Introduction
Glioma is one of the most common primary malignant
tumors in the central nervous system (CNS), which accounted for
~40% of all intracranial tumors (1). According to the world health
organization (WHO), gliomas can be divided into four grades (I, II,
III and IV) (2). Glioblastoma
(GBM), the highest grade (grade IV) of glioma tumor, is the most
malignant form of astrocytoma. Although there have been great
advances in treatment options, including surgery, radiotherapy and
chemotherapy, in recent decades, the prognosis of GBM remains very
poor. At present, the 5-year survival rate of patients with GBM is
only 9.8% (3–5). In order to improve the prognosis of
patients with GBM, it is urgent to identify novel therapeutic
targets.
Cell adhesion molecules (CADM) consists of a protein
family, which has been reported to maintain cell polarity. The
majority of the CADMs belong to the immunoglobulin superfamily, and
recent research has suggested that CADM also acts as a tumor
suppressor (6–9). CADM2 is a member of the CADM family,
and previous studies have demonstrated that CADM2 inhibits human
renal clear cell carcinoma by promoting DNA promoter methylation
and/or loss of heterozygosity. Loss of CADM2 causes higher tumor
pathological stages (6). In
addition, CADM2 has been observed to be downregulated in prostate
(10) and ovarian cancer (11), and lymphoma and melanoma (12,13).
However, to the best of our knowledge, the present study was the
first to have reported the role of CADM2 in human glioma. In the
present study, the expression level of CADM2 in human glioma was
analyzed by bioinformatics (Oncomine database) and tissue
microarrays. The results demonstrated that CADM2 was markedly
downregulated in human glioma tissues. The expression level of
CADM2 was decreased, while glioma pathological grades were
increased. CADM2 inhibited GBM cell proliferation, migration and
invasion in vitro and in vivo. Furthermore, CADM2
regulated the glioma cell G1/S transition and affected glioma cell
epithelial-mesenchymal transition (EMT). Taken together, the
results of the present study implicated CADM2 as a novel tumor
suppressor candidate in human glioma, and provided a novel
molecular target for the treatment of human glioma.
Materials and methods
Tissues
Human glioma tissues were obtained from the surgical
branch of Tangdu Hospital, Fourth Military Medical University,
China. Normal brain tissues, which were preserved in liquid
nitrogen, were obtained from the patients who had undergone partial
brain resection. The tissue samples were obtained with the written
informed consent of the patients, and the process was approved by
the Ethics Committee of Tangdu Hospital Institutional Review
Board.
Cell lines and lentiviral
transfection
The established U251 and U87MG cell lines used in
the present study were purchased from the Cell Bank of Chinese
Academy of Sciences (14). The
authenticity of the cancer cell lines was tested by short tandem
repeat (STR) profiling. The STR result demonstrated that the U87
cell line used in the present study matched the U87MG cell from
ATCC. However, the U87MG cell line from ATCC may not be the
original GBM cell line from the University of Uppsala established
in 1968. It has been reported that the U87MG cell line from ATCC is
of CNS origin and is likely to be derived from another patient with
glioma, although its source is unknown (15). Therefore, the aforementioned
misidentifications of U87MG-ATCC did not affect the outcomes of the
present study. All the cell lines were cultured in 10% fetal bovine
serum (FBS) Dulbecco's modified Eagle's medium (DMEM; both Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA). The CADM2 coding
sequence (CDS) was sub-cloned into a pCDH1-overexpression vector
using the restriction sites of Bamh1 and Xho1. U251
and U87 cells were transfected with CADM2-overexpression
lentivirus, and stable CADM2-overexpression cell lines were
established by puromycin screening at 2 µg/ml.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol reagent (Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
Reverse transcription was performed using a Takara reverse
transcription kit (Takara Bio, Inc., Otsu, Japan). The sequences of
the primers used were as follows: CADM2 forward,
5′-CCTCAATGCCACCCCTCAG-3′ and reverse, 5′-TTCTCCGCCATCCTTTGTCC-3′;
and β-actin forward, 5′-TCCCTGGAGAAGAGCTACG-3′ and reverse,
5′-GTAGTTTCGTGGATGCCACA-3′. The reverse transcription process was
as follows: 37°C for 1 h and 85°C for 15 sec. PCR conditions were
95°C for 5 min, followed by 40 cycles at 95°C for 20 sec, 60°C for
30 sec, 72°C for 60 sec. Gene expression in each sample was
normalized to relative to that of β-actin mRNA. qPCR was performed
using SYBRGreen (Takara Bio, Inc.) with the ABI PRISM 7700 Sequence
Detection system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The expression was normalized to human β-Actin expression
using the 2−ΔΔCq method (16).
Western blot assay
Radioimmunoprecipitation assay (RIPA) lysis buffer
was used to lyse glioma tissues and different glioma cell lines,
and complete protease inhibitor cocktail (Beyotime Institute of
Biotechnology, Haimen, China) was supplemented with RIPA lysis
buffer prior to lysis. Bicinchoninic acid protein assay kit
(Beyotime Institute of Biotechnology) was used to detect protein
concentration. A total of 30 µg protein samples were loaded onto
the gel and separated by 12% SDS-PAGE gel. Protein was subsequently
transferred to polyvinylidene difluoride membranes. Following
blocking with 5% skimmed milk for 1 h at room temperature, the
membranes were incubated with the following primary antibodies at
4°C overnight: CADM2 (dilution, 1:4,000; cat. no. ab64873; Abcam,
Cambridge, UK), β-actin (dilution, 1:4,000; cat. no. M20011; Abmart
Co., Ltd., Shanghai, China), CDK2 [dilution, 1:2,000; cat. no.
2546; Cell Signaling Technology (CST)], CDK4 (dilution, 1:2,000;
cat. no. 12790; CST), E-cadherin (dilution, 1:2,000; cat. no.
14472; CST), β-catenin (dilution, 1:2,000; cat. no. 8480; CST),
cyclin D1 (dilution, 1:1,000; cat. no. sc-70899; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) and cyclin E (dilution,
1:1,000; cat. no. sc-377100; Santa Cruz Biotechnology, Inc.). On
the second day, the membranes were incubated with horseradish
peroxidase (HRP)-conjugated secondary antibodies (goat anti-mouse
IgG, cat. no. ab97023 or goat anti-rabbit IgG, cat. no. ab6721;
dilution, 1:1,000) at room temperature for 2 h. Next, the membranes
were exposed using a Millipore enhanced chemiluminescence kit.
MTT assay
The proliferation rates of different U87 and U251
cells (Blank, vector and CADM2 groups) were detected using an MTT
assay. A total of 1×104 cells/well were suspended in 200
µl culture DMEM supplemented with 10% FBS and seeded in 96-well
plates. Every 24 h after cell seeding, the culture medium of the
96-well plate was replaced with DMEM, and then incubated at 37°C
for 4 h. Next, DMEM was replaced with isopyknic dimethyl sulfoxide.
Following 5 shocks, each of 1 sec duration, the absorbance at 490
nm was detected by SpectraMax 190 (Molecular Devices, LLC,
Sunnyvale, CA, USA) to measure the optical density (OD) value.
Wound-healing assay
Wound-healing assay was performed in 6-well culture
plates. In brief, 5×105 cells were suspended in medium
containing 10% FBS, and were then seeded into 6-well plates until
monolayer cells were nearly confluent. A 200-µl sterile pipette tip
was used to equably wound the monolayer cells, and the cells were
washed with PBS twice to remove the floating cells. Following
incubation in DMEM supplemented with 1% FBS for 24 h, images of the
wounded monolayers of blank, vector and CADM2 glioma cell groups
were captured under a phase-contrast microscope (magnification,
×20; Olympus Corporation, Tokyo, Japan).
Transwell invasion assay
QCM 24-Well Cell Invasion assay kit (EMD Millipore,
Billerica, MA, USA) was used for the Transwell invasion assay
processed by a Transwell filter coated with Matrigel. Cells were
seeded onto the top side of the Transwell filter on the top
chamber. DMEM supplemented with 10% FBS was added to the bottom
chamber. Cells (0.5×105) were suspended in 250 µl
serum-free DMEM. After 72 h incubation, the residual cells were
removed from the top chamber with a cotton swab, and the invaded
cells were dislodged from the lower membrane surface using
detachment solution. The detachment cells were stained with Hoechst
33342 at 37°C for 30 min, and then images of the detached cells
were directly captured and the number of cells was counted under a
fluorescence microscope (magnification, ×20). Cells were counted in
at least 5 randomly selected fields of view for each group. The
number of invaded cells per field was calculated and data are
presented as the mean ± standard error of the mean.
Animal studies
The stably transfected negative control (NC) and
CADM2 lentivirus U87 cells (2×106 of each) were
suspended in PBS and then implanted subcutaneously into 10 male
Balb/c nude mice aged 4 weeks. The mice were purchased from Vital
River Laboratories Co., Ltd. (Beijing, China). The length (a) and
width (b) of NC and CADM2 mice tumors were monitored every 2 days.
After 3 days of experiments, the nude mice were sacrificed and the
weights of the animals ranged between 24.3 and 25.8 g. Tumor volume
(V) was calculated using the formula V = ab2/2. Animal
experiments in the present study had been approved by the Ethics
Committee of Tangdu Hospital Institutional Review Board.
Statistical analysis
Data are presented as the mean ± standard deviation
of at least three independent experiments. Significance between
groups was analyzed by one-way analysis of variance, followed by a
Newman-Keuls comparison test using GraphPad Prism version 5.03
(GraphPad Software, Inc., La Jolla, CA, USA). *P<0.05,
**P<0.01 and ***P<0.001 were considered to indicate
statistically significant differences.
Results
CADM2 expression is downregulated in
human glioma and predicts a poor prognosis in patients with
glioma
To investigate the candidate role of CADM2 in human
glioma, the data of CADM2 in the Oncomine database was initially
referred to. It was demonstrated that the mRNA expression level of
CADM2 was markedly decreased in tumor tissues (98 samples) compared
with normal brain tissues (3 samples; Fig. 1A). Tissue microarrays were performed
on 60 glioma tissues (grade I, II, III and IV), 10 cancer adjacent
non-cancerous brain tissues and 10 normal brain tissues to compare
the protein expression level of CADM2 in normal tissues and
different grades of the malignancy (Fig. 1A). Tissue microarray results
demonstrated that CADM2 expression was much lower in higher grade
tissues, particularly grade II, III and IV glioma samples, than in
the grade I samples (Fig. 1B). The
CADM2 expression levels in different grades of glioma tissues and
normal tissues were studied by immunohistochemistry. Representative
images of these tissues are presented in Fig. 1C. Next, survival analysis was
performed to study the association between CADM2 expression and
survival times in patients with glioma. The Kaplan-Meier survival
curves presented in Fig. 1D
revealed that glioma patients with high CADM2 expression had a
longer overall survival time than those with low CADM2 expression
(P<0.05). Western blot analysis also verified the lower
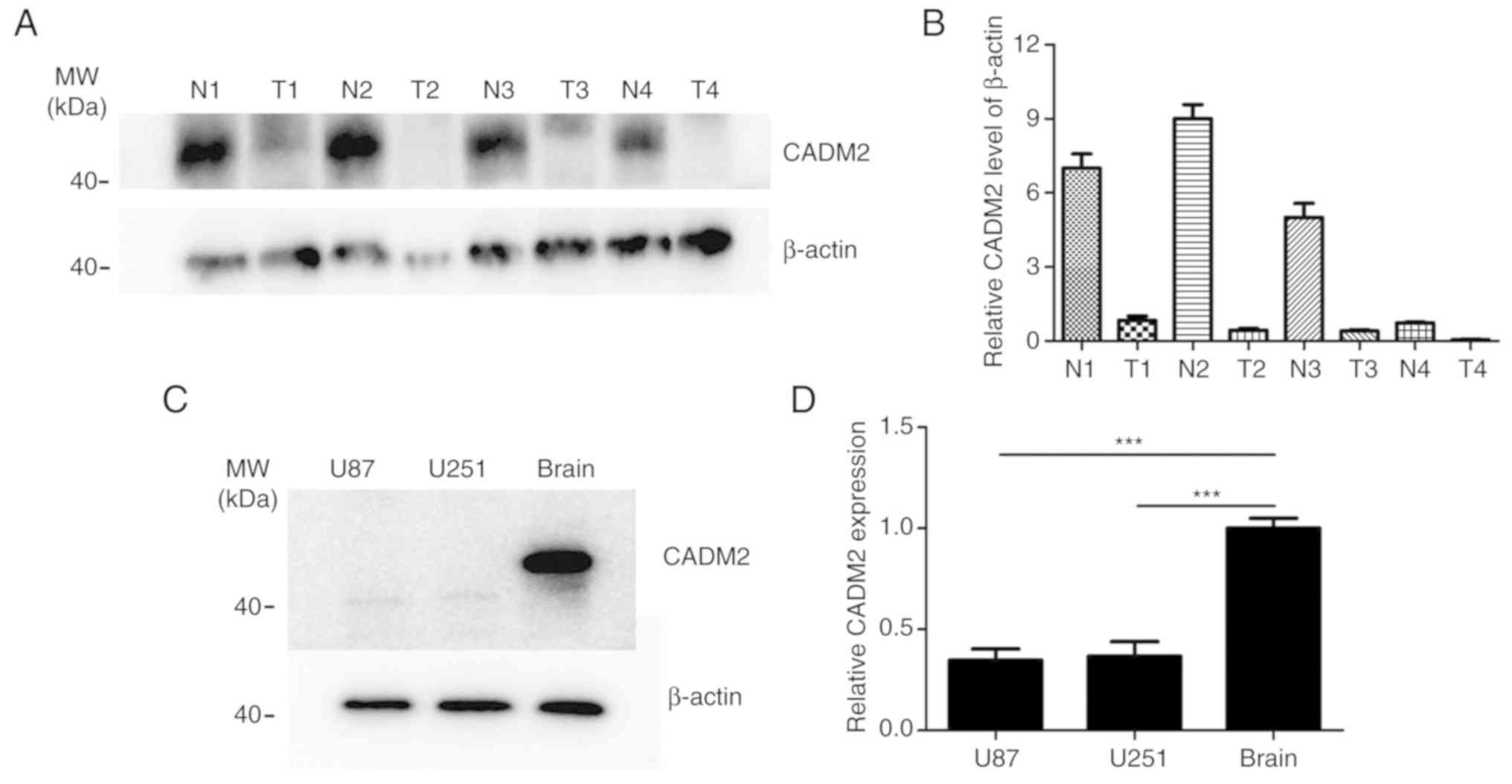
expression level of CADM2 in 4 GBM tissues, compared with the 4
corresponding adjacent non-cancerous brain tissues (Fig. 2A and B). The results also
demonstrated that CADM2 exhibited a much lower expression in glioma
tumor tissues. In addition, CADM2 protein expression in the U87 and
U251 cells was almost undetectable, while in normal brain cells it
was particularly high (Fig. 2C),
and the mRNA expression level of CADM2 in the GBM U87 and U251 cell
lines was ~60% lower than in the normal brain (Fig. 2D). Taken together, these data
indicated that the low expression level of CADM2 at the mRNA and
protein expression levels was observed in human glioma.
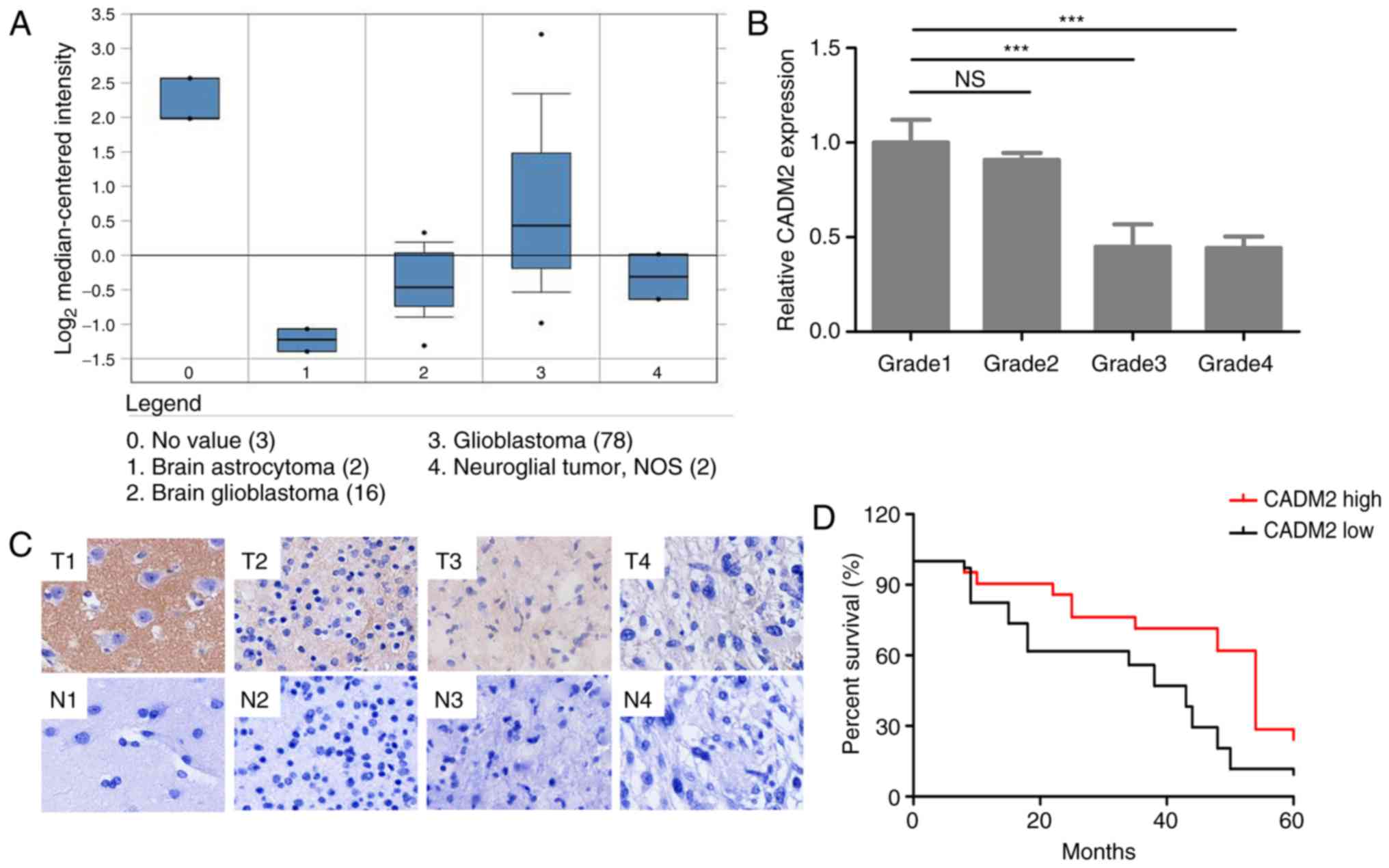 | Figure 1.Bioinformatics analysis of CADM2
expression in human glioma. (A) CADM2 expression in the Oncomine
database. (B) Tissue microarray analysis of CADM2 expression in
human tissues. (C) Representative images of tissue microarray
glioma tissue immunohistochemical staining (T1, T2, T3 and T4
represent no. 79, no. 20, no. 23 and no. 41 of the microarray
tissues, which are normal brain, and grade II, grade III and grade
IV glioma tissues, respectively. N1, N2, N3 and N4 represent the
corresponding adjacent normal tissues). (D) Kaplan-Meier survival
curves of patients with glioma based on the protein expression
levels of CADM2. *P<0.05 and ***P<0.001. CADM2, cell adhesion
molecule 2; ns, not significant; T, tumor; N, non-tumor. |
Downregulation of CADM2 is associated
with aggressive tumor progression of human gliomas
To verify the association between CADM2 expression
and GBM development, all 98 patients were stratified into CADM2-low
and CADM2-high groups using the median value of CADM2 expression.
Of the 98 patients with gliomas, 80 (81.63%) belonged to the
CADM2-low and 18 (18.37%) to the CADM2-high groups, respectively.
As demonstrated in Table I, low
CADM2 expression occurred more frequently in glioma patients with
advanced WHO grade (II–IV). However, no statistically significant
differences were observed between CADM2 expression and other
demographic characteristics, including patient age and sex
(P>0.05; Table I).
 | Table I.Associations between CADM2 protein
expression and various clinicopathological characteristics of
patients with glioma. |
Table I.
Associations between CADM2 protein
expression and various clinicopathological characteristics of
patients with glioma.
| Clinicopathological
feature | No. cases | Low CADM2 expression,
n (%) | High CADM2
expression, n (%) | P-value |
|---|
| WHO grade |
|
|
| <0.001 |
| IV | 38 | 36 (94.74) | 2 (5.26) |
|
| III | 29 | 26 (89.66) | 3 (10.34) |
|
| II | 17 | 14 (82.35) | 3 (17.65) |
|
| I | 14 | 4 (28.57) | 10 (71.43) |
|
| Age, years |
|
|
| 0.352 |
|
<55 | 44 | 39 (88.64) | 5 (11.36) |
|
|
≥55 | 54 | 47 (87.04) | 7 (12.96) |
|
| Sex |
|
|
| 0.438 |
|
Male | 55 | 49 (89.09) | 6 (10.91) |
|
|
Female | 43 | 38 (88.37) | 5 (11.63) |
|
| KPS score |
|
|
| 0.025 |
|
<80 | 45 | 38 (84.44) | 7 (5.56) |
|
|
≥80 | 53 | 45 (84.91) | 8 (5.09) |
|
CADM2 inhibits glioma cell
proliferation in vitro and in vivo
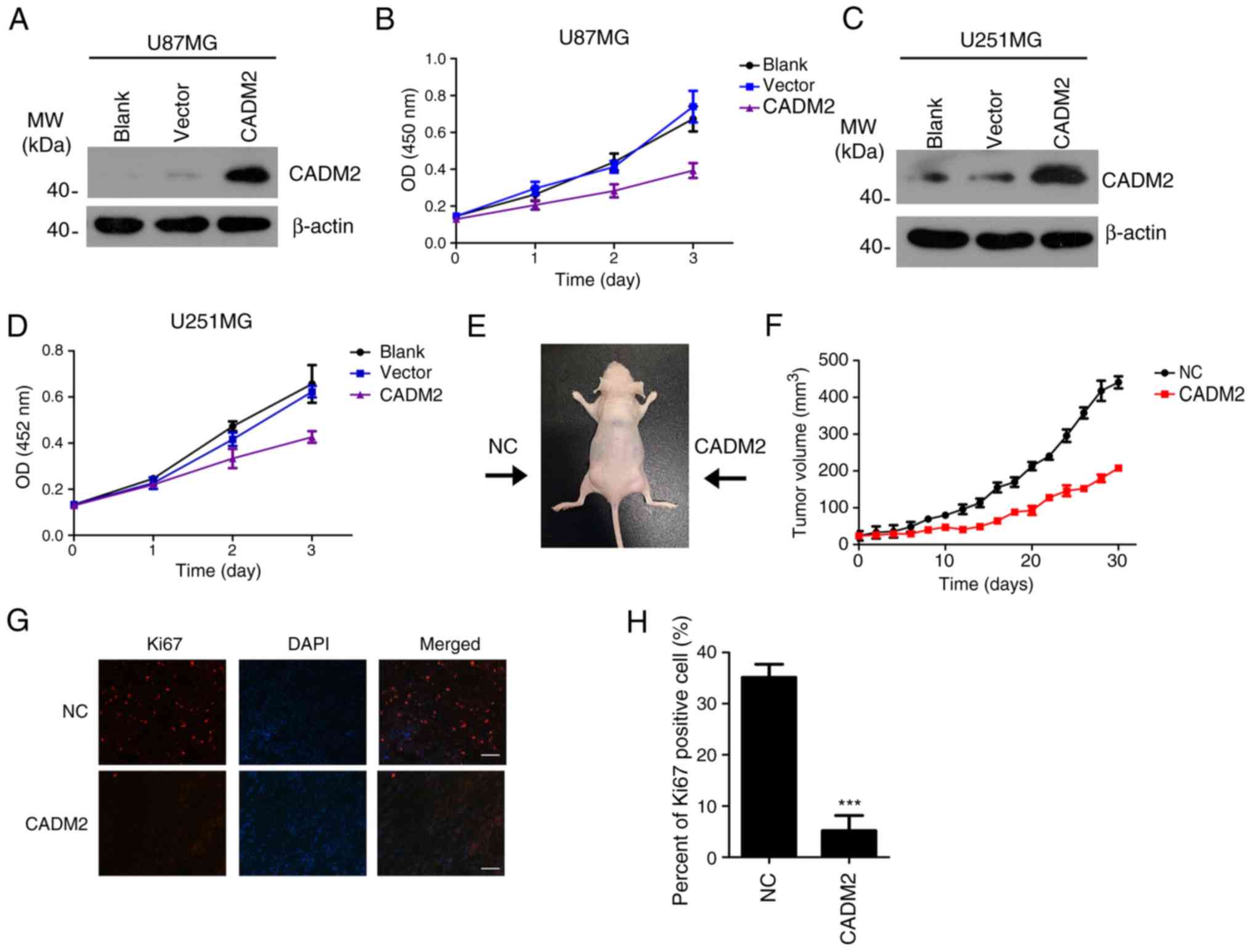
To investigate the biological function of CADM2 on
glioma tumorigenesis, CADM2 was overexpressed in U87 and U251 cell
lines, respectively (Fig. 3A and
C). The effect of CADM2 on glioma cell proliferation was
measured using an MTT cell proliferation assay. The results
demonstrated that the overexpression of CADM2 significantly
inhibited the proliferation of U87 and U251 compared with the blank
group and the vector group (Fig. 3B and
D). Nude mice xenograft tumors were created to verify the
effect of CADM2 on glioma proliferation in vivo. The
CADM2-overexpressing U87MG cells and the control U87MG cells were
suspended in PBS and implanted into two sides of the mice. After 30
days, the tumor volume results demonstrated that CADM2
significantly inhibited U87MG xenograft tumor growth in
vivo. The mean volumes of CADM2-U87 tumors cells were almost
half the size of that of the control U87MG cells (n=10 animals per
group, Fig. 3E and F). Ki-67
staining revealed that there were fewer Ki-67-positive cells in
CADM2-U87MG tumors than in U87MG control tumors (Fig. 3G and H). Therefore, these results
demonstrated that the CADM2 overexpression significantly inhibited
glioma proliferation in vitro and in vivo.
CADM2 overexpression inhibits glioma
cell migration and invasion
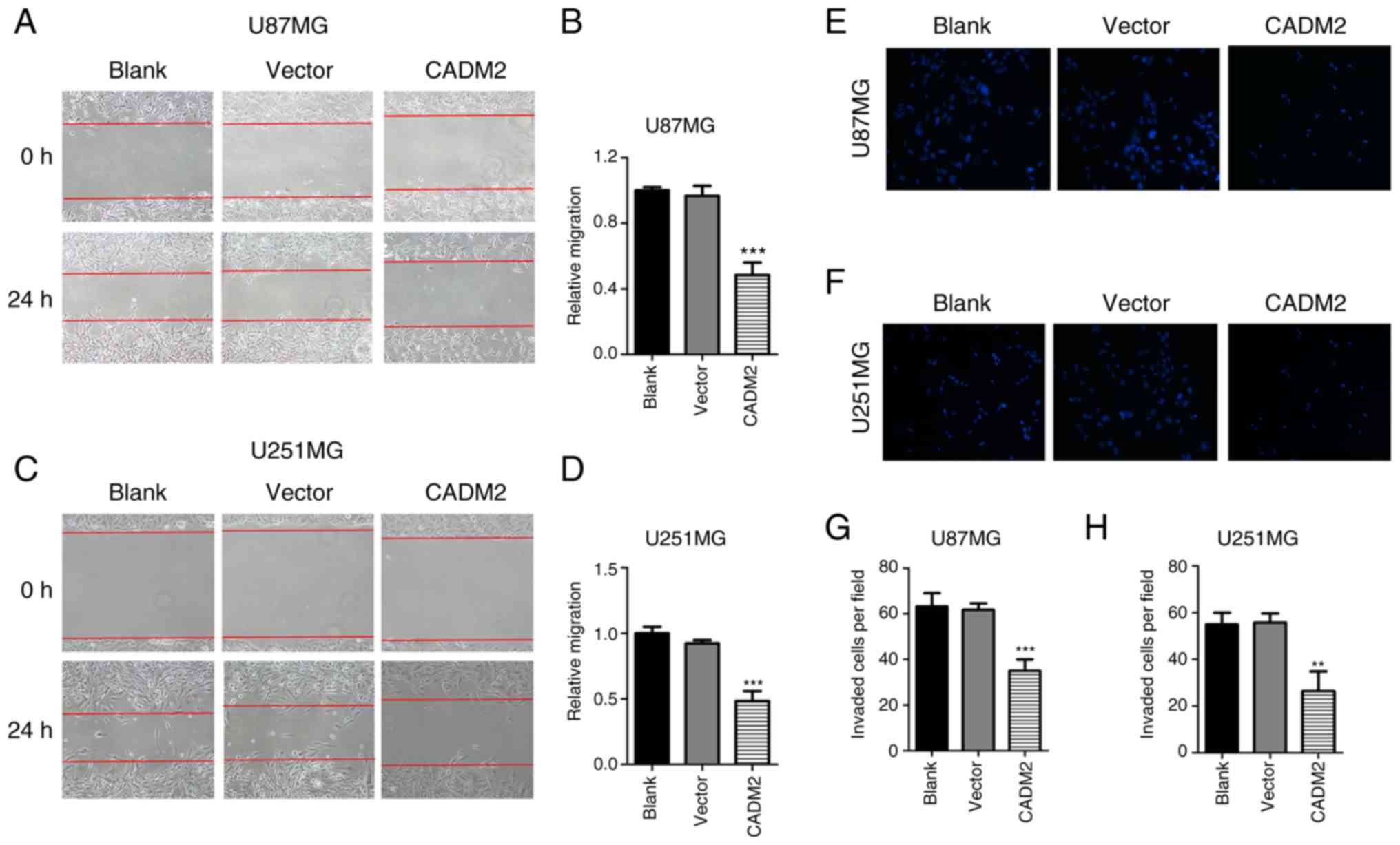
Next, a wound-healing assay and a Transwell assay
were performed to study the role of CADM2 on glioma cell migration
and invasion. In the wound-healing assay, the healing rate was
calculated by the healing distance divided by the initial distance
in order to demonstrate the cell migration ability. The results
indicated that CADM2 inhibited the healing rate of glioma cells,
and the inhibition rate of U87 and U251 cells was 57.6% (Fig. 4A and B) and 52.5% (Fig. 4C and D), respectively. The Transwell
experiment was processed to examine the migratory capacity of
glioma cells, and at least three wells were used for each
experiment. The results demonstrated that the overexpression of
CADM2 markedly attenuated the invasive properties of U87 and U251
cells; the number of invaded cells per field declined from 62 to
38/field for U87 cells (Fig. 4E and
G) and from 52 to 25/field for U251 cells (Fig. 4F and H), respectively. The results
indicated that CADM2 could inhibit the migratory and invasive
abilities of glioma cells.
CADM2 regulates the cell cycle and
EMT-related genes in glioma cells
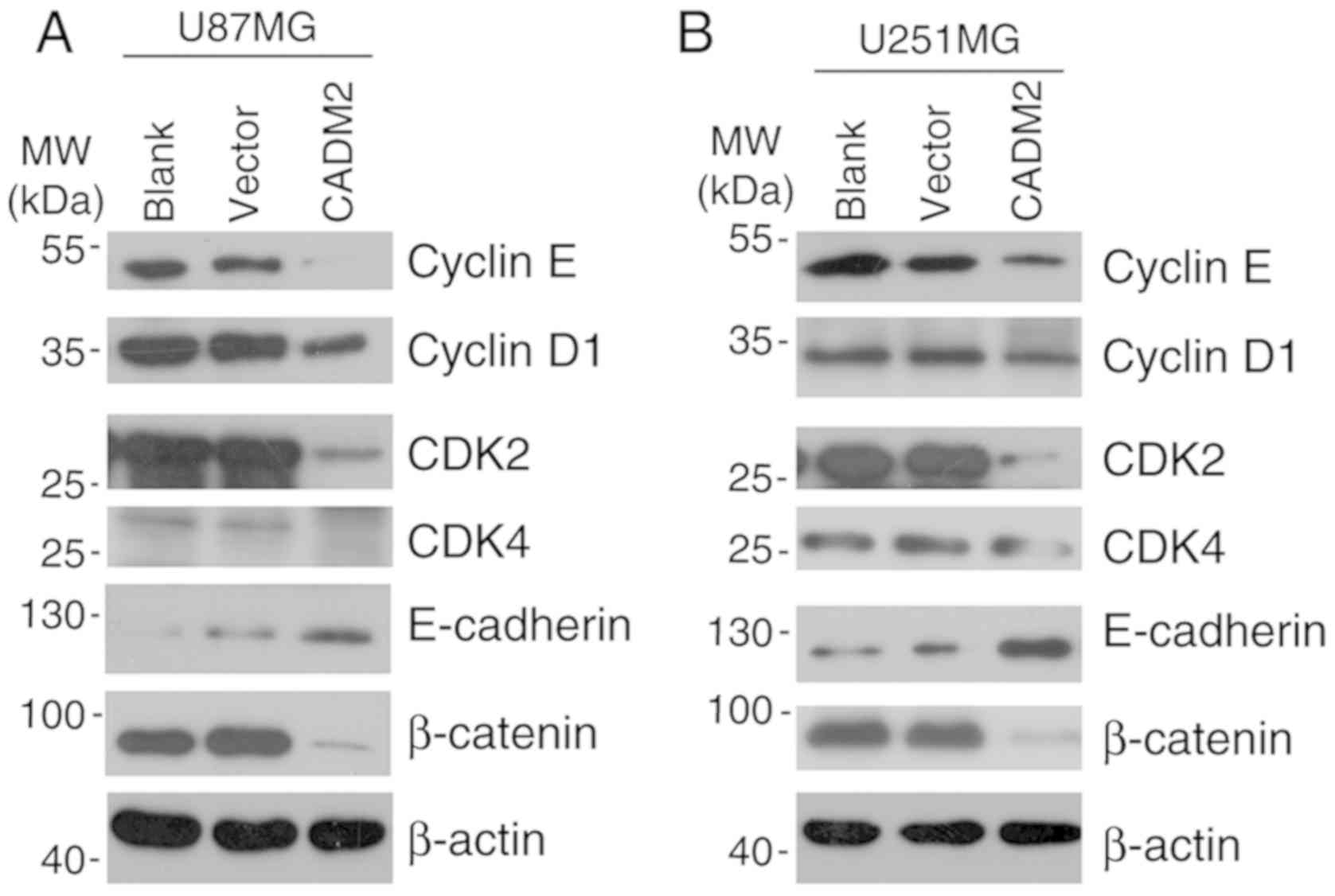
CADMs were reported as key regulators in G1/S
transition. The results of the present study demonstrated that
CADM2 overexpression significantly decreased the protein expression
levels of cyclin D1, CDK4, cyclin E1 and CDK2, compared with the
blank group (Fig. 5A and B). The
EMT is a crucial process for the invasion of cancer cells (17,18).
Next, the expression of two EMT markers, E-cadherin and β-catenin,
was detected by western blot analysis. The results demonstrated
that CADM2 significantly downregulated the protein expression level
of β-catenin and markedly increased the expression level of
E-cadherin in U87 cells (Fig. 5A)
and U251 cells (Fig. 5B). These
data suggested that CADM2 may inhibit glioma cell proliferation by
regulating cell cycle related genes and inhibit glioma cell
invasion via regulating the glioma cell EMT process.
Discussion
Glioma is one of the most common primary malignant
tumors of the brain parenchyma, which is also one of the most
difficult types of cancer to cure. Although a large number of
advances have been achieved in surgery, radiation therapy and
chemotherapy over recent decades, the prognosis of malignant glioma
remains poor. Comprehensive treatment with specific molecular
targets is thought to be a promising therapeutic strategy.
Therefore, it is urgent to identify novel and efficient targets for
glioma therapy.
Recent studies have reported that the CADMs family
function as tumor suppressors (6,7). In
numerous types of malignant tumors, the CADMs family are not
expressed or are only expressed at a low level. For example, CADM1
is reported to be downregulated in lung (19), prostate (20), esophageal (21) and breast cancer (22). CADM3 and CADM4 have also been
reported to suppress the growth of numerous types of cancer
(23–25). Among the CADMs family, the role of
CADM2, as well as that of other members, in cancer has not yet been
characterized (10). According to
the limited number of previous studies, CADM2 may participate in
maintaining cell polarity and decrease the adhesion of renal cell
carcinoma (cRCC) (6). The
downregulation of CADM2 in cRCC is probably due to promoter
methylation, and the CADM2 promoter methylation is associated with
tumor grade in primary cRCC (6).
Promoter methylation may mediate the mechanism by which CADM2
downregulation occurs in cRCC. A previous study demonstrated that
CADM2 promoter methylation was correlated with tumor stage and
grade in primary cRCC, and the expression of CADM2 was restored in
selected cell lines following 5-azadC treatment (6). However, there is no clear conclusion
regarding the expression and clinical significance of CADM2 in
human brain gliomas so far.
To the best of our knowledge, the present study was
the first regarding the biological function of CADM2 in human
glioma. Based on tissue microarray analysis, combined with qPCR
detection in glioma tissues and cell lines, it was demonstrated
that CADM2 is significantly downregulated in human glioma. The
expression intensity of CADM2 was significantly correlated with
patient pathological grade. The present study demonstrated that
CADM2 overexpression inhibited glioma cell proliferation in
vitro and in vivo. In addition, CADM2 also markedly
inhibited the migration and invasion of glioma cells. It was
further demonstrated that CADM2 regulated the expression of cell
cycle associated genes and EMT associated genes. In conclusion,
CADM2 inhibits glioma cell proliferation, migration and invasion by
regulating the glioma cell cycle and EMT. CADM2 may be a novel
therapeutic target for glioma treatment.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Scientific Foundation of China (grant no. 81572983), the
Foundation of Science Innovation and Development in Tangdu
Hospital, Fourth Military Medical University (grant no.
2016JCYJ013), the Natural Scientific Foundation of Shaanxi Province
(grant no. 2014JM4148) and the Beijing Key Laboratory of Brain
Major Diseases Open Project (grant no. 2015NZDJ02).
Availability of data and materials
All data generated and/or analyzed during this study
are included in this published article.
Authors' contributions
YT and YC conceived and designed the study; NL
performed the majority of the experiments and wrote the majority of
the manuscript; CY and WB collected clinical information, performed
statistical analyses and wrote part of the manuscript; XW and MJ
also gave academic guidance during the process of experimental
design; HY gave guidance and constructive suggestions in the animal
experiment section; ZW, WW, PZ and HL assisted with PCR, western
blotting and in vitro experiments. All authors read and
approved the manuscript, and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Tangdu Hospital, and written informed consent was
obtained from all patients prior to sample collection.
Patient consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
References
|
1
|
Dunn GP, Rinne ML, Wykosky J, Genovese G,
Quayle SN, Dunn IF, Agarwalla PK, Chheda MG, Campos B, Wang A, et
al: Emerging insights into the molecular and cellular basis of
glioblastoma. Genes Dev. 26:756–784. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Oike T, Suzuki Y, Sugawara K, Shirai K,
Noda SE, Tamaki T, Nagaishi M, Yokoo H, Nakazato Y and Nakano T:
Radiotherapy plus concomitant adjuvant temozolomide for
glioblastoma: Japanese mono-institutional results. PLoS One.
8:e789432013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fiorica F, Carau B, Ursino S and Cartei F:
Radiotherapy plus concomitant and adjuvant temozolomide for
unresectable glioblastoma: A retrospective analysis of our
experience. Radiother Oncol. 81:S297. 2006.
|
|
5
|
Hargreaves SJ, Williams M, Liu Z, et al:
Survival in patients receiving radiotherapy plus concomitant and
adjuvant temozolomide (RCAT) for glioblastoma (GBM). Int J Radiat
Oncol Biol Phys. 78:S281–S282. 2010. View Article : Google Scholar
|
|
6
|
He W, Li X, Xu S, Ai J, Gong Y, Gregg JL,
Guan R, Qiu W, Xin D, Gingrich JR, et al: Aberrant methylation and
loss of CADM2 tumor suppressor expression is associated with
human renal cell carcinoma tumor progression. Biochem Biophys Res
Commun. 435:526–532. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang S, Yan HL, Tao QF, Yuan SX, Tang GN,
Yang Y, Wang LL, Zhang YL, Sun SH and Zhou WP: Low CADM2 expression
predicts high recurrence risk of hepatocellular carcinoma patients
after hepatectomy. J Cancer Res Clin Oncol. 140:109–116. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Takai Y, Irie K, Shimizu K, Sakisaka T and
Ikeda W: Nectins and nectin-like molecules: Roles in cell adhesion,
migration, and polarization. Cancer Sci. 94:655–667. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Murakami Y: Involvement of a cell adhesion
molecule, TSLC1/IGSF4, in human oncogenesis. Cancer Sci.
96:543–552. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chang GM, Xu SP, Dhir R, Chandran U,
O'Keefe DS, Greenberg NM and Gingrich JR: Hypoexpression and
epigenetic regulation of candidate tumor suppressor gene
CADM-2 in human prostate cancer. Clin Cancer Res.
16:5390–5401. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cody NA, Shen Z, Ripeau JS, Provencher DM,
Mes-Masson AM, Chevrette M and Tonin PN: Characterization of the
3p12.3-pcen region associated with tumor suppression in a
novel ovarian Cancer cell line model genetically modified by
chromosome 3 fragment transfer. Mol Carcinog. 48:1077–1092. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lake SL, Coupland SE, Taktak AF and Damato
BE: Whole-genome microarray detects deletions and loss of
heterozygosity of chromosome 3 occurring exclusively in
metastasizing uveal melanoma. Invest Ophthalmol Vis Sci.
51:4884–4891. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Roy D, Sin SH, Damania B and Dittmer DP:
Tumor suppressor genes FHIT and WWOX are deleted in primary
effusion lymphoma (PEL) cell lines. Blood. 118:e32–e39. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tu YY, Gao XC, Li G, Fu H, Cui D, Liu H,
Jin W and Zhang Y: MicroRNA-218 inhibits glioma invasion,
migration, proliferation, and cancer stem-like cell self-renewal by
targeting the polycomb group gene Bmi1. Cancer Res. 73:6046–6055.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Allen M, Bjerke M, Edlund H, Nelander S
and Westermark B: Origin of the U87MG glioma cell line: Good news
and bad news. Sci Transl Med. 8:354re32016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Micalizzi DS, Farabaugh SM and Ford HL:
Epithelial-mesenchymal transition in cancer: Parallels between
normal development and tumor progression. J Mammary Gland Biol
Neoplasia. 15:117–134. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee MY, Chou CY, Tang MJ and Shen MR:
Epithelial mesenchymal transition in cervical cancer: Correlation
with tumor progression, epidermal growth factor receptor
overexpression, and snail up-regulation. Clin Cancer Res. 14:47–43.
2008. View Article : Google Scholar
|
|
19
|
Kuramochi M, Fukuhara H, Nobukuni T, Kanbe
T, Maruyama T, Ghosh HP, Pletcher M, Isomura M, Onizuka M, Kitamura
T, et al: TSLC1 is a tumor-suppressor gene in human
non-small-cell lung cancer. Nat Genet. 27:427–430. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fukuhara H, Kuramochi M, Fukami T,
Kasahara K, Furuhata M, Nobukuni T, Maruyama T, Isogai K, Sekiya T,
Shuin T, et al: Promoter methylation of TSLC1 and tumor
suppression by its gene product in human prostate cancer. Jap J
Cancer Res. 93:605–609. 2002. View Article : Google Scholar
|
|
21
|
Ito T, Shimada Y, Hashimoto Y, Kaganoi J,
Kan T, Watanabe G, Murakami Y and Imamura M: Involvement of TSLC1
in progression of esophageal squamous cell carcinoma. Cancer Res.
63:6320–6326. 2003.PubMed/NCBI
|
|
22
|
Takahashi Y, Iwai M, Kawai T, Arakawa A,
Ito T, Sakurai-Yageta M, Ito A, Goto A, Saito M, Kasumi F, et al:
Aberrant expression of tumor suppressors CADM1 and 4.1B in invasive
lesions of primary breast cancer. Breast Cancer. 19:242–252. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Williams YN, Masuda M, Sakurai-Yageta M,
Maruyama T, Shibuya M and Murakami Y: Cell adhesion and prostate
tumor-suppressor activity of TSLL2/IGSF4C, an immunoglobulin
superfamily molecule homologous to TSLC1/IGSF4. Oncogene.
25:1446–1453. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Raveh S, Gavert N, Spiegel I and Ben-Ze'ev
A: The cell adhesion nectin-like molecules (Necl) 1 and 4 suppress
the growth and tumorigenic ability of colon cancer cells. J Cell
Biochem. 108:326–36. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nagata M, Sakurai-Yageta M, Yamada D, Goto
A, Ito A, Fukuhara H, Kume H, Morikawa T, Fukayama M, Homma Y, et
al: Aberrations of a cell adhesion molecule CADM4 in renal clear
cell carcinoma. Int J Cancer. 130:1329–1337. 2012. View Article : Google Scholar : PubMed/NCBI
|