Introduction
Ovarian cancer is the leading cause of death among
women (1). Currently, the main
treatment for ovarian cancer is surgical excision followed by
combination chemotherapy. Cisplatin (CDDP) is the most effective
chemotherapy drug for the treatment of ovarian cancer (2). However, CDDP chemoresistance remains a
large obstacle for the successful treatment of ovarian cancer
(3). Therefore, it is important to
explore the molecular basis of CDDP resistance to improve the
success rate of chemotherapy.
MicroRNAs (miRNAs) are noncoding, single stranded
RNAs that regulate gene expression by binding the 3′-untranslated
regions (3′-UTRs) of target mRNAs (4). Ovarian cancer patients who were
resistant to chemotherapy frequently had different miRNA expression
patterns compared to those of the sensitive patients (5). For example, microarray identified that
let-7i was the first miRNA associated with CDDP resistance of
ovarian cancer (6). Moreover,
miR-595 (7), miR-186 (8) and miR-141 (9) increased the sensitivity of ovarian
cancer cells to CDDP by targeting multiple signaling pathways,
whereas miR-130a and miR-374a contribute to CDDP resistance in
ovarian cancer (10). However, the
role and mechanism of miRNA involved in CDDP resistance in ovarian
cancer remains largely unknown.
In the present study, we explored the expression
pattern and biological effects of miR-199a-3p in CDDP
chemosensitivity in ovarian cancer. We found that miR-199a-3p was
significantly downregulated in CDDP-resistant ovarian cancer
tissues and cell lines. Overexpression of miR-199a-3p results in
enhanced CDDP sensitivity. ITGB8 was a direct target of miR-199a-3p
and ectopic expression of ITGB8 alleviated the CDDP sensitivity.
Importantly, miR-199a-3p also enhanced CDDP sensitivity in ovarian
cancer in vivo. Collectively, our findings suggest that
miR-199a-3p plays a critical role in regulating CDDP
chemosensitivity by targeting ITGB8 in ovarian cancer.
Materials and methods
Tissue specimens, cell lines and cell
culture
Thirty cases of CDDP-sensitive and 28 cases of
CDDP-resistant human ovarian cancer tissues were collected from the
Department of Obstetrics and Gynecology, Xijing Hospital, The
Fourth Military Medical University between Jan 2008 and Dec 2012.
Tissues were frozen in liquid nitrogen and stored at −80°C until
use. All patients were treated with the standard care of
platinum-based therapy after surgery at least for 6 months.
Platinum resistance or platinum sensitivity was defined as a
relapse or progression within 6 months or 6 months after the last
platinum-based chemotherapy. The present study was approved by the
Ethics Committee of Xi'an No. 1 Hospital and written informed
consent was obtained from each patient. Clinicopathological
characteristics of chemoresistant and chemosensitive ovarian cancer
patients are shown in Table I.
Human ovarian cancer cell line SKOV3 was purchased from Shanghai
Institute of Cell Biology (Shanghai, China). Cells were maintained
in RPMI-1640 supplemented with 10% FBS at 37°C in a humidified
incubator with 5% CO2.
 | Table I.Clinicopathological characteristics of
chemoresistant and chemosensitive ovarian cancer patients. |
Table I.
Clinicopathological characteristics of
chemoresistant and chemosensitive ovarian cancer patients.
| Characteristics | Cases | Chemosensitive, n
(%) | Chemoresistant, n
(%) | χ2 | P-value |
|---|
| Age (years) |
| 34 | 24 |
|
|
|
<50 | 39 | 23 (59.0) | 16 (41.0) | 0.793 | 0.459 |
|
≥50 | 19 | 11 (57.8) | 8 (42.2) |
|
|
| FIGO stage |
|
|
|
|
|
| I | 14 | 10 (71.40) | 4 (28.6) | 1.926 | 0.503 |
| II | 18 | 10 (55.6) | 8 (44.4) |
|
|
|
III | 17 | 9 (52.9) | 8 (47.1) |
|
|
| IV | 9 | 5 (55.6) | 4 (44.4) |
|
|
| Histological
grade |
|
|
|
|
|
| G1 | 6 | 4 (66.7) | 2 (33.7) | 0.732 | 0.625 |
| G2 | 20 | 11 (55.0) | 9 (45.0) |
|
|
| G3 | 32 | 19 (59.3) | 13 (40.7) |
|
|
| Histology |
|
|
|
|
|
|
Serous | 30 | 19 (63.3) | 11 (36.7) | 2.037 | 0.561 |
|
Mucinous | 11 | 6 (54.5) | 5 (35.5) |
|
|
|
Endometrioid | 10 | 6 (60) | 4 (40) |
|
|
| Clear
cell | 3 | 1 (33.3) | 2 (66.7) |
|
|
|
Anaplastic | 4 | 2 (50.0) | 2 (50.0) |
|
|
| Lymph node
metastasis |
|
|
|
|
|
|
Yes | 26 | 11 (42.3) | 15 (57.7) | 2.453 | 0.471 |
| No | 32 | 23 (71.8) | 9 (28.2) |
|
|
Ethics approval
All procedures performed in studies involving human
participants were in accordance with the ethical standards of the
institutional and/or national research committee and with the 1964
Helsinki declaration and its later amendments or comparable ethical
standards. All applicable international, national, and/or
institutional guidelines for the care and use of animals were
followed. All applicable international, national, and/or
institutional guidelines for the care and use of animals were
followed.
Establishment of CDDP-resistant
ovarian cancer cell lines
The CDDP-resistant ovarian cancer cell lines were
induced using progressive concentration of CDDP as previously
described (11). Briefly, SKOV3
cells were first treated with 0.5 µg/ml of CDDP, CDDP was withdrawn
48 h later and cells were cultured without CDDP until they
recovered. The same treatment was performed with CDDP, the
concentration was gradually increased to 1, 2, 3, 4 and finally to
6 µg/ml. When the induced cells still survived in 6 µg/ml of CDDP,
the cells were confirmed to be CDDP-resistant and named
SKOV3/CDDP.
Cell transfection
miR-199a-3p mimics, miR-199a-3p inhibitor and
negative control (NC) were designed and synthesized by
GeneBioPharma (Shanghai, China). Transient transfection was
performed using HiPerFect Transfection Reagent (Qiagen, Hilden,
German) as instructed by the manufacturer. Full-length ITGB8 cDNA
was amplified and cloned into the pcDNA3.1 vector, primer for
amplication ITGB8 was as follows: forward,
5′-CCGGAATTCATGTCCCGCCCACGCTG-3′ and reverse,
5′-CGGGATCCTTAATTAATCCAGTCATCT-3′. Transfection was performed using
Lipofectamine LTX (Thermo Fisher Scientific, Waltham, MA, USA)
according to the manufacturer's protocol. Transfection efficiencies
were assessed using qRT-PCR.
Cytotoxicity assay
The CDDP cytotoxicity was measured using a Cell
Counting Kit-8 (CCK-8) assay (Dojindo Laboratories, Kumamoto,
Japan). Briefly, cells were collected and reseeded in triplicate in
200 µl of medium in a 96-well plate at 5×103 cells/well
with 0, 0.0625, 0.125, 0.25, 0.05, 1, 2, 4, 8 µg/ml of CDDP.
Seventy-two hours after culture, 20 µl of CCK-8 was added and
cultured for additional 4 h. The absorbance at 490 nm was measured
using microplate reader (Bio-Tek, Winooski, VT, USA). The
concentration of CDDP that caused 50% inhibition of ovarian cancer
cell activity was defined as IC50. The inhibition rate
was calculated using the following formula: Inhibition rate (%) =
(1 - ODtreated/ODcontrol) × 100%.
Cell apoptosis assay
Cell apoptosis was evaluated by flow cytometry using
Annexin V-FITC/PI detection kit (KeyGen Biotech Co., Nanjing,
China) according to the manufacturer's instructions. Data were
acquired on a FACSCalibur flow cytometer (BD Biosciences, San Jose,
CA, USA) and analyzed using FlowJo software (Tree Star, Inc.,
Ashland, OR, USA).
Cell cycle analysis
Cell cycle was measured using propidium iodide (PI)
staining. Briefly, 24 h post transfection, cells were collected and
reseeded in triplicate in a 6-well plate at 1×105
cells/well, cells were harvested 24 h later, and washed with PBS,
fixed in ice-cold 70% ethanol overnight. Cells were then washed
with ice-cold PBS, resuspended in 500 µl of a PI master mix
containing 50 µg/ml PI and 100 µg/ml RNAse A (both from Sigma, St.
Louis, MO, USA) diluted in PBS and incubated for 30 min in the dark
at 37°C. Cell cycle was measured using BD FACSCalibur flow
cytometer (BD Biosciences, San Diego, CA, USA), and analyzed using
FlowJo software (Tree Star, Inc.).
Wound healing assay
SKOV3 or SKOV3/CDDP cells were seeded into 6-well
plates at 3×105 cells/well and cultured until confluent.
A P200 pipette tip was used to make a straight scratch, and then
incubated for 48 h. The gap width of scratch re-population was
recorded under the microscope.
Cell invasion assay
Cell migration assays were performed using 24-well
Transwells (8-µm pore size; Millipore Corporation, Billerica, MA,
USA) pre-coated with 100 µl of Matrigel (BD Pharmingen, San Jose,
CA, USA). Briefly, miRNA transfected or control cells were
suspended in 200 µl RPMI-1640 medium then added to the upper
chamber. A volume of 600 µl of RPMI-1640 containing 10% FBS was
placed in the lower chamber. After incubating for 48 h, the
Matrigel and the cells remaining in the upper chamber were removed
using cotton swabs. Cells on the lower surface of the membrane were
fixed with 4% paraformaldehyde and stained with Giemsa
(Sigma-Aldrich, St. Louis, MO, USA). Five microscopic fields (at a
magnification of ×200) were used to count and photograph the
cells.
RNA extraction and real-time
quantitative PCR
Total RNA were extracted from ovarian cancer
tissues, transfected SKOV3 cells or xenografts using the RNeasy
Mini kit (Qiagen, Shanghai, China) following the manufacturer's
instructions. The concentration and quality of the RNA was examined
by NanoDrop 2000 (Thermo Fisher Scientific). Total RNA of 1 µg was
reversely transcribed into cDNA using miScript II RT kit (Qiagen).
cDNA was then used for real-time PCR with miScript SYBR-Green PCR
kit (Qiagen). The amplification was performed on an Applied
Biosystems 7500 Fast Real-Time PCR System (Applied Biosystems,
Foster City, CA, USA), with a first step at 95°C for 3 min and then
40 cycles with 95°C for 15 sec, 60°C for 5 sec, 68°C for 30 sec
with a fluorescent reading at the end of this step. Primers used
were as follows: RT primes for miR-199a-3p,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTAACCAAT-3′; miR-199a-3p
forward, 5′-ACACTCCAGCTGGGACAGTAGTCTGCACAT-3′ and reverse,
5′-TGGTGTCGTGGAGTCG-3′; U6 forward, 5′-ATTGGAACGATACAGAGAAGATT-3′
and reverse, 5′-GGAACGCTTCACGAATTTG-3′; ITGB8 forward,
5′-CCCGTGACTTTCGTCTTGGA-3′ and reverse, 5′-CCTTTCGGGGTGGATGCTAA-3′;
β-actin forward, 5′-CGTCTTCCCCTCCATCGT-3′ and reverse,
5′-GCCTCGTCGCCCACATAG-3′. The relative expression of the gene was
quantified using the comparative 2−ΔΔCt method relative
to the appropriate housekeeping gene.
Protein extraction and western
blotting
Protein was extracted from transfected cells or
xenografts using radioimmunoprecipitation assay (RIPA) lysis buffer
(Beyotime Biotechnology, Shanghai, China). Protein concentration
was assessed using a BCA protein assay kit (KeyGen Biotech Co.).
Protein (50 µg) was separated by 10% SDS-PAGE, then transferred
onto polyvinylidene fluoride membranes (Millipore Corporation,
Bedford, MA, USA). The membrane was washed in 0.1% Tween-20 in
Tris-buffered saline solution (TBST), and blocked using 5% non-fat
milk. The membrane was incubated with primary antibodies against
ITGB8 (#88300) and β-actin (#8457) (both from Cell Signaling
Technology, Inc., Danvers, MA, USA) overnight at 4°C. After washing
3 times, the membrane was incubated with horseradish
peroxidase-conjugated secondary antibody for 1 h at room
temperature in a dilution of 1:2,000. Protein bands were visualized
with ECL™ Western Blotting Detection Reagents (Amersham; GE
Healthcare, Chalfont, UK) and captured using Bio-Rad Imaging
Systems (Bio-Rad).
Dual-luciferase reporter assay
Two 3′-UTR of ITGB8 containing miR-199a-3p binding
sequences was amplified by PCR from genomic DNA. The PCR product
was cloned into psiCHECK-reporter vector (Promega, Madison, WI,
USA). Mutant construct of the ITGB8 3′-UTR-2, containing a mutated
sequence within the miR-199a-3p target site was generated using a
site-directed mutagenesis kit (Takara, Tokyo, Japan).
psiCHECK-ITGB8-control or psiCHECK-ITGB8-3′-UTR-1 or 2, or
psiCHECK-ITGB8-3′-UTR-2 Mut (500 ng) and 50 nM miR-199a-3p mimics
or negative control were co-transfected into SKOV3/CDDP cells using
Lipofectamine LTX. Luciferase signals were measured 48 h after
transfection using a Dual-Luciferase Reporter Assay kit
(Promega).
Orthotopic mouse ovarian cancer
model
Orthotopic mouse ovarian cancer model was generated
in nude mice (female, 6–8 weeks old) as previously described
(12). Briefly, mice were
anesthetized with pentobarbital, a 1.5 cm dorsal incision was
performed in the kidney region, the left ovarian bursa was exposed
and injected with 1×106 SKOV3/CDDP or
SKOV3/CDDP/miR-199a-3p cells in 50 µl PBS (n=5/group). Animals were
euthanized at 10 weeks, tumors were harvested and photographed. The
number of peritoneal metastasis was recorded. The mouse experiments
were approved by the Institutional Animal Care and Use Committee of
Xi'an No. 1 Hospital.
Statistical analysis
Data are shown as means ± SD. Differences between
groups were evaluated using the Student's t-test or one-way ANOVA.
GraphPad Prism version 6.00 (GraphPad Software, San Diego, CA, USA)
was used for all analyses. P<0.05 was considered as statistical
significance.
Results
miR-199a-3p is downregulated in
cisplatin-resistant ovarian cancer tissues and cell lines
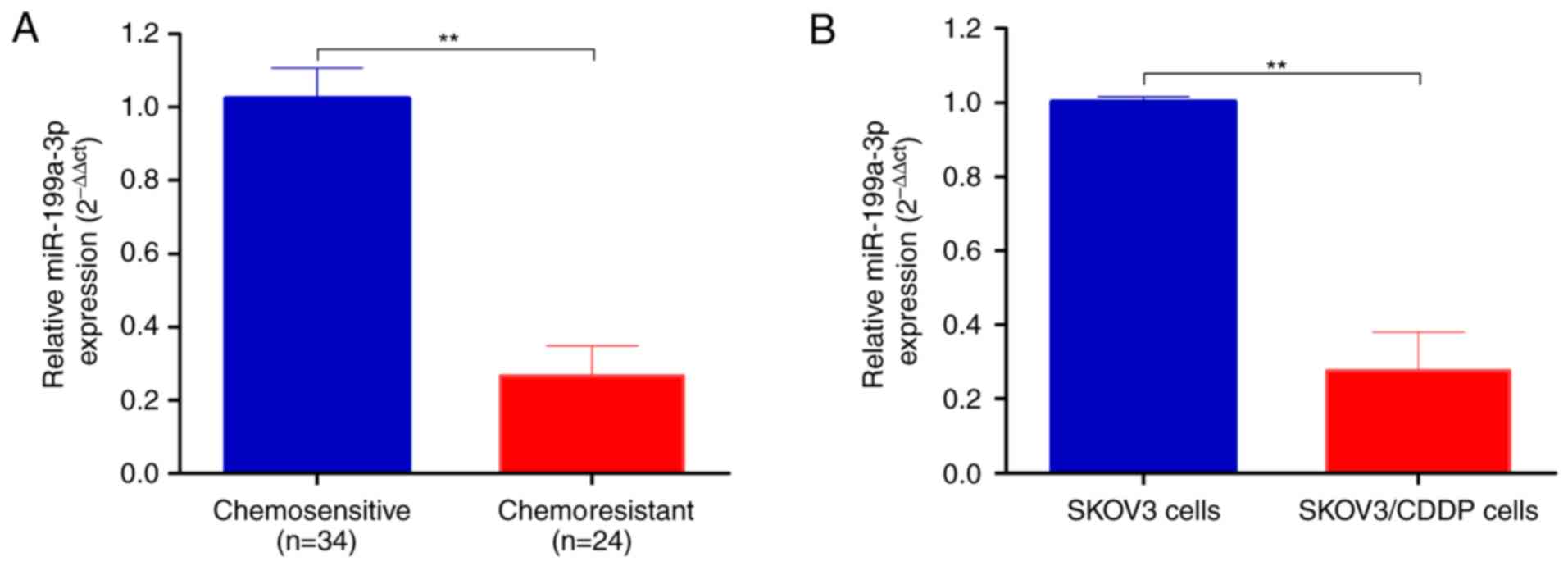
Total RNA was extracted from chemosensitive (n=34)
and chemoresistant (n=24) ovarian cancer tissues. As shown in
Fig. 1A, miR-199a-3p was
significantly decreased in chemoresistant ovarian cancer tissues
compared to chemosensitive carcinomas (2−ΔΔCt,
0.271±0.037 vs. 1.006±0.124, P<0.01).
We then established the CDDP-resistant SKOV3/CDDP
cells by progressive concentration of CDDP induction as described
in Materials and methods. The cell proliferation, metastasis and
invasion capability of CDDP-resistant ovarian cancer SKOV3/CDDP
cell lines were tested. The IC50 value of CDDP for SKOV3
cells was 0.75 and 2.03 µg/ml for SKOV3/CDDP cells. In agreement
with the ovarian cancer tissues, the expression level of
miR-199a-3p was significantly downregulated in CDDP resistant
SKOV3/CDDP cells compared to SKOV3 cells (2−ΔΔCt,
0.07±0.01 vs. 1.001±0.042, P<0.01; Fig. 1B).
miR-199a-3p enhances the sensitivity
of ovarian cancer cell lines to cisplatin
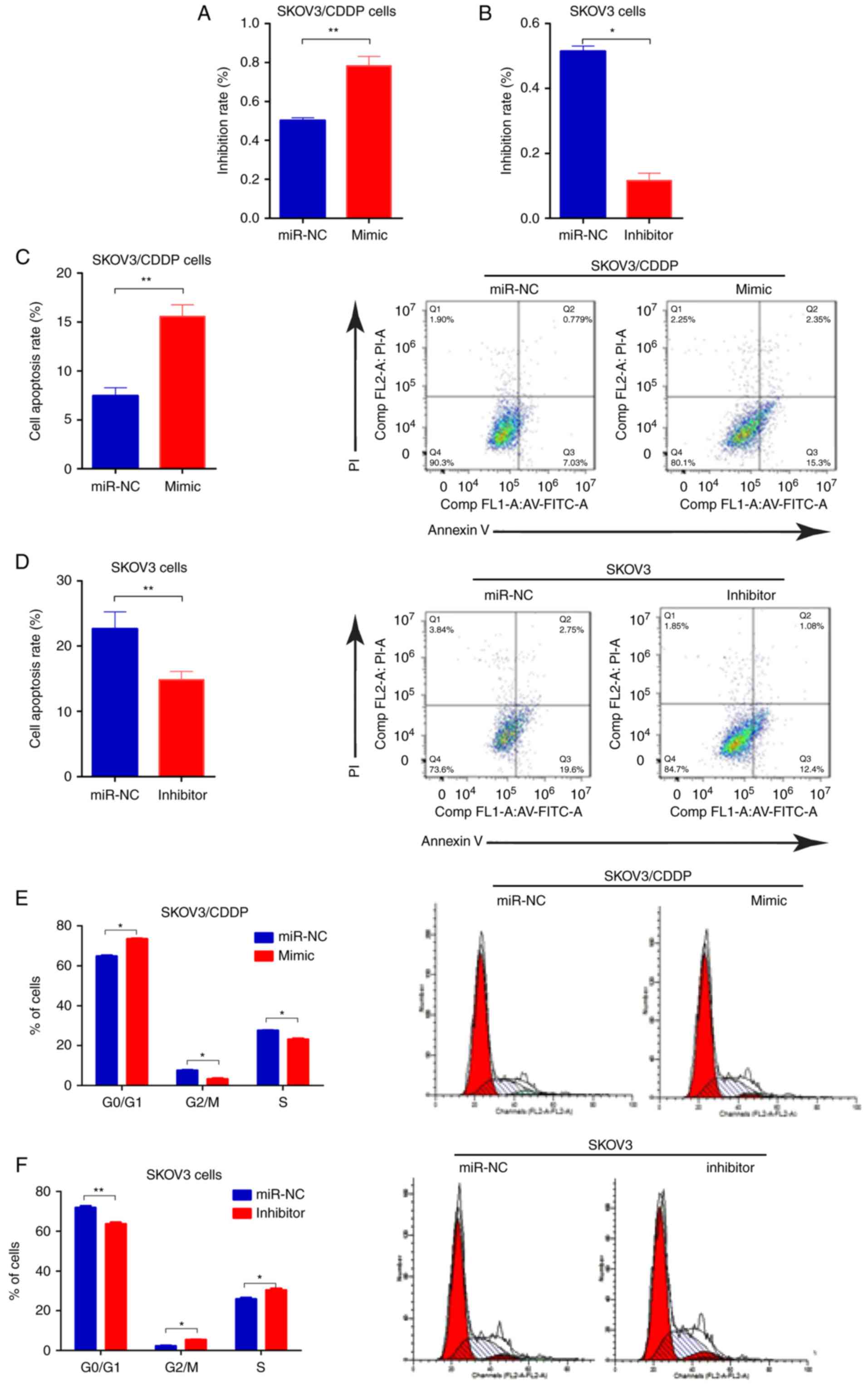
To determine whether miR-199-3p contributed to
cisplatin sensitivity of ovarian cancer cells, we transfected the
SKOV3/CDDP cells with miR-199-3p mimic, and SKOV3 cells with
miR-199-3p inhibitor, respectively. We found that inhibition of
miR-199-3p reduced the inhibition rate of CDDP (0.75 µg/ml) in
SKOV3 cells, compared to the negative controls, whereas miR-199a-3p
mimics improved the toxicity of CDDP (2.03 µg/ml) in SKOV3/CDDP
cells (Fig. 2A and B). Apoptosis of
miR-199a-3p-overexpressing SKOV3/CDDP cells was significantly
increased compared with the negative control, while miR-199a-3p
inhibition could decrease the apoptosis rate in SKOV3 cells
(Fig. 2C and D). Cell cycle assay
indicated that ectopic miR-199a-3p resulted in significantly
increased G1 phase but a reduced S phase in SKOV3/CDDP cells
compared to the negative controls, while miR-199a-3p inhibition
showed the opposite effect (Fig. 2E and
F). In addition, Transwell assay showed that miR-199a-3p
overexpression markedly suppressed the capacity for cell migration
in SKOV3/CDDP cells, while miR-199a-3p inhibition promoted cell
migration in SKOV3 cells (Fig. 2G and
H). These results indicate that miR-199a-3p enhanced the
sensitivity of ovarian cancer cell lines to CDDP.
ITGB8 is a direct target of
miR-199a-3p
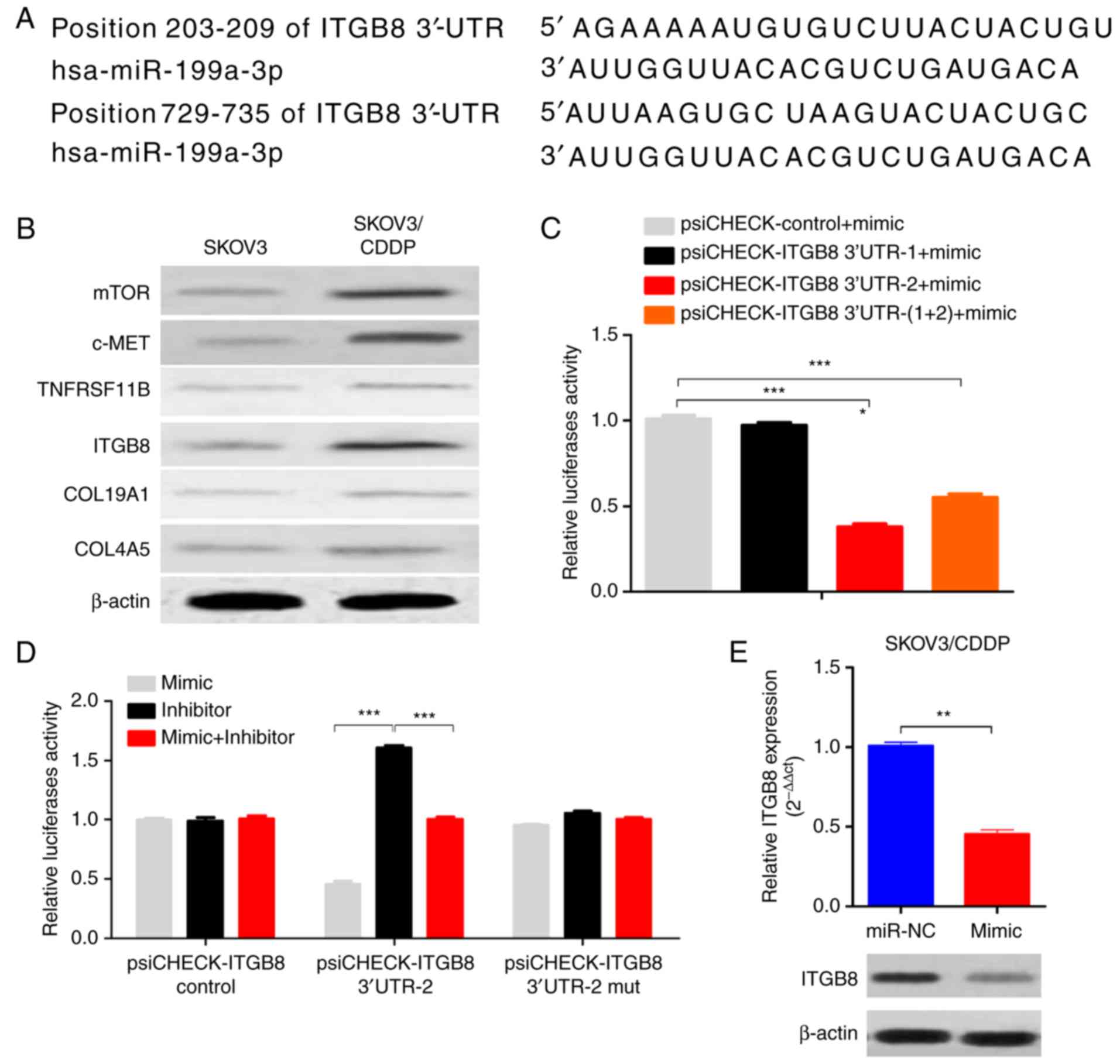
The computer-aided algorithms (TargetScan Release
5.2, http://www.targetscan.org; and PicTar,
(http://pictar.mdc-berlin.de/cgi-bin/new_PicTar_vertebrate.cgi)
predicted that integrin β8 (ITGB8), one of the integrins involved
in the regulation of cell cycle and motility, was a potential
target gene of miR-199a-3p (Fig.
3A). There were two putative binding sites for miR-199a-3p on
ITGB8 3′-UTR, one located at position 203–209 (3′-UTR-1), and the
other located at position 729–735 of ITGB8 3′-UTR (3′-UTR-2)
(Fig. 3A). Importantly, the ITGB8
level was also upregulated in SKOV3/CDDP cells compared to SKOV3
cells (Fig. 3B). Luciferase assay
showed that the luciferase activities decreased significantly in
the ITGB8 3′-UTR-2-transfected cells when co-transfected with
miR-199a-3p, while ITGB8 3′-UTR-1 transfected cells had no effect
on the luciferase activities (Fig.
3C). In addition, the inhibition of luciferase activities by
miR-199a-3p was abolished when the ITGB8 3′-UTR-2 site was mutated
(Fig. 3D). miR-199a-3p
overexpression reduced the ITGB8 mRNA and protein levels in
SKOV3/CDDP cells (Fig. 3E). These
results indicate that ITGB8 is a functional direct target of
miR-199a-3p in ovarian cancer cells.
Overexpression of ITGB8 enhances CDDP
resistance in miR-199a-3p overexpressed ovarian cancer cells
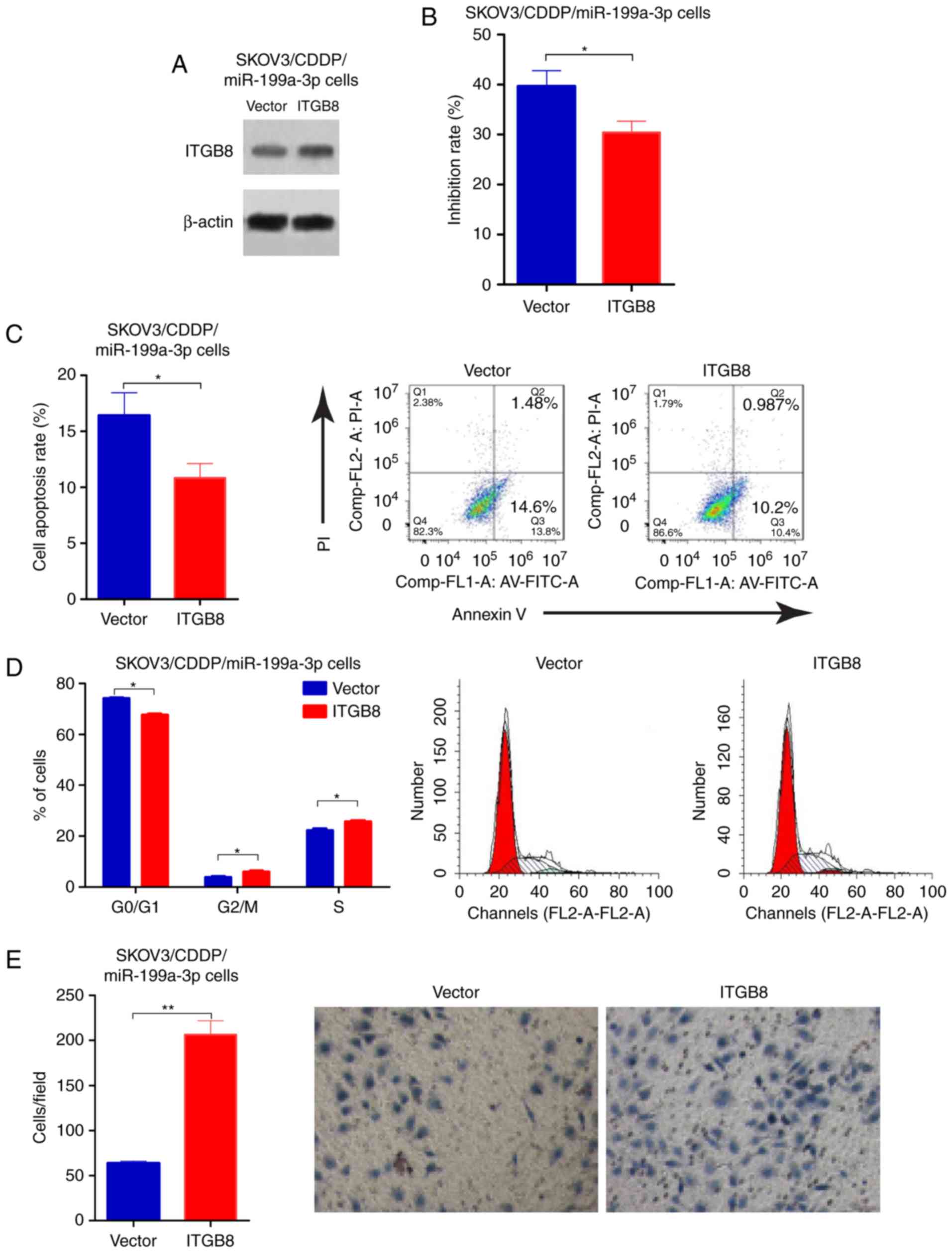
To determine the functional involvement of ITGB8 in
miR-199a-3p-mediated CDDP resistance, miR-199a-3p-overexpression
SKOV3/CDDP cells were transfected with empty vector or ITGB8
plasmid, the transfection efficiency was validated by western
blotting (Fig. 4A). ITGB8
overexpression enhances the CDDP resistance that was inhibited by
miR-199a-3p (Fig. 4B). Cell
apoptosis was reduced in SKOV3/CDDP/miR-199a-3p cells with ITGB8
overexpression compared to the cell line transfected with empty
vector (Fig. 4C). In addition,
ectopic ITGB8 resulted in significantly decreased G1 phase, but an
increased S phase in SKOV3/CDDP/miR-199a-3p cells compared to the
vector controls (Fig. 4D).
Transwell assay showed that ITGB8 overexpression markedly
suppressed the capacity for cell migration in
SKOV3/CDDP/miR-199a-3p cells compared to the vector controls
(Fig. 4E). These results
demonstrated that miR-199a-3p regulated cisplatin sensitivity in
ovarian cancer cells by via suppression of ITGB8.
miR-199a-3p enhances CDDP sensitivity
of ovarian cancer in vivo
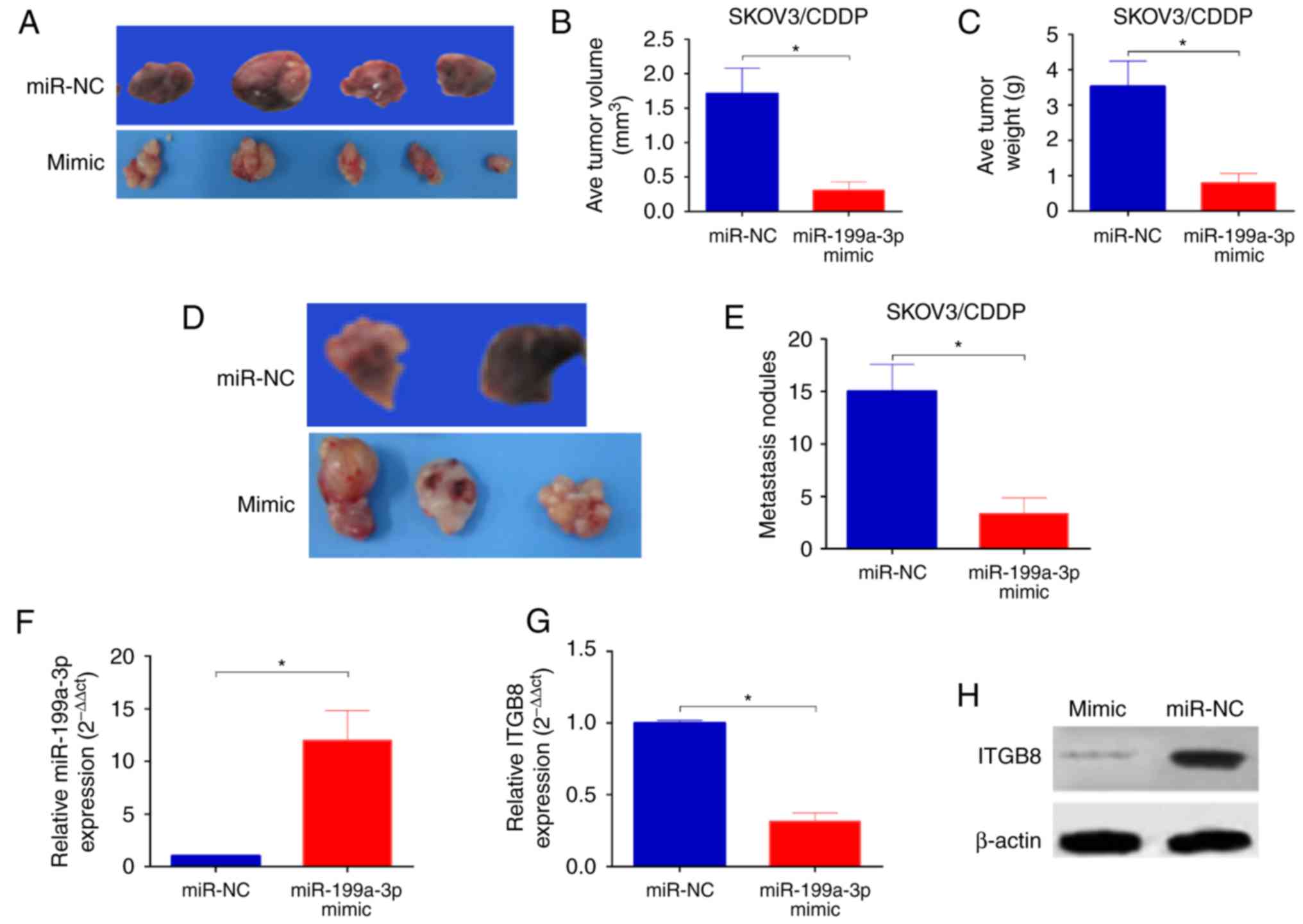
To determine whether the delivery of miR-199a-3p
in vivo enhanced the cytotoxicity of CDDP we established an
orthotopic ovarian cancer model in nude mice by injecting
miR-199a-3p- or miR-NC-overexpression SKOV3/CDDP cells into the
ovary of mice. Ten weeks after injection, significantly reduced
tumor sizes and lower tumor weight were observed in mice injected
with SKOV3/CDDP/miR-199a-3p cells in comparison with those of mice
injected with SKOV3/CDDP/miR-NC (Fig.
5A-C). In addition, the number of peritoneal metastasis was
also markedly decreased in mice injected with
SKOV3/CDDP/miR-199a-3p cells than that in mice injected with
SKOV3/CDDP/miR-NC (Fig. 5D and E).
In addition, the miR-199a-3p levels was significantly upregulated
(Fig. 5F), while the ITGB8
expression was reduced in SKOV3/CDDP/miR-199a-3p xenografts
(Fig. 5G and H). Taken together,
these results indicate that miR-199a-3p enhances the CDDP
sensitivity in ovarian cancer in vivo.
Discussion
miR-199a-3p was first identified in 2003 by computer
analysis of mouse and Fugu rubripes sequences (13). Previous studies showed that
miR-199a-3p was dysregulated in several types of cancers (14–17),
including ovarian cancer (18–20),
and may serve as a tumor suppressor in cancer development (21,22).
Recent studies have reported the essential role of miR-199a-3p in
regulating drug resistance. For example, Li et al, reported
that miR-199a-3p enhanced CDDP sensitivity of cholangiocarcinoma
cells (23). Wang et al,
demostreated that miR-199a could reverse cisplatin resistance in
human ovarian cancer cells through the inhibition of mTOR (19). miR-199a-3p also regulated
doxorubicin sensitivity of human hepatocarcinoma cells (24). However, the role and mechanism of
miR-199a-3p involved in CDDP chemosensitivity in ovarian cancer
remains largely unknown.
In the present study, we found significant
downregulation of miR-199a-3p in chemoresistant ovarian cancer
tissues, as well as CDDP-resistant cell line. Since miR-199a-3p
downregulation was strongly related to CDDP resistant breast cancer
and cholangiocarcinoma cells (23,25),
as well as ovarian cancer cells (19). miR-199a-3p may be involved in
CDDP-resistance in ovarian cancer. Functional experiments
demonstrated that forced miR-199-3p significantly reduced cell
viability, G1 phase cell cycle arrest, cell invasion and promoted
apoptosis in cisplatin-resistant SKOV3/CDDP cells, compared to
negative controls, whereas downregulation of miR-199a-3p showed the
opposite effect, indicating that the role of miR-199a-3p in
regulating cell growth, apoptosis, as well as metastasis in ovarian
cancer may be correlated with the maintenance of CDDP sensitivity.
However, the underlying molecular mechanism of miR-199a-3p in
cisplatin resistance remains unclear.
We then analyzed the potential targets of
miR-199a-3p and found that ITGB8 was significantly downregulated by
miR-199a-3p. ITGB8 is a member of the integrin β-chain family and
encodes a single-pass type I membrane protein with a VWFA domain
and four cysteine-rich repeats (26). It binds to an α subunit to form a
heterodimeric integrin complex (27). They also play critical roles in
signal transduction that was involved in the regulation of cell
growth and motility (28). In
addition to cell motility, ITGB8 was recently reported associated
with gefitinib resistance in hepatic cancer. Wang et al,
showed that ITGB8 silencing decreased cell proliferation and
increased gefitinib sensitivity in HepG2/G cells (29). Wala et al, showed that
miR-199a-3p regulated ITGB8 expression in papillary renal cell
carcinoma (17). In the present
study, we showed a significantly negative correlation between ITGB8
and miR-199a-3p levels in ovarian cancer cells. In addition, no
significant effect was observed in the luciferase activity of ITGB8
mutation between miR-199a-3p overexpressed and miR-199a-3p normally
expressed cell lines, overexpression of miR-199a-3p remarkably
decreased both mRNA and protein level of ITGB8 in SKOV3/CDDP cells,
indicating that ITGB8 was a direct and functional target of
miR-199a-3p.
In summary, the present study demonstrates, for the
first time, that miR-199a-3p increases ovarian cancer cell
sensitivity to CDDP by decreasing ITGB8 expression. The combination
of miR-199a-3p with chemotherapy agents may prevent the development
of drug resistance in ovarian cancer with CDDP-resistance. However,
the precise molecular mechanisms of miR-199a-3p, and the
opportunities for its clinical use, require further
investigation.
Acknowledgements
The present study was supported by the Programs for
Science and Technology Development of Shaanxi Province
(2016SF138).
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jayson GC, Kohn EC, Kitchener HC and
Ledermann JA: Ovarian cancer. Lancet. 384:1376–1388. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cress RD, Chen YS, Morris CR, Petersen M
and Leiserowitz GS: Characteristics of long-term survivors of
epithelial ovarian cancer. Obstet Gynecol. 126:491–497. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hammond SM: An overview of microRNAs. Adv
Drug Deliv Rev. 87:3–14. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Eitan R, Kushnir M, Lithwick-Yanai G,
David MB, Hoshen M, Glezerman M, Hod M, Sabah G, Rosenwald S and
Levavi H: Tumor microRNA expression patterns associated with
resistance to platinum based chemotherapy and survival in ovarian
cancer patients. Gynecol Oncol. 114:253–259. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang N, Kaur S, Volinia S, Greshock J,
Lassus H, Hasegawa K, Liang S, Leminen A, Deng S, Smith L, et al:
MicroRNA microarray identifies Let-7i as a novel biomarker and
therapeutic target in human epithelial ovarian cancer. Cancer Res.
68:10307–10314. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tian S, Zhang M, Chen X, Liu Y and Lou G:
MicroRNA-595 sensitizes ovarian cancer cells to cisplatin by
targeting ABCB1. Oncotarget. 7:87091–87099. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhu X, Shen H, Yin X, Long L, Xie C, Liu
Y, Hui L, Lin X, Fang Y, Cao Y, et al: miR-186 regulation of Twist1
and ovarian cancer sensitivity to cisplatin. Oncogene. 35:323–332.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
van Jaarsveld MT, Helleman J, Boersma AW,
van Kuijk PF, van Ijcken WF, Despierre E, Vergote I, Mathijssen RH,
Berns EM, Verweij J, et al: miR-141 regulates KEAP1 and modulates
cisplatin sensitivity in ovarian cancer cells. Oncogene.
32:4284–4293. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li N, Yang L, Wang H, Yi T, Jia X, Chen C
and Xu P: miR-130a and miR-374a function as novel regulators of
cisplatin resistance in human ovarian cancer A2780 cells. PLoS One.
10:e01288862015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fu X, Tian J, Zhang L, Chen Y and Hao Q:
Involvement of microRNA-93, a new regulator of PTEN/Akt signaling
pathway, in regulation of chemotherapeutic drug cisplatin
chemosensitivity in ovarian cancer cells. FEBS Lett. 586:1279–1286.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang Y, Luo L, Zheng X and Yu T: An
advanced orthotopic ovarian cancer model in mice for therapeutic
trials. Biomed Res Int. 2016:25857872016.PubMed/NCBI
|
|
13
|
Lim LP, Lau NC, Weinstein EG, Abdelhakim
A, Yekta S, Rhoades MW, Burge CB and Bartel DP: The microRNAs of
Caenorhabditis elegans. Genes Dev. 17:991–1008. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shatseva T, Lee DY, Deng Z and Yang BB:
MicroRNA miR-199a-3p regulates cell proliferation and survival by
targeting caveolin-2. J Cell Sci. 124:2826–2836. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Minna E, Romeo P, De Cecco L, Dugo M,
Cassinelli G, Pilotti S, Degl'Innocenti D, Lanzi C, Casalini P,
Pierotti MA, et al: miR-199a-3p displays tumor suppressor functions
in papillary thyroid carcinoma. Oncotarget. 5:2513–2528. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu C, Xing M, Wang L and Zhang K:
miR-199a-3p downregulation in thyroid tissues is associated with
invasion and metastasis of papillary thyroid carcinoma. Br J Biomed
Sci. 74:90–94. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wala SJ, Karamchandani JR, Saleeb R, Evans
A, Ding Q, Ibrahim R, Jewett M, Pasic M, Finelli A, Pace K, et al:
An integrated genomic analysis of papillary renal cell carcinoma
type 1 uncovers the role of focal adhesion and extracellular matrix
pathways. Mol Oncol. 9:1667–1677. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen R, Alvero AB, Silasi DA, Kelly MG,
Fest S, Visintin I, Leiser A, Schwartz PE, Rutherford T and Mor G:
Regulation of IKKbeta by miR-199a affects NF-kappaB activity in
ovarian cancer cells. Oncogene. 27:4712–4723. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Z, Ting Z, Li Y, Chen G, Lu Y and Hao
X: microRNA-199a is able to reverse cisplatin resistance in human
ovarian cancer cells through the inhibition of mammalian target of
rapamycin. Oncol Lett. 6:789–794. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Z, Liu Q, Gao Q, Luo Y, Li Q and Chen
J: Involvement of miR-199a in cisplatin resistance of ovarian
cancer cell through modulating expression of mTOR. Zhonghua Yi Xue
Za Zhi. 95:2705–2708. 2015.(In Chinese). PubMed/NCBI
|
|
21
|
Qu F, Zheng J, Gan W, Lian H, He H, Li W,
Yuan T, Yang Y, Li X, Ji C, et al: MiR-199a-3p suppresses
proliferation and invasion of prostate cancer cells by targeting
Smad1. Oncotarget. 8:52465–52473. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ghosh A, Dasgupta D, Ghosh A, Roychoudhury
S, Kumar D, Gorain M, Butti R, Datta S, Agarwal S, Gupta S, et al:
MiRNA199a-3p suppresses tumor growth, migration, invasion and
angiogenesis in hepatocellular carcinoma by targeting VEGFA,
VEGFR1, VEGFR2, HGF and MMP2. Cell Death Dis. 8:e27062017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li Q, Xia X, Ji J, Ma J, Tao L, Mo L and
Chen W: MiR-199a-3p enhances cisplatin sensitivity of
cholangiocarcinoma cells by inhibiting mTOR signaling pathway and
expression of MDR1. Oncotarget. 8:33621–33630. 2017.PubMed/NCBI
|
|
24
|
Fornari F, Milazzo M, Chieco P, Negrini M,
Calin GA, Grazi GL, Pollutri D, Croce CM, Bolondi L and Gramantieri
L: MiR-199a-3p regulates mTOR and c-Met to influence the
doxorubicin sensitivity of human hepatocarcinoma cells. Cancer Res.
70:5184–5193. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fan X, Zhou S, Zheng M, Deng X, Yi Y and
Huang T: MiR-199a-3p enhances breast cancer cell sensitivity to
cisplatin by downregulating TFAM (TFAM). Biomed Pharmacother.
88:507–514. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
LaPointe VL, Verpoorte A and Stevens MM:
The changing integrin expression and a role for integrin β8 in the
chondrogenic differentiation of mesenchymal stem cells. PLoS One.
8:e820352013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cabodi S, Di Stefano P, Leal Mdel P,
Tinnirello A, Bisaro B, Morello V, Damiano L, Aramu S, Repetto D,
Tornillo G and Defilippi P: Integrins and signal transduction. Adv
Exp Med Biol. 674:43–54. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cabodi S, del Pilar Camacho-Leal M, Di
Stefano P and Defilippi P: Integrin signalling adaptors: Not only
figurants in the cancer story. Nat Rev Cancer. 10:858–870. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang WW, Wang YB, Wang DQ, Lin Z and Sun
RJ: Integrin beta-8 (ITGB8) silencing reverses gefitinib resistance
of human hepatic cancer HepG2/G cell line. Int J Clin Exp Med.
8:3063–3071. 2015.PubMed/NCBI
|