Introduction
Pancreatic cancer was revealed to be the fourth
leading cause of cancer-associated mortality in the USA in 2016,
and the estimated number of deaths caused by pancreatic cancer for
that year is 21,450 in men and 20,330 in women (1). Numerous efforts and chemotherapeutic
advancements have been made to improve the efficacy of pancreatic
cancer treatment; however, the results remain unsatisfactory
(2–4). Therefore, there is an urgent need to
develop novel therapeutic agents for pancreatic cancer.
It is well documented that transmembrane tyrosine
kinases are strongly associated with the proliferation and
metastasis of numerous types of human cancer (5,6). The
phosphoinositide 3-kinase (PI3K)/protein kinase B (AKT)/mammalian
target of rapamycin (mTOR) and Ras/Raf/mitogen-activated protein
kinase kinase (MEK)/extracellular signal-regulated kinase (ERK)
pathways are two of the most frequently dysregulated kinase
cascades in human cancer. These pathways signify important signal
transduction mechanisms, which accelerate the proliferation and
survival of cancers via activation of growth factor receptors, such
as insulin-like growth factor 1 receptor (IGF1R), vascular
endothelial growth factor receptor (VEGFR) and epidermal growth
factor receptor (EGFR) (7,8). Either through somatic mutations or
epigenetic modifications, the individual downstream constituents of
these signaling cascades can induce tumorigenesis and resistance to
anticancer treatments, and these constituents have been reported to
be frequently transformed in malignant tumors (9).
Tanshinone-IIA (Tan-IIA) is an active component of
the plant-derived traditional Chinese medicine Danshen, which has
been reported to possess anticancer potential (2,3).
Tan-IIA is able to inhibit the protein expression levels of MCL1,
B-cell lymphoma 2 (Bcl-2) family apoptosis regulator; tumor
protein, translationally-controlled 1 and Bcl-extra large, in order
to destroy mitochondrial function and increase Bcl-2-associated X
protein and caspase-3 expression, thus inducing apoptosis of BxPC-3
human pancreatic cancer cells (10). Furthermore, Tan-IIA increases
protein kinase R-like endoplasmic reticulum kinase, activating
transcription factor 6, caspase-12 and CCAAT-enhancer-binding
protein homologous protein expression, in order to induce apoptosis
of BxPC-3 pancreatic cancer cells in vitro (11). It has also been reported that
Tan-IIA may exert cytotoxic effects on MiaPaCa-2 human pancreatic
cancer cells (12). The present
study aimed to detect the protein expression levels of IGF1R,
VEGFR, EGFR, PI3K, AKT, mTOR, Ras, Raf, MEK, ERK and phosphatase
and tensin homolog (PTEN) in MiaPaCa-2 human pancreatic cancer
cells following treatment with Tan-IIA.
Materials and methods
Chemicals and reagents
Tan-IIA (molecular formula,
C19H18O3; CAS No., 568-72-9) was
purchased from Sigma-Aldrich; Merck KGaA (Darmstadt, Germany). The
MiaPaCa-2 human pancreatic cancer cell line (BCRC No. 60139) was
obtained from the Food Industry Research and Development Institute
(Hsinchu, Taiwan). MTT, sodium deoxycholate, leupeptin, Triton
X-100, Tris/HCl, RNase A, sodium orthovanadate, sodium pyruvate,
HEPES and mouse anti-β-actin (cat no. A5441, MW 43 kDa) were
obtained from Sigma-Aldrich; Merck KGaA. Dimethyl sulfoxide (DMSO),
potassium phosphate and TE buffer were purchased from Merck KGaA.
Fetal bovine serum (FBS), Dulbecco's modified Eagle's medium
(DMEM), trypsin-EDTA, penicillin-streptomycin and glutamine were
obtained from Gibco; Thermo Fisher Scientific, Inc. (Waltham, MA,
USA). Buffer (10X TG-SDS), Tween-20 and glycine were obtained from
Amresco, LLC (Solon, OH, USA). BioMax film was obtained from Kodak
(Rochester, NY, USA). Other materials and reagents not specified
were obtained from Sigma-Aldrich; Merck KGaA or Merck KGaA.
Antibodies
Anti-VEGFR (cat. no. NB100-527, MW 151 kDa) was
obtained from Novus Biologicals, LLC (Littleton, CO, USA);
anti-EGFR (cat. no. 2239, MW 175 kDa), anti-IGF1R (cat. no. 3018,
MW 95 kDa), anti-Ras (cat. no. 3339, MW 21 kDa), anti-Raf (cat. no.
12552, MW 75 kDa), anti-MEK (cat. no. 9126, MW 45 kDa), anti-ERK
(cat. no. 4695, MW 42–44 kDa), anti-PI3K (cat. no. 4292, MW 85
kDa), anti-AKT (cat. no. 3063, MW 60 kDa), anti-mTOR (cat. no.
2983, MW 289 kDa) and anti-PTEN cat. no. 9559, MW 54 kDa)
antibodies were all obtained from Cell Signaling Technology, Inc.
(Danvers, MA, USA).
Cell culture
The MiaPaCa-2 cells were maintained in DMEM
containing 10% FBS and 1% penicillin/streptomycin (10,000 U/ml
penicillin, 10 mg/ml streptomycin) at 37°C in a humidified
atmosphere containing 5% CO2.
Cytotoxicity assay
The cytotoxicity of Tan-IIA in MiaPaCa-2 cells was
evaluated by MTT assay in triplicate, as previously described
(10). Briefly, the MiaPaCa-2 cells
were plated in 96-well plates at a density of 1×104
cells/well and were treated with various concentrations (0, 1, 3,
9, 15, 30 and 60 µg/ml) of Tan-IIA for different durations (24, 48
and 72 h). Subsequently, the cells were incubated with 100 µl MTT
(1 mg/ml) in fresh complete DMEM for 2 h. The surviving cells
converted MTT to formazan, as presented by a blue-purple color when
dissolved in DMSO at 37°C for 2 h. Absorbance was measured using an
ELISA microplate reader at 590 nm. The relative percentage of cell
viability was calculated by dividing the absorbance of treated
cells by that of the control in each experiment, using the
following formula: Proliferation rate (%) = (OD test - OD blank) ×
100, where OD test and OD blank are the optical density of the test
substance and the blank control, respectively.
Western blotting
The effects of Tan-IIA on the protein expression
levels of EGFR, IGF1R, VEGFR, Ras, PI3K, AKT, mTOR, Raf, MEK, ERK
and PTEN were detected in MiaPaCa-2 cells. The MiaPaCa-2 cells were
treated with various concentrations of Tan-IIA (0, 3, 6 and 12
µg/ml) for 48 h, or with various concentrations of Tan-IIA (0, 1.5,
3 and 6 µg/ml) for 72 h, after which the expression levels of EGFR,
IGF1R, VEGFR, Ras, PI3K, AKT, mTOR, Raf, MEK, ERK and PTEN were
evaluated by western blotting. Alternatively, MiaPaCa-2 cells were
treated with Tan-IIA (6 µg/ml) for various durations (0, 24, 48 and
72 h), after which the proteins expression levels of EGFR, IGF1R,
VEGFR, Ras, PI3K, AKT, mTOR, Raf, MEK, ERK and PTEN were evaluated
by western blotting.
Western blot analysis was conducted, as previously
described (10). Briefly, after
treatment, the cells were lysed in ice-cold whole cell extract
buffer containing protease inhibitors (cat. no. 20-188; Merck
KGaA). The lysate was agitated for 30 min at 4°C and centrifuged at
12,281 × g for 10 min. Protein concentration was measured using a
bicinchoninic acid protein assay kit (Pierce; Thermo Fisher
Scientific, Inc.). Equal amounts of protein (10 µg) were then
subjected to electrophoresis using 10–15% SDS-polyacrylamide gels.
To verify equal protein loading (10 µg) and transfer, proteins were
transferred to polyvinylidene difluoride membranes, which were
blocked overnight at 4°C using blocking buffer [5% non-fat dried
milk in solution containing 50 mM Tris/HCl (pH 8.0), 2 mM
CaCl2, 80 mM sodium chloride, 0.05% Tween-20 and 0.02%
sodium azide]. The membranes were then incubated with the specific
primary antibodies (1:1,000) for 2 h at 25°C and were washed three
times with Tris-buffered saline-0.05% Tween-20 (TBST).
Subsequently, the membranes were incubated with anti-rabbit (cat.
no. sc-2004) or anti-mouse (cat. no. sc-2005) immunoglobulin
G-horseradish peroxidase-conjugated secondary antibodies (1:5,000;
Santa Cruz Biotechnology Inc., Dallas, TX, USA). The membranes were
then washed a further three times for 10 min with TBST. Finally,
the protein bands were visualized on X-ray film and were analyzed
using the enhanced chemiluminescence detection system (PerkinElmer,
Inc., Waltham, MA, USA) and ImageJ 1.4.4p (National Institutes of
Health, Bethesda, MD, USA) software analysis. β-actin was used as
an internal control in all western blots. Results are presented as
the means ± standard deviation of three experiments.
Statistical analysis
The statistical significance of the differences
between groups was assessed using SPSS software version 20 (IBM
Corp., Armonk, NY, USA). Data were analyzed using one-way analysis
of variance followed by Dunnett's test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Cytotoxicity of Tan-IIA in MiaPaCa-2
cells
MiaPaCa-2 cells were cultured with various
concentrations (0, 1, 3, 9, 15, 30 and 60 µg/ml) of Tan-IIA for
different durations (24, 48 and 72 h). Following Tan-IIA treatment
for 24, 48 and 72 h, the half maximum inhibitory concentration
values for Tan-IIA were 14.2, 6.4 and 2.9 µg/ml, respectively.
These results revealed that Tan-IIA may inhibit the proliferation
of MiaPaCa-2 human pancreatic cancer cells in a time- and
dose-dependent manner (Fig. 1).
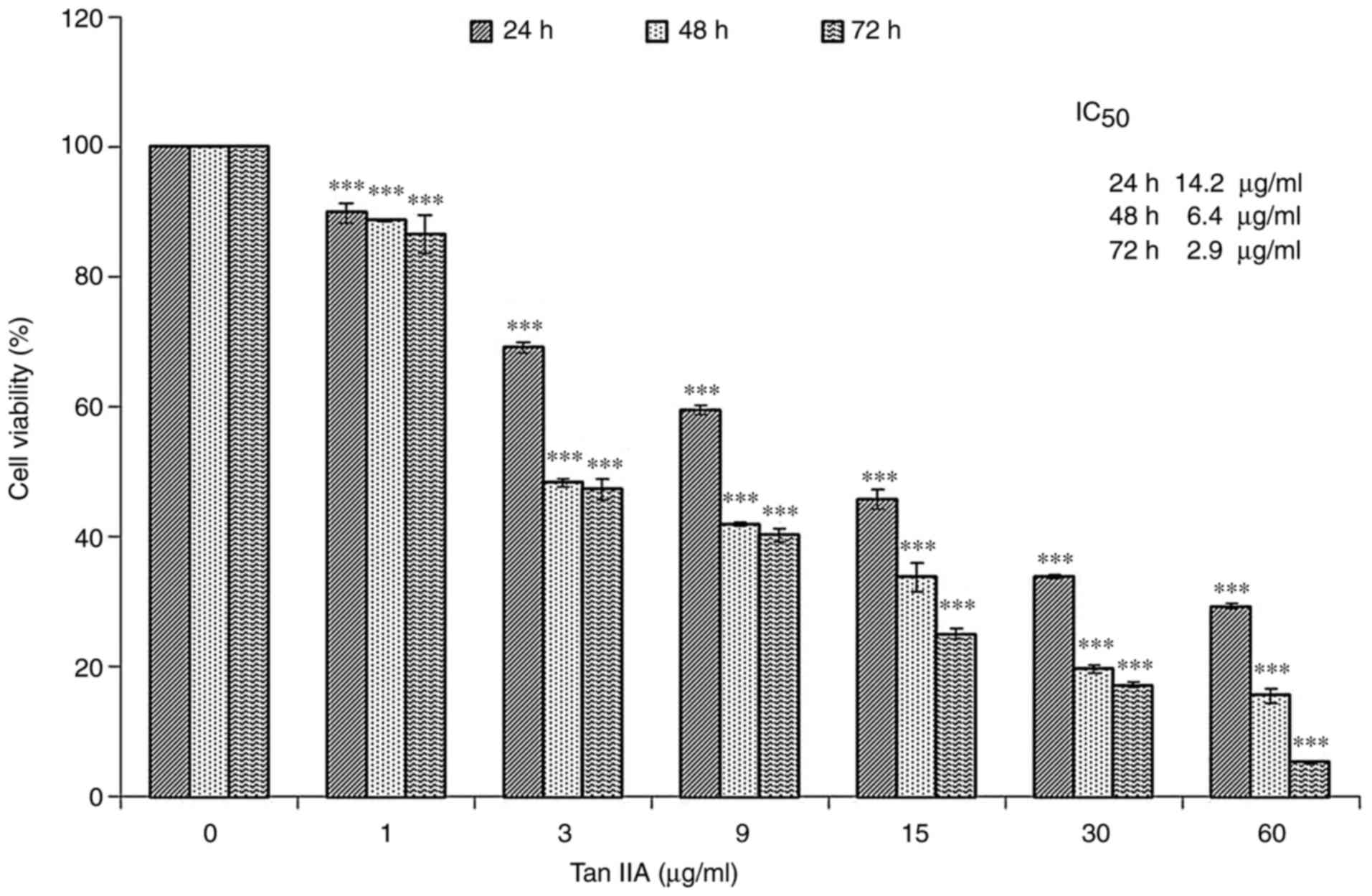 | Figure 1.Cytotoxicity of Tan-IIA in MiaPaCa-2
cells. Cytotoxicity of Tan-IIA in MiaPaCa-2 cells was determined
using the MTT assay. MiaPaCa-2 cells were cultured with various
concentrations (0, 1, 3, 9, 15, 30 and 60 µg/ml) of Tan-IIA for
different durations (24, 48 and 72 h). The results revealed that
Tan-IIA inhibited the proliferation of MiaPaCa-2 human pancreatic
cancer cells in a time- and dose-dependent manner. The
IC50 values were 14.2, 6.4 and 2.9 µg/ml, respectively.
Results are presented as the means ± standard deviation of three
experiments. ***P<0.001 vs. the control group. IC50,
half-maximum inhibitory concentration; Tan-IIA, Tanshinone-IIA. |
Dose-dependent effects of Tan-IIA on
the protein expression levels of EGFR, IGF1R, VEGFR, Ras, Raf, MEK,
ERK, PI3K, AKT, mTOR and PTEN in MiaPaCa-2 cells
MiaPaCa-2 cells were treated with various
concentrations (0, 3, 6 and 12 µg/ml) of Tan-IIA for 48 h or with
various concentrations (0, 1.5, 3 and 6 µg/ml) of Tan-IIA for 72 h,
after which the protein expression levels were evaluated by western
blot analysis (Fig. 2). The results
revealed that Tan-IIA significantly decreased the protein
expression levels of Ras (Fig. 2D and
M), Raf (Fig. 2E and L), MEK
(Fig. 2F and M), ERK (Fig. 2G and M), AKT (Fig. 2I and L) and PTEN (Fig. 2K and L) at 48 and 72 h. However, the
protein expression levels of IGF1R (Fig. 2C and M) and PI3K (Fig. 2H and L) were significantly decreased
only in response to 48 h treatment. The protein expression levels
of mTOR (Fig. 2J and L) were
significantly decreased only in response to 72 h treatment. The
protein expression levels of EGFR (Fig.
2A and L) were significantly decreased in response to all
concentrations of Tan-IIA after 72 h and in response to 3 µg/ml
Tan-IIA after 48 h.
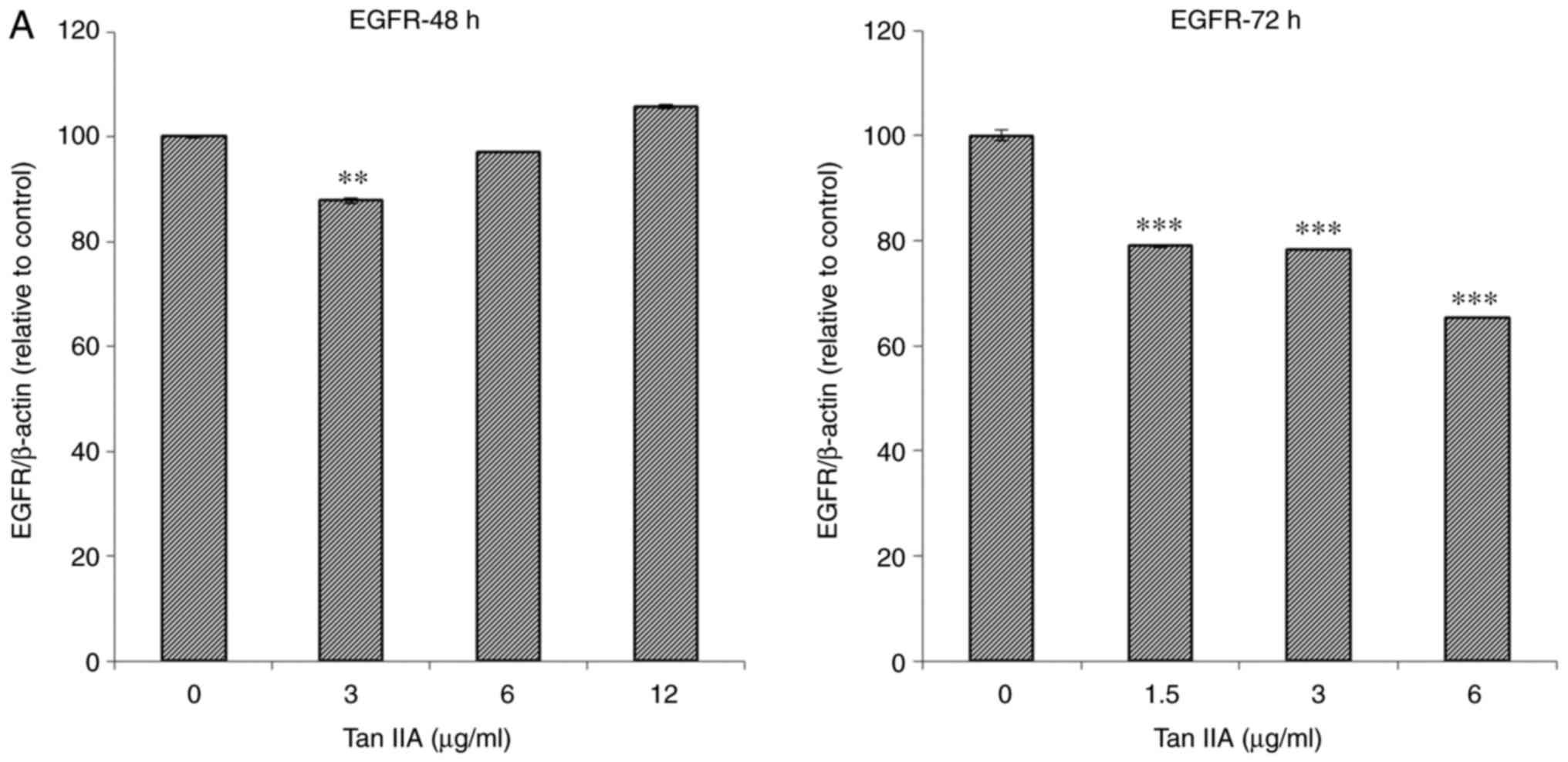 | Figure 2.Dose-dependent effects of Tan-IIA on
the protein expression levels of EGFR, IGF1R, VEGFR, Ras, Raf, MEK,
ERK, PI3K, AKT, mTOR and PTEN in MiaPaCa-2 cells. The MiaPaCa-2
cells were treated with various concentrations (0, 3, 6 and 12
µg/ml) of Tan-IIA for 48 h or with various concentrations (0, 1.5,
3 and 6 µg/ml) of Tan-IIA for 72 h, and the protein expression
levels were evaluated by western blot analysis. Protein expression
levels of (A) EGFR. **P<0.01, ***P<0.001 compared with the
control group. AKT, protein kinase B; EGFR, epidermal growth factor
receptor; ERK, extracellular signal-regulated kinase; IGF1R,
insulin-like growth factor 1 receptor; MEK, mitogen-activated
protein kinase kinase; mTOR, mammalian target of rapamycin; PI3K,
phosphoinositide 3-kinase; PTEN, phosphatase and tensin homolog;
VEGFR, vascular endothelial growth factor receptor. Protein
expression levels of (B) VEGFR, (C) IGF1R, (D) Ras, (E) Raf.
***P<0.001 compared with the control group. AKT, protein kinase
B; EGFR, epidermal growth factor receptor; ERK, extracellular
signal-regulated kinase; IGF1R, insulin-like growth factor 1
receptor; MEK, mitogen-activated protein kinase kinase; mTOR,
mammalian target of rapamycin; PI3K, phosphoinositide 3-kinase;
PTEN, phosphatase and tensin homolog; VEGFR, vascular endothelial
growth factor receptor. Protein expression levels of (F) MEK, (G)
ERK, (H) PI3K, (I) AKT **P<0.01, ***P<0.001 compared with the
control group. AKT, protein kinase B; EGFR, epidermal growth factor
receptor; ERK, extracellular signal-regulated kinase; IGF1R,
insulin-like growth factor 1 receptor; MEK, mitogen-activated
protein kinase kinase; mTOR, mammalian target of rapamycin; PI3K,
phosphoinositide 3-kinase; PTEN, phosphatase and tensin homolog;
VEGFR, vascular endothelial growth factor receptor. (J) mTOR and
(K) PTEN. (L and M) Representative images of western blots.
**P<0.01, ***P<0.001 compared with the control group. AKT,
protein kinase B; EGFR, epidermal growth factor receptor; ERK,
extracellular signal-regulated kinase; IGF1R, insulin-like growth
factor 1 receptor; MEK, mitogen-activated protein kinase kinase;
mTOR, mammalian target of rapamycin; PI3K, phosphoinositide
3-kinase; PTEN, phosphatase and tensin homolog; VEGFR, vascular
endothelial growth factor receptor. |
Time-dependent effects of Tan-IIA on
the protein expression levels of EGFR, IGF1R, VEGFR, Ras, Raf, MEK,
ERK, PI3K, AKT, mTOR and PTEN in MiaPaCa-2 cells
MiaPaCa-2 cells were treated with Tan-IIA (6 µg/ml)
for various durations (0, 24, 48 and 72 h) and the protein
expression levels were evaluated by western blot analysis (Fig. 3). The results revealed that Tan-IIA
significantly decreased the protein expression levels of EGFR
(Fig. 3A and L), IGF1R (Fig. 3B and L), Raf (Fig. 3E and L) and MEK (Fig. 3F and L) in a time-dependent manner.
However, the protein expression levels of ERK (Fig. 3G and L) and AKT (Fig. 3I and M) were significantly decreased
only at 48 and 72 h; the protein expression levels of VEGFR
(Fig. 3C and L) and mTOR (Fig. 3J and M) were significantly decreased
only at 72 h; and PTEN (Fig. 3K and
M) was only significantly decreased at 24 h. Tan-IIA
significantly decreased the protein expression levels of Ras
(Fig. 3D and L) and PI3K (Fig. 3H and M) at 24, 48 and 72 h, but
expression was slightly higher at 48 h than at 24 and 72 h.
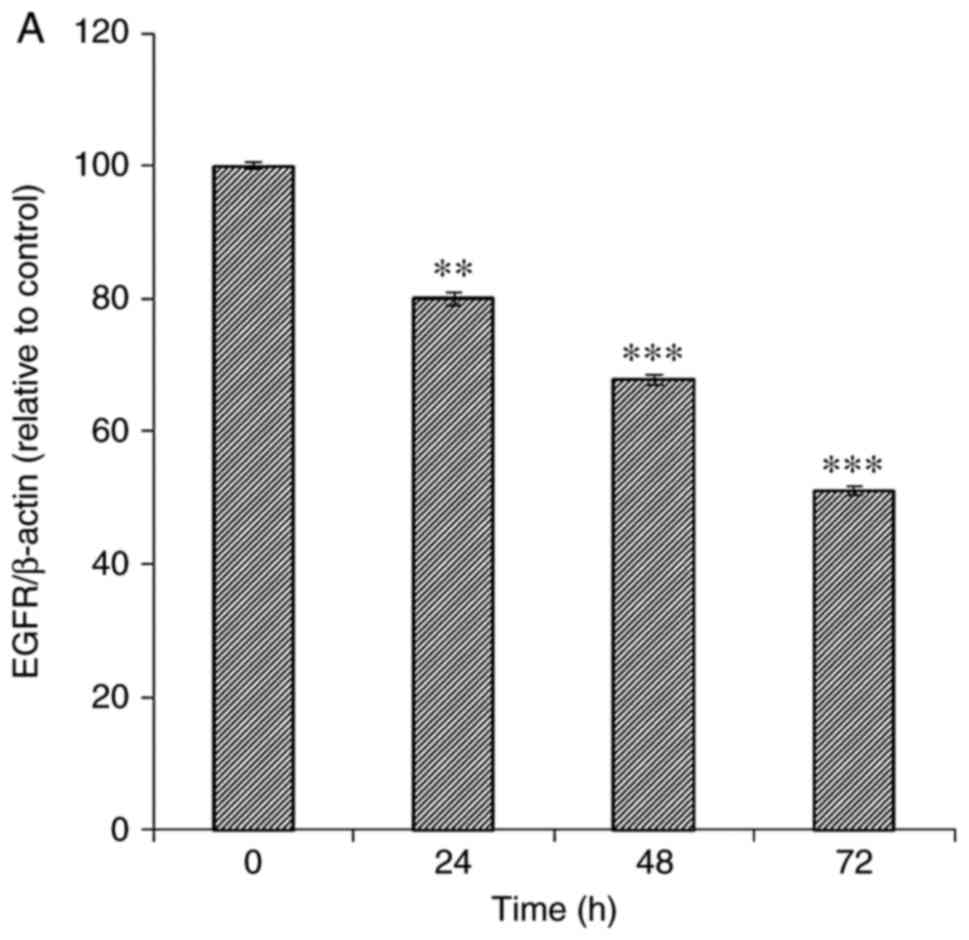 | Figure 3.Time-dependent effects of Tan-IIA on
the protein expression levels of EGFR, IGF1R, VEGFR, Ras, Raf, MEK,
ERK, PI3K, AKT, mTOR and PTEN in MiaPaCa-2 cells. MiaPaCa-2 cells
were treated with Tan-IIA (6 µg/ml) for various durations (0, 24,
48 and 72 h) and the protein expression levels were evaluated by
western blotting. Protein expression levels of (A) EGFR, (B) IGF1R,
(C) VEGFR, (D) Ras, (E) Raf, (F) MEK *P<0.05, **P<0.01,
***P<0.001 compared with the control group. AKT, protein kinase
B; EGFR, epidermal growth factor receptor; ERK, extracellular
signal-regulated kinase; IGF1R, insulin-like growth factor 1
receptor; MEK, mitogen-activated protein kinase kinase; mTOR,
mammalian target of rapamycin; PI3K, phosphoinositide 3-kinase;
PTEN, phosphatase and tensin homolog; VEGFR, vascular endothelial
growth factor receptor. Protein expression levels of (G) ERK, (H)
PI3K, (I) AKT, (J) mTOR and (K) PTEN. (L and M) Representative
images of western blots. *P<0.05, ***P<0.001 compared with
the control group. AKT, protein kinase B; EGFR, epidermal growth
factor receptor; ERK, extracellular signal-regulated kinase; IGF1R,
insulin-like growth factor 1 receptor; MEK, mitogen-activated
protein kinase kinase; mTOR, mammalian target of rapamycin; PI3K,
phosphoinositide 3-kinase; PTEN, phosphatase and tensin homolog;
VEGFR, vascular endothelial growth factor receptor. |
Discussion
Ras is the most frequently mutated oncogene in human
cancer (13). ERK belongs to the
mitogen-activated protein kinase (MAPK) pathway, and dysregulation
of the Ras/Raf/MAPK signal pathway has been well documented in
tumorigenesis (14–16). AKT has an important role in
apoptosis and cell production. Furthermore, PI3K can stimulate
cellular survival and proliferation, and is required for activation
of AKT (7,8). It is well known that the PI3K/Akt/mTOR
pathway enables the proliferation and survival of cancer driven by
growth factor receptors. The PI3K/Akt/mTOR pathway is another of
the most frequently dysregulated kinase cascades in human cancer
(9). Novel PI3K/AKT/mTOR
pathway-targeting drugs can overcome everolimus resistance in
pancreatic neuroendocrine tumors and cell lines (17). In pre-clinical and clinical studies,
both Ras/Raf/MEK/ERK and PI3K/Akt/mTOR cascade inhibitors have been
examined for possible uses (18–20).
Natural products and their derivatives can overpower MAPK signaling
and the PI3K pathways activated by oncogenic Ras (21).
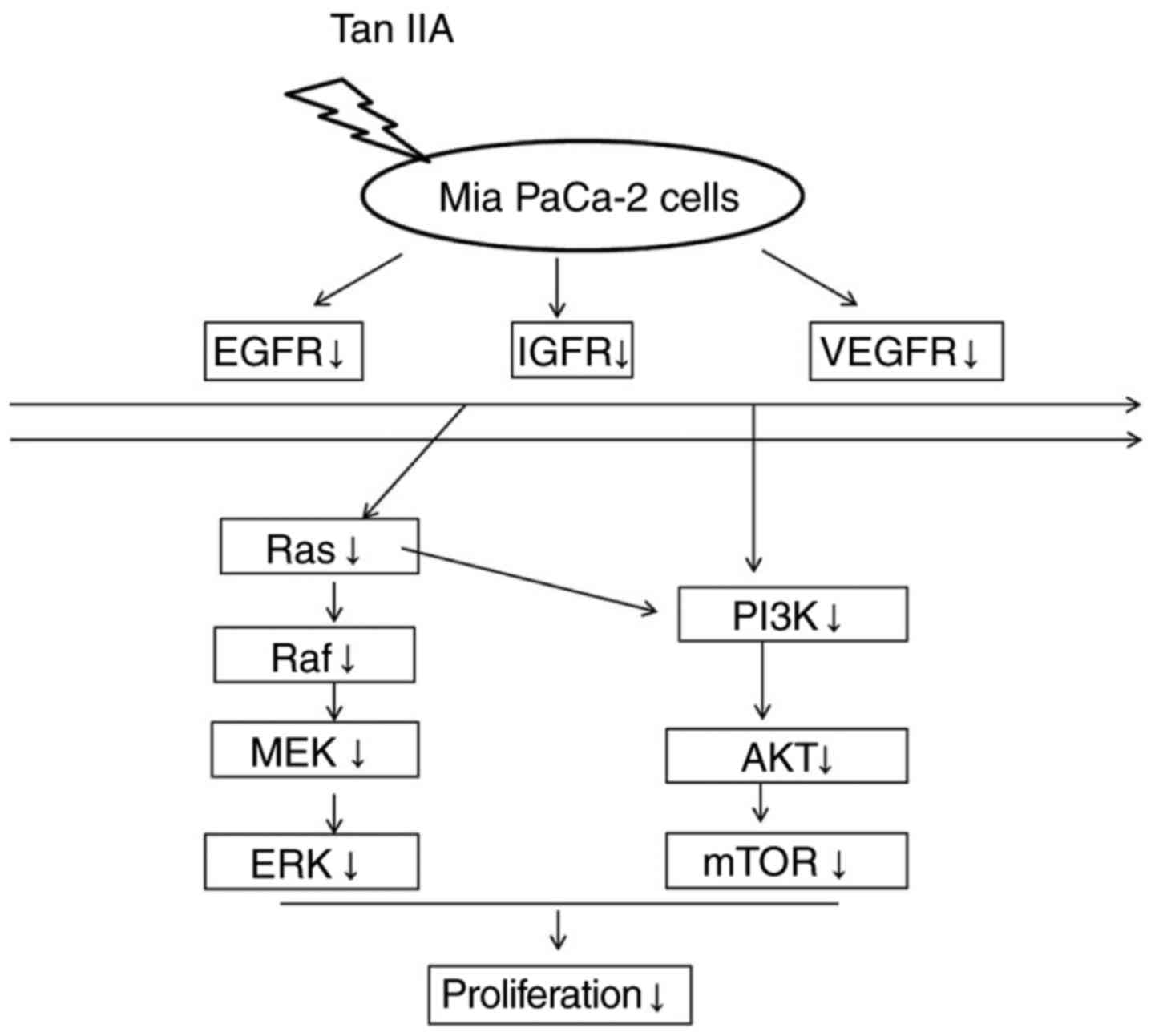
The present results demonstrated that MiaPaCa-2
cells treated with Tan-IIA exhibited decreased protein expression
levels of EGFR, IGF1R, VEGFR, Ras, Raf, MEK, ERK, PI3K, AKT and
mTOR. These findings indicated that one of the molecular mechanisms
through which Tan-IIA inhibits MiaPaCa-2 cells may be via
decreasing EGFR, IGFR and VEGFR expression, and inhibiting the
Ras/Raf/MEK/ERK and PI3K/Akt/mTOR pathway cascades. To the best of
our knowledge, the present study is the first to reveal that
Tan-IIA may inhibit MiaPaCa-2 pancreatic cancer cells through
decreasing the protein expression levels of EGFR, IGFR and VEGFR,
and inducing a dual blockade of Ras/Raf/MEK/ERK and PI3K/Akt/mTOR
pathways (Fig. 4). Although
advances have been made in systemic therapies, the use of adjuvant
treatment in pancreatic cancer remains uncertain; and the 5-year
survival rate for patients with early stage pancreatic cancer
remains <25% (22). Further
in vivo studies regarding the chemotherapeutic potential of
Tan-IIA in human pancreatic cancer are required.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from
Changhua Christian Hospital (grant no. 103-CCHICO-003).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Author contributions
CCS was involved in designing, performing the
experiments and manuscript submission.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The author declares that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2016. Cancer J Clinical. 66:7–30. 2016. View Article : Google Scholar
|
|
2
|
Che AJ, Zhang JY, Li CH, Chen XF, Hu ZD
and Chen XG: Separation and determination of active components in
Radix Salviae miltiorrhizae and its medicinal preparations by
nonaqueous capillary electrophoresis. J Sep Sci. 27:569–575. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhou L, Zuo Z and Chow MS: Danshen: An
overview of its chemistry, pharmacology, pharmacokinetics, and
clinical use. J Clin Pharmacol. 45:1345–1359. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Conroy T, Desseigne F, Ychou M, Bouché O,
Guimbaud R, Bécouarn Y, Adenis A, Raoul JL, Gourgou-Bourgade S, de
la Fouchardière C, et al: FOLFIRINOX versus gemcitabine for
metastatic pancreatic cancer. N Engl J Med. 364:1817–1825. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yuen JS and Macaulay VM: Targeting the
type 1 insulin-like growth factor receptor as a treatment for
cancer. Expert Opin Ther Targets. 12:589–603. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chitnis MM, Yuea JS, Protheroe AS, Pollak
M and Macaulay VM: The type 1 insulin-like growth factor receptor
pathway. Clin Cancer Res. 14:6364–6370. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Santarpia L, Lippman SM and El-Naggar AK:
Targeting the MAPK-RAS-RAF signaling pathway in cancer therapy.
Expert Opin Ther Targets. 16:103–119. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu P, Cheng H, Roberts TM and Zhao JJ:
Targeting the phosphoinositide 3-kinase pathway in cancer. Nat Rev
Drug Discov. 8:627–644. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
McCubrey JA, Steelman LS, Kempf CR,
Chappell WH, Abrams SL, Stivala F, Malaponte G, Nicoletti F, Libra
M, Basecke J, et al: Therapeutic resistance resulting from
mutations in Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR signaling pathways.
J Cell Physiol. 226:2762–2781. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang CY, Chiu TL, Kuo SJ, Chien SY, Chen
DR and Su CC: Tanshinone IIA inhibits the growth of pancreatic
cancer BxPC-3 cells by decreasing protein expression of TCTP, MCL-1
and Bcl-xL. Mol Med Rep. 7:1045–1049. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Su CC: Tanshinone IIA could inhibit
pancreatic cancer BxPC-3 cells through increasing PERK, ATF6,
caspase-12 and CHOP expression to induce apoptosis. J Biomedical
Sci Engineering. 8:149–159. 2015. View Article : Google Scholar
|
|
12
|
Fronza M, Murillo R, Ślusarczyk S, Adams
M, Hamburger M, Heinzmann B, Laufer S and Merfort I: In vitro
cytotoxic activity of abietane diterpenes from Peltodon
longipes as well as Salvia miltiorrhiza and Salvia
sahendica. Bioorg Med Chem. 19:4876–4881. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kranenburgm O, Gebbink MF and Voest EE:
Stimulation of angiogenesis by Ras proteins. Biochim Biophys Acta.
1654:23–37. 2004.PubMed/NCBI
|
|
14
|
Boonstra J, Rijken P, Humbel B, Cremers F,
Verkleij A and van Bergen en Henegouwen P: The epidermal growth
factor. Cell Biol Int. 19:413–430. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cary LA, Han DC and Guan JL:
Integrin-mediated signal transduction pathways. Histol Histopathol.
14:1001–1009. 1999.PubMed/NCBI
|
|
16
|
Stacey DW: Cyclin D1 serves as a cell
cycle regulatory switch in actively proliferating cells. Curr Opin
Cell Biol. 15:158–163. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Vandamme T, Beyens M, de Beeck KO, Dogan
F, van Koetsveld PM, Pauwels P, Mortier G, Vangestel C, de Herder
W, Van Camp G, et al: Long-term acquired everolimus resistance in
pancreatic neuroendocrine tumours can be overcome with novel
PI3K-AKT-mTOR inhibitors. Br J Cancer. 114:650–658. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
McCubrey JA, Steelman LS, Chappell WH,
Abrams SL, Franklin RA, Montalto G, Cervello M, Nicoletti F, Fagone
P, Malaponte G, et al: Mutations and deregulation of
Ras/Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR cascades which alter therapy
response. Oncotarget. 3:954–987. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sebolt-Leopold JS: Advances in the
development of cancer therapeutics directed against the
Ras-mitogen-activated protein kinase pathway. Clin Cancer Res.
14:3651–3656. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Haura EB, Ricart AD, Larson TG, Stella PJ,
Bazhenova L, Miller VA, Cohen RB, Eisenberg PD, Selaru P, Wilner
KD, et al: A phase II study of PD-0325901, an oral MEK inhibitor,
in previously treated patients with advanced non-small cell lung
cancer. Clin Cancer Res. 16:2450–2457. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Quah SY, Tan MS, Teh YH and Stanslas J:
Pharmacological modulation of oncogenic Ras by natural products and
their derivatives: Renewed hope in the discovery of novel anti-Ras
drugs. Pharmacol Ther. 162:35–57. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Goodman MD and Saif MW: Adjuvant therapy
for pancreatic cancer. JOP. 15:87–90. 2014.PubMed/NCBI
|