Introduction
Mesenchymal stem cells (MSCs) are multipotent
progenitors that constitute a small proportion of many tissues,
including bone marrow, umbilical cord, amniotic fluid, adipose
tissue and fetal lung tissue (1).
MSCs are described as fibroblast-like cells with the property of
plastic adherence during in vitro culture. MSCs are able to
differentiate into numerous cell lineages (such as adipocytes,
osteoblasts, chondrocytes, tenocytes and cells of visceral
mesoderm) in the presence of different conditional media (2). MSCs are positive for the expression of
CD105, CD73 and CD90 markers, and negative for the expression of
the co-stimulatory factors CD34, CD80, CD86 and HLA-II (3). MSCs also have immunosuppressive
capabilities, meaning that they can modulate the proliferation and
function of all the components of the immune system. Increasing
evidence indicates that MSCs inhibit the maturation of Tregs and
dendritic cells and suppress the functions of memory and naïve T
cells, as well as CD4+ and CD8+ T cells.
Similar effects were also observed in B cells and natural killer
cells (4).
To date, MSCs are considered to be optimal
candidates for use in cell therapy based on these advantages. The
in vivo immunomodulatory properties of MSCs play an
important role in the maintenance of peripheral tolerance and the
induction of transplantation tolerance, which can protect
solid-organ grafts from being rejected (5). Transfusion of MSCs in systemic lupus
erythematosus patients has been revealed to improve the levels of
serological markers and stabilize renal function without the
occurrence of serious adverse events (6). In addition, increasing evidence
indicates that MSCs are involved in various diseases, such as type
I diabetes (7), liver cirrhosis
(8), cerebral palsy (9) and other autoimmune diseases (10). Collectively, these studies indicate
that MSCs have potentially significant clinical applications.
However, the effect of MSCs on malignant tumors has
remained controversial. Certain studies have indicated that MSCs
could inhibit breast tumors (11)
and lung cancer (12), whereas
other studies have demonstrated that MSCs promote the metastasis of
pancreatic (13) and prostate
cancer cells (14). In the present
study, MSCs isolated from umbilical cord were co-cultured with
melanoma A375 cells, and the effect of MSCs on the function of A375
cells was investigated.
Materials and methods
Cell culture
The UCMSCs were purchased from the Sichuan Umbilical
Cord Blood Stem Cell Bank (Chengdu, China). Following dissolving in
a 37°C water bath, UCMSCs were maintained in Dulbecco's modified
Eagle's medium (DMEM; Invitrogen, Carlsbad, CA, USA) supplemented
with 10% fetal bovine serum (FBS; Invitrogen) at 1×105
cells/well in a 6-well plate (BD Falcon; BD Biosciences, Bedford,
MA, USA). The medium was changed every two days, and adherent cells
were harvested after two weeks using 0.25% trypsin (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA). Only UCMSCs passaged
<6 times were used for co-culture since in our previous study
(15) we observed that the
differentiation of UCMSCs increased dramatically when they were
passaged more than 6 times. Melanoma cells A375 [obtained from the
American Type Culture Collection (ATCC; Manassas, VA, USA)] were
cultured in RPMI-1640 (Gibco; Thermo Fisher Scientific) with 10%
FBS. The co-culture ratio of UCMSCs to A375 cells was 2:1.
Cell proliferation assay
Cell proliferation was assessed by an MTT assay at
24, 48 and 72 h following co-culture.
Analysis of apoptosis and cell cycle
distribution by flow cytometry
An apoptosis assay was performed by incubating the
cells with 3 µl Annexin V-FITC (Beyotime Institute of
Biotechnology, Beijing, China) for 10 min and then counterstaining
with 5 µl PI, and detecting the apoptotic cells by flow cytometry.
The cell cycle distribution of the A375 cells was also assessed by
flow cytometry.
RNA extraction and quantitative
PCR
Total RNA was extracted from A375 cells using an
RNeasy Mini kit (Qiagen, Dusseldorf, Germany) at 24, 48 and 72 h
post co-culture with UCMSCs. cDNA was synthesized using a ReverTra
Ace qPCR RT kit (FSQ-101; Toyobo Co., Ltd., Kagoshima, Japan) with
the following transcription conditions: 65°C (5 min), 37°C (15 min)
and 98°C (5 min). Real Master Mix (SYBR Green; FP202; Tiangen
Biotech, Beijing, China) was used for the qPCR with an iCycler
iQ™ Optical Module (Beckman Coulter, Fullerton, CA, USA)
under the following conditions: 95°C for 30 sec, followed by 40
cycles at 95°C for 30 sec, 58°C for 30 sec and 72°C for 30 sec. A
melt curve analysis followed, consisting of increasing temperatures
from 55 to 95°C in 0.5°C increments at 10-sec intervals for 40
cycles. The primers used are listed in Table I. All amplifications were performed
three times.
 | Table I.List of primers in quantitative PCR
detection. |
Table I.
List of primers in quantitative PCR
detection.
| Gene | Forward primer | Reverse primer | GenBank number |
|---|
| BCL2 |
CAGGTTATATCTCATCTTTGAG |
GTTGAGTAACGAGCTGACCCC | KY098794 |
| Survivin |
ACCATAACCCACCACAGC |
CAGTTCGTCCCTTTCCAG | DQ508249 |
| MCL1 |
ATTCCAGAACAGGAGTACAGCTGT |
CAGATGTACCCCTTGTTGTAGAGT | NM_021960 |
| Bax |
GACTTGAGTTGGGAGGGGAA |
GAGGCTCAGCGCCAGGGCTGGG | KJ890756 |
| Fas |
TGGCAACGCTGTCCTGTG |
CCTTTTGCCAGTAGATGCGAG | KR709619 |
| TNFα |
GGTGCTTGTTCCTCAGCCTC |
CAGGCAGAAGAGCGTGGTG | M10988 |
| GAPDH |
GAAGGTGAAGGTCGGAGTC |
GAAGATGGTGATGGGATTTC | J04038 |
Invasion assay
An invasion assay was performed using 24-well (8-µm
pore size) Transwell plates (Corning, Lowell, MA, USA). The A375
cells were plated in the upper chambers, which were pre-coated with
Matrigel (20%; BD Biosciences, Sparks, MD, USA), while UCMSCs were
maintained in the bottom chamber. Toluidine blue (Leagene, Beijing,
China)was used to stain the invaded A375 cells at 24 and 48 h post
co-culture.
Western blot analysis
Collected A375 cells were washed twice with cold PBS
and proteins were extracted using a protein extraction reagent
(Pierce, Rockford, IL, USA) containing protease inhibitors (Roche
Applied Science, Indianapolis, IN, USA). Protein concentration was
assessed using the Micro BCA Protein Assay kit (Pierce). Protein
samples (20 µg) were separated by 12% SDS-polyacrylamide gel
electrophoresis and transferred onto nitrocellulose membranes
(Invitrogen). Following blocking, the membranes were incubated with
primary antibodies (Table II)
followed by horseradish peroxidase-conjugated secondary antibodies
(1:5,000; cat. no. ab97040; Abcam, Cambridge, UK). Antigen-antibody
complexes were visualized using an enhanced chemiluminescence
reagent (Amersham Biosciences, Fairfield, CT, USA).
| Table II.The antibodies used in western blot
analysis. |
Table II.
The antibodies used in western blot
analysis.
| Antibody
(dilutions) | Catalogue number | Company |
|---|
| AKT (1:1,000) | AF6261 | Affinity
Biosciencesa |
| p-AKT (1:1,000) | AF0016 | Affinity
Biosciences |
| PI3K (1:500) | AF6242 | Affinity
Biosciences |
| p-PI3K (1:500) | AF3241 | Affinity
Biosciences |
| STAT3 (1:1,000) | AF6294 | Affinity
Biosciences |
| P-STAT3
(1:1,000) | AF3294 | Affinity
Biosciences |
| ERK (1:1,000) | AF0155 | Affinity
Biosciences |
| P-ERK (1:1,000) | AF1015 | Affinity
Biosciences |
| MTOR (1:1,000) | AF6308 | Affinity
Biosciences |
| P-MTOR (1:1,000) | AF3310 | Affinity
Biosciences |
| GAPDH (1:1,000) | 200608 | Zen
BioScienceb |
Fluorescence detection
A375 cells were maintained on glass slides and
co-cultured with UCMSCs for 48 h. Primary antibodies against
E-cadherin and vimentin were applied to the cells for 4 h at room
temperature. Following secondary antibody staining, A375 cells were
viewed under a confocal scanning microscope (Carl Zeiss, Jena,
Germany)at 488 and 525 nm.
Statistical analysis
qPCR and western blotting results were analyzed with
Bio-Rad CFX manager software (Bio-Rad Laboratories, Inc., Hercules,
CA, USA) and Image Lab software 3.0 (Bio-Rad Laboratories),
respectively. Cell cycle distribution was analyzed by ModFit 3.2
(Verity Software House, Topsham, ME, USA). All data are expressed
as the mean ± standard error using Student's t-test. SPSS 19.0 (IBM
SPSS Statistics, Armonk, NY, USA) was used for statistical
analysis. Values of P<0.05 and P<0.01 were considered to
indicate statistically significant differences compared with the
control group. All figures were generated with GraphPad Prism 5
(GraphPad Software, Inc., La Jolla, CA, USA).
Results
Apoptosis of A375 cells can be induced
by UCMSCs
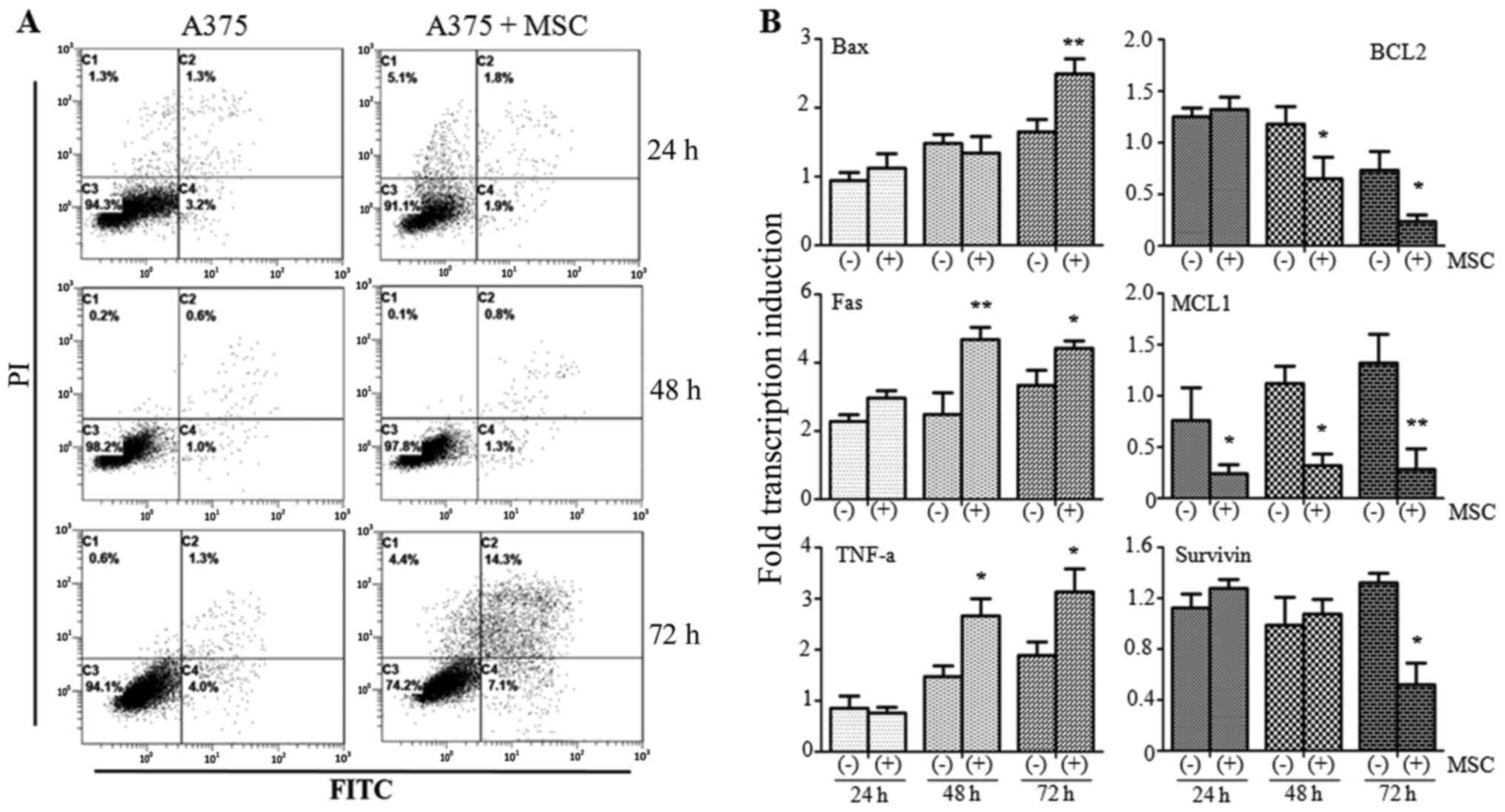
Flow cytometry was performed to detect the
occurrence of apoptosis in A375 cells co-cultured with UCMSCs. The
results indicated no obvious variation in the apoptotic rate
between the A375 monoculture and A375+MSC co-culture groups at 24 h
(4.5 vs. 3.7%, respectively) or 48 h (1.6 vs. 2.1%, respectively).
However, the apoptotic rate increased markedly in the A375+MSC
co-culture group (21.4%) compared with the A375 monoculture group
(5.3%) at 72 h (Fig. 1A).
qPCR was then used to detect the levels of
apoptosis-associated molecules in the different groups. Among the
apoptosis-promoting factors, the expression of Bax was found to be
increased at 72 h (P<0.05) while Fas was markedly increased at
48 h (P<0.01) and 72 h (P<0.05) post co-culture compared with
the A375 monoculture group. TNF-α was also found to be increased at
48 and 72 h (both P<0.05) in the presence of UCMSCs. We also
investigated three anti-apoptotic molecules: BCL2 was suppressed at
48 and 72 h (both P<0.05) while the expression of MCL1 was
inhibited at 24 h (P<0.05), 48 h (P<0.05) and 72 h
(P<0.01) post co-culture. Survivin was only decreased after 72 h
(P<0.05) (Fig. 1B). Our results
indicated that UCMSCs could induce apoptosis in A375 cells.
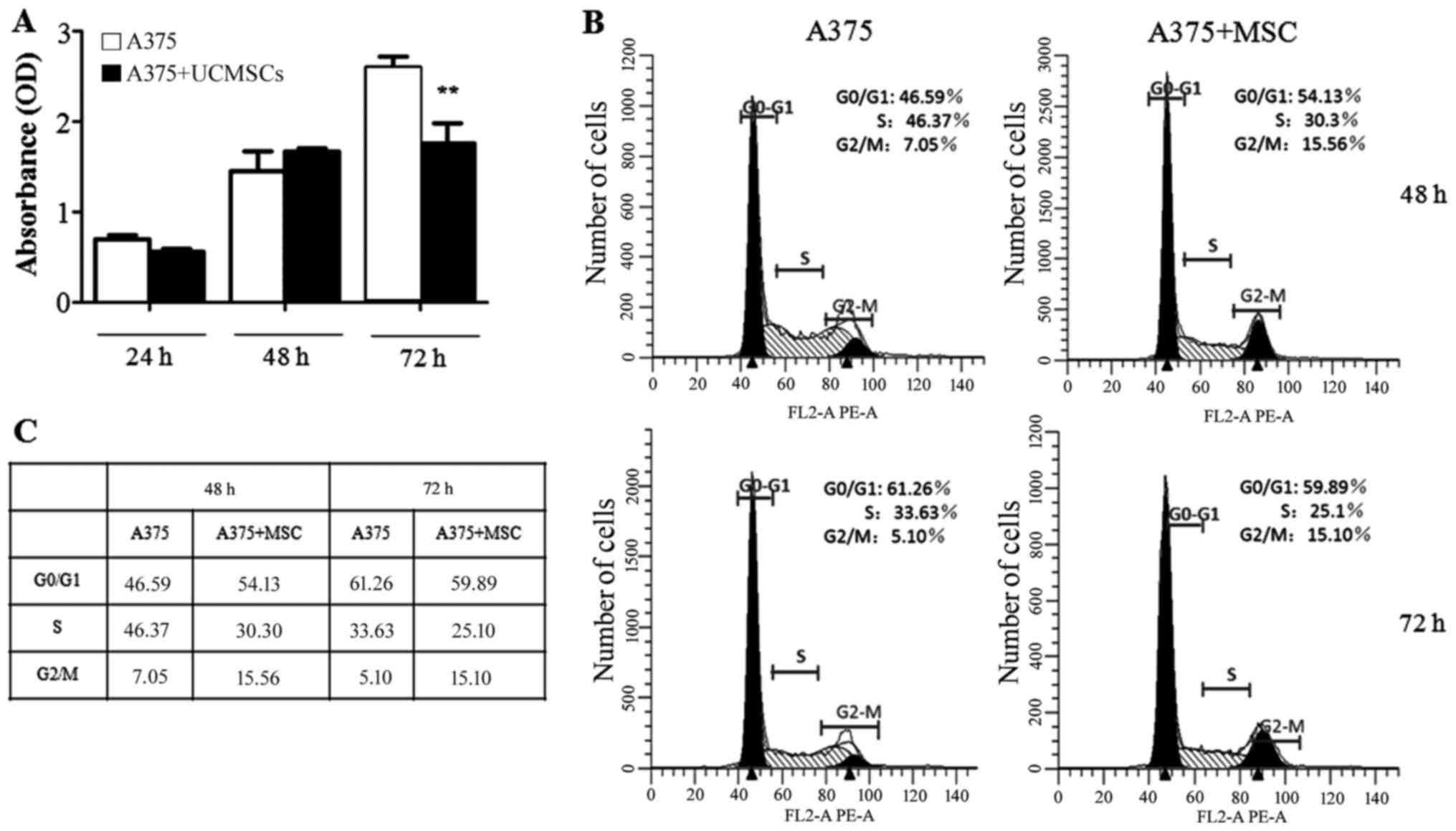
UCMSCs inhibit the proliferation of
A375 cells, however, have no effect on the cell cycle
Subsequently, we detected whether the proliferation
and the cell cycle of A375 cells could be influenced by UCMSCs. In
the MTT assay, the proliferation did not significantly differ
between the A375 monoculture and the A375+UCMSCs co-culture groups
at 24 and 48 h. However, the proliferation of A375 cell was
markedly inhibited (P<0.01) at 72 h post co-culture with UCMSCs
(Fig. 2A). Flow cytometry was then
used to assess the cell cycle distribution in various groups of
A375 cells. The results indicated that UCMSCs had no effect on the
cell cycle at 48 or 72 h post co-culture (Fig. 2B and C).
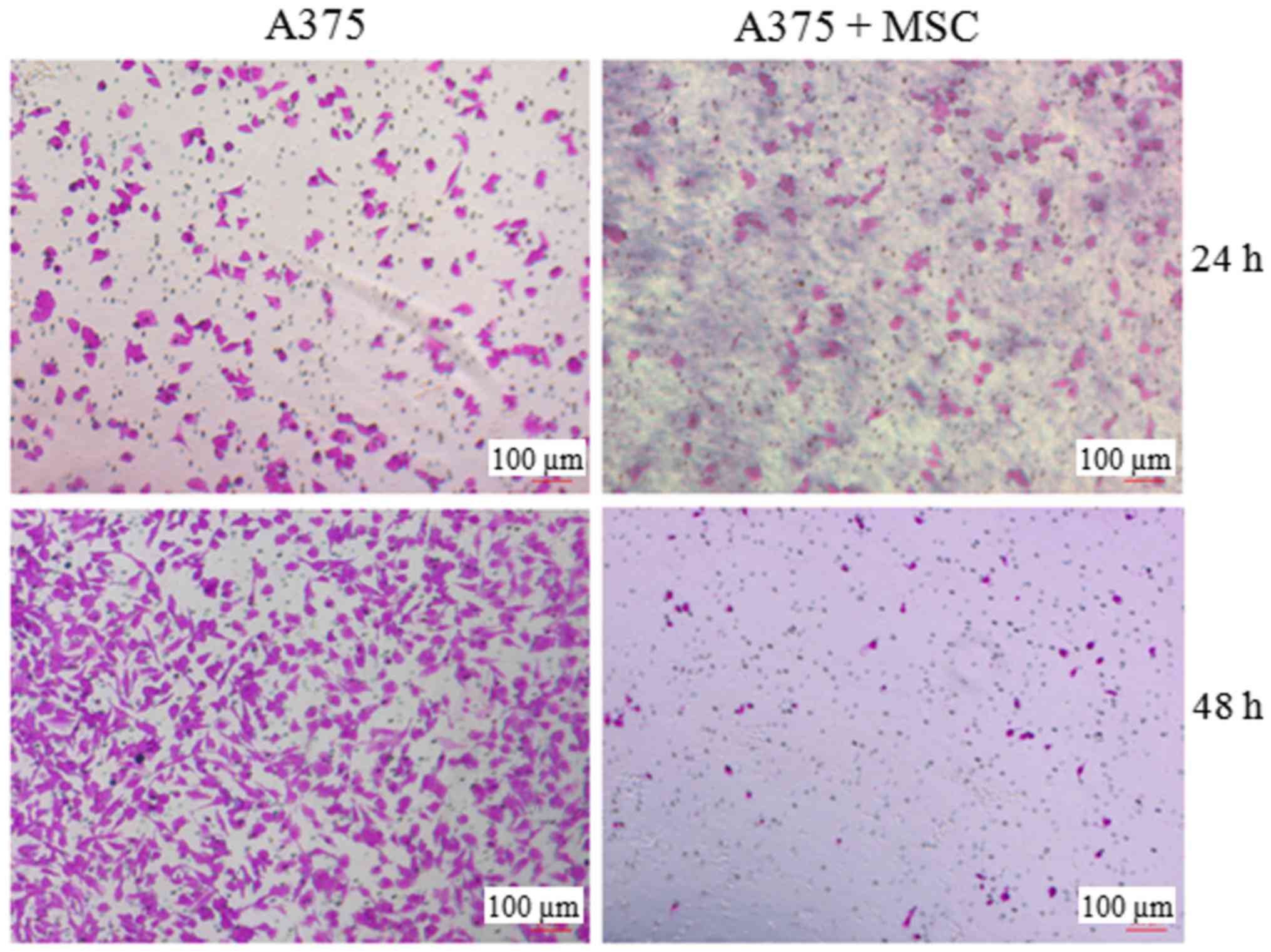
UCMSCs suppress the invasive ability
of A375 cells
As invasion is another important property of
malignant tumor cells, we investigated the influence of UCMSCs on
the invasive ability of A375 cells in a Transwell chamber
pre-coated with Matrigel. As displayed in Fig. 3, the invaded cell numbers showed no
obvious difference between the A375 and A375+MSC groups at 24 h
post co-culture. However, the invasion of the A375 cells was
significantly suppressed in the presence of UCMSCs after 48 h of
co-culture.
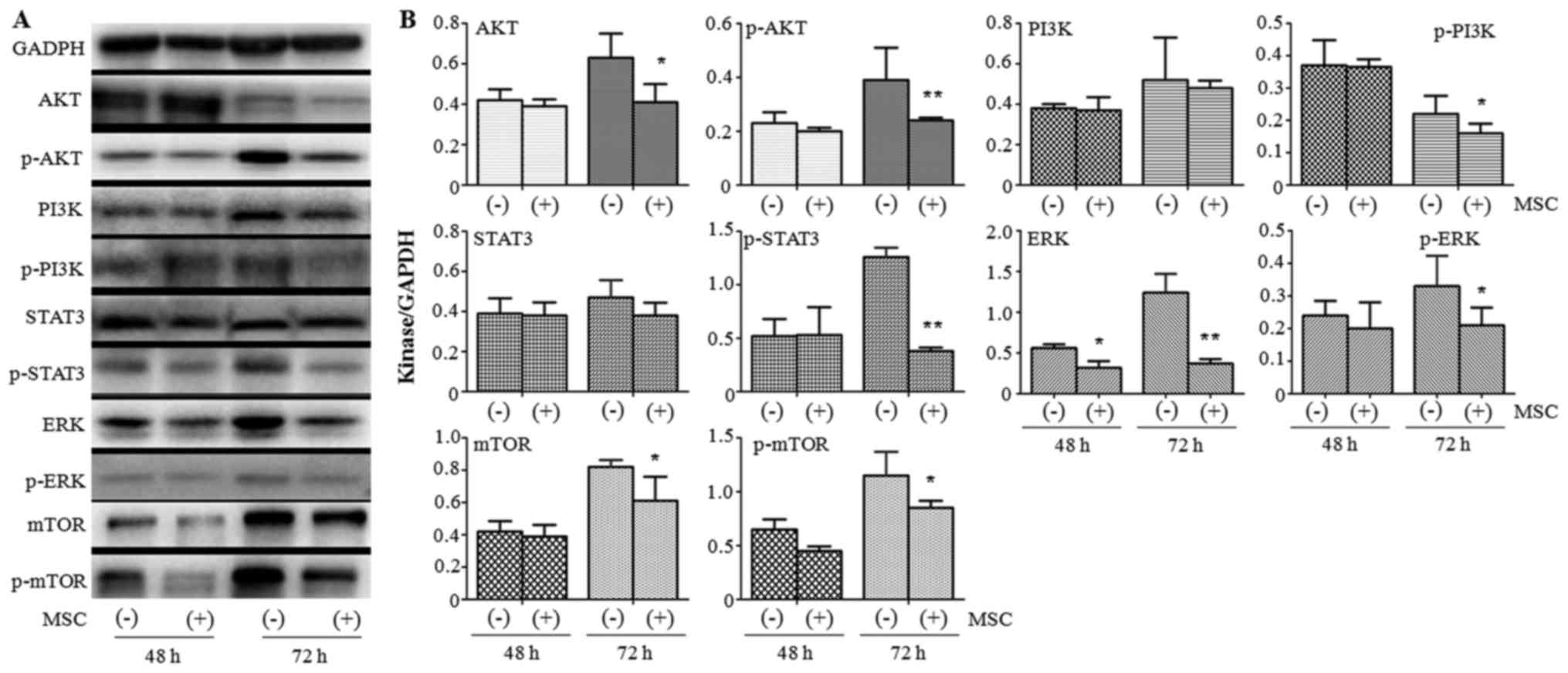
UCMSCs inhibit the expression of
several AKT/STAT3/PI3K/mTOR pathway components in A375 cells
To identify the mechanism by which the functions of
A375 cells were attenuated by UCMSCs, western blotting was
performed to detect the protein expression of several important
kinases at 48 and 72 h post co-culture. We found that all tested
kinases (AKT/p-AKT, PI3K/p-PI3K, STAT3/p-STAT3, ERK/p-ERK and
mTOR), except ERK (P<0.05), demonstrated no significant
variation between the A375 monoculture and A375+MSC co-culture
groups at 48 h (Fig. 4). Following
72 h of co-culture, UCMSCs led to the downregulation of the
expression of AKT (P<0.05), p-AKT (P<0.01), p-PI3K
(P<0.05), p-STAT3 (P<0.01), ERK (P<0.01), p-ERK
(P<0.05), mTOR (P<0.05) and p-mTOR (P<0.05) (Fig. 4). These results indicated the
important role of the AKT/STAT3/PI3K/mTOR pathway in the regulation
of the function of A375 cells by UCMSCs.
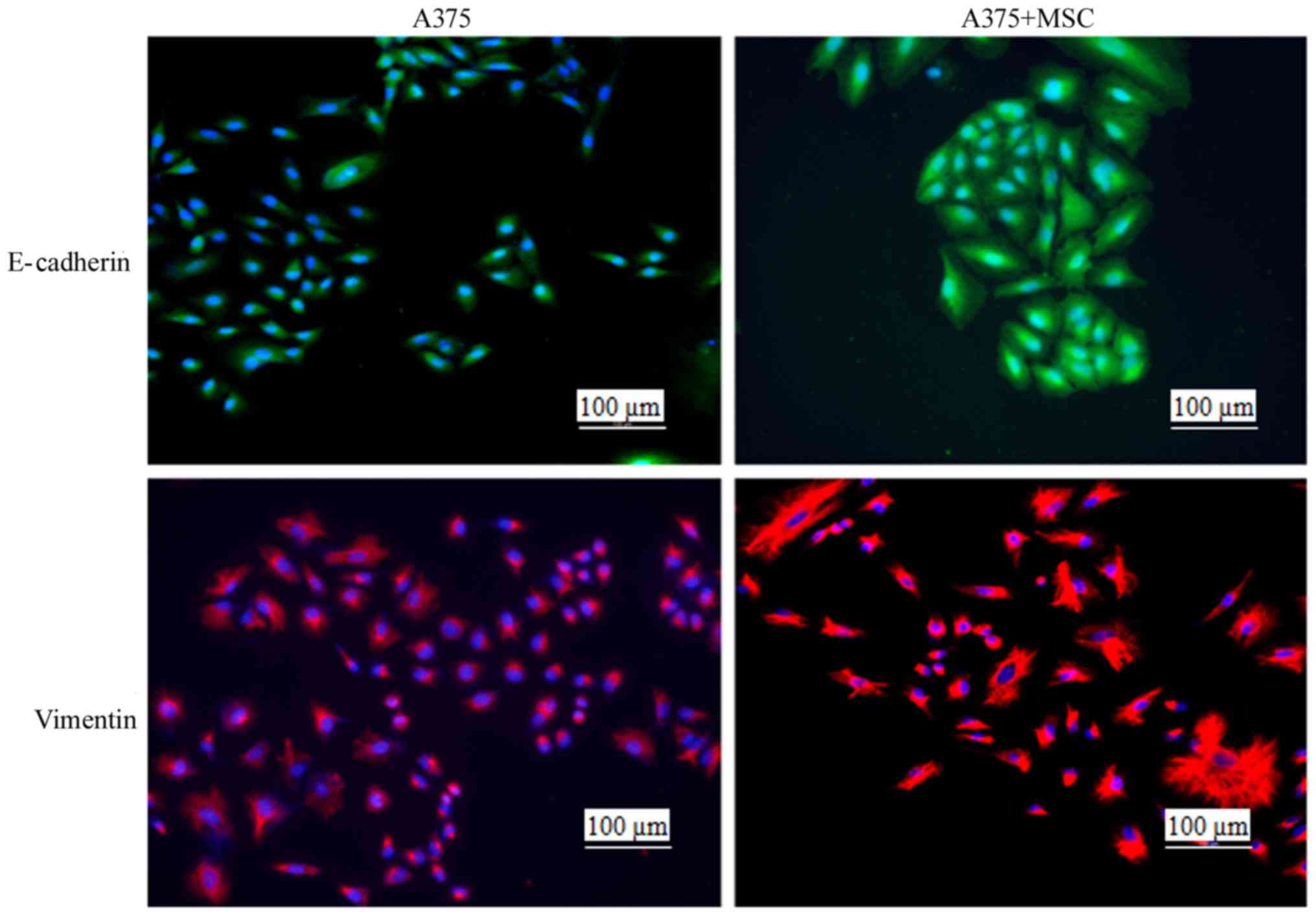
UCMSCs promote cytoskeletal remodeling
in A375 cells
The invasion of malignant tumor cells involves
decreased adherence and enhanced migration properties, and these
changes are part of the epithelial-mesenchymal transition (EMT)
(16). During the EMT process,
cytoskeletal molecules (including E-cadherin, N-cadherin, laminin
and vimentin) play important regulatory roles (17). In the present study, we detected the
occurrence of cytoskeletal rearrangement by staining for E-cadherin
and vimentin at 48 h post co-culture with UCMSCs. The results
indicated that the cytoskeletal rearrangement was obvious in the
A375+MSC co-culture group compared with the A375 monoculture group
(Fig. 5), indicating that in the
presence of UCMSCs, the invasion of A375 cells may be regulated by
certain cytoskeletal molecules.
Discussion
In the present study, we co-cultured melanoma A375
cells with MSCs isolated from umbilical cord to study the role of
UCMSCs in regulating the functions of A375 cells. Although UCMSCs
had no effect on the cell cycle of A375 cells, this study clearly
indicated that UCMSCs inhibited the cell proliferation and
invasion, as well as induced the apoptosis of A375 cells. In the
mechanistic analysis, the expression levels of AKT/PI3K/STAT3/mTOR
pathway components were downregulated following UCMSC co-culture.
Furthermore, marked cytoskeletal rearrangement was observed in the
A375+UCMSC co-culture group. Collectively, these results confirmed
that UCMSCs can exert an antitumor effect on A375 melanoma
cells.
The effect of MSCs on malignant tumor cells remains
controversial. The reported discrepancies may be due to the
heterogeneity of MSCs or the timing and dose of MSC treatments, or
the involvement of other unknown molecules and mechanisms. Liu
et al (18) reported that
UCMSCs could inhibit the growth of human cholangiocarcinoma
HCCC-9810 cells in xenograft models, and that UCMSC-conditioned
medium suppressed cell proliferation (inhibition rate: 6.21 vs.
49.86%) and induced cell apoptosis (9.3 vs. 48.1%) in a dose- and
time-dependent manner. In an immunoblot analysis, the same authors
also found that p-PDK1, p-AKT, β-catenin, cyclin-D1 and c-Myc
mediated the functional changes of HCCC-9810 cells induced by
UCMSCs (18). In another study,
pancreatic cancer cells were co-implanted with MSCs in NOD/SCID
mice, and it was demonstrated that MSC-derived myofibroblast-like
cells could maintain the stemness of tumor-initiating stem cells
among pancreatic cancer cells. The mechanistic analysis indicated
that the Notch-signaling pathway appeared to contribute to the
regulation of stemness by MSCs (13). Wang et al (19) explored the effect of the fusion of
MSCs with esophageal carcinoma cells, and observed that MSCs
markedly decreased tumor cell growth, increased apoptosis and
suppressed tumorigenicity. They also observed that the expression
of DUSP6/MKP3 in the MAPK pathway increased and the exogenous
overexpression confirmed the growth suppression. Sun et al
(20) engineered MSCs stably
transfected with TNF-related apoptosis-inducing ligand (TRAIL) and
co-cultured them with hepatoblastoma HepG2 cells directly, or
applied MSC-conditioned media to the HepG2 cells. The survival rate
of HepG2 cells was markedly decreased by the co-culture conditions,
and TRAIL was indicated to serve an important role in promoting
cell death. All of these studies revealed the uncertainty regarding
the effect of MSCs on malignant tumors.
However, the present study clearly demonstrated the
antitumor effect of MSCs on A375 melanoma cells in a co-culture
system. We not only observed inhibition of proliferation, induction
of apoptosis and suppression of invasion, but also further
investigated the possible mechanism of the MSC-mediated antitumor
effect. Certain important kinases (including AKT, PI3K and STAT3)
were downregulated in A375 cells upon MSC co-culture, and this
finding was consistent with previous research. Of note was the
observation of the rearrangement of the cytoskeleton of A375 cells
co-cultured with MSCs. Based on this, we hypothesized that MSCs
could inhibit tumor invasiveness by regulating the EMT. However,
further research is required to confirm this hypothesis. The
limitation of the present study was that only one cell line was
included, since this was just a preliminary study concerning the
role of UCMSCs in regulating malignant tumor functions. Our
following study will include tumor cell lines from lung, cervical,
breast and prostate cancer.
Acknowledgements
The authors thank the professor Li Zhang (Laboratory
of Pathology, West China Hospital, Sichuan University) for his
continuous support to this work.
Funding
The present study was supported by Sichuan Province
Science and Technology Support Program (nos. 2014SZ0206 and
2014SZ0001).
Availability of data and materials
The datasets used during the study are available
from the corresponding author upon reasonable request.
Authors' contributions
WW carried out fluorescence detection, statistical
analysis and drafted the manuscript. LL was responsible for
completing all the cell related experiments. FC carried out PCR
experiments and western blot analysis. YY conceived of the study
and participated in its design and coordination and helped to draft
the manuscript. All authors read and approved the manuscript and
agree to be accountable for all aspects of the research in ensuring
that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
All experimental protocols have been approved by the
Ethics Committee of West China Hospital, Sichuan University
(Chengdu, China).
Consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
Authors' information
WW is a professor of the Department of Pathology,
West China Second University Hospital, Chengdu. LL is a laboratory
director at the Laboratory of Pathology, West China Hospital,
Sichuan University, Chengdu. FC is an assistant laboratory director
at the Laboratory of Pathology, West China Hospital, Sichuan
University, Chengdu. YY is a professor of the Department of Medical
Oncology, Cancer Center of West China Hospital, Sichuan University,
Chengdu, P.R. China.
References
|
1
|
Pittenger MF, Mackay AM, Beck SC, Jaiswal
RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S and
Marshak DR: Multilineage potential of adult human mesenchymal stem
cells. Science. 284:142–147. 1999. View Article : Google Scholar
|
|
2
|
Phinney DG and Prockop DJ: Concise review:
Mesenchymal stem/multipotent stromal cells: The state of
transdifferentiation and modes of tissue repair-current views. Stem
Cells. 25:2896–2902. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bassi EJ, Aita CA and Camara NO: Immune
regulatory properties of multipotent mesenchymal stromal cells:
Where do we stand? World J Stem Cell. 3:1–8. 2011. View Article : Google Scholar
|
|
4
|
Shi M, Liu ZW and Wang FS:
Immunomodulatory properties and therapeutic application of
mesenchymal stem cells. Clin Exp Immunol. 164:1–8. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sensebe L, Krampera M, Schrezenmeier H,
Bourin P and Giordano R: Mesenchymal stem cells for clinical
application. Vox Sang. 98:93–107. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun L, Wang D, Liang J, Zhang H, Feng X,
Wang H, Hua B, Liu B, Ye S, Hu X, et al: Umbilical cord mesenchymal
stem cell transplantation in severe and refractory systemic lupus
erythematosus. Arthritis Rheum. 62:2467–2475. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang HS, Shu JF, Shen WS, Hsu HC, Chi TC,
Chen CP, Huang SW, Shyr YM, Tang KT and Chen TH: Transplantation of
insulin-producing cells derived from umbilical cord stromal
mesenchymal stem cells to treat NOD mice. Cell Trnsplant.
20:455–466. 2011. View Article : Google Scholar
|
|
8
|
Zhang Z, Lin H, Shi M, Xu R, Fu J, Lv J,
Chen L, Lv S, Li Y, Yu S, et al: Human umbilical cord mesenchymal
stem cells improve liver function and ascites in decompensated
liver cirrhosis patients. J Gastroenterol Hepaol. 27 Suppl
2:S112–S120. 2012. View Article : Google Scholar
|
|
9
|
Wang J, Yin Z, Wen H, Wang L and Chen Z:
Application of umbilical cord mesenchymal stem cells in cerebral
palsy treatment: Report of 51 cases. Chinse Gen Pract.
14:2446–2447. 2011.
|
|
10
|
Liu Y, Mu R, Wang S, Long L, Liu X, Li R,
Sun J, Guo J, Zhang X, Guo J, et al: Therapeutic potential of human
umbilical cord mesenchymal stem cells in the treatment of
rheumatoid arthritis. Arthritis Res Ther. 12:R2102010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ma Y, Hao X, Zhang S and Zhang J: The in
vitro and in vivo effects of human umbilical cord mesenchymal stem
cells on the growth of breast cancer cells. Breast Cancer Res
Treat. 133:473–485. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Matsuzuka T, Rachakatla RS, Doi C, Maurya
DK, Ohta N, Kawabata A, Pyle MM, Pickel L, Reischman J, Marini F,
et al: Human umbilical cord matrix-derived stem cells expressing
interferon-beta gene significantly attenuate bronchioloalveolar
carcinoma xenografts in SCID mice. Lung Cancer. 70:28–36. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kabashima-Niibe A, Higuchi H, Takaishi H,
Masugi Y, Matsuzaki Y, Mabuchi Y, Funakoshi S, Adachi M, Hamamoto
Y, Kawachi S, et al: Mesenchymal stem cells regulate
epithelial-mesenchymal transition and tumor progression of
pancreatic cancer cells. Cancer Sci. 104:157–164. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang T, Lee YW, Rui YF, Cheng TY, Jiang
XH and Li G: Bone marrow-derived mesenchymal stem cells promote
growth and angiogenesis of breast and prostate tumors. Stem Cell
Res Ther. 4:702013. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang Y, Wang Y, Li L, Chen F and Zhang P:
Activation of the Toll-like receptor 8 pathway increases the
immunogenicity of mesenchymal stem cells from umbilical cord. Mol
Med Rep. 16:2061–2068. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Thiery JP: Epithelial-mesenchymal
transition in tumor progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu J, Han GQ, Liu H and Qin C:
Suppression of cholangiocarcinoma cell growth by human umbilical
cord mesenchyaml stem cells: A possible role of wnt and Akt
signaling. PLoS One. 8:e628442013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Y, Fan H, Zhou B, Ju Z, Yu L, Guo L,
Han J and Lu S: Fusion of human umbilical cord mesenchymal stem
cells with esophageal carcinoma cells inhibits the tumorigenecity
of esophageal carcinoma cells. Int J Oncol. 40:370–377.
2012.PubMed/NCBI
|
|
20
|
Sun XY, Nong J, Qin K, Lu H, Moniri MR,
Dai LJ and Warnock GL: MSC (TRAIL)-mediated HepG2 cell death in
direct and indirect co-culture. Anticancer Res. 31:3705–3712.
2011.PubMed/NCBI
|