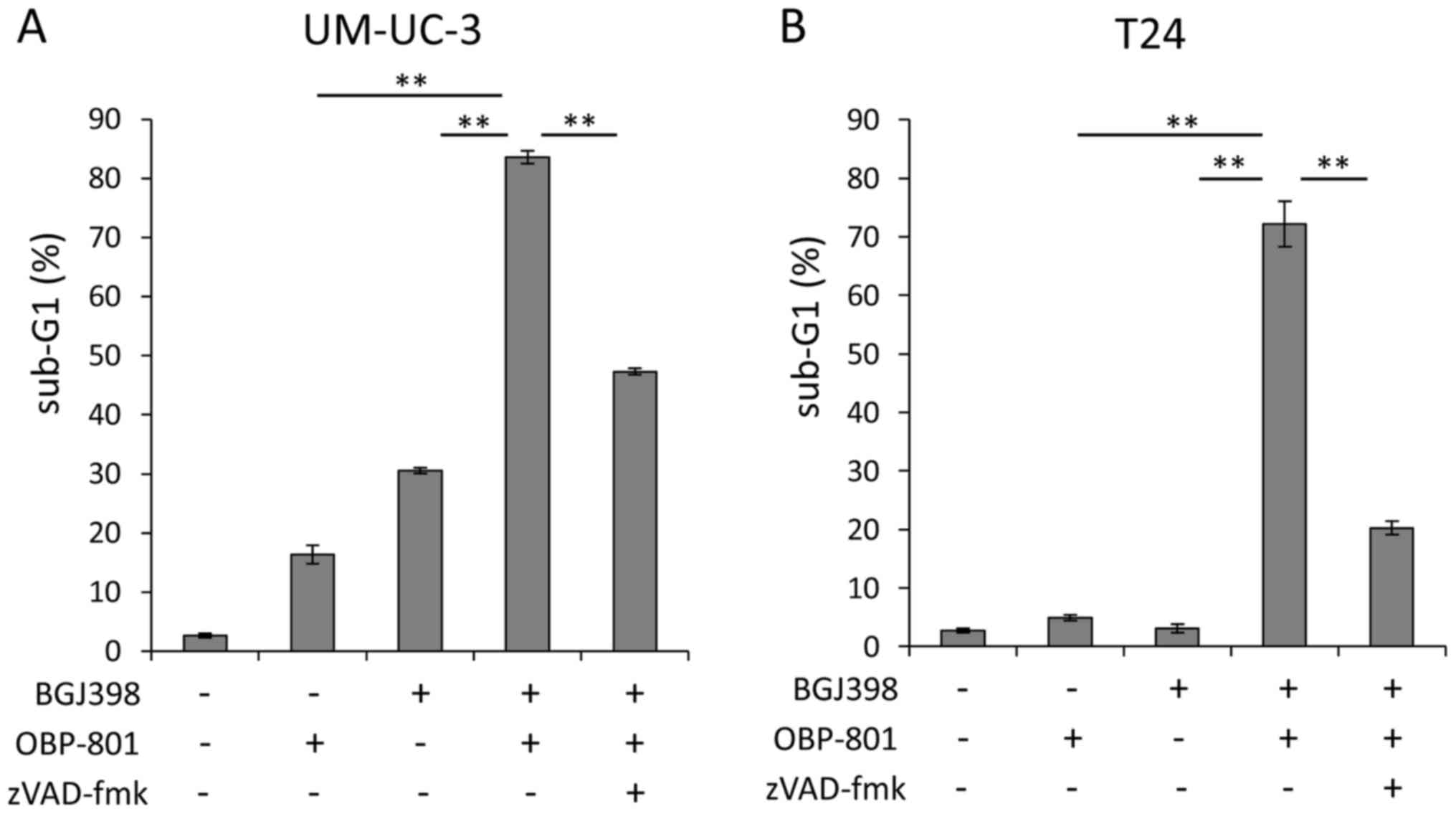
To clarify the mechanisms of the synergistic
inhibitory effects on cell growth of bladder cancer cells by the
combined treatment with BGJ398 and OBP-801, we next investigated
the effect of the combination on apoptosis by measuring the sub-G1
populations, using flow cytometry after treatment for 72 h. The
co-treatment with BGJ398 and OBP-801 more markedly induced
apoptosis than that of each agent alone in the UM-UC-3 and T24
cells (Fig. 3). To examine whether
the apoptosis induced by the combination of the agents is
caspase-dependent, we analyzed the effect of a caspase inhibitor.
The pan-caspase inhibitor zVAD-fmk effectively blocked apoptosis
induced by the co-treatment with BGJ398 and OBP-801 (Fig. 3). These results suggest that the
combination of BGJ398 and OBP-801 synergistically induces
caspase-dependent apoptosis in human bladder cancer cells.
Combined treatment with BGJ398 and
OBP-801 causes apoptosis via intrinsic and extrinsic pathways in
human bladder cancer cells
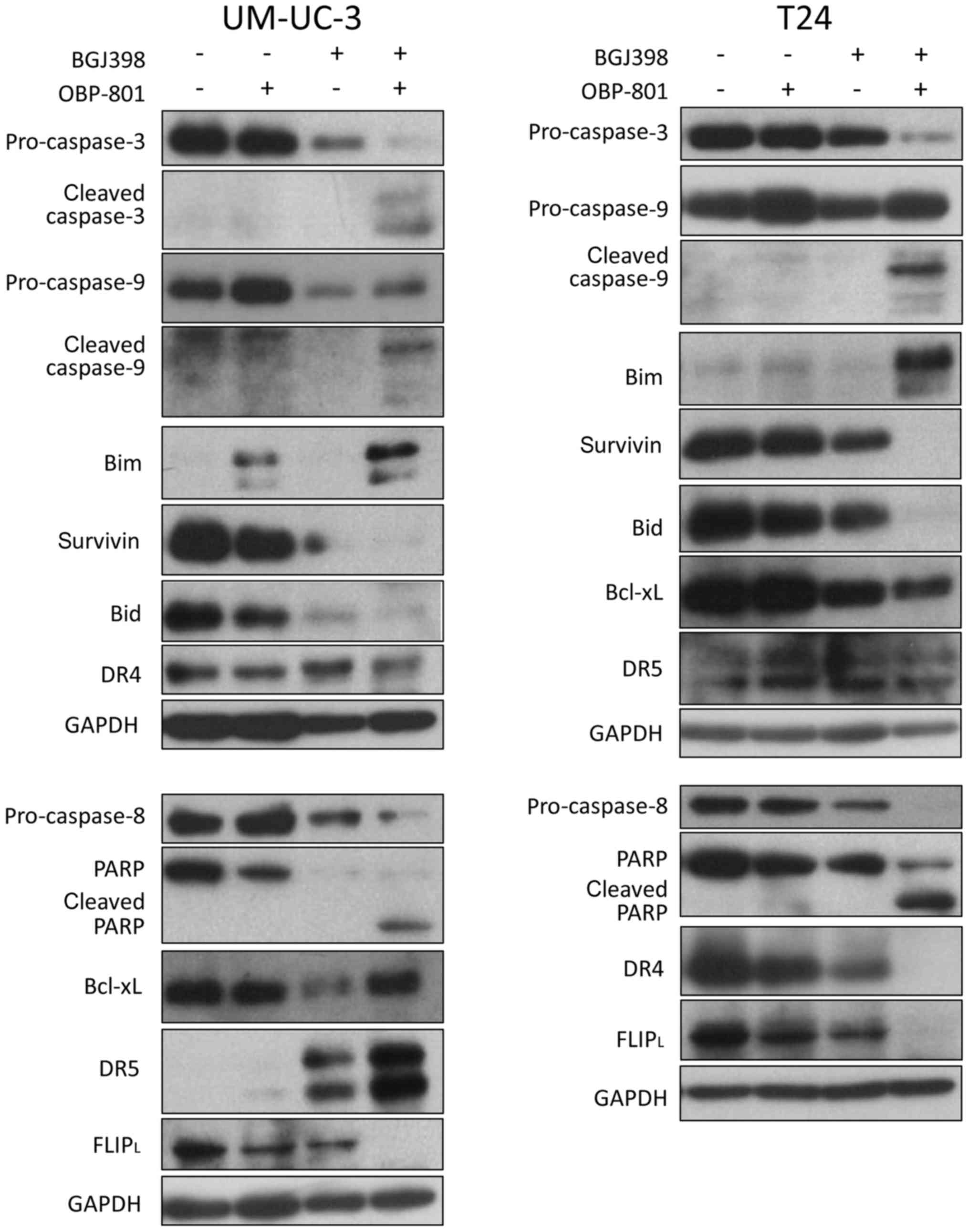
We examined the expression of apoptosis-related
proteins in the combined treatment. As shown in Fig. 4, the concurrent treatment induced
the cleavage of caspase-8, caspase-9, caspase-3 and PARP. The
co-treatment increased the expression of pro-apoptotic protein Bim,
and reduced the expression of anti-apoptotic proteins, survivin and
FLIPL, in UM-UC-3 and T24 cells (Fig. 4). The expression of DR5, one of the
pro-apoptotic proteins, was upregulated by co-treatment in UM-UC-3
cells, but not in T24 cells. These results suggest that the
combined treatment with BGJ398 and OBP-801 induced apoptosis
through the activation of both intrinsic and extrinsic pathways in
human bladder cancer cells.
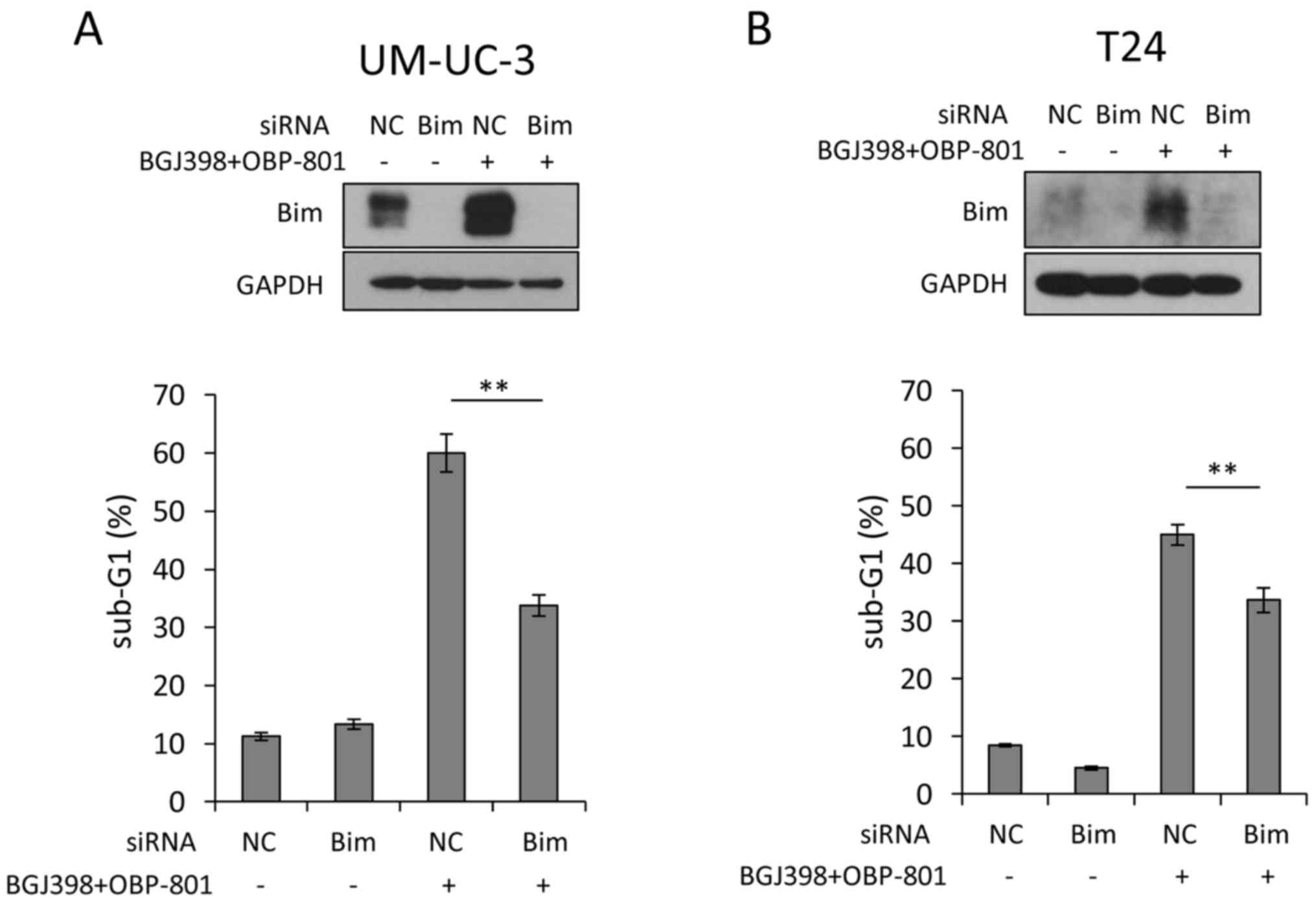
Bim contributes to apoptosis by the
combined treatment in bladder cancer cells
The combined treatment upregulated the expression of
Bim in the UM-UC-3 and T24 cells (Fig.
4). We examined whether Bim contributed to the induction of
apoptosis by co-treatment with BGJ398 and OBP-801. The effects of
the Bim knockdown were confirmed by western blotting (Fig. 5). Bim siRNA significantly suppressed
apoptosis induced by the combination compared with the control
(Fig. 5). These results suggest
that the combined treatment with BGJ398 and OBP-801 caused
apoptosis at least partially through the upregulation of Bim in
bladder cancer cells.
Discussion
In the present study, we showed for the first time
the synergistic effect of the combined treatment with FGFR
inhibitor BGJ398 and the novel HDAC inhibitor OBP-801 on cell
growth arrest and apoptosis against high grade bladder cancer
cells. Furthermore, the apoptosis was associated with the
upregulation of Bim and DR5, and downregulation of survivin and
FLIP by the combined treatment as the molecular mechanisms. In
these apoptosis-related proteins, we showed that Bim knockdown
significantly inhibited apoptosis induced by the combination
(Fig. 5). The results suggest that
the combined treatment causes apoptosis at least partially through
the upregulation of Bim in bladder cancer cells.
Bim is a BH3-only pro-apoptotic member of the Bcl-2
family (18), and its expression is
regulated by ERK1/2 and PI3K/AKT pathways (19). The activation of the ERK1/2 pathway
induces the phosphorylation of Bim and promotes
proteasome-dependent degradation of phosphorylated Bim (20). In the present study, OBP-801
slightly inhibited the phosphorylation of ERK1/2 in UM-UC-3 cells
(Fig. 2B). A previous report showed
that HDAC inhibitors decreased phosphorylation of ERK by
upregulating dual-specificity phosphatase (DUSP) (21). DUSP may be involved in the
downregulation of ERK1/2 by OBP-801 in UM-UC-3 cells. On the other
hand, the effect of BGJ398 was different in the UM-UC-3 and in T24
cells. BGJ398 clearly inhibited the phosphorylation of ERK1/2 in
UM-UC-3 cells (Fig. 2B). The
phosphorylation of ERK1/2 was more clearly inhibited by the
co-treatment of BGJ398 and OBP-801 in UM-UC-3 cells (Fig. 2B). These results suggest that the
expression of Bim was upregulated by the combination in
ERK1/2-dependent pathways in UM-UC-3 cells.
In the present study, BGJ398 alone and co-treatment
with OBP-801 inhibited the phosphorylation of FRS2 in T24 cells
(Fig. 2B). On the other hand, the
phosphorylation of ERK was not suppressed by these treatments in
T24 cells (Fig. 2B). Therefore, we
next examined whether the combination regulated Shp2 downstream of
FGFR-FRS2 signaling independent of ERK. Shp2, a non-receptor
phosphotyrosine phosphatase, promotes tumor progression in various
cancer types (22). Furthermore,
previous studies reported that Shp2 is a key downregulator of Bim
through ERK1/2 activation (23,24).
Notably, in this study, the combination inhibited phosphorylation
of Shp2 but not the phosphorylation of ERK in T24 cells. These data
showed that Bim was upregulated by Shp2 independent of ERK in T24
cells treated by the combination, suggesting the existence of an
unknown pathway between Shp2 and Bim independent of ERK.
Moreover, the expression of DR5 was upregulated by
the combination in UM-UC-3 cells (Fig.
4). We previously reported that OBP-801 and celecoxib
synergistically induced apoptosis via the DR5-dependent pathway,
and that Bim partially acts as one of the key molecules downstream
of DR5 in bladder cancer cells (17). Therefore, the DR5-dependent pathway
may be partially involved in the caspase-dependent apoptosis by the
combination of BGJ398 and OBP-801 in UM-UC-3 cells.
In conclusion, this is the first report showing the
synergistic effect of the combination of an FGFR inhibitor and an
HDAC inhibitor against human bladder cancer cells. We suggest that
the combined treatment is a promising novel therapeutic approach to
treat muscle-invasive bladder cancer.
Acknowledgements
The present study was supported by the Research
Funding of Kyoto Prefectural University of Medicine.
Glossary
Abbreviations
Abbreviations:
|
FGFR
|
fibroblast growth factor receptor
|
|
HDAC
|
histone deacetylase
|
|
MAPK
|
mitogen-activated protein kinase
|
|
PI3K
|
phosphoinositide 3-kinase
|
|
ERK
|
extracellular signal-related
kinase
|
|
CI
|
combination index
|
|
FRS2
|
fibroblast growth factor receptor
substrate 2
|
References
|
1
|
Fitzmaurice C, Dicker D, Pain A, Hamavid
H, Moradi-Lakeh M, MacIntyre MF, Allen C, Hansen G, Woodbrook R,
Wolfe C, et al Global Burden of Disease Cancer Collaboration, : The
Global Burden of Cancer 2013. JAMA Oncol. 1:505–527. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
von der Maase H, Sengelov L, Roberts JT,
Ricci S, Dogliotti L, Oliver T, Moore MJ, Zimmermann A and Arning
M: Long-term survival results of a randomized trial comparing
gemcitabine plus cisplatin, with methotrexate, vinblastine,
doxorubicin, plus cisplatin in patients with bladder cancer. J Clin
Oncol. 23:4602–4608. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Drayton RM and Catto JW: Molecular
mechanisms of cisplatin resistance in bladder cancer. Expert Rev
Anticancer Ther. 12:271–281. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tomlinson DC, Baldo O, Harnden P and
Knowles MA: FGFR3 protein expression and its relationship to
mutation status and prognostic variables in bladder cancer. J
Pathol. 213:91–98. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tomlinson DC, Lamont FR, Shnyder SD and
Knowles MA: Fibroblast growth factor receptor 1 promotes
proliferation and survival via activation of the mitogen-activated
protein kinase pathway in bladder cancer. Cancer Res. 69:4613–4620.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Turner N and Grose R: Fibroblast growth
factor signalling: From development to cancer. Nat Rev Cancer.
10:116–129. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Klint P and Claesson-Welsh L: Signal
transduction by fibroblast growth factor receptors. Front Biosci.
4:D165–D177. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cancer Genome Atlas Research Network, .
Comprehensive molecular characterization of urothelial bladder
carcinoma. Nature. 507:315–322. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lim S, Koh MJ, Jeong HJ, Cho NH, Choi YD,
Cho Y, Lee HY and Rha SY: Fibroblast growth factor receptor 1
overexpression is associated with poor survival in patients with
resected muscle invasive urothelial carcinoma. Yonsei Med J.
57:831–839. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nogova L, Sequist LV, Perez Garcia JM,
Andre F, Delord JP, Hidalgo M, Schellens JH, Cassier PA, Camidge
DR, Schuler M, et al: Evaluation of BGJ398, a fibroblast growth
factor receptor 1–3 kinase inhibitor, in patients with advanced
solid tumors harboring genetic alterations in fibroblast growth
factor receptors: Results of a global phase I, dose-escalation and
dose-expansion study. J Clin Oncol. 35:157–165. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cheng T, Roth B, Choi W, Black PC, Dinney
C and McConkey DJ: Fibroblast growth factor receptors-1 and −3 play
distinct roles in the regulation of bladder cancer growth and
metastasis: Implications for therapeutic targeting. PLoS One.
8:e572842013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li QQ, Hao JJ, Zhang Z, Hsu I, Liu Y, Tao
Z, Lewi K, Metwalli AR and Agarwal PK: Histone deacetylase
inhibitor-induced cell death in bladder cancer is associated with
chromatin modification and modifying protein expression: A
proteomic approach. Int J Oncol. 48:2591–2607. 2016.PubMed/NCBI
|
|
13
|
Jeon HG, Yoon CY, Yu JH, Park MJ, Lee JE,
Jeong SJ, Hong SK, Byun SS and Lee SE: Induction of caspase
mediated apoptosis and down-regulation of nuclear factor-κB and Akt
signaling are involved in the synergistic antitumor effect of
gemcitabine and the histone deacetylase inhibitor trichostatin A in
human bladder cancer cells. J Urol. 186:2084–2093. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang D, Jing Y, Ouyang S, Liu B, Zhu T,
Niu H and Tian Y: Inhibitory effect of valproic acid on bladder
cancer in combination with chemotherapeutic agents in vitro and in
vivo. Oncol Lett. 6:1492–1498. 2013.PubMed/NCBI
|
|
15
|
Li DR, Zhang H, Peek E, Wang S, Du L, Li G
and Chin AI: Synergy of histone-deacetylase inhibitor AR-42 with
cisplatin in bladder cancer. J Urol. 194:547–555. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shindoh N, Mori M, Terada Y, Oda K, Amino
N, Kita A, Taniguchi M, Sohda KY, Nagai K, Sowa Y, et al: YM753, a
novel histone deacetylase inhibitor, exhibits antitumor activity
with selective, sustained accumulation of acetylated histones in
tumors in the WiDr xenograft model. Int J Oncol. 32:545–555.
2008.PubMed/NCBI
|
|
17
|
Toriyama S, Horinaka M, Yasuda S,
Taniguchi T, Aono Y, Takamura T, Morioka Y, Miki T, Ukimura O and
Sakai T: A histone deacetylase inhibitor, OBP-801, and celecoxib
synergistically inhibit the cell growth with apoptosis via a
DR5-dependent pathway in bladder cancer cells. Mol Cancer Ther.
15:2066–2075. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Youle RJ and Strasser A: The BCL-2 protein
family: Opposing activities that mediate cell death. Nat Rev Mol
Cell Biol. 9:47–59. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Balmanno K and Cook SJ: Tumour cell
survival signalling by the ERK1/2 pathway. Cell Death Differ.
16:368–377. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ley R, Balmanno K, Hadfield K, Weston C
and Cook SJ: Activation of the ERK1/2 signaling pathway promotes
phosphorylation and proteasome-dependent degradation of the
BH3-only protein, Bim. J Biol Chem. 278:18811–18816. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ferguson BS, Harrison BC, Jeong MY, Reid
BG, Wempe MF, Wagner FF, Holson EB and McKinsey TA:
Signal-dependent repression of DUSP5 by class I HDACs controls
nuclear ERK activity and cardiomyocyte hypertrophy. Proc Natl Acad
Sci USA. 110:pp. 9806–9811. 2013; View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang J, Zhang F and Niu R: Functions of
Shp2 in cancer. J Cell Mol Med. 19:2075–2083. 2015.PubMed/NCBI
|
|
23
|
Yang W, Klaman LD, Chen B, Araki T, Harada
H, Thomas SM, George EL and Neel BG: An Shp2/SFK/Ras/Erk signaling
pathway controls trophoblast stem cell survival. Dev Cell.
10:317–327. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Selimoglu-Buet D, Gallais I, Denis N,
Guillouf C and Moreau-Gachelin F: Oncogenic kit triggers
Shp2/Erk1/2 pathway to down-regulate the pro-apoptotic protein Bim
and to promote apoptosis resistance in leukemic cells. PLoS One.
7:e490522012. View Article : Google Scholar : PubMed/NCBI
|