Introduction
Glioma is the most frequent and aggressive primary
malignant tumor of the central nervous system (CNS), and accounts
for ~80% of primary CNS tumors. According to the 2016 World Health
Organization (WHO) classification, gliomas are classified into four
histopathological grades based on the degree of malignancy,
including low-grade glioma (grade I and II) and high-grade glioma
(grade III and IV), and glioblastoma (GBM) which is the highest
grade of glioma (grade IV) (1).
Despite significant improvements in the treatment of glioma over
recent decades, including surgical resection, local radiotherapy
and systemic chemotherapy, the prognosis of glioma patients remains
poor, with a 5-year survival rate of 20–30% (2,3).
Therefore, investigations into the molecular mechanisms and
therapeutic targets of glioma have attracted extensive attention
and have potential uses in the treatment of glioma (4,5).
With novel technologies having provided accelerating
depths of RNA sequencing, recent studies have demonstrated that the
human genome contains thousands of long non-coding RNAs (lncRNAs)
(6). lncRNAs are >200
nucleotides in length and regulate gene expression by chromatin
modification, at transcriptional and post-transcriptional levels
(7,8). Functional studies for a number of
lncRNAs have demonstrated that they participate in various aspects
of cell biology, including cell proliferation, apoptosis,
differentiation, invasion and metastasis, and potentially
contribute to tumor initiation and malignant progression (9,10).
lncRNAs have also been demonstrated to be involved in hematological
malignancies (11) and multiple
types of solid tumors, including hepatocellular carcinoma (12), breast (13), lung (14), gastric (15) and gynecological cancer (16), as well as melanoma (17). Downregulation or upregulation of
lncRNA expression has been revealed to contribute to the
transformed cancer phenotype, further affecting the
clinicopathological appearance, prognosis and outcome of the cancer
(18–20).
lncRNAs have been implicated in the oncogenesis of
gliomas and are increasingly being considered as potential
therapeutic targets. According to the function of lncRNAs in
glioma, they are classified into oncogenes or tumor suppressors.
HOTAIR (21,22), CRNDE (23), H19 (24,25)
and XIST (26) are notable
oncogenic lncRNAs, which can facilitate tumorigenesis. ADAMTS9-AS2
(27), CASC2 (28), MEG3 (29) and MALAT1 (30) are well-known tumor suppressor
lncRNAs. Dysregulation of tumor suppressor lncRNAs leads to tumor
formation. Therefore, advances in identifying glioma-related
lncRNAs and elucidating their precise molecular mechanisms is
critical for the diagnosis and treatment of glioma.
The present study identified a novel lncRNA,
HOXA10-AS, which is transcribed from the antisense strand of the
homeobox A10 (HOXA10) gene locus at chromosome
7p15.3. HOXA10-AS was markedly upregulated in glioma, and its
increased expression levels were correlated with the malignancy of
glioma. Furthermore, the present study addressed HOXA10-AS as a
regulator of cell proliferation and apoptosis in glioma, and
identified HOXA10 as a target of HOXA10-AS. Additionally, the
oncogene HOXA10 had an important role in promoting the
tumorigenesis of glioma.
Materials and methods
Patients and samples
A total of 59 glioma and 20 normal brain samples
were obtained from the Department of Neurosurgery at the First
Hospital of Jilin University (Changchu, China) from January to
December of 2014. The 59 glioma patients (age range, 35–72 years;
mean age, 46.6 years; 41 males and 28 females) consisted of 32
cases of low-grade glioma (WHO grade I and II) and 24 cases of
high-grade glioma (WHO grade III and IV). All tissue samples were
frozen in liquid nitrogen immediately after resection and stored in
liquid nitrogen until use. All clinical pathological and biological
data were available for these patients. The present study was
approved by the Ethics Committee of The First Hospital of Jilin
University and written informed consent was obtained from all
patients. All the tumor tissues were obtained at primary resection,
and none of the patients had undergone chemotherapy or radiation
therapy prior to surgery.
Cell culture
The human glioma A172 and U251 cell lines were
purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA) and normal human astrocytes (HA) were obtained
from ScienCell Research Laboratories (San Diego, CA, USA). A172 and
U251 cells were maintained in Dulbecco's modified Eagle's medium
(DMEM; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific,
Inc.), and HA cells were maintained in astrocyte medium (ScienCell
Research Laboratories) at 37°C in humidified atmosphere with 5%
CO2.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was isolated from tissues or cultured
cells using TRIzol reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. RNA quantity was
determined using a NanoDrop 2000c Spectrophotometer (Thermo Fisher
Scientific, Inc.). cDNA synthesis was performed with 1 µg total
RNA, using the PrimeScript™ RT reagent kit (Takara Biotechnology,
Dalian, China), according to the manufacturer's protocol. Real-time
PCR was performed on the Takara system, using the SYBR®
Premix Ex Taq™ II Kit (Takara Biotechnology). The cycling
conditions were the following: 30 sec at 95°C, followed by 40
cycles at 95°C for 5 sec and 60°C for 30 sec. GAPDH was used as the
endogenous control. The relative expression was calculated using
the 2−ΔΔCq method (31).
The primers of HOXA10-AS, HOXA10 and GAPDH are listed in Table I.
 | Table I.Primers for real-time qPCR. |
Table I.
Primers for real-time qPCR.
| Primer | Sequences |
|---|
| HOXA10-AS | F:
CCCAGTAAGCCAAAGTCAAGCC |
|
| R:
CTGAGGTCAATGGTGCAAAGG |
| HOXA10 | F:
CAACTGGCTCACGGCAAAGA |
|
| R:
TTCAGTTTCATCCTGCGGTTC |
| GAPDH | F:
GCACCGTCAAGGCTGAGAAC |
|
| R:
TGGTGAAGACGCCAGTGGA |
Lentiviral packaging and stable cell
line establishment
A piLenti-shRNA-GFP lentiviral vector (Applied
Biological Materials, Inc., Richmond, BC, Canada) was used for the
HOXA10-AS-knockdown experiment. In brief, lentiviral particles
expressing HOXA10-AS-specific or control scrambled shRNAs were
co-transfected into 293T cells with the mixed set of packaging
plasmids (SPAX2 and MD2G) using Lipofectamine 3000 reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). The medium was
changed after 8 h and the supernatant containing lentiviruses was
collected 48 h later. Produced lentiviruses were concentrated using
the Centricon Plus-20 centrifugal filter device (EMD Millipore,
Billerica, MA, USA). Lentiviral stock was titred and stored at
−80°C. A172 and U251 cells were infected with the HOXA10-AS shRNA
construct, and GFP+ cells were sorted using a flow
cytometer (FACSAria II; Becton-Dickinson, Mountain View, CA, USA).
Subsequently, the cells were cultured with regular complete medium.
Finally, the cells were analyzed for mRNA expression by RT-qPCR.
The following two targets were used for HOXA10-AS knockdown: Target
1, ACAGGAAACTACCTAAATCACCGACCAGT; and target 2,
GTTCTGGTGCTGCCCGCGAAGGGCTGCCT.
MTT cell viability analysis
MTT assay (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) was performed to determine cell viability. A total of
1×103 cells were seeded into each well of 96-well plates
and cultured in 200 µl growth medium. The cells were incubated with
fresh complete medium every 2 days and after 6 days, 20 µg MTT was
added to each well, followed by incubation at 37°C for 4 h. The
reaction was stopped by adding 150 µl dimethyl sulfoxide (DMSO) to
each well and incubating at 37°C for 10 min. The absorbance was
then determined at 450 nm using a microplate reader (Molecular
Devices LLC, Sunnyvale, CA, USA).
BrdU cell proliferation assay
Cell proliferation was evaluated using BrdU Flow kit
(BD Biosciences, San Diego, CA, USA). Cells were plated into 6-well
plates with 2 ml complete growth medium/well, and prior to adding
BrdU, the complete medium was replaced with serum-free medium.
After 12 h, the serum-free medium was replaced with complete
medium, and BrdU (1 mM/ml) was added to each well and incubated for
4 h. Subsequently, the cells were harvested, fixed and incubated
with anti-BrdU and 7-AAD according to the manufacturer's protocol.
Cell cycle distribution was determined using a flow cytometer
(FACSAria II; BD Biosciences).
Cell apoptosis analysis
The Annexin V-PE assay kit (BD Biosciences) was used
to analyze cell apoptosis in glioma cells. Cells were harvested at
their exponential growth phase and were resuspended in 100 µl
binding buffer at a density of 1×106 cells/ml.
PE-conjugated Annexin V and 7-AAD reagent staining was performed at
the concentrations and times recommended by the manufacturer.
Stained cells were analyzed using a flow cytometer (FACSAria II; BD
Biosciences), and data were analyzed using FlowJo V10 software
(FlowJo LLC, Ashland, OR, USA).
Western blot analysis
Total protein was isolated from cells using
radioimmunoprecipitation assay lysis buffer (Pulilai Gene
Technology, Co., Ltd., Beijing, China). Protein concentrations were
determined with a BCA protein assay kit (Pulilai Gene Technology,
Co., Ltd.) and the mass of protein loaded per lane was 40 µg. Total
proteins were separated by 12% SDS-PAGE and transferred onto a
polyvinylidene difluoride (PVDF) membrane. The membrane was blocked
with 5% (W/V) non-fat milk (Pulilai Gene Technology, Co., Ltd.) at
room temperature for 1 h, and incubated with the following primary
antibodies: Anti-caspase-3 (1:1,000; cat. no. 9662; Cell Signaling
Technology, Inc., Danvers, MA, USA), anti-cleaved caspase-3
(1:1,000; cat. no. 9664; Cell Signaling Technology, Inc.),
anti-Bcl-2 (1:1,000; cat. no. 4223; Cell Signaling Technology,
Inc.) and anti-β-actin (1:1,000; cat. no. 4970; Cell Signaling
Technology, Inc.) at 4°C overnight and then incubated with
secondary antibody (anti-rabbit IgG, HRP-linked antibody; 1:2,000;
cat. no. 7074; Cell Signaling Technology, Inc.) at room temperature
for 1 h. Bands were visualized using an enhanced chemiluminescence
reagent (Thermo Fisher Scientific, Inc.). β-actin was used as a
loading control.
Small interfering RNA
transfection
HOXA10-knockdown in glioma cells was performed by
siRNA transfection. siRNA oligonucleotides targeting HOXA10 were
designed and synthesized by Shanghai GenePharma Co., Ltd.
(Shanghai, China), and the non-specific siRNA oligonucleotides
(Shanghai GenePharma Co., Ltd.) were used as a negative control.
Cells were transfected with 150 pmol siRNA/well in 6-well plates,
using Lipofectamine RNAiMax Reagent (Invitrogen.), according to the
manufacturer's protocol. All siRNA oligonucleotide sequences are
listed in Table II.
| Table II.siRNA oligonucleotides. |
Table II.
siRNA oligonucleotides.
| siRNA | Sequences |
|---|
| siHOXA10-1 | S:
GUCAGCCAGAAAGGGCUAUTT |
|
| A:
AUAGCCCUUUCUGGCUGACTT |
| siHOXA10-2 | S:
CCAUAGACCUGUGGCUAGATT |
|
| A:
UCUAGCCACAGGUCUAUGGTT |
| siHOXA10-3 | S:
CGCAGAACAUCAAAGAAGATT |
|
| A:
UCUUCUUUGAUGUUCUGCGTT |
| siControl | S:
UUCUCCGAACGUGUCACGUTT |
|
| A:
ACGUGACACGUUCGGAGAATT |
Statistical analysis
All data are presented as the mean ± standard error
of the mean (SEM) and were analyzed using Student's t-test.
Spearman's correlation analysis was performed to investigate the
correlation between HOXA10 and HOXA10-AS. All
experiments were performed at least three times. All statistical
analyses were performed using SPSS 19.0 (IBM Corp., Armonk, NY,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
HOXA10-AS is overexpressed in human
glioma samples and cell lines
The human homeobox A (HOXA10) gene is
located in the HOXA gene cluster at chromosome 7p15.3. A
GeneBank search identified the gene HOXA10-AS in a
tail-to-tail orientation relative to the HOXA gene, on the
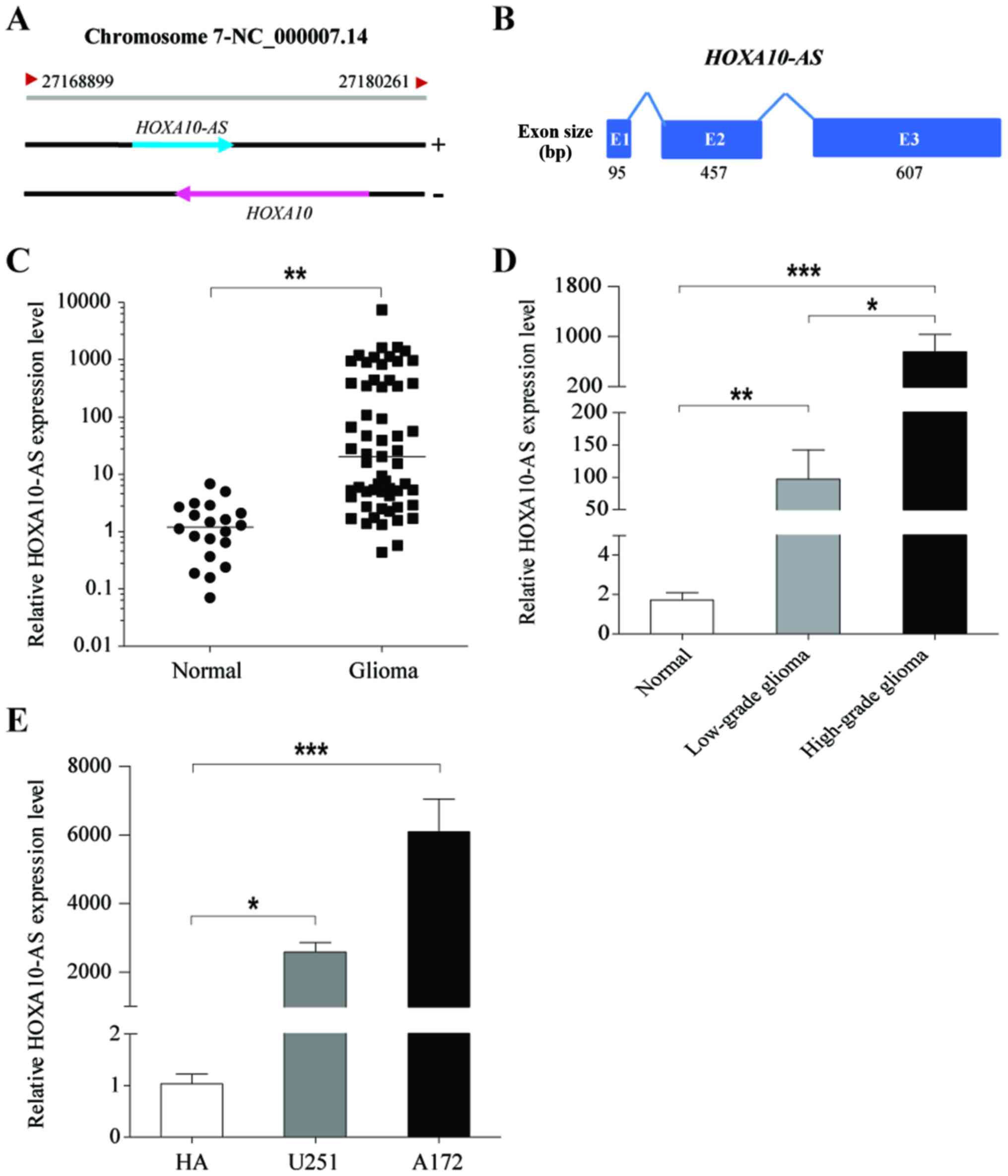
opposite strand of the HOXA10 gene (Fig. 1A). HOXA10-AS is a 1,159-bp long
non-coding antisense transcript of HOXA10, consisting of three
exons with a 3′ polyadenylation tail (Fig. 1B).
To investigate the expression levels of HOXA10-AS in
glioma, tumor tissues were collected from 59 patients with glioma
and normal brain tissues were obtained from 20 healthy patients,
and the expression levels of HOXA10-AS were examined using qPCR.
The results demonstrated that HOXA10-AS was significantly
upregulated in glioma tissues compared with expression in normal
brain tissues (Fig. 1C).
Furthermore, it was demonstrated HOXA10-AS expression was
associated with the malignancy grade of brain tumors, and that
HOXA10-AS expression in cases of high-grade glioma (WHO III and IV,
n=27) was significantly higher than in cases of low-grade glioma
(WHO I and II, n=32; Fig. 1D).
Additionally, HOXA10-AS expression was analyzed in two human glioma
cell lines, A172 and U251 cells, and normal human astrocytes (HA).
Data demonstrated that HOXA10-AS expression was significantly
increased in glioma cell lines compared with that in HA (Fig. 1E). Therefore, these results
indicated that HOXA10-AS was frequently upregulated in glioma, and
may serve an important role in the progression of malignant
glioma.
HOXA10-AS promotes glioma cell
proliferation
To investigate the involvement of HOXA10-AS in
glioma cell growth, the changes in cell viability and proliferation
following silencing of HOXA10-AS were analyzed. The effects of
HOXA10-AS-knockdown were determined through lentivirus-mediated
expression of shRNA targeting HOXA10-AS. These target sequences
avoid overlap with the HOXA10 sequences, and do not cause silencing
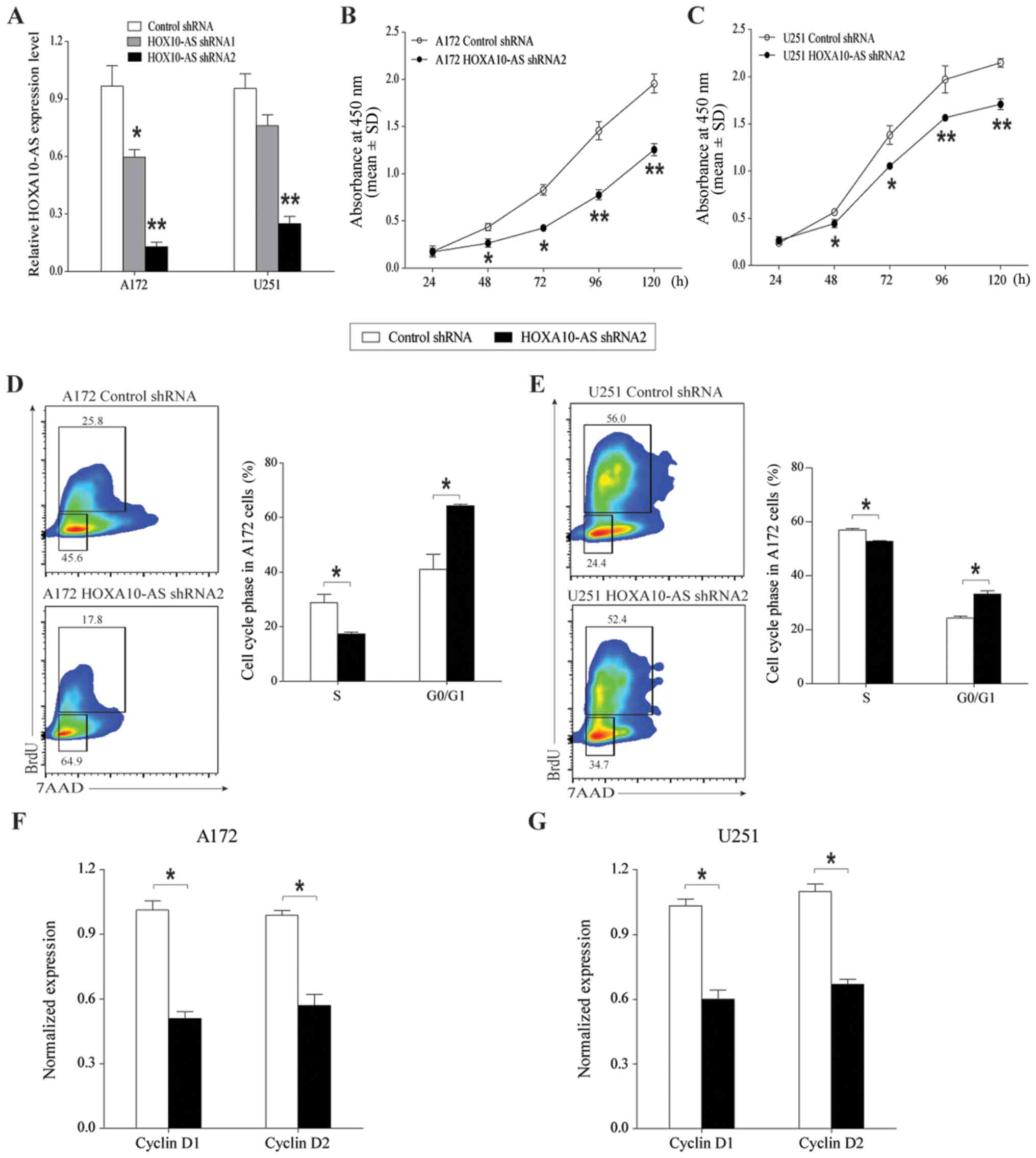
of the HOXA10 gene. The efficiency of HOXA10-AS
downregulation was determined by RT-qPCR analysis. The results
indicated that HOXA10-AS shRNA1 and shRNA2 induced significant
reductions in the expression levels of HOXA10-AS, with a greater
inhibitory effect with shRNA2 compared with shRNA1 in the two cell
lines (Fig. 2A).
The growth inhibitory effects of HOXA10-AS-knockdown
were confirmed by assessing the proliferation rate of cells for 5
consecutive days using MTT assays. In the scrambled control
shRNA-infected cells, the number of cells increased by >10-fold
over a 5-day culture period, whereas the number of
HOXA10-AS-knockdown cells increased more slowly than that of the
control cells. HOXA10-AS-knockdown cells cultured for 5 consecutive
days exhibited statistically significantly lower proliferation
rates than those of scrambled vector-containing cells (Fig. 2B and C).
To illustrate the underlining mechanisms by which
HOXA10-AS regulated cell growth, the cell cycle of A172 and U251
cell lines was then investigated using a BrdU array.
HOXA10-AS-knockdown markedly decreased the proportion of S phase
cells and increased the proportion of G0/G1
phase cells (Fig. 2D and E).
HOXA10-AS-knockdown led to arrest in the growth of cells (A172
cells: 56.7%; U251 cells: 36.6%), compared with the control cells.
Additionally, RT-qPCR demonstrated that the RNA expression levels
of cyclin D1 and cyclin D2, cell-cycle regulators, were decreased
in HOXA10-AS-knockdown glioma cells (Fig. 2F and G). These data indicated that
knockdown of HOXA10-AS inhibited glioma cell proliferation via
G0/G1 phase arrest and a decrease in DNA
synthesis.
Therefore, HOXA10-AS functions by promoting tumor
cell growth in glioma and may be a tumor oncogene.
Knockdown of HOXA10-AS induces glioma
cell apoptosis
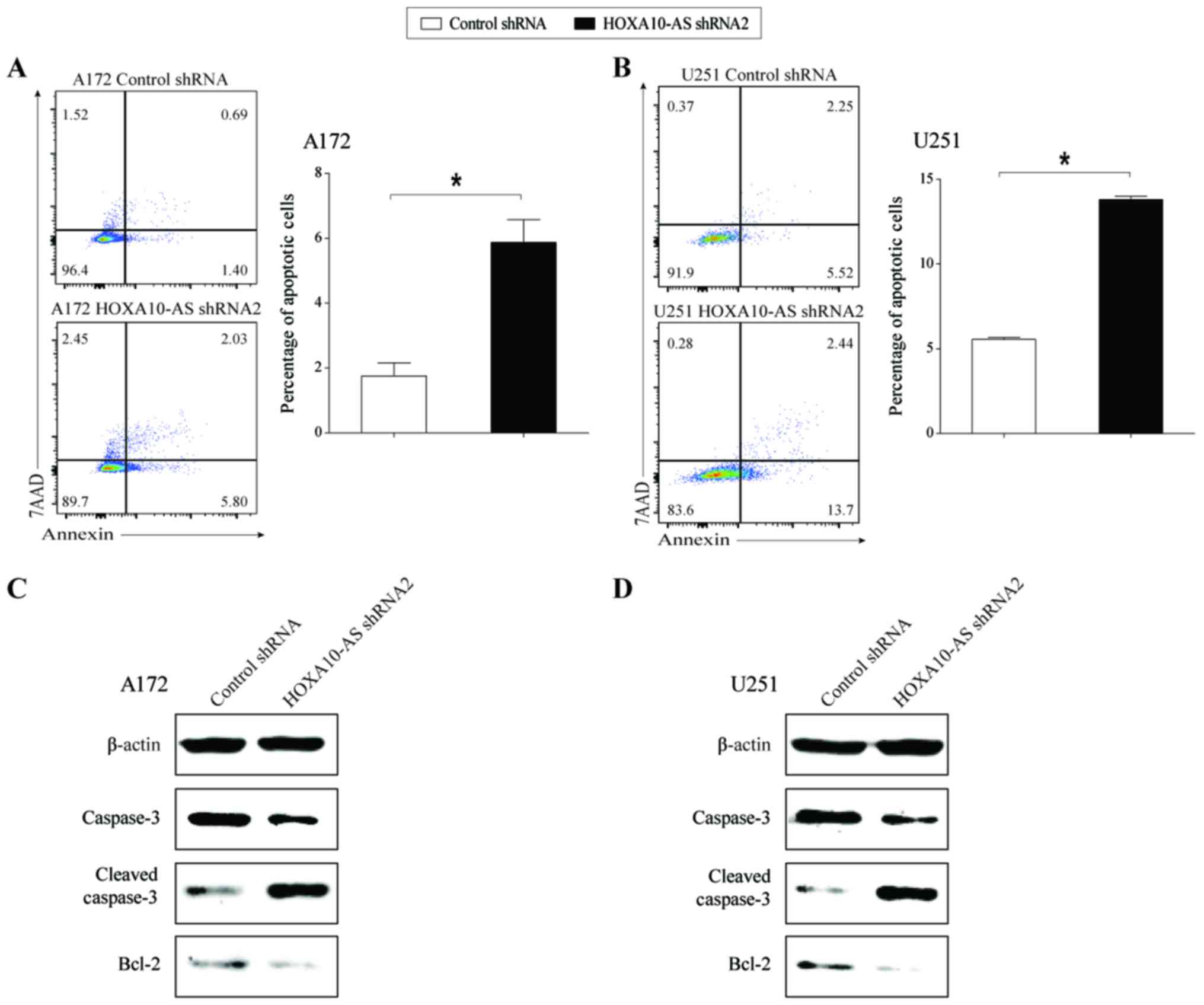
In order to examine the effect of
HOXA10-AS-knockdown on tumor cell apoptosis, cell apoptosis
analysis was performed using flow cytometry in HOXA10-AS-knockdown
and shRNA-control glioma cell lines. Knockdown of HOXA10-AS
resulted in a marked increase in the proportion of Annexin
V-positive and 7AAD-negative cells in knocked down A172 and U251
cells, compared with that of the scrambled shRNA-expressing cells
(Fig. 3A and B). This result
demonstrated that the knockdown of HOXA10-AS induced glioma cell
apoptosis.
In order to confirm the effect of HOXA10-AS on cell
apoptosis, the protein expression levels of apoptotic regulators,
including caspase, cleaved-caspase and Bcl-2 were investigated in
HOXA10-AS-knockdown cells. Western blot analysis demonstrated that
total caspase-3 and Bcl-2 protein expression levels were decreased
and the protein expression level of cleaved-caspase-3 was increased
following knockdown of HOXA10-AS in glioma cells (Fig. 3C and D). The expression trends of
three apoptosis-related proteins were consistent with the change in
the cell apoptosis rate following knockdown of HOXA10-AS.
These findings indicated that HOXA10-AS may serve a
role in promoting glioma cell survival.
HOXA10-AS activates HOXA10 gene
expression
In order to investigate the molecular mechanisms
underlying the tumor promoting effects of HOXA10-AS on glioma
cells, the ability of HOXA10-AS to regulate the transcription
activity of the HOXA10 gene was investigated in A172 and
U251 cells.
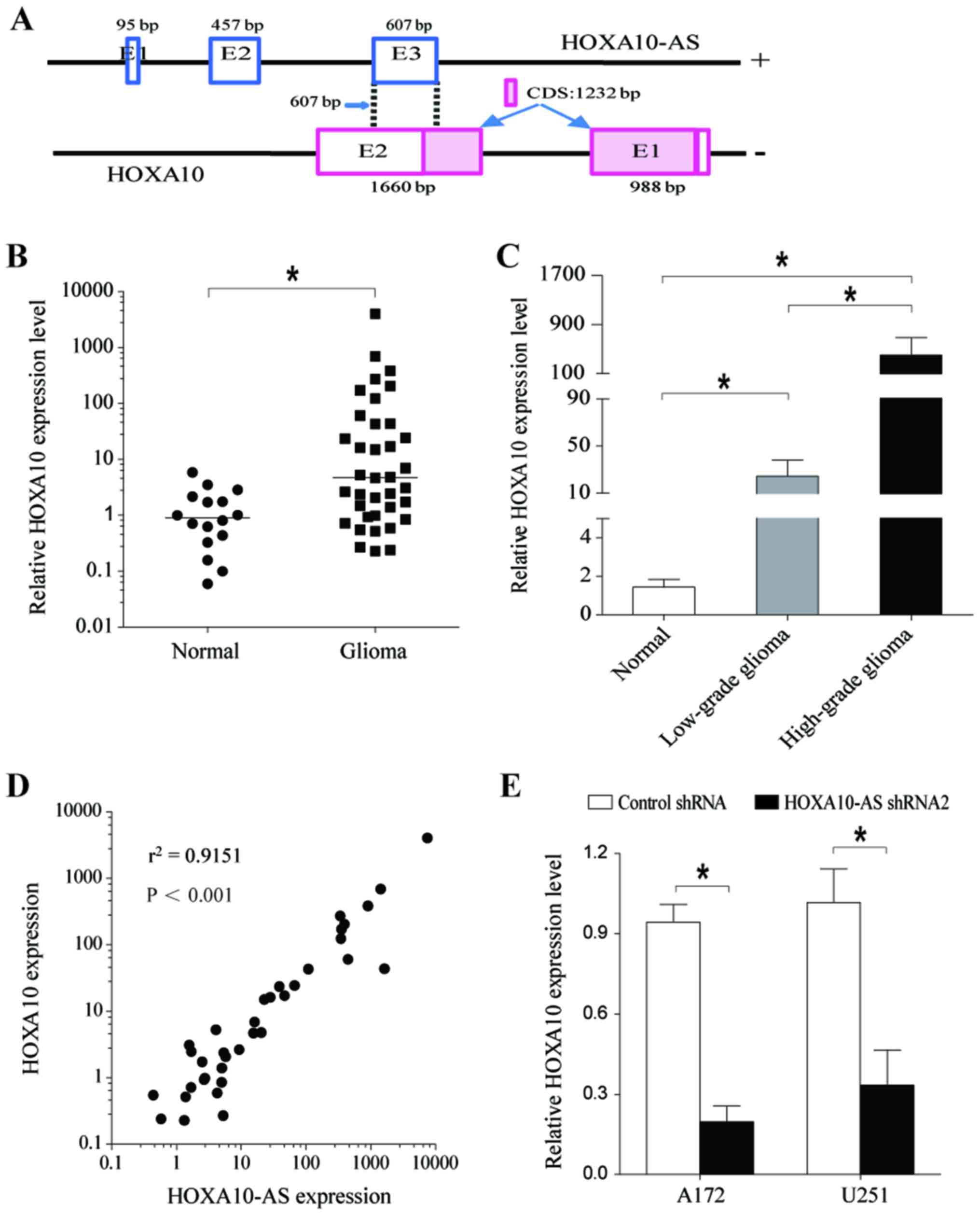
Sequence analysis demonstrated that exon 3 of
HOXA10-AS overlaps with the second exon of HOXA10 by 607 bp in an
antisense manner. The 607 bp of overlapping sequences is mostly
located in 3′-UTR of HOXA10 mRNA, excluding 15 bp (Fig. 4A). Antisense lncRNAs have been
implicated in the regulation of their overlapping sense transcripts
(32,33). As a first step in determining
whether HOXA10-AS regulates the expression of the HOXA10
gene, the present study analyzed the expression of HOXA10 in human
glioma tissues and compared it with that in normal brain tissues.
The results indicated that HOXA10 expression levels were
significantly increased in glioma tissues (Fig. 4B). Additionally, HOXA10 mRNA
expression levels in high-grade glioma tissues were higher than
those in low-grade glioma tissues, and HOXA10 expression was
associated with the degree of malignancy of the brain tumor
(Fig. 4C). Further statistical
analysis demonstrated a significant and positive correlation
between HOXA10 and HOXA10-AS gene expression
(Fig. 4D).
RT-qPCR analysis confirmed that HOXA10 expression
was significantly decreased at the mRNA level in A172 and U251
cells transfected with shHOXA10-AS (Fig. 4E). These results indicated that
HOXA10-AS regulated HOXA10 gene expression.
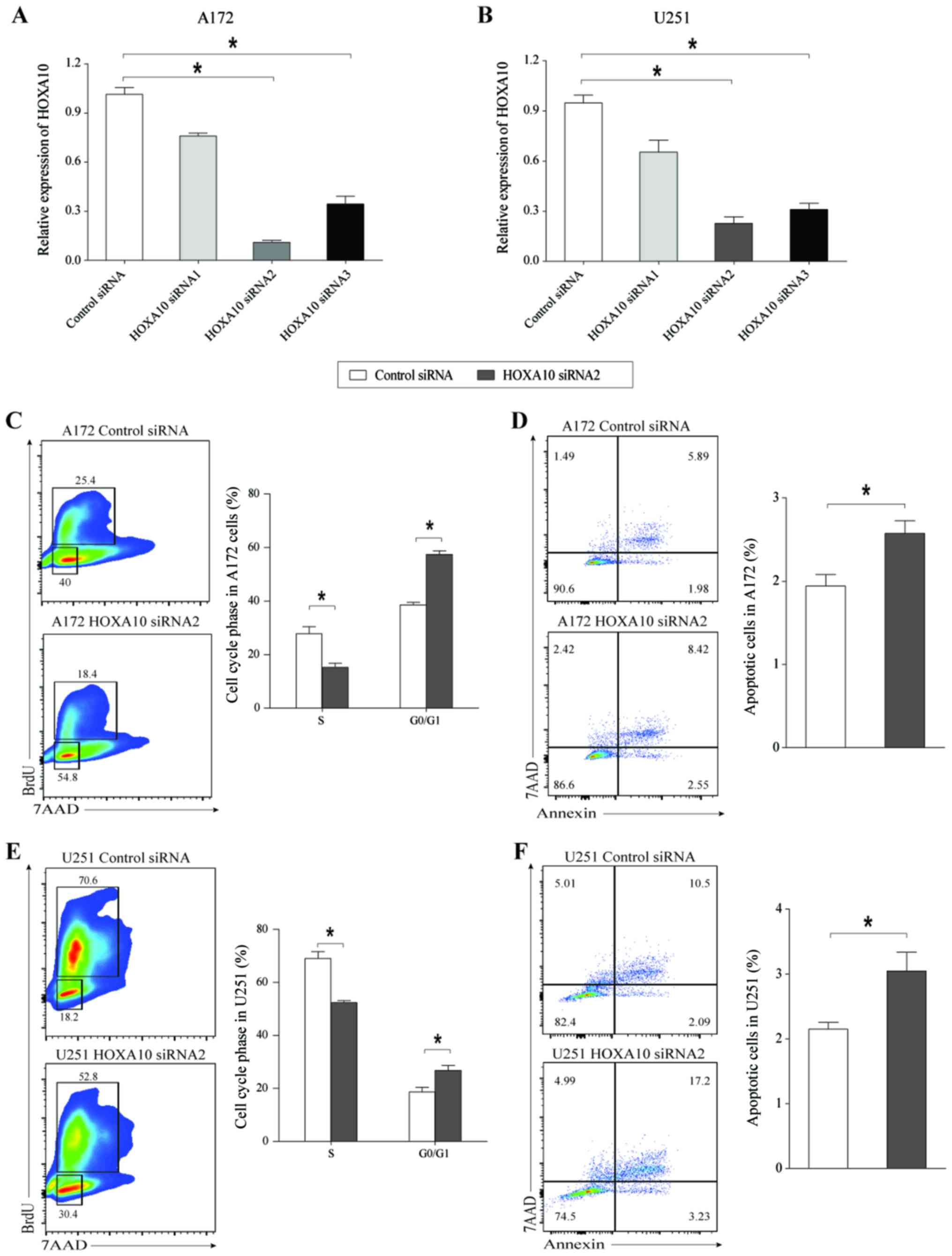
Knockdown of HOXA10 suppresses glioma
cell proliferation and induces cell apoptosis
To further examine the effect of HOXA10 on glioma
cell growth, the present study analyzed the changes in cell
proliferation and apoptosis following silencing of HOXA10 using
transcript-specific siRNAs in A172 and U251 cells. These siRNA
sequences avoid overlap with the sequences of HOXA10-AS, and do not
cause gene silencing of HOXA10-AS.
The present study first assessed HOXA10 mRNA
expression levels in A172 and U251 cells treated with siHOXA10 for
48 h. The results indicated that siHOXA10-2 and siHOXA10-3 induced
a significant and visible decrease in HOXA10 expression levels,
with a greater inhibitory effect achieved with siHOXA10-2 compared
with siHOXA10-3 in the two cell lines (Fig. 5A and B). Following knockdown of
HOXA10 in A172 cells, the percentage of cells in the S phase was
decreased and the percentage of cells in the
G0/G1 phase was increased (Fig. 5C), and caused a significant increase
in the rate of cell apoptosis (Fig.
5D). Similar results were also obtained in glioma cell line
U251 (Fig. 5E and F). These results
indicated that knockdown of HOXA10 inhibited glioma cell growth by
inducing G0/G1 phase arrest and cell
apoptosis. Therefore, we hypothesized that HOXA10-AS was able to
promote tumor cell growth by targeting HOXA10 in human glioma
cells.
Discussion
It is currently well-known that >75% of the human
genome is functional and encodes a large number of ncRNAs (34). Based on the ENCODE project, it is
estimated that the human genome encodes >28,000 distinct
lncRNAs, a number of which continue to be discovered and are yet to
be given a functional annotation (35). Therefore, revealing the biological
function of lncRNAs and identifying the disease-related lncRNAs are
one of the most investigated areas in the life sciences and medical
research fields.
To the best of our knowledge, the present study was
the first to report the role of HOXA10-AS in glioma. The original
results of qPCR analysis revealed significantly increased HOXA10-AS
expression in glioma tissues and cell lines, and demonstrated that
it was strongly associated with the histological tumor grades of
glioma tissues. We hypothesized that HOXA10-AS functions as a tumor
oncogene in glioma. In order to verify this hypothesis, the tumor
oncogenic roles of HOXA10-AS were investigated in glioma cell
lines. The present study investigated the function of HOXA10-AS in
glioma by HOXA10-AS-knockdown. Compared with the control group,
cell proliferation was significantly suppressed and the cell
apoptosis rate was increased in shHOXA10-AS-transfected glioma
cells. These data indicated that HOX10-AS served a crucial role in
glioma tumorigenesis.
Certain lncRNAs are cis-acting, affecting the
expression of neighboring genes. HOXA10-AS is a 1,161-bp spliced
and polyadenylated RNA, which transcripts the antisense strand of
the HOXA10 gene with a tail-to-tail overlap with HOXA10
transcript. Based on the results obtained in the clinical sample
analysis, the expression of HOXA10 was upregulated in glioma and
the expression of HOXA10-AS was correlated with that of HOXA10.
HOXA10, a member of the HOXA gene cluster, is a
pivotal transcriptional regulator of early embryonic development.
Previous studies have demonstrated that high expression of HOXA10
in several types of cancer is involved in regulating cell
proliferation, migration and invasion, and is correlated with a
poor prognosis (36–40). For example, HOXA10 significantly
promoted nasopharyngeal carcinoma cell proliferation and invasion
(36). Overexpression of HOXA10
predicted a poor prognosis in a subgroup of patients with
epithelial ovarian cancer (37) or
gastric cancer (38). HOXA10 serves
a role in the migration processes of medulloblastoma cells
(39).
Furthermore, Kurscheid et al (41) identified high expression of HOXA10
in glioblastoma tissues, compared with non-cancerous brain tissues
based on Affymetrix array data from TCGA, and demonstrated that
HOXA10 levels were significantly higher in glioblastoma stem cells.
The present study revealed that HOXA10 exerted effects on promoting
the proliferation and inducing the apoptosis of glioma cells. It
was further verified that the expression of HOXA10 was decreased in
HOXA10-AS-knockdown glioma cells compared with the control group,
indicating that HOXA10 may be activated by high HOXA10-AS
expression levels.
However, further studies are required to elucidate
the detailed mechanism of how HOXA10-AS regulates HOXA10. The
results of the present study demonstrated that HOXA10-AS is
overlapped with the 3′-UTR of HOXA10 mRNA. Therefore, the potential
HOXA10-AS/HOXA10 RNA hybrid may act to stabilize the HOXA10
transcript. It has been confirmed that Wrap53, as a natural p53
antisense transcript, is required for p53 stabilization via
Wrap53/p53 RNA interaction (42).
In addition, previous studies have reported that DNA methylation at
key regulatory CpGs in the promoters of HOXA10 was significantly
associated with HOXA10-signature expression (41). Antisense lncRNAs have been proposed
to cause DNA methylation (43,44).
Therefore, we hypothesized that the transcriptional activity of the
HOXA10 gene may be regulated in part by HOXA10-AS-directed
DNA hypomethylation. Several miRNAs were also known to regulate
HOXA10 expression in colorectal cancer (45), oral cancer (46), hepatocellular carcinoma (47) and breast cancer (48), suggesting that HOXA10-AS may
function as a competing endogenous RNA (ceRNA) to regulate
HOXA10 gene expression.
In conclusion, the results of the present study
provided the first evidence of HOXA10-AS values in glioma tissues,
which is a tumor oncogene. Increased expression of HOXA10-AS was
associated with malignancy status in glioma and knockdown of
HOXA10-AS may be a potential therapeutic strategy for glioma. In
vitro experiments demonstrated that silencing of HOXA10-AS
suppressed glioma cell proliferation and induced cell apoptosis.
Furthermore, the present study identified HOXA10 as a target of
HOXA10-AS and demonstrated that knockdown of HOXA10-AS
downregulated the mRNA expression levels of HOXA10. Additionally,
HOXA10 exhibited definitive oncogenic effects in glioma. Further
evaluation of the interactions between HOXA10-AS and HOXA10 is
required. The results of the present study indicated an important
role of HOXA10-AS in the promotion of the proliferation and
survival of glioma cells and that it may serve as a novel biomarker
and therapeutic target for glioma.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural
Science Foundation of Beijing City (7152098) and the Health Special
Science Foundation of Jilin Province (grant no.
2018SCZWSZX-014).
Availability of data and materials
The datasets supporting the findings of this study
are included within the article.
Authors' contributions
CYD, QL and JC performed the experiments. CYD, JC
and DHL collected and analyzed the data. QL and XYH conceived and
designed the study. QL and XYH wrote and edited the paper. CYD, JC
and DHL reviewed and revised the paper. All authors read and
approved the manuscript and agree to be accountable for all aspects
of the research in ensuring that the accuracy or integrity of any
part of the work were appropriately investigated and resolved.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the First Hospital of Jilin University and written
informed consent was obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Louis DN, Perry A, Reifenberger G, von
Deimling A, Figarella-Branger D, Cavenee WK, Ohgaki H, Wiestler OD,
Kleihues P and Ellison DW: The 2016 world health organization
classification of tumors of the central nervous system: A summary.
Acta Neuropathol. 131:803–820. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Omuro A and Deangelis LM: Glioblastoma and
other malignant gliomas: A clinical review. JAMA. 310:1842–1850.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Taylor LP: Diagnosis, treatment, and
prognosis of glioma: Five new things. Neurology. 75 18 Suppl
1:S28–S32. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hegi ME, Diserens AC, Gorlia T, Hamou MF,
de Tribolet N, Weller M, Kros JM, Hainfellner JA, Mason W, Mariani
L, et al: MGMT gene silencing and benefit from temozolomide
in glioblastoma. N Engl J Med. 352:997–1003. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mellinghoff IK, Wang MY, Vivanco I,
Haas-Kogan DA, Zhu S, Dia EQ, Lu KV, Yoshimoto K, Huang JH, Chute
DJ, et al: Molecular determinants of the response of glioblastomas
to EGFR kinase inhibitors. N Engl J Med. 353:2012–2024. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Claverie JM: Fewer genes, more noncoding
RNA. Science. 309:1529–1930. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Iyer MK, Niknafs YS, Malik R, Singhal U,
Sahu A, Hosono Y, Barrette TR, Prensner JR, Evans JR, Zhao S, et
al: The landscape of long noncoding RNAs in the human
transcriptome. Nature Genet. 47:199–208. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Derrien T, Johnson R, Bussotti G, Tanzer
A, Djebali S, Tilgner H, Guernec G, Martin D, Merkel A, Knowles DG,
et al: The GENCODE v7 catalog of human long noncoding RNAs:
Analysis of their gene structure, evolution, and expression. Genome
Res. 22:1775–1789. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Weidle UH, Birzele F, Kollmorgen G and
Rüger R: Long noncoding RNAs and their role in metastasis. Cancer
Genomics Proteomics. 14:143–160. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bhan A, Soleimani M and Mandal SS: Long
noncoding RNA and cancer: A new paradigm. Cancer Res. 77:3965–3981.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Alvarez-Dominguez JR and Lodish HF:
Emerging mechanisms of long noncoding RNA function during normal
and malignant hematopoiesis. Blood. 130:1965–1975. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Klingenberg M, Matsuda A, Diederichs S and
Patel T: Non-coding RNA in hepatocellular carcinoma: Mechanisms,
biomarkers and therapeutic targets. J Hepatol. 67:603–618. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang J, Ye C, Xiong H, Shen Y, Lu Y, Zhou
J and Wang L: Dysregulation of long non-coding RNA in breast
cancer: An overview of mechanism and clinical implication.
Oncotarget. 8:5508–5522. 2017.PubMed/NCBI
|
|
14
|
Xie W, Yuan S, Sun Z and Li Y: Long
noncoding and circular RNAs in lung cancer: Advances and
perspectives. Epigenomics. 8:1275–1287. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sun W, Yang Y, Xu C, Xie Y and Guo J:
Roles of long noncoding RNAs in gastric cancer and their clinical
applications. J Cancer Res Clin Oncol. 142:2231–2237. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hosseini ES, Meryet-Figuiere M,
Sabzalipoor H, Kashani HH, Nikzad H and Asemi Z: Dysregulated
expression of long noncoding RNAs in gynecologic cancers. Mol
Cancer. 16:1072017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Richtig G, Ehall B, Richtig E,
Aigelsreiter A, Gutschner T and Pichler M: Function and clinical
implications of long non-coding RNAs in melanoma. Int J Mol Sci.
18:pii: E715. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Brunner AL, Beck AH, Edris B, Sweeney RT,
Zhu SX, Li R, Montgomery K, Varma S, Gilks T, Guo X, et al:
Transcriptional profiling of long non-coding RNAs and novel
transcribed regions across a diverse panel of archived human
cancers. Genome Biol. 13:R752012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bartonicek N, Maag JL and Dinger ME: Long
noncoding RNAs in cancer: Mechanisms of action and technological
advancements. Mol Cancer. 15:432016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shi T, Gao G and Cao Y: Long noncoding
RNAs as novel biomarkers have a promising future in cancer
diagnostics. Dis Markers. 2016:90851952016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang JX, Han L, Bao ZS, Wang YY, Chen LY,
Yan W, Yu SZ, Pu PY, Liu N, You YP, et al: HOTAIR, a cell
cycle-associated long noncoding RNA and a strong predictor of
survival, is preferentially expressed in classical and mesenchymal
glioma. Neuro Oncol. 15:1595–1603. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang B, Wei ZY, Wang BQ, Yang HC, Wang JY
and Bu XY: Down-regulation of the long noncoding RNA-HOX transcript
antisense intergenic RNA inhibits the occurrence and progression of
glioma. J Cell Biochem. 119:2278–2287. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Wang Y, Li J, Zhang Y, Yin H and
Han B: CRNDE, a long-noncoding RNA, promotes glioma cell growth and
invasion through mTOR signaling. Cancer Lett. 367:122–128. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jiang X, Yan Y, Hu M, Chen X, Wang Y, Dai
Y, Wu D, Wang Y, Zhuang Z and Xia H: Increased level of H19 long
noncoding RNA promotes invasion, angiogenesis, and stemness of
glioblastoma cells. J Neurosurg. 124:129–136. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Matouk IJ, Mezan S, Mizrahi A, Ohana P,
Abu-Lail R, Fellig Y, Degroot N, Galun E and Hochberg A: The
oncofetal H19 RNA connection: Hypoxia, p53 and cancer. Biochim
Biophys Acta. 1803:443–451. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yao Y, Ma J, Xue Y, Wang P, Li Z, Liu J,
Chen L, Xi Z, Teng H, Wang Z, et al: Knockdown of long non-coding
RNA XIST exerts tumor-suppressive functions in human glioblastoma
stem cells by up-regulating miR-152. Cancer Lett. 359:75–86. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yao J, Zhou B, Zhang J, Geng P, Liu K, Zhu
Y and Zhu W: A new tumor suppressor LncRNA ADAMTS9-AS2 is regulated
by DNMT1 and inhibits migration of glioma cells. Tumour Biol.
35:7935–7944. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang P, Liu YH, Yao YL, Li Z, Li ZQ, Ma J
and Xue YX: Long non-coding RNA CASC2 suppresses malignancy in
human gliomas by miR-21. Cell Signal. 27:275–282. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang P, Ren Z and Sun P: Overexpression of
the long non-coding RNA MEG3 impairs in vitro glioma cell
proliferation. J Cell Biochem. 113:1868–1874. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Han Y, Wu Z, Wu T, Huang Y, Cheng Z, Li X,
Sun T, Xie X, Zhou Y and Du Z: Tumor-suppressive function of long
noncoding RNA MALAT1 in glioma cells by downregulation of MMP2 and
inactivation of ERK/MAPK signaling. Cell Death Dis. 7:e21232016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Katayama S, Tomaru Y, Kasukawa T, Waki K,
Nakanishi M, Nakamura M, Nishida H, Yap CC, Suzuki M, Kawai J, et
al: Antisense transcription in the mammalian transcriptome.
Science. 309:1564–1566. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Werner A, Carlile M and Swan D: What do
natural antisense transcripts regulate? RNA Biol. 6:43–48. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sanfilippo PG and Hewitt AW: Translating
the ENCyclopedia of DNA elements project findings to the clinic:
ENCODE's implications for eye disease. Clin Exp Ophthalmol.
42:78–83. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tragante V, Moore JH and Asselberg FW: The
ENCODE project and perspectives on pathways. Genet Epidemiol.
38:275–280. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shen ZH, Zhao KM and Du T: HOXA10 promotes
nasopharyngeal carcinoma cell proliferation and invasion via
inducing the expression of ZIC2. Eur Rev Med Pharmacol Sci.
21:945–952. 2017.PubMed/NCBI
|
|
37
|
Eoh KJ, Kim HJ, Lee JY, Nam EJ, Kim S, Kim
SW and Kim YT: Dysregulated expression of homeobox family genes may
influence survival outcomes of patients with epithelial ovarian
cancer: Analysis of data from The Cancer Genome Atlas. Oncotarget.
8:70579–70585. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lim JY, Yoon SO, Seol SY, Hong SW, Kim JW,
Choi SH, Lee JS and Cho JY: Overexpression of miR-196b and HOXA10
characterize a poor-prognosis gastric cancer subtype. World J
Gastroenterol. 19:7078–7088. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bonfim-Silva R, Melo Ferreira FU, Thomé
CH, Abraham KJ, De Souza FAL, Ramalho FS, Machado HR, De Oliveira
RS, Cardoso AA, Covas DT, et al: Functional analysis of
HOXA10 and HOXB4 in human medulloblastoma cell lines.
Int J Oncol. 51:1929–1940. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Carrera M, Bitu CC, de Oliveira CE,
Cervigne NK, Graner E, Manninen A, Salo T and Coletta RD: HOXA10
controls proliferation, migration and invasion in oral squamous
cell carcinoma. Int J Clin Exp Pathol. 8:3613–3623. 2015.PubMed/NCBI
|
|
41
|
Kurscheid S, Bady P, Sciuscio D, Samarzija
I, Shay T, Vassallo I, Criekinge WV, Daniel RT, van den Bent MJ,
Marosi C, et al: Chromosome 7 gain and DNA hypermethylation at the
HOXA10 locus are associated with expression of a stem cell related
HOX-signature in glioblastoma. Genome Biol. 16:162015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Mahmoudi S, Henriksson S, Corcoran M,
Méndez-Vidal C, Wiman KG and Farnebo M: Wrap53, a natural p53
antisense transcript required for p53 induction upon DNA damage.
Mol Cell. 33:462–471. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tufarelli C, Stanley JA, Garrick D, Sharpe
JA, Ayyub H, Wood WG and Higgs DR: Transcription of antisense RNA
leading to gene silencing and methylation as a novel cause of human
genetic disease. Nat Genet. 34:157–165. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sun BK, Deaton AM and Lee JT: A transient
heterochromatic state in Xist preempts X inactivation choice
without RNA stabilization. Mol Cell. 21:617–628. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sun S, Su C, Zhu Y, Li H, Liu N, Xu T, Sun
C and Lv Y: MicroRNA-544a regulates migration and invasion in
colorectal cancer cells via regulation of homeobox A10. Dig Dis
Sci. 61:2535–2544. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Libório-Kimura TN, Jung HM and Chan EK:
miR-494 represses HOXA10 expression and inhibits cell proliferation
in oral cancer. Oral Oncol. 51:151–157. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xiao ZD, Jiao CY, Huang HT, He LJ, Zhao
JJ, Lu ZY and Liu LX: miR-218 modulate hepatocellular carcinoma
cell proliferation through PTEN/AKT/PI3K pathway and HoxA10. Int J
Clin Exp Pathol. 7:4039–4044. 2014.PubMed/NCBI
|
|
48
|
Chen Y, Zhang J, Wang H, Zhao J, Xu C, Du
Y, Luo X, Zheng F, Liu R, Zhang H, et al: miRNA-135a promotes
breast cancer cell migration and invasion by targeting
HOXA10. BMC Cancer. 12:1112012. View Article : Google Scholar : PubMed/NCBI
|