Introduction
Liver cancer is the fifth most common cancer and the
third leading cause of cancer-related mortality worldwide (1). Despite recent advances in disease
diagnosis and treatment, the long-term prognosis of liver cancer
patients remains poor. For advanced-stage liver cancer patients,
the overall 5-year survival rate is <5%. The main challenges for
liver cancer treatment include intrahepatic recurrence and
metastasis, which leads to the poor outcome for liver cancer
patients (2). The molecular
mechanisms underlying liver cancer recurrence and metastasis have
not been fully elucidated.
The important roles of microRNAs (miRNAs) in health
and diseases have been revealed in the past decade. miRNAs are
important non-coding RNAs that negatively regulate gene expression
at the post-transcriptional or translational levels. miRNAs
participate in the regulation of many cellular processes, including
cell proliferation, apoptosis, migration and invasion. Increasing
evidence suggests that a variety of miRNAs are aberrantly expressed
in human malignancies, including liver cancer (3,4). The
deregulated expression of miRNAs is critically involved in the
pathogenesis of cancer. miRNAs may function as tumor-suppressor
genes or oncogenes, contributing to the development and progression
of liver cancer (5–7). Therefore, further study of the
expression pattern and roles of miRNAs may provide novel diagnostic
and therapeutic targets for liver cancer.
miR-498 has been previously shown to be abnormally
expressed in several human cancers. For instance, miR-498 is
downregulated in non-small cell lung (8), ovarian (9), esophageal (10) and colorectal cancer (11). Low expression level of miR-498 is
associated with disease progression and poor clinical outcome in
cancer patients, indicating that miR-498 may act as a potential
tumor-suppressor gene. miR-498 has also been shown to inhibit
ovarian cancer cell growth by targeting human telomerase reverse
transcriptase (hTERT) (12) and
FOXO3 (13). However, it remains
unknown whether miR-498 is involved in liver cancer growth and
metastasis.
In the present study, we reported that miR-498
expression was low in human liver cancer and miR-498 overexpression
suppressed liver cancer cell growth in vitro and in
vivo. miR-498 overexpression induced cell cycle arrest and cell
apoptosis in liver cancer cells. Furthermore, miR-498 inhibited
liver cancer cell migration and invasion by reversing EMT.
Mechanistically, miR-498 targets zinc finger E-box binding homeobox
2 (ZEB2), which consequently inactivates the TGF-β/SMAD and
Wnt/β-catenin pathways. These findings provide a basis for the
better understanding of the roles of miRNAs in liver cancer and a
new target for the diagnosis and therapy of liver cancer.
Materials and methods
Microarray data analysis
Human microarray datasets GSE59856 (14) and GSE26323 (15) were obtained from the Gene Expression
Omnibus (GEO database, http://www.ncbi.nlm.nih.gov/geo/). The GSE59856
dataset contained the transcriptome data of 202 serum samples from
52 liver cancer patients and 150 healthy controls. The GSE26323
dataset contained 6 samples of paired primary liver cancer tissues
and lung metastatic tissues. The data were normalized by the Robust
Multichip Average (RMA) algorithm (16). The z-score of log2 format of
normalized data was used for further analysis.
Cell culture
Human normal liver cell line (HL-7702), human liver
cancer cell lines [HepG2 (hepatoblastoma) and HCC-LM3
(hepatocellular carcinoma)] and human 293T cells were purchased
from the Institutes for Biological Sciences at the Chinese Academy
of Sciences (Shanghai, China). The cells were cultured in
high-glucose Dulbecco's modified Eagle's medium (DMEM) supplemented
with 10% fetal bovine serum (FBS; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). All the cells were cultured in a humidified
incubator with 5% CO2 at 37°C.
Gene transfection
Cells were seeded in 6-well plates at a density of
2×105/well and cultured overnight. The overexpressing
plasmid and knockdown shRNA (Hanbio, Shanghai, China) were
transfected into the cells using LipoFiter transfection reagent
(Hanbio) in serum-free medium. Cells were added to complete medium
at 6 h after transfection and cultured for another 36 h.
Luciferase reporter assay
293T cells were co-transfected with miR-498 mimics
and the luciferase reporter vector containing wild-type (WT) or
mutant (MUT) 3′-UTR of ZEB2 as indicated. At 36 h after
transfection, the cells were lysed and the luciferase activity was
detected using the Dual-Luciferase Assay Kit (Promega Corporation,
Madison, WI, USA).
Cell cycle analysis
Cell cycle analysis was conducted with a cell cycle
detection kit (Fcmacs, Jiangsu, China). The transfected cells were
collected and fixed in 95% ethanol overnight. Afterwards, the cells
were stained with 50 µg/ml propidium iodide (PI) for 30 min in the
dark. The cell cycle distribution was analyzed on a flow cytometer
(BD FACSCalibur; BD Biosciences, San Jose, CA, USA) using CellQuest
software (BD Biosciences).
Cell apoptosis assay
Cell apoptosis was determined using the Annexin
V-Alexa Fluor 647/PI apoptosis detection kit (Fcmacs). The
transfected cells were digested with collagenase, collected, and
stained with Annexin V-Alexa Fluor 647 and PI for 15 min at room
temperature. The cell apoptosis rate was analyzed using flow
cytometry.
Cell counting and cell colony
formation assays
For the cell counting assay, the transfected cells
were seeded in 24-well plates (1×104/well) and were
counted for 6 days. For the cell colony formation assay, the
transfected cells were seeded in 6-well plates
(1×103/well) and cultured for 10 days at 37°C. The
medium was changed every 3 days. At the end of the experiments, the
cells were fixed with 4% paraformaldehyde and stained with 1%
crystal violet. The number of colonies was calculated under a Nikon
Eclipse Ti-S inverted microscope (Nikon Corp., Tokyo, Japan).
Transwell migration assay
The transfected cells were collected and seeded into
the upper Transwell chamber (8 µm) (Corning Inc., Corning, NY, USA)
at a density of 1×105 cells/well. The lower chamber was
filled with 500 µl culture medium supplemented with 10% FBS. The
cells were let to migrate for 12 h at 37°C. Then, the cells on the
upper surface of the membrane were removed with a cotton swab.
Then, the lower cells were fixed with formaldehyde and stained with
1% crystal violet for 30 min at room temperature. The number of
migrated cells was counted under a Nikon Eclipse Ti-S inverted
microscope (Nikon Corp.).
Cell invasion assay
The diluted basement Matrigel (BD Biosciences,
Franklin Lakes, NJ, USA) was added into each chamber and let to
polymerize at 37°C for 30 min. The transfected cells were seeded
into the upper chamber at a density of 2×105 cells/well.
The lower chamber was filled with 500 µl culture medium
supplemented with 10% FBS. The cells were allowed to invade to the
lower membrane for 24 h. Subsequently, the cells on the upper
surface of the membrane were removed with a cotton swab. The lower
cells were then fixed with formaldehyde and stained with 1% crystal
violet for 30 min at room temperature. The number of invaded cells
was counted under a Nikon Eclipse Ti-S inverted microscope (Nikon
Corp.).
Quantitative real-time PCR
Total RNA was extracted using TRIzol reagent (Thermo
Fisher Scientific, Inc.) and reverse transcribed into cDNA using
miScript reverse transcription kit (Bio-Rad Laboratories, Hercules,
CA, USA). The relative expression of target genes was detected on a
Bio-Rad CFX96 quantitative real-time PCR system with the SYBR-Green
method (Qiagen, Inc., Valencia, CA, USA). PCR was conducted at 95°C
for 15 min, followed by 40 cycles of 94°C for 15 sec, 55°C for 30
sec, and 63°C for 30 sec. U6 and β-actin served as internal
controls. The expression of miR-498 was analyzed by
2−ΔΔCq method. The sequences of the primers are listed
in Table I.
 | Table I.Primers used for qRT-PCR. |
Table I.
Primers used for qRT-PCR.
| Gene | Sequence
(5′-3′) | Product size
(bp) | Annealing
temperature (°C) |
|---|
| E-cadherin | F:
5′-CGCATTGCCACATACACTCT-3′ | 252 | 55 |
|
| R:
5′-TTGGCTGAGGATGGTGTAAG-3′ |
|
|
| N-cadherin | F:
5′-AGTCAACTGCAACCGTGTCT-3′ | 337 | 55 |
|
| R:
5′-AGCGTTCCTGTTCCACTCAT-3′ |
|
|
| Vimentin | F:
5′-GAGCTGCAGGAGCTGAATG-3′ | 344 | 55 |
|
| R: 5′-
AGGTCAAGACGTGCCAGAG-3′ |
|
|
| Slug | F:
5′-CCTGGTTGCTTCAAGGACAC-3′ | 395 | 55 |
|
| R:
5′-TCCATGCTCTTGCAGCTCTC-3′ |
|
|
| Twist | F:
5′-ACGAGCTGGACTCCAAGATG-3′ | 484 | 55 |
|
| R:
5′-GGCACGACCTCTTGAGAATG-3′ |
|
|
| Snail | F:
5′-GCGAGCTGCAGGACTCTAAT-3′ | 310 | 55 |
|
| R:
5′-GCCTCCAAGGAAGAGACTGA-3′ |
|
|
| Bcl-2 | F:
5′-GGATCCAGGATAACGGAGGC-3′ | 150 | 55 |
|
| R:
5′-CCAGATAGGCACCCAGGGT-3′ |
|
|
| Cyclin D1 | F:
5′-CCGAGAAGCTGTGCATCTAC-3′ | 221 | 55 |
|
| R:
5′-CTTCACATCTGTGGCACAGAG-3′ |
|
|
Western blot analysis
The cells were collected and lysed with RIPA buffer
containing 1% protease inhibitors. Equal amounts of proteins were
separated on 12% SDS-polyacrylamide gels and transferred onto
polyvinylidene fluoride (PVDF) membranes, followed by blocking with
5% non-fat milk for 1 h. The membranes were incubated with primary
antibodies overnight at 4°C. The following primary antibodies were
used: Anti-E-cadherin (cat. no. 4695S; Cell Signaling Technology,
Inc., Inc., Beverly, MA, USA), anti-N-cadherin (cat. no. 4370S;
Cell Signaling Technology, Inc.), anti-Bcl2 (cat. no. H-108; Santa
Cruz Biotechnology, Dallas, TX, USA), anti-cyclin D1 (cat. no.
H-108; Santa Cruz Biotechnology), anti-Slug (cat. no. 9585S; Cell
Signaling Technology, Inc., Inc.), anti-Snail (cat. no. 3879S; Cell
Signaling Technology, Inc.), anti-vimentin (cat. no. 5741S; Cell
Signaling Technology, Inc.), anti-β-catenin (cat. no. 8480S; Cell
Signaling Technology, Inc.), anti-c-Myc (cat. no. 5605S; Cell
Signaling Technology, Inc.), anti-TGF-β1 (3711S; Cell Signaling
Technology, Inc.), anti-SMAD2/3 (cat. no. 8685S; Cell Signaling
Technology, Inc.), anti-p-SMAD2 (cat. no. 3108S; Cell Signaling
Technology, Inc.), anti-p-SMAD3 (cat. no. 9520S; Cell Signaling
Technology, Inc.) and anti-GAPDH (cat. no. MB001; Bioworld
Technology, St. Louis Park, MN, USA). After incubation with the
secondary antibodies (anti-rabbit secondary antibody, cat. no.
31460; and anti-mouse secondary antibody, cat. no. 31430; both from
Bioworld Technology) at 37°C for 1 h, specific protein bands were
visualized using an ECL chemiluminescent detection system
(Millipore, Shanghai, China).
Clinical samples
The serum samples from liver cancer (n=20; 12 male
and 8 female patients; age range, 56–78 years old) and sex- and
age-matched healthy controls (n=20, 10 male and 10 female patients;
age range, 53–72 years old) were obtained from the Affiliated
People's Hospital of Jiangsu University between January 2018 and
April 2018. Written informed consent was obtained from all the
patients and this study was approved by the Institutional Ethics
Committee of Jiangsu University (ref. no. 2017003). The patients
included in the present study had not received any preoperative
therapy.
Animal study
Ten male BALB/c nude mice (weight, 21–25 g; aged 4–6
weeks) were purchased from the Model Animal Research Center of
Nanjing University (Nanjing, Jiangsu, China) and maintained in
specific pathogen-free (SPF) conditions in accordance with the
institutional policies. The mice received sterile rodent chow and
water ad libitum and were housed in sterile filter-top cages
with 12-h light/dark cycles. Control or miR-498-transfected HepG2
cells were collected in PBS and subcutaneously injected into the
mice (2×106 cells/mice, n=5). The mice were regularly
fed and the tumors were measured twice a week. The tumor volume was
calculated using the following formula: V (cm3) = 1/2 ×
length × width2. The protocol was approved by the
Laboratory Animal Management Committee of Jiangsu University.
Statistical analysis
All the results are expressed as the mean ± SD.
Differences between experimental groups were assessed by the
Student's t-test or one-way analysis of variance (ANOVA) with the
least significant difference (LSD) t-test using GraphPad Prism
version 5.0 software (GraphPad Software, La Jolla, CA, USA).
P<0.05 was considered as statistically significant.
Results
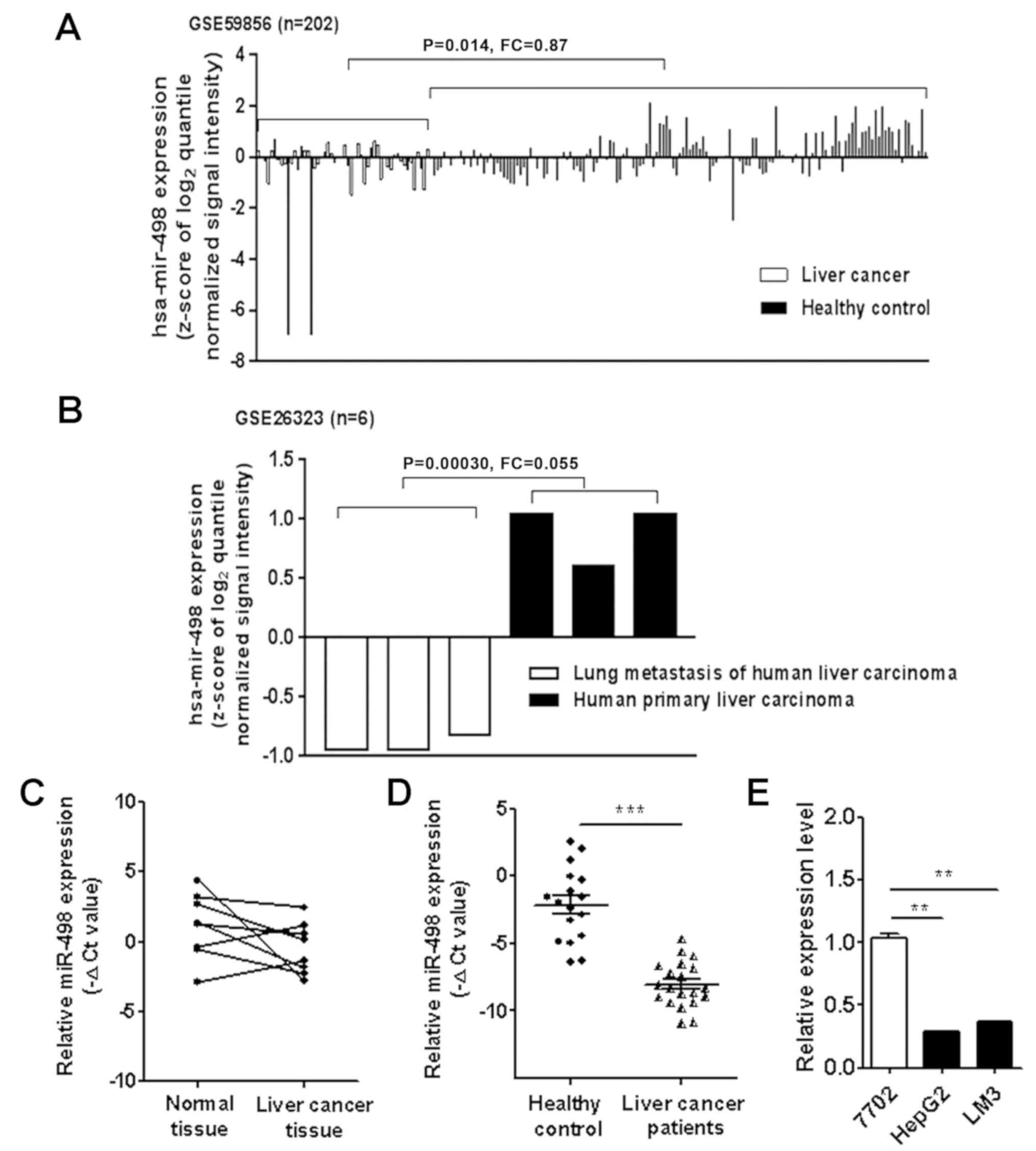
miR-498 is downregulated in human
liver cancer
We first analyzed the expression levels of miR-498
in liver cancer patients using the microarray data downloaded from
GEO (GSE59856 and GSE26323). The results showed that miR-498
expression level was downregulated in the serum of liver cancer
patients compared to that from healthy controls (Fig. 1A). miR-498 expression level was also
lower in the metastatic tumor tissues than that in the primary
tumor tissues (Fig. 1B). To
validate the findings of the GEO data analysis, we detected the
expression of miR-498 in 8 pairs of liver cancer tissues and
adjacent normal tissues using qRT-PCR. As shown in Fig. 1C, the expression of miR-498 was
downregulated in 6 liver cancer tissues compared to that noted in
the adjacent normal tissues. We further examined the expression of
miR-498 in serum samples from liver cancer patients and healthy
controls. The results showed that the expression levels of serum
miR-498 were significantly lower in liver cancer patients than that
in healthy controls (Fig. 1D).
Moreover, miR-498 expression levels were detected in the normal
liver cell line (HL-7702) and liver cancer cell lines [HepG2
(hepatoma) and HCC-LM3 (hepatocellular carcinoma)]. The expression
levels of miR-498 in HepG2 and HCC-LM3 cells were significantly
lower than that in the HL-7702 cells (Fig. 1E). Taken together, these findings
suggest that miR-498 is downregulated in liver cancer.
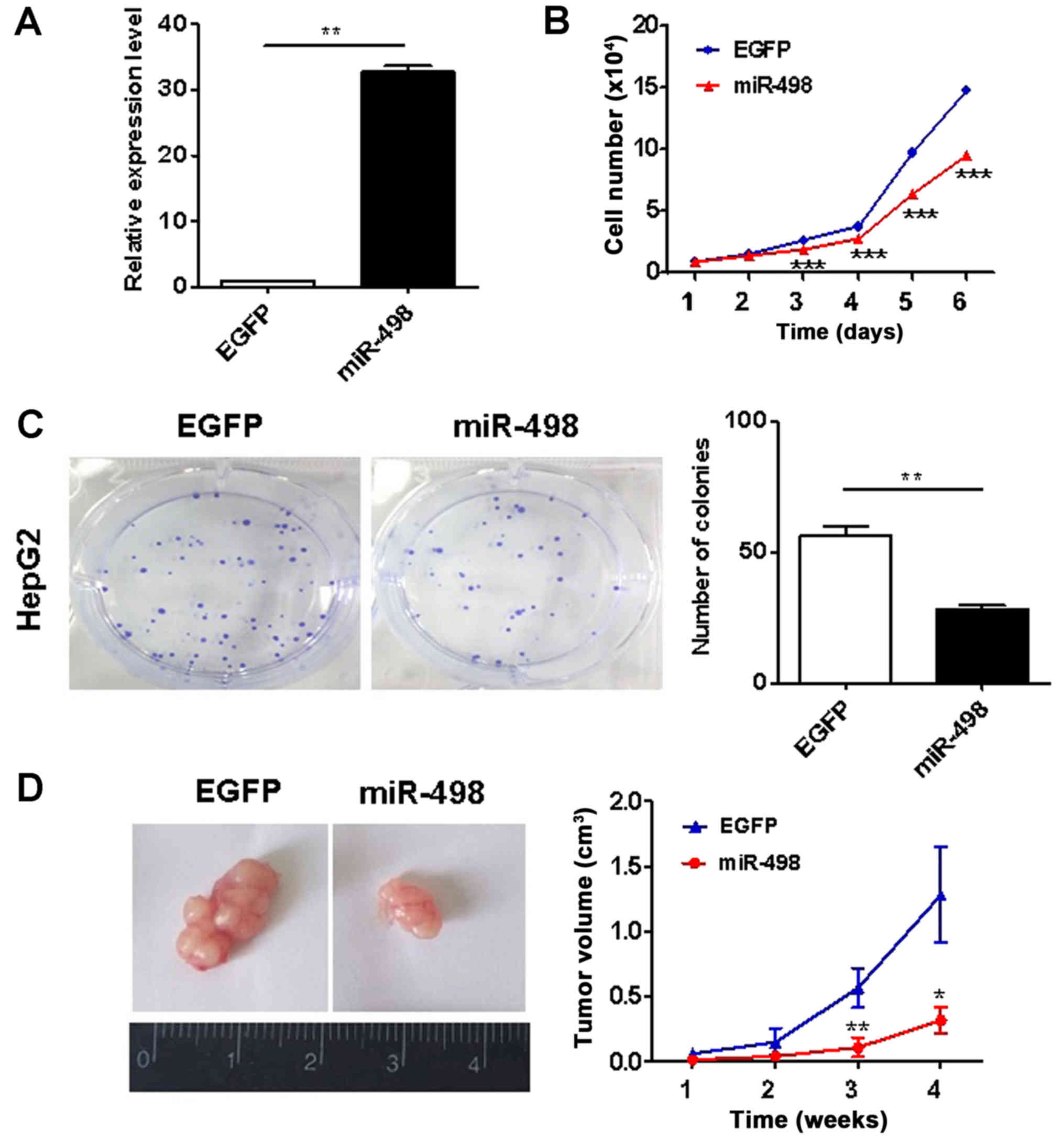
miR-498 overexpression inhibits the
growth of liver cancer cells
To investigate the roles of miR-498 in liver cancer,
we overexpressed miR-498 in HepG2 cells using gene transfection.
The efficacy of gene overexpression was validated (Fig. 2A). We then determined the
proliferation abilities of HepG2 cells using cell counting and
colony formation assays. The ectopic expression of miR-498
significantly inhibited the proliferation rate of HepG2 cells
(Fig. 2B). The results of colony
formation assay showed that HepG2 cells with miR-498 overexpression
formed significantly less colonies than the control cells
(P<0.01; Fig. 2C). Thus, these
findings indicate that miR-498 suppresses liver cancer cell
proliferation in vitro. We further determined the effects of
miR-498 on liver cancer growth in vivo. As shown in Fig. 2D, the growth rate of xenograft
tumors in the miR-498 overexpression group were significantly
slower than that of xenograft tumors in in the control group.
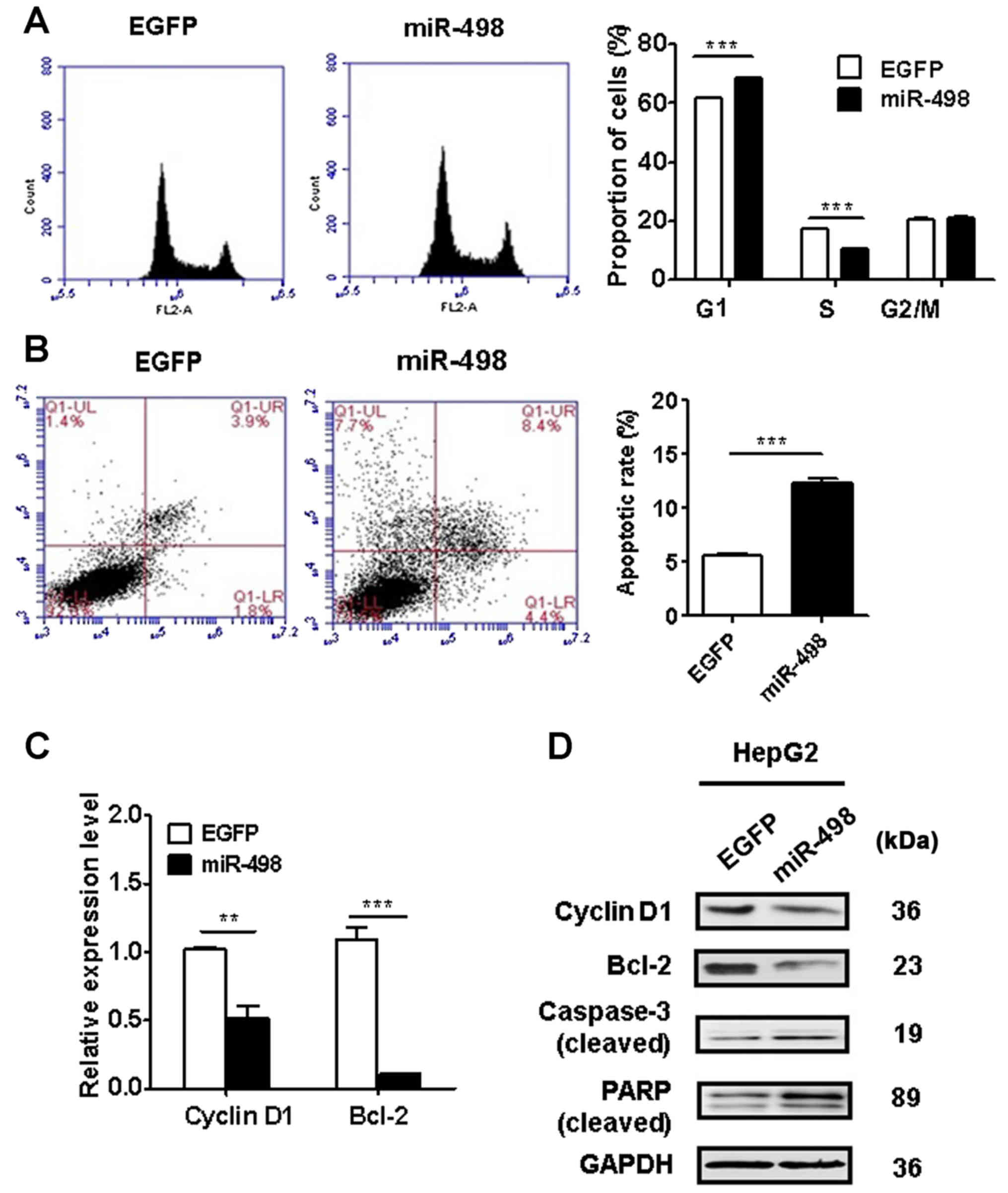
miR-498 overexpression induces cell
cycle arrest and apoptosis in liver cancer cells
We performed flow cytometric analyses to determine
the cell cycle distribution and apoptosis of HepG2 cells with or
without miR-498 overexpression. The results of cell cycle analysis
revealed that miR-498 overexpression in HepG2 cells significantly
increased the proportion of cells in the G1 phase (P<0.01) while
decreased that in the S phase (P<0.01) (Fig. 3A). We then evaluated the effect of
miR-498 overexpression on cell apoptosis. As shown in Fig. 3B, miR-498 overexpression in HepG2
cells significantly increased the percentage of apoptotic cells. We
determined the expression of cell cycle and apoptosis-related genes
and proteins using qRT-PCR and western blot analysis. As shown in
Fig. 3C and D, miR-498
overexpression led to a decrease in the expression of cyclin D1 and
Bcl-2 in HepG2 cells. Moreover, western blot results showed that
miR-498 overexpression promoted the cleavage of PARP and caspase-3
in HepG2 cells (Fig. 3D). Taken
together, these results indicate that miR-498 overexpression
inhibited liver cancer cell proliferation by inducing cell cycle
arrest and apoptosis.
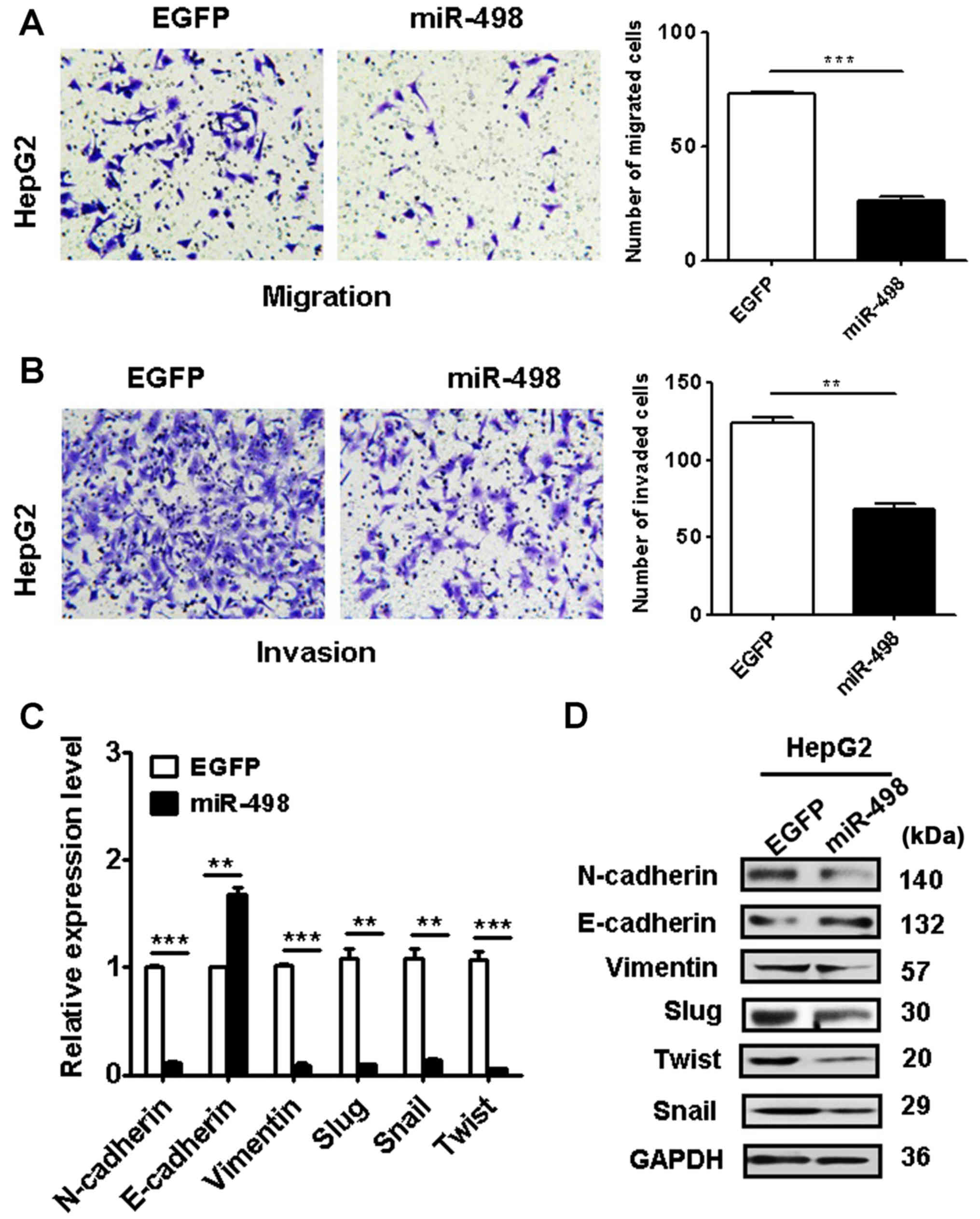
miR-498 overexpression suppresses the
metastatic abilities of liver cancer cells
We next determined the effects of miR-498
overexpression on the metastatic potential of liver cancer cells.
Our Transwell migration assay results showed that the miR-498
overexpression group had less migrated cells than that of the
control group (Fig. 4A). In
addition, the results of Matrigel invasion assays showed that
miR-498 overexpression also suppressed the invasion abilities of
HepG2 cells (Fig. 4B). Together,
these data suggest that miR-498 has an inhibitory role in liver
cancer cell migration and invasion.
To further understand the mechanisms by which
miR-498 overexpression inhibits liver cancer cell migration and
invasion, we determined the expression of EMT-related factors in
HepG2 cells with or without miR-498 overexpression. We found that
miR-498 overexpression upregulated the expression of E-cadherin
gene while downregulated that of N-cadherin, vimentin, Slug, Snail
and Twist genes in HepG2 cells (Fig.
4C). Western blot results also confirmed the same changes
(Fig. 4D). These data indicate that
miR-498 overexpression may inhibit EMT in liver cancer cells.
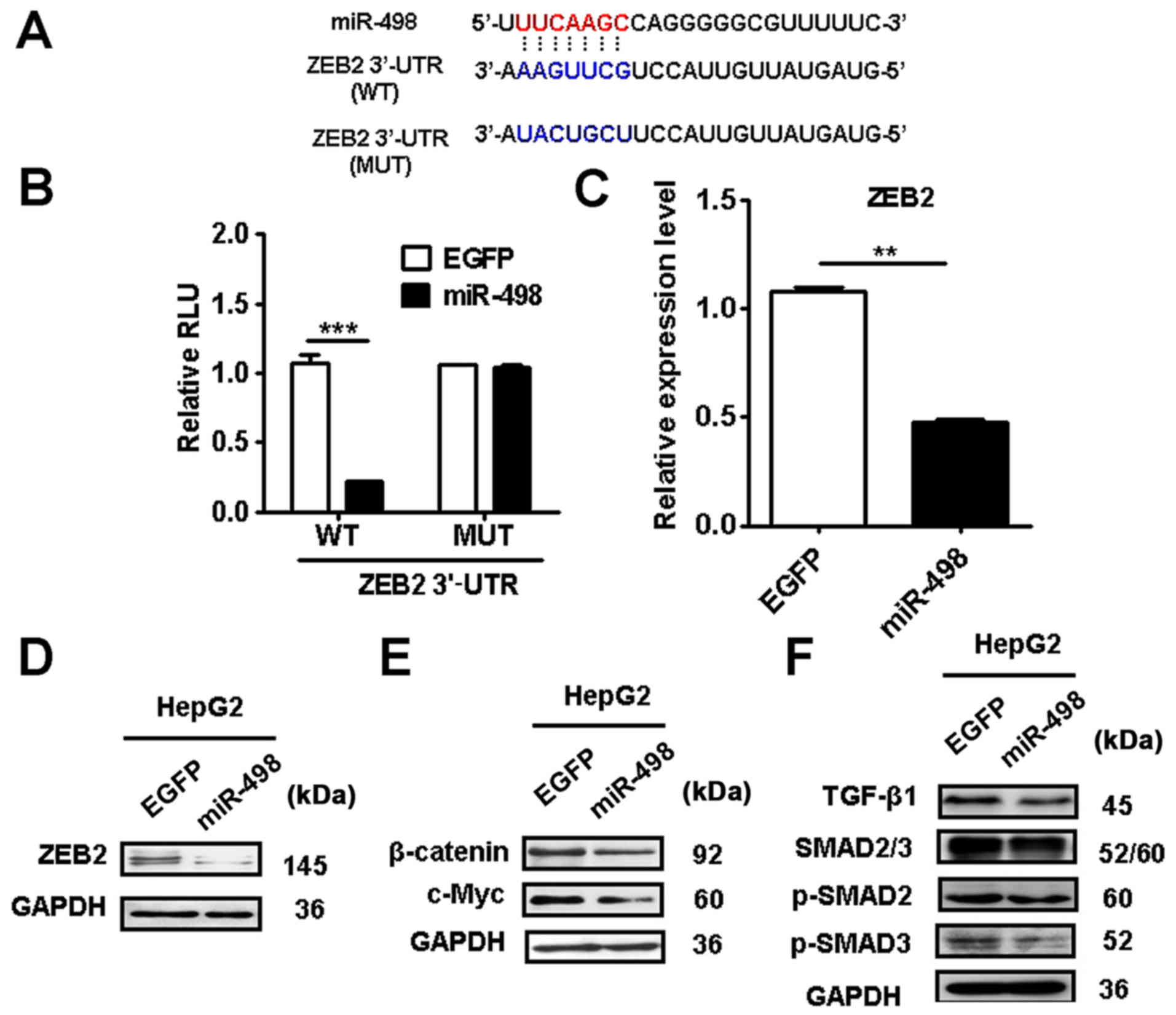
miR-498 targets ZEB2 in liver cancer
cells
We searched for potential target genes of miR-498
using the bioinformatic algorithm TargetScan (http://www.targetscan.org) and miRDB (http://www.miRDB.org) and identified ZEB2 as a
putative target of miR-498 (Fig.
5A). We further validated the prediction using luciferase
reporter assay. The 3′-UTR of ZEB2 mRNA containing the wild-type
(WT) or mutant (MT) miR-498 potential binding site were cloned and
inserted into the luciferase reporter vector. The luciferase
reporter assay results showed that miR-498 overexpression
significantly decreased the activity of luciferase reporter gene in
wild-type but not the mutant group (Fig. 5B). In addition, miR-498
overexpression significantly reduced the expression of ZEB2 gene
and protein in HepG2 cells (Fig. 5C and
D). Moreover, we found that miR-498 overexpression inhibited
the expression of β-catenin and its downstream target c-Myc in
HepG2 cells (Fig. 5E). The
expression levels of TGF-β1, p-SMAD2 and p-SMAD3 were also
suppressed by miR-498 overexpression (Fig. 5F). These results indicate that
miR-498 targets ZEB2 to inactivate the Wnt/β-catenin and TGF-β/SMAD
pathways in liver cancer cells.
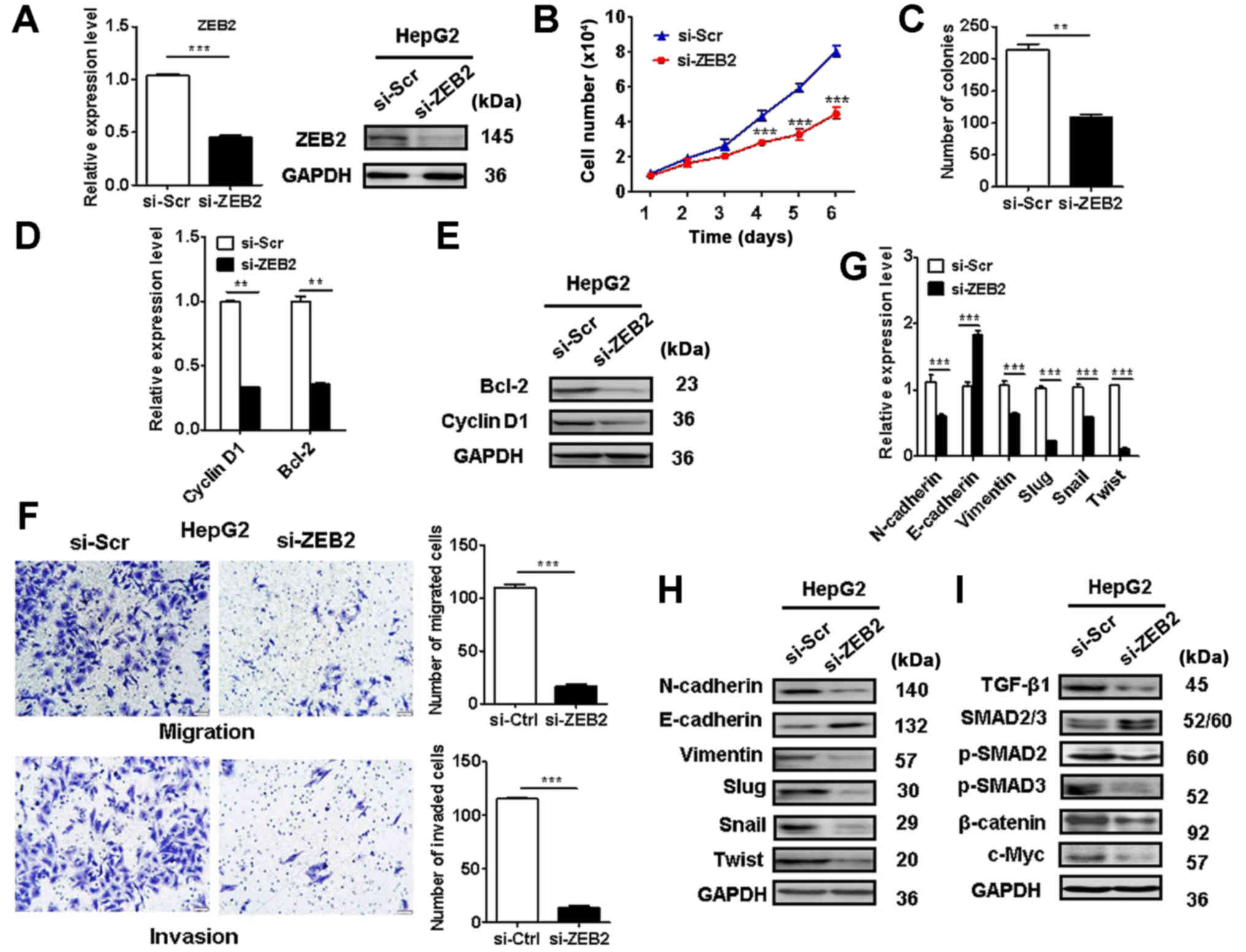
ZEB2 knockdown re-capitulates the
inhibitory roles of miR-498 in liver cancer cell proliferation,
migration and invasion
We further determined the role of ZEB2 in liver
cancer progression. The efficacy of ZEB2 knockdown in liver cancer
cells was validated (Fig. 6A). Cell
counting and colony formation assay results showed that ZEB2
knockdown suppressed HepG2 cell proliferation (Fig. 6B and C). In addition, ZEB2 knockdown
in HepG2 cells decreased the expression of cyclin D1 and Bcl-2
(Fig. 6D and E). ZEB2 knockdown
also suppressed the migration and invasion of HepG2 cells (Fig. 6F). ZEB2 knockdown in liver cancer
cells increased the expression of E-cadherin while decreased that
of N-cadherin, vimentin, Slug, Snail, and Twist (Fig. 6G and H). Moreover, ZEB2 knockdown
inactivated Wnt/β-catenin and TGF-β/SMAD pathways in HepG2 cells
(Fig. 6I). These findings suggest
that ZEB2 knockdown recapitulates the inhibitory roles of miR-498
in liver cancer cells.
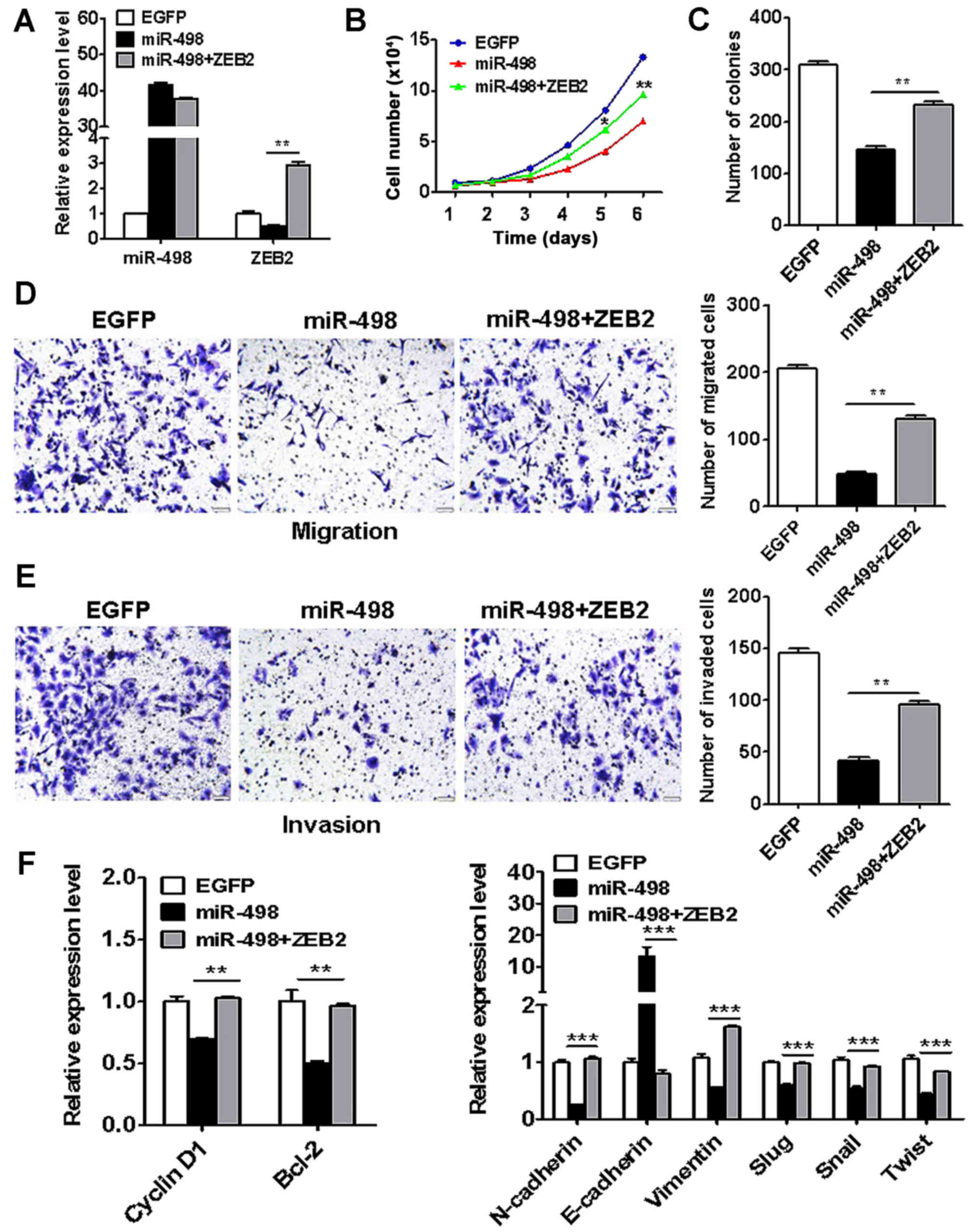
ZEB2 overexpression reverses the
suppressive effects of miR-498 on liver cancer cells
To further demonstrate that ZEB2 is a key target of
miR-498 in liver cancer, we co-transfected ZEB2 and miR-498 into
HepG2 cells (Fig. 7A). The results
of cell counting and colony formation assays showed that ZEB2
overexpresvicesion reversed the inhibition of proliferation by
miR-498 in liver cancer cells (Fig. 7B
and C). In addition, ZEB2 overexpression restored the migratory
and invasive abilities of HepG2 cells that were inhibited by
miR-498 (Fig. 7D and E). Moreover,
ZEB2 overexpression reversed the cyclin D1, Bcl-2, N-cadherin,
vimentin, Slug, Snail and Twist expression in the
miR-498-transfected HepG2 cells (Fig.
7F). These results suggest that ZEB2 is key target of miR-498
in liver cancer.
Discussion
In the past decade, various miRNAs have been
identified to play critical roles in the pathogenesis of liver
cancer. For example, miR-146a suppresses liver cancer metastasis by
inhibiting the nuclear accumulation of β-catenin and downregulating
NF-κB p65 (17). Wu et al
demonstrated that miR-655-3p targets ADAM10 to inhibit liver cancer
growth (18). Yu et al found
that miR-195 targets YAP to inhibit EMT in liver cancer cells
(19). In the present study, we
showed that miR-498 was frequently downregulated in liver cancer
tissues and serum samples. Moreover, we found that decreased
miR-498 expression was associated with liver cancer progression. We
further investigated the function of miR-498 in liver cancer and
found that miR-498 overexpression significantly inhibited the
growth and metastasis of liver cancer cells in vitro and
in vivo, indicating that miR-498 acts as a tumor suppressor
in liver cancer. Although our preliminary results is in support of
this notion, the detection of miR-498 expression in a large cohort
of liver cancer tissue and serum samples need to be performed in
future studies.
Our data suggest that the downregulation of miR-498
may contribute to the advanced progression of human liver cancer.
Similar findings have been reported in other types of human
cancers. For instance, Wang et al found that miR-498
inhibited the proliferation of non-small cell lung cancer (NSCLC)
cells (8). Gopalan et al
found that miR-498 suppressed cell proliferation in colorectal
cancer (11). However, miR-498 is
found to act as an oncogenic miRNA that targets cell cycle
progression 1 (CCPG1) in retinoblastoma (20). miR-498 was found to be consistently
upregulated in triple-negative breast cancer (TNBC) cells and
miR-498 knockdown led to reduced proliferation in TNBC cells
(21,22), suggesting that the functional roles
of miR-498 is tumor type-specific and cell context-dependent.
Recurrence or metastasis is the major cause of the
poor prognosis of liver cancer patients. miRNAs have been
previously suggested to play important roles in cancer metastasis.
We found that miR-498 overexpression decreased the metastatic
ability of liver cancer cells. EMT is a key mechanism for cancer
metastasis. During the process of EMT, the cancer cells lose the
characteristics of epithelial cells and acquire the properties of
mesenchymal cells. The previous studies demonstrated that miRNAs
can regulate the occurrence of EMT in liver cancer. We found that
miR-498 overexpression increased E-cadherin expression and
decreased N-cadherin and vimentin expression. Furthermore, we
confirmed that liver cancer cells with miR-498 overexpression had
significantly lower level of several important EMT-related
transcription factors. These findings suggest that miR-498 could
suppress liver cancer metastasis by inhibiting EMT.
ZEB2 is highly expressed in human cancers and plays
oncogenic roles (23,24). Several other studies have shown that
ZEB2 is regulated by miRNAs. miRNA-30a-5p targets ZEB2 to promote
the aggressiveness of clear cell renal cell carcinoma (25). In papillary thyroid cancer, miR-335
expression is decreased and its re-expression suppresses cancer
cell growth, migration and invasion by directly targeting ZEB2
(26). Pang et al
demonstrated that miR-154 targets ZEB2 to inhibit liver cancer cell
proliferation, migration and invasion (27). Hu et al demonstrated that
miR-204 targets ZEB2 to suppress liver cancer cell growth and
metastasis (28). Our results
showed that miR-498 overexpression markedly decreased the
expression of ZEB2 in liver cancer cells. We identified ZEB2 as a
novel target gene of miR-498. We further showed that ZEB2 knockdown
could re-capitulate the suppressive roles of miR-498 in the
proliferation, migration, and invasion of liver cancer cells. In
glioma cells, ZEB2 knockdown was found to decrease β-catenin
expression (29). Furthermore, Zhou
et al reported that miRNA-145 targets ZEB2 to inactivate the
Wnt/β-catenin pathway, inhibiting hepatic stellate cell activation
and proliferation (30). In the
present study, we found that miR-498 overexpression inactivated
Wnt/β-catenin and TGF-β/SMAD pathways in liver cancer cells. miRNAs
have been suggested to regulate multiple molecular pathways that
are involved in the development and progression of tumorigenesis
(31). Whether miR-498 could
regulate other targets and pathways in liver cancer warrants
further investigation in future studies.
In conclusion, we demonstrated in the present study
that miR-498 acts as a novel tumor suppressor in liver cancer.
miR-498 inhibits the proliferation, migration, and invasion of
liver cancer cells, partly at least, via targeting ZEB2. The
present study not only provides novel insights into the mechanism
underlying the development and progression of liver cancer, but
also suggests a promising diagnostic and therapeutic target for
liver cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (nos. 81672416 and 81572075),
the Key Research and Development Project of Zhenjiang (no.
SH2015034), the Major Research and Development Project of Jiangsu
Province (no. BE2015667), the Project of Jiangsu Provincial
Commission of Health and Family Planning (no. LGY2017024), the
Starting Foundation for Senior Talents of Jiangsu University (no.
13JDG086), the Qing Lan Project of Jiangsu Province, the ‘333’
project of Jiangsu Province and the Foundation for Young Academic
Leader of Jiangsu University.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
XZh conceived the idea of this study; XZh, XX, GG,
XZa, MS, SZ, YZ, ZM, JZ and FM performed the experiments and
acquired the data; XZh, HQ and WX analyzed and interpreted the
data; XZh, XX, HQ and WX drafted and revised the manuscript. All
authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Written informed consent was obtained from all the
patients and this study was approved by the Institutional Ethics
Committee of Jiangsu University (ref no. 2017003). The protocol for
the animal study was approved by the Laboratory Animal Management
Committee of Jiangsu University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Singal AG and El-Serag HB: Hepatocellular
carcinoma from epidemiology to prevention: Translating knowledge
into practice. Clin Gastroenterol Hepatol. 13:2140–2151. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mizuguchi Y, Takizawa T, Yoshida H and
Uchida E: Dysregulated miRNA in progression of hepatocellular
carcinoma: A systematic review. Hepatol Res. 46:391–406. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Morishita A and Masaki T: miRNA in
hepatocellular carcinoma. Hepatol Res. 45:128–141. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chang RM, Xiao S, Lei X, Yang H, Fang F
and Yang LY: miRNA-487a promotes proliferation and metastasis in
hepatocellular carcinoma. Clin Cancer Res. 23:2593–2604. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dietrich P, Koch A, Fritz V, Hartmann A,
Bosserhoff AK and Hellerbrand C: Wild type Kirsten rat sarcoma is a
novel microRNA-622-regulated therapeutic target for hepatocellular
carcinoma and contributes to sorafenib resistance. Gut.
67:1328–1341. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kabir TD, Ganda C, Brown RM, Beveridge DJ,
Richardson KL, Chaturvedi V, Candy P, Epis M, Wintle L, Kalinowski
F, et al: A microRNA-7/growth arrest specific 6/TYRO3 axis
regulates the growth and invasiveness of sorafenib-resistant cells
in human hepatocellular carcinoma. Hepatology. 67:216–231. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang M, Zhang Q, Wang J and Zhai Y:
MicroRNA-498 is downregulated in non-small cell lung cancer and
correlates with tumor progression. J Cancer Res Ther. 11 Suppl
1:C107–C111. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cong J, Liu R, Wang X, Wang J, Wang H and
Hou J: Low miR-498 expression levels are associated with poor
prognosis in ovarian cancer. Eur Rev Med Pharmacol Sci.
19:4762–4765. 2015.PubMed/NCBI
|
|
10
|
Islam F, Gopalan V, Law S, Tang JC, Chan
KW and Lam AK: MiR-498 in esophageal squamous cell carcinoma:
Clinicopathological impacts and functional interactions. Hum
Pathol. 62:141–151. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gopalan V, Smith RA and Lam AK:
Downregulation of microRNA-498 in colorectal cancers and its
cellular effects. Exp Cell Res. 330:423–428. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kasiappan R, Shen Z, Tse AK, Jinwal U,
Tang J, Lungchukiet P, Sun Y, Kruk P, Nicosia SV, Zhang X, et al:
1,25-Dihydroxyvitamin D3 suppresses telomerase expression and human
cancer growth through microRNA-498. J Biol Chem. 287:41297–41309.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu R, Liu F, Li L, Sun M and Chen K:
MiR-498 regulated FOXO3 expression and inhibited the proliferation
of human ovarian cancer cells. Biomed Pharmacother. 72:52–57. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kojima M, Sudo H, Kawauchi J, Takizawa S,
Kondou S, Nobumasa H and Ochiai A: MicroRNA markers for the
diagnosis of pancreatic and biliary-tract cancers. PLoS One.
10:e01182202015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tao ZH, Wan JL, Zeng LY, Xie L, Sun HC,
Qin LX, Wang L, Zhou J, Ren ZG, Li YX, et al: miR-612 suppresses
the invasive-metastatic cascade in hepatocellular carcinoma. J Exp
Med. 210:789–803. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fu M, Huang Z, Zang X, Pan L, Liang W,
Chen J, Qian H, Xu W, Jiang P and Zhang X: Long noncoding RNA
LINC00978 promotes cancer growth and acts as a diagnostic biomarker
in gastric cancer. Cell Prolif. 5:2017.doi: 10.1111/cpr.12425.
|
|
17
|
Zhang Z, Zhang Y, Sun XX, Ma X and Chen
ZN: microRNA-146a inhibits cancer metastasis by downregulating VEGF
through dual pathways in hepatocellular carcinoma. Mol Cancer.
14:52015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu G, Zheng K, Xia S, Wang Y, Meng X, Qin
X and Cheng Y: MicroRNA-655-3p functions as a tumor suppressor by
regulating ADAM10 and β-catenin pathway in hepatocellular
carcinoma. J Exp Clin Cancer Res. 35:892016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yu S, Jing L, Yin XR, Wang MC, Chen YM,
Guo Y, Nan KJ and Han LL: MiR-195 suppresses the metastasis and
epithelial-mesenchymal transition of hepatocellular carcinoma by
inhibiting YAP. Oncotarget. 8:99757–99771. 2017.PubMed/NCBI
|
|
20
|
Yang L, Wei N, Wang L, Wang X and Liu QH:
miR-498 promotes cell proliferation and inhibits cell apoptosis in
retinoblastoma by directly targeting CCPG1. Childs Nerv Syst.
34:417–422. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Matamala N, Vargas MT, González-Cámpora R,
Arias JI, Menéndez P, Andrés-León E, Yanowsky K, Llaneza-Folgueras
A, Miñambres R, Martínez-Delgado B, et al: MicroRNA deregulation in
triple negative breast cancer reveals a role of miR-498 in
regulating BRCA1 expression. Oncotarget. 7:20068–20079. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chai C, Wu H, Wang B, Eisenstat DD and
Leng RP: MicroRNA- 498 promotes proliferation and migration by
targeting the tumor suppressor PTEN in breast cancer cells.
Carcinogenesis. 39:1185–1196. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dai YH, Tang YP, Zhu HY, Lv L, Chu Y, Zhou
YQ and Huo JR: ZEB2 promotes the metastasis of gastric cancer and
modulates epithelial mesenchymal transition of gastric cancer
cells. Dig Dis Sci. 57:1253–1260. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li MZ, Wang JJ, Yang SB, Li WF, Xiao LB,
He YL and Song XM: ZEB2 promotes tumor metastasis and correlates
with poor prognosis of human colorectal cancer. Am J Transl Res.
9:2838–2851. 2017.PubMed/NCBI
|
|
25
|
Chen Z, Zhang J, Zhang Z, Feng Z, Wei J,
Lu J, Fang Y, Liang Y, Cen J, Pan Y, et al: The putative tumor
suppressor microRNA-30a-5p modulates clear cell renal cell
carcinoma aggressiveness through repression of ZEB2. Cell Death
Dis. 8:e28592017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kan Q, Su Y and Yang H: MicroRNA-335 is
downregulated in papillary thyroid cancer and suppresses cancer
cell growth, migration and invasion by directly targeting ZEB2.
Oncol Lett. 14:7622–7628. 2017.PubMed/NCBI
|
|
27
|
Pang X, Huang K, Zhang Q, Zhang Y and Niu
J: miR-154 targeting ZEB2 in hepatocellular carcinoma functions as
a potential tumor suppressor. Oncol Rep. 34:3272–3279. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hu B, Sun M, Liu J, Hong G and Lin Q:
MicroRNA-204 suppressed proliferation and motility capacity of
human hepatocellular carcinoma via directly targeting zinc finger
E-box binding homeobox 2. Oncol Lett. 13:3823–3830. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Qi S, Song Y, Peng Y, Wang H, Long H, Yu
X, Li Z, Fang L, Wu A, Luo W, et al: ZEB2 mediates multiple
pathways regulating cell proliferation, migration, invasion, and
apoptosis in glioma. PLoS One. 7:e388422012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou DD, Wang X, Wang Y, Xiang XJ, Liang
ZC, Zhou Y, Xu A, Bi CH and Zhang L: MicroRNA-145 inhibits hepatic
stellate cell activation and proliferation by targeting ZEB2
through Wnt/β-catenin pathway. Mol Immunol. 75:151–160. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vasuri F, Visani M, Acquaviva G, Brand T,
Fiorentino M, Pession A, Tallini G, D'Errico A and de Biase D: Role
of microRNAs in the main molecular pathways of hepatocellular
carcinoma. World J Gastroenterol. 24:2647–2660. 2018. View Article : Google Scholar : PubMed/NCBI
|