Introduction
Neuroblastoma (NB) is the most common extracranial
solid tumor in children and is responsible for 10% of the mortality
resulting from all pediatric tumors (1,2). The
prognosis of many children's malignant tumors has improved by
effective treatments. However, high-risk NB remains intractable,
with merely 40% of patients achieving long-term survival despite
the availability of multiple therapeutic methods (3,4). The
tumorigenesis and progression of NB is reported to be complex, and
is influenced by both the external environment and internal genetic
factors (4,5). Early accurate diagnosis and timely
intervention is important to improve prognosis. Therefore, further
research investigating the biological targets and underlying
molecular mechanisms is essential for improving the diagnosis and
therapy of NB (6).
Long non-coding RNAs (lncRNAs) are generally defined
as RNA transcripts that are >200 nt with no protein products
(7–9).
Non-coding RNAs, once considered as transcriptional waste, are
actually a part of the regulatory network of transcriptional and
post-transcriptional processes (1,10,11). They are ubiquitously expressed in
mammalian genomes, and participate in the regulation of numerous
biological processes, including DNA methylation, chromatin
remodeling, transcription regulation and translation (10,12). It is
reported that several lncRNAs are closely linked to NB initiation
and progression, and involved in a variety of tumor-associated
biological processes (13,14), such as cancer susceptibility 15
(CASC15) (15), MYCN upstream
transcript (lncUsMycN) (16) and ETS1
antisense RNA 1 (pancEts-1) (17).
In our previous study, genome-wide lncRNA analysis
was performed to detect NB-associated lncRNAs. Linc01105 was
identified as differentially expressed between NB and normal
adrenal gland tissues. It was also demonstrated that upregulation
of linc01105 in NB was correlated with the International
Neuroblastoma Staging System (INSS) stage (14). The present study found that hypoxia
inducible factor-1α (HIF-1α) may bind at the linc01105 promoter to
activate its transcription. In addition, linc01105 knockdown
significantly suppressed proliferation and induced apoptosis via
the p53/caspase signaling pathway. Furthermore, it is well
established that, lncRNAs can act as microRNA (miRNA) sponges, and
thus regulate their ability to target mRNAs (18,19). In
the present study, it was demonstrated that linc01105 may inhibit
tumor angiogenesis via the linc01105/miR-6769b-5p/vascular
endothelial growth factor A (VEGFA) network.
Materials and methods
Human tissues and NB cell line
Human NB and adjacent normal adrenal gland tissues
were collected from patients who underwent surgery between January
2011 and January 2017 in the Children's Hospital of Fudan
University, Shanghai, China. Tumor tissues were diagnosed as NB by
the pathology department and all patients were stage III/IV
according to INSS. For the present study, 32 patients were
enrolled,18 boys and 14 girls. The age of the patients ranged from
two months to ten years. Informed consent was acquired from every
patient's legal guardians. This study was approved by the Institute
Research Ethics Committee at the Children's Hospital of Fudan
University.
The NB cell line SH-SY5Y and SK-N-BE (2), the human umbilical vein endothelial
cells (HUVECs) and 293T cells were obtained from the Cell Bank of
Type Culture Collection of the Chinese Academy of Sciences in
Shanghai. CHLA15, LA-N-5, and CHLA136 were gifts from professor Li
Kai's lab of FuDan University. The SH-SY5Y and SK-N-BE (2) cells were cultured in DMEM/F12
(Biological Industries) with 10% fetal bovine serum (Biological
Industries) in a humidified incubator with 5% CO2 at
37°C. HUVEC, 293T, and other NB cell lines(CHLA15, LA-N-5, and
CHLA136 cells) were cultured in DMEM with 10% FBS (both Gibco;
Thermo Fisher Scientific, Inc.). All cell culture dishes and
culture plates were purchased from Hangzhou Xinyou Biotechnology
Co., Ltd, China.
Nuclear and cytoplasmic extract
preparation
Cells were grown to 90–100% confluency. Pre-cooled
PBS was used to wash the cells twice. Cytoplasmic extraction buffer
(0.5 ml; Invent Biotechnologies, Inc.) was then added to the cells,
and set on ice for 5 min. The lysates were transferred to tubes and
mixed rigorously for 15 sec. After centrifuging at 12,000 × g for 5
min at 4°C, TRIzol was added to the nuclear (pellet) and
cytoplasmic fragments (supernatant) for RNA extraction.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted using TRIzol reagent (Takara
Bio, Inc.) and cDNA was synthesized using a cDNA Reverse
Transcription kit (Takara Bio, Inc.). qPCR reactions were performed
using SYBR Green PCR Master Mix (Takara Bio, Inc.) on the Step One
Real-Time PCR system. The PCR program was: 94°C for 10 min, 40
cycles of 94°C for 60 sec and 60°C for 15 sec. GAPDH was used as
internal control. Data was analyzed using GraphPad software
(GraphPad Software, Inc.) and relative fold changes in expression
were calculated using the formula 2−ΔΔCq (20). The following primer sequences were
used in this study: Linc01105, forward 5′-TGTGCCATTCCATGTTATA-3′
and 5′-TTGAGGCTGAAGACCAAA-3; and GAPDH, forward
5′-GGAGCGAGATCCCTCCAAAAT-3′ and reverse
5′-GGCTGTTGTCATACTTCTCATGG-3′. U1 and actin were used as controls
for the nuclear and the cytoplasmic extracts, respectively, and
their primes were: U1, forward 5′-GACGGGAAAAGATTGAGCGG-3′ and
reverse 5′-GCCACGAAGAGAGTCTTGAAGG-3′; actin, forward
5′-CATGTACGTTGCTATCCAGGC-3′ and reverse,
5′-CTCCTTAATGTCACGCACGAT-3′.
To quantitatively measure miRNAs, the All-in-OneTM
miRNA qRT-PCR Detection kit (GeneCopoeia, Inc.) was used. The
experimental procedure included two major steps. Firstly,
single-step cDNA synthesis was performed. Poly-A polymerase was
used to add poly-A tails to the 3′ end of miRNAs, and M-MLV RTase
was used simultaneously, with a unique oligodT adaptor primer, to
reverse transcribe the miRNA. Secondly, qPCR detection was
performed. The All-in-OneTM miRNA qRT-PCR Mix, containing SYBR
Green, specifically detects the reverse transcribed miRNA. The
miR-6769b-5p-specific primer (cat. no. HmiRQP3816) and the U6
control primer (cat. no. HmiRQP9001) were purchased from
GeneCopoeia, Inc.
Construction of shRNA and stable
transfected cell lines
Linc01105 shRNA plasmids were designed and
constructed by Genomeditech Co., using the pGMLV-SC5 RNAi vector.
The control target sequence was 5′-TTCTCCGAACGTGTCACGT-3′. Three
different shRNA target sequences were designed as follows: sh1,
5′-GCTCAGGAGAAAGAGCAAATG-3′; sh2, 5′-GCCTGCTGAGAAGGCTCATCT-3′; and
sh3, 5′-GCAGCAACTCCTGTGCATGT-3′. Purified, endotoxin-free
lentiviral vector and its auxiliary packaging vector plasmids were
co-transfected into 293T cells (Cell Bank of Type Culture
Collection of the Chinese Academy of Sciences) using the HG
transgene reagent (Genomeditech Co.) for 10–12 h. Enhancing buffer
(Genomeditech Co.) was added, and the medium was replaced with
fresh medium after 8 h. After 48 h of incubation, the supernatant
of the cell line, containing the lentiviral particles, was
collected and concentrated to obtain a high titer of lentivirus
concentrate, with a final virus titer of 5×108
transduction units/ml. SH-SY5Y cells were infected with different
shRNA-expressing lentivirus, and stable infected cells were
acquired following puromycin selection.
Cell proliferation and viability
assay
Cell proliferation and viability were detected using
the Cell Counting Kit-8 assay (Yeasen Biotechnology Co., Ltd.) and
the cell viability kit (Beyotime Institute of Biotechnology),
according to the suppliers' protocols. Cells were cultured in a
96-well plate at a concentration of 2,000 cells per well in 100 µl
medium. The Cell-Light EdU DNA cell proliferation kit (Guangzhou
RiboBio Co., Ltd.) was also used to assess cell proliferation. For
the EdU assay, 10,000 cells were seeded in 24-well plates. Cells
were cultured for 2 h with 50 µM EdU, then 4% formaldehyde was used
to fix the cells for 20 min at room temperature. Cells were
permeabilized with 0.5% Triton X-100 for 10 min, stained with 200
µl of Hoechst 33342 for 30 min, and visualized under a fluorescent
microscope.
Cell apoptosis assay
After digestion with EDTA-free trypsin, cells were
collected and washed twice with pre-cooled PBS. Cells
(106) were transferred into 5 ml flow tubes. Then, 5 µl
of Annexin V-APC and 10 µl of 7-AAD (Yeasen Biotechnology Co.,
Ltd.) were added on the cells in the dark at room temperature for
15 min. Apoptosis rates (percentages of Q2+Q3) were then analyzed
by flow cytometry (BD Celesta; Beckman Coulter, Inc.) within 1
h.
Cell migration and invasion assay
Twelve-well culture plates with 8 µm micropore
inserts were used for cell migration and invasion assays. Cells
were serum-starved for 24 h prior to the assays. For the migration
assay, 2×105 NB cells were placed into the upper wells
in DMEM/F12 without FBS for 24 h. For the cell invasion assay, the
upper side of the insert was coated with Matrigel (BD Biosciences),
and 4×105 NB cells were placed into the upper wells
without FBS for 24 h. The cells on the lower sides of the inserts
were then fixed with 4% paraformaldehyde for 10 min and stained
with 0.1% crystal violet for 30 min. Five fields were randomly
selected and captured using a light microscope (Olympus
Corporation), and the average count of the five fields was
calculated.
Wound healing assay
Cells (1×105) were seeded in the two
wells of culture inserts (Ibidi GmbH). After the cells were
attached and the insert was removed, a standard gap was left
between the cells. The gap closure was observed at ×100
magnification and photographs were captured using light microscopy
at 0, 6, 12 and 24 h. Image J (National Institutes of Health) was
used to measure the gap width.
Caspase-3 activity assay
Caspase-3 activity was detected using the caspase-3
activity kit (Beyotime Institute of Biotechnology). According to
the manufacturer's instructions, cells were mixed with buffers
containing caspase-3 substrate and incubated at 37°C for 2 h.
Samples were the detected at 450 nm using a plate reader.
Western blotting
Cell proteins were extracted by lysis buffer
(Beyotime Institute of Biotechnology), and western blotting was
performed with standard procedures (21). Briefly, after blocking, the membranes
were incubated with primary antibodies at 4°C overnight. The
primary antibodies were as follows: Anti-BCL2 (cat. no. 60178;
1:2,000; ProteinTech Group, Inc.), anti-Bax (cat. no. 50599;
1:2,000; ProteinTech Group, Inc.), anti-caspase-3 (cat. no. 19677;
1:500; ProteinTech Group, Inc.), anti-caspase-9 (cat. No 10380;
1:500; ProteinTech Group, Inc.), anti-active caspase-3 (cat. no.
F021507; 1:500; Abways Technology), anti-cleaved caspase-9 (cat.
no. F016210; 1:500; Abways Technology), anti-p53 (cat. no. F024201;
1:500 Abways Technology), anti-poly (ADP-ribose) polymerase (PARP;
cat. no. AP102-1; 1:1,000; Beyotime Institute of Biotechnology),
anti-VEGFA (cat. no 19003; 1:1,000; ProteinTech Group, Inc.), and
anti-tubulin (cat. no. ab210797; 1:1,000; Abcam). After washing
with TBS/0.5% Tween 20 three times, the membranes were incubated
with the relevant secondary antibody (goat anti-rabbit cat. no.
CW0156, 1:2,000; or goat anti-mouse cat. no. CW0110, 1:5,000; both
ComWin Biotech Co., Ltd.) for 1 h at room temperature. The protein
signals were detected using an enhanced chemiluminescence substrate
(EMD Millipore).
Transient transfections
Cells (104) were seeded in a 24-well
plate one day in advance, and the cell confluence was ~30% at the
time of transfection. Small interfering RNA (siRNA) or miRNA mimics
or inhibitor (50 pmol) were added to OPTI-MEM (Gibco; Thermo Fisher
Scientific, Inc.) to a final volume of 25 µl and mixed well. Then,
1 µl of Lipofectamine 2000 (Thermo Fisher Scientific, Inc.) was
added to OPTI-MEM to a final volume of 25 µl. The two solutions
were then thoroughly mixed at room temperature for 15 min. The 50
µl of transfection complex was added to cells in 0.45 ml media, and
incubated for 48 h. The sequences were as follows: siVEGFA,
5′-GUGCUACUGUUUAUCCGUA-3′; negative control (NC) siNC,
5′-UUCUCCGAACGUGUCACGU-3′; miR-107 mimics,
5′-GCCUUCUGACUCCAAGUCCAGU-3′, miR-378a-3p mimics,
5′-UGAUAGCCCUGUACAAUGCUGCU-3′, miR-6769b-5p mimics,
5′-UCUACUCUUUCUAGGAGGUUGUGA-3′; mimics NC,
5′-GCACUUCUCCUCCCCACCCACCA-3′; miR-6769b-5p inhibitor,
5′-GCACUUCCUCCCCACCCACCA-3′; and inhibitor NC,
5′-UCACAACCUCCUAGAAAGAGUAGA-3′. HIF-1a and VEGFA plasmids (both in
pCDH vector) were purchased from Shanghai Generay Biotech Co, Ltd,
and transfected (4 µg) into cells with Lipofectamine 2000 (Thermo
Fisher Scientific, Inc.). After 48 h of transfection, subsequent
experiments were performed.
Luciferase assays
293T cells were seeded in 24-well plates. When cells
reached 75–85% density, they were transfected with luciferase
reporter plasmids containing the 3′ untranslated region (UTR) of
VEGF or the promoter of linc01105 (both constructed by Generay
Biotech Co., Ltd.) using Lipofectamine 2000, according to the
manufacturer's instructions. After 48 h, luciferase activity (Dual
Luciferase Reporter Gene Assay kit; Yeasen Biotechnology Co., Ltd)
was measured using a microplate reader and Renilla
luciferase activity was used for normalization.
Tube formation assay
HUVECs (5×103) were seeded into a 96-well
plate that was pre-coated with Matrigel (BD Biosciences).
Conditioned media (CM) supernatant from the stable-infected SH-SY5Y
cells were added to the HUVEC cells. A VEGFA antibody (cat. no.
19003; 1:500; ProteinTech Group, Inc.) was used for 6 h
pre-treatment in the tube formation assay, as indicated. After 6 h
of incubation, tube formation was observed using phase-contrast
light microscopy.
Database used for exploring
miRNAs
To determine whether linc01105 functions as a miRNA
sponge, first the DIANA-LncBase database was used to search for
potential miRNA recognition elements on linc01105 (http://diana.imis.athena-innovation.gr).
The search term was linc01105, and multiple miRNAs were predicted
to bind. The CircNet database (http://circnet.mbc.nctu.edu.tw) was used to search for
miRNAs that may bind with VEGFA. Finally, three miRNAs, miR-107,
miR-6769b-5p, miR-378a-3p were identified as potential miRNAs that
could bind with both linc01105 and VEGFA.
Chromatin immunoprecipitation
(ChIP)
ChIP assay was performed as described by Morelli
et al (22). The cells of one
10 cm dish were sonicated 4 times for 10 sec at pre-cooled
conditions (Fisher Sonic Dismembrator; Thermo Fisher Scientific,
Inc.) and then treated according to the standard protocol.
Anti-HIF-1 (cat. no. 39665; Active Motif, Inc.) or control rabbit
IgG (cat. no. 294670; Abmart, Inc.) antibodies were used to capture
chromatin fragments from cell extracts. PCR was used to amplify the
DNA fragment with the antibody and the input DNA was used as
control.
Chromatin isolation by RNA
purification (CHIRP)
Linc01105 antisense DNA (asDNA), β-galactosidase
(lacZ) asDNA and linc01105 sense DNA probes were designed using an
online probe designer (singlemoleculefish.com). Oligonucleotides were
biotinylated at the 3′ end with an 18-carbon spacer arm. Cells were
collected and subjected to CHIRP, using the method previously
described by Chu et al (23).
GO and KEGG analysis were performed using the DAVID Functional
Annotation web-based tool (http://david.ncifcrf.gov).
Statistical analysis
All results are presented as mean ± standard
deviation. Results from different groups were compared using the
Student's test or one-way ANOVA followed by Turkey multiple
comparisons test. Statistical analysis was performed using GraphPad
Prism 5 (GraphPad Software, Inc.). P<0.05 was considered to
indicate a statistically significant difference.
Results
Expression and localization of
linc01105
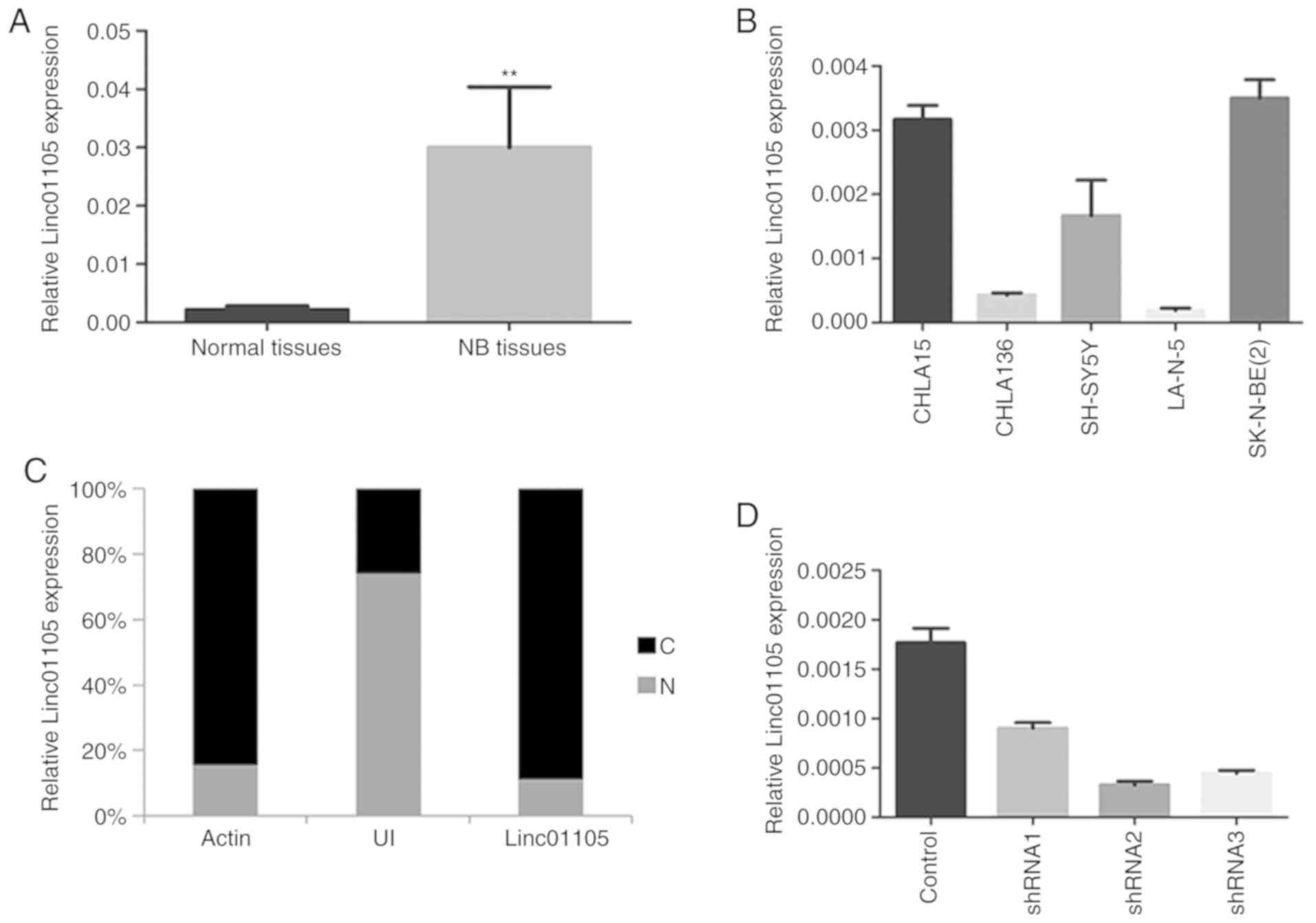
Linc01105 was significantly upregulated in the 32 NB
specimens compared with the matched normal adrenal gland tissues
(Fig. 1A). The SH-SY5Y (non-MYCN
amplification) cell line was selected as the in vitro model
for NB in the present study (Fig.
1B), as our previous study had used SK-N-BE (2)C (MYCN amplification) and CHLA15 was too
difficult to culture. Nuclear and cytosolic RNA of SH-SY5Y was
extracted and detected by RT-qPCR. The differential enrichments of
U1 small nuclear RNA and actin were used as controls for the
nuclear and the cytoplasmic extracts, respectively. The results
demonstrated that linc01105 expression levels were clearly
increased in the cytosol relative to the nucleus (Fig. 1C), which suggested that linc01105 was
mainly localized in the cytosol and thus may have a key role in
post-transcriptional events. In order to investigate the function
of linc01105, three individual shRNAs and a negative control shRNA
were purchased from Genomditech. Following lentiviral infection,
the RT-qPCR results confirmed that shRNA2 had the highest knockdown
efficiency, and thus was selected for subsequent experiments
(Fig. 1D).
Knockdown of linc01105 inhibits
proliferation and viability
To further explore the biological function of
linc01105 in NB cells, a shRNA2 stable-infected line was generated
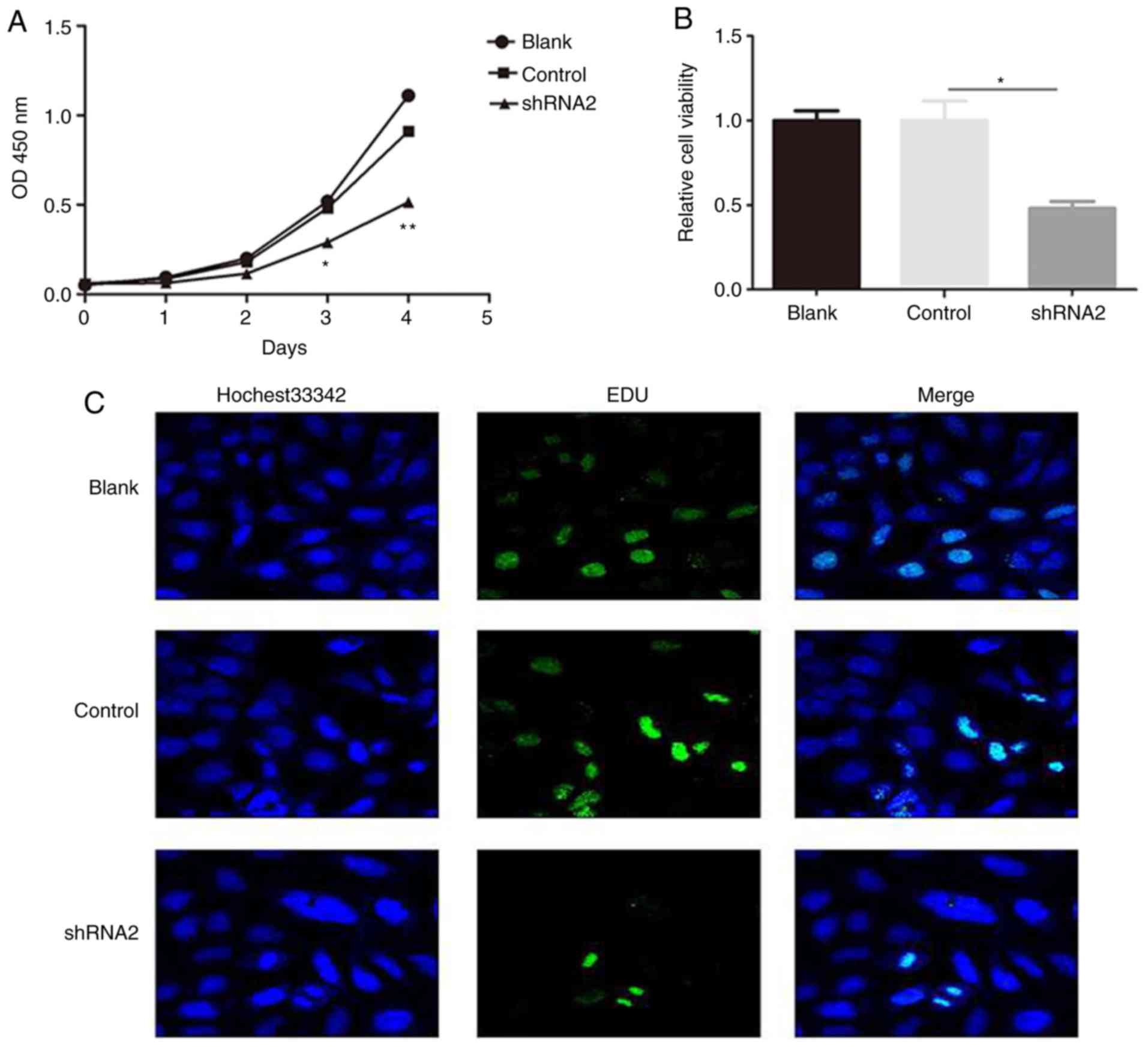
in SH-SY5Y cells. A CCK-8 assay was performed to investigate the
effect of linc01105 on cell proliferation. Silencing of linc01105
significantly inhibited NB cell proliferation compared with
negative control (Fig. 2A). In
addition, cell viability was reduced following linc01105 silencing
(Fig. 2B). Finally, similar results
were observed with the Cell-Light EdU DNA cell proliferation assay
(Fig. 2C). Taken together, these
results revealed that downregulation of linc01105 may inhibit cell
proliferation.
Knockdown of linc01105 contributes to
increased apoptosis
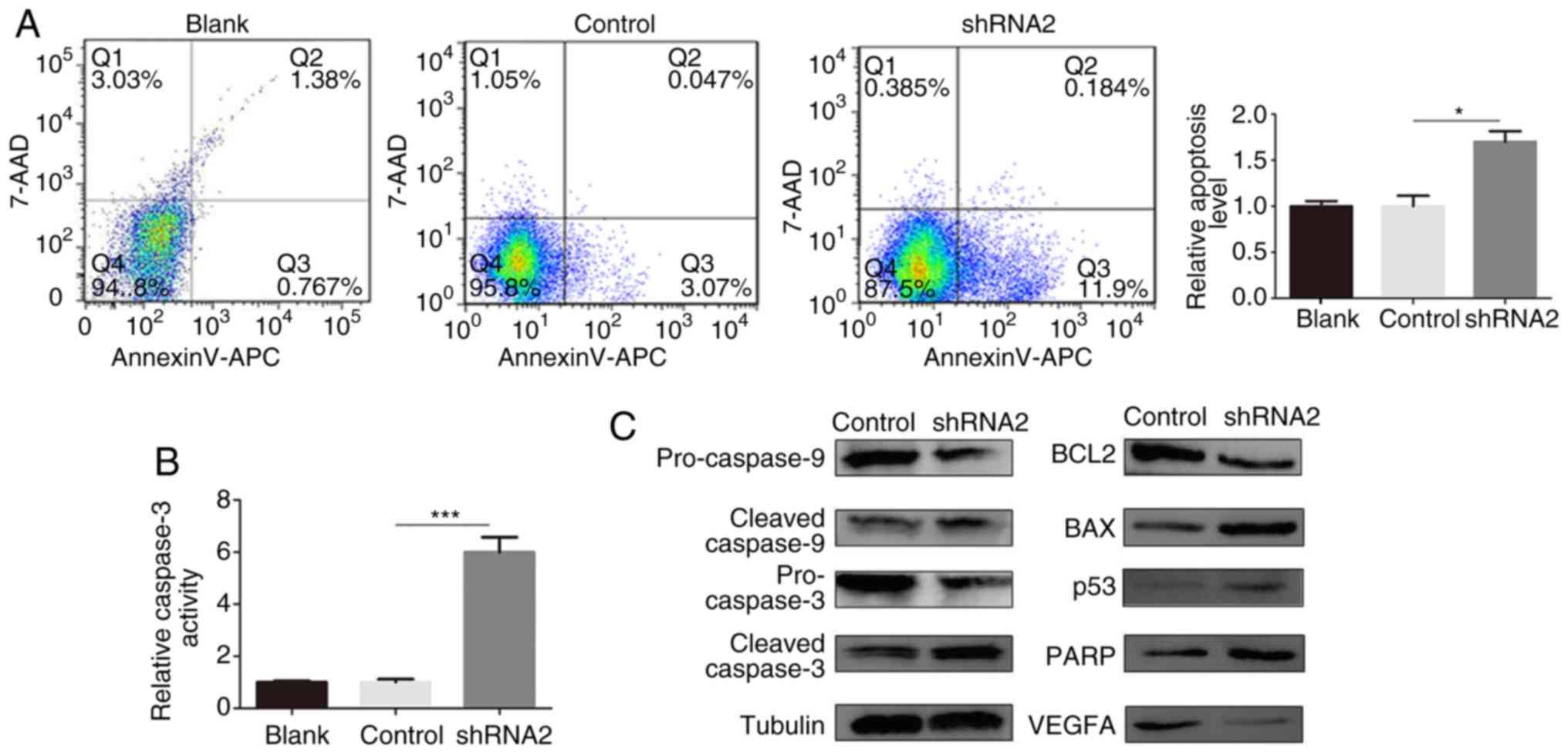
Compared with the control group, knockdown of
linc01105 induced cell apoptosis, as evidenced by flow cytometry
analysis (Fig. 3A). To investigate
this further, a caspase-3 activity assay was performed to evaluate
the activation of caspase-3 (Fig.
3B). Knockdown of linc01105 resulted in increased caspase-3
activation compared with the control group. Western blot analysis
for proteins associated with the Bcl-2 family and the p53/caspase
pathway further confirmed that linc01105 knockdown resulted in an
obvious apoptotic effect in NB cells (Fig. 3C). Bcl-2 family proteins and the
p53/caspase pathway are important regulatory factors of apoptosis
(24–26). As shown in Fig. 3C, the expression levels of the
antiapoptotic protein Bcl-2 were decreased, while the expression
levels of the pro-apoptotic protein Bax were increased following
Linc01105 knockdown. Consistent with this, pro-caspase9/3
expression levels were downregulated, while cleaved caspase9/3 and
PARP, a marker of caspase3 activation, were upregulated.
Knockdown of linc01105 inhibits
migration and invasion
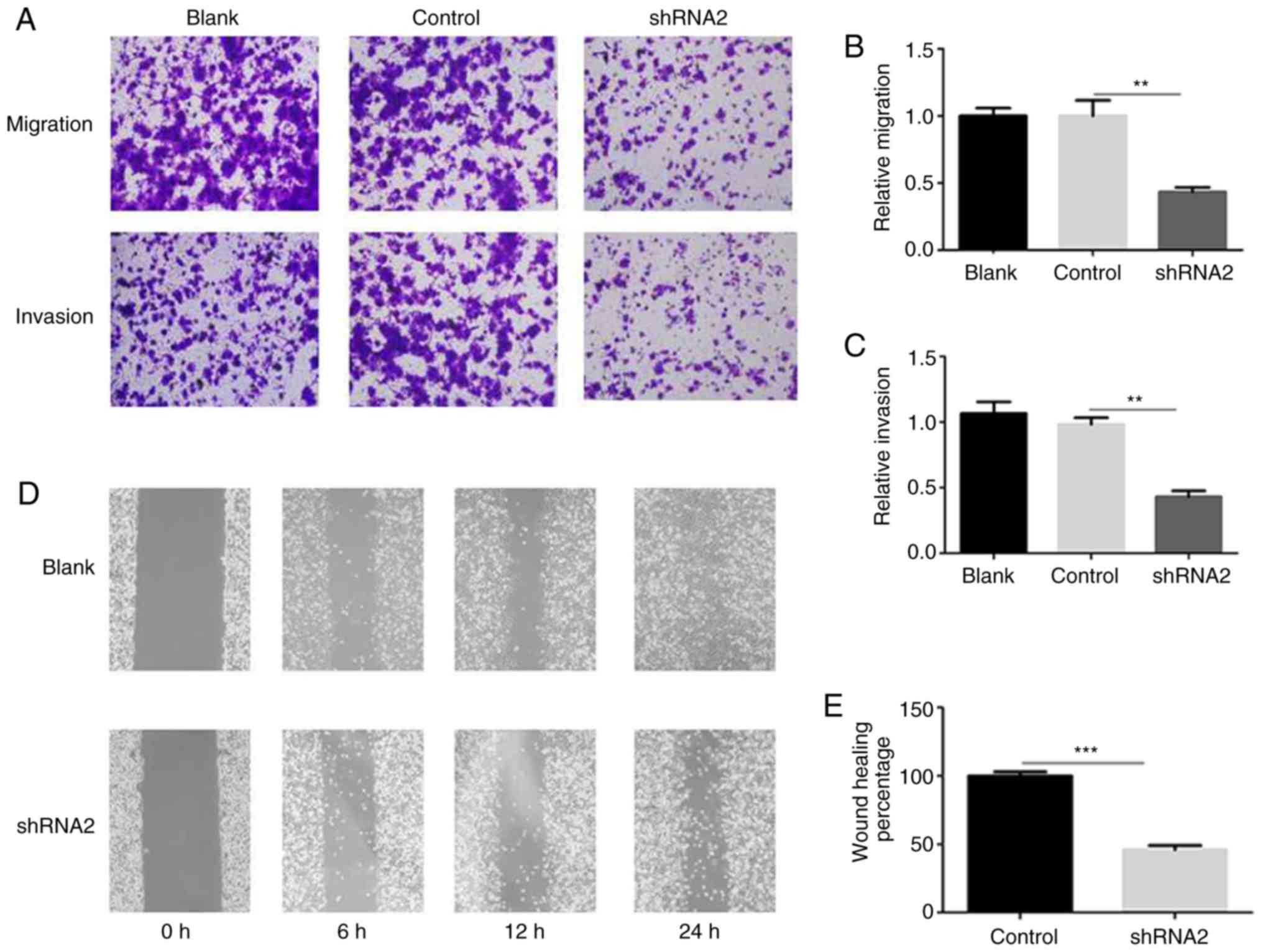
Transwell assays were performed to evaluate the
effect of linc01105 knockdown on the migration and invasion of NB
cells. The results demonstrated that silencing linc01105
significantly inhibited NB cell migration (Fig. 4A and B) and invasion (Fig. 4A and C), compared with the control
group. Furthermore, analysis of the wound healing assay indicated
that knockdown of linc01105 significantly suppressed NB cell
migration (Fig. 4D and E).
Linc01105 regulates the expression of
miR-6769b-5p targeting VEGFA in NB cells
To explore the mechanism of linc01105 in NB
progression, a potential link with VEGFA, a key angiogenic factor
involved in NB angiogenesis, was explored. DIANA-LncBase and CirNet
database analyses were used to search for potential miRNAs binding
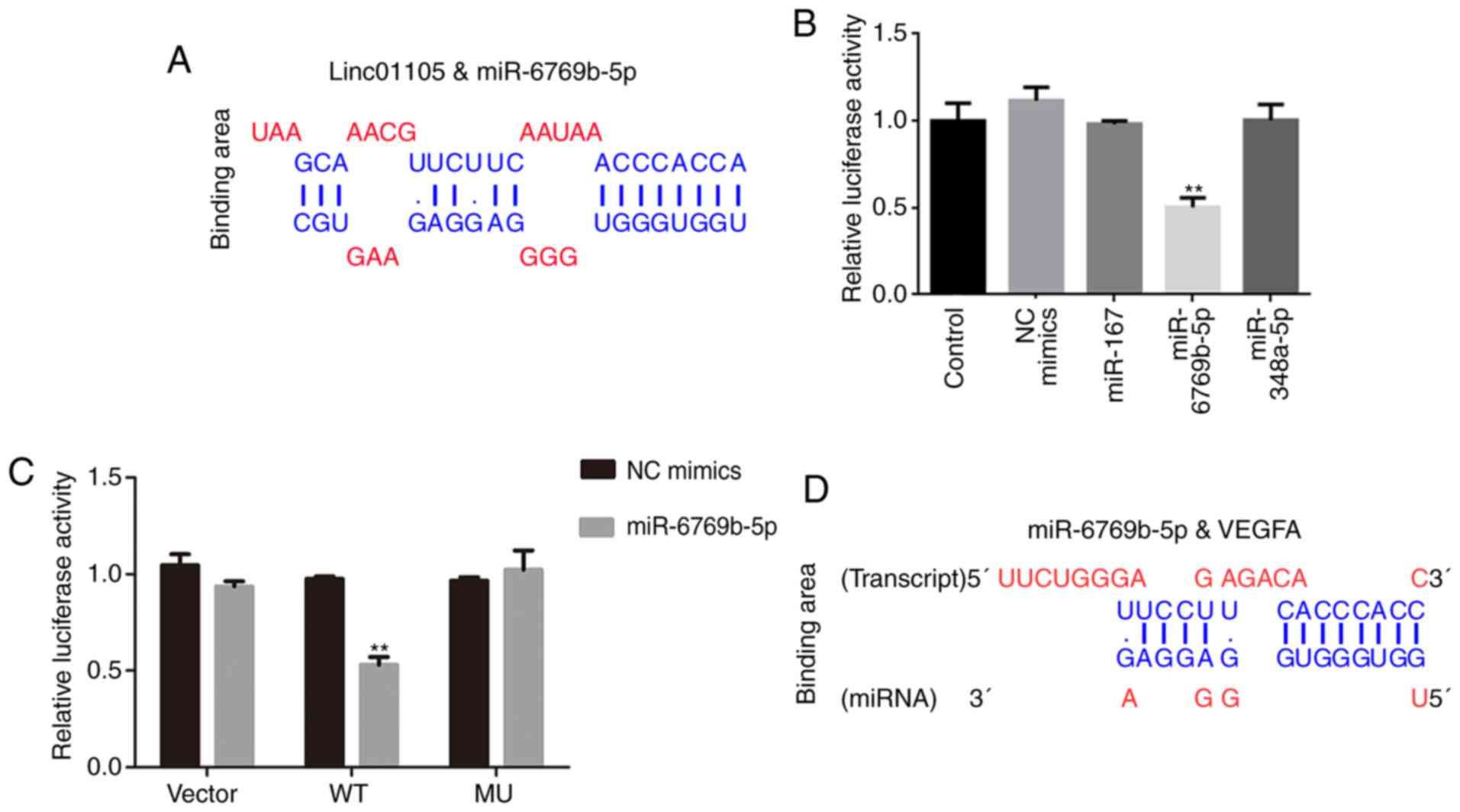
with both linc01105 and VEGFA (Fig. 5A
and D). miR-107, miR-6769b-5p and miR-348a-3p were identified
as potential miRNAs involved in this process. To examine this, the
cDNA of linc01105 was cloned into a luciferase reporter plasmid
(RLuc-Linc01105-WT). Subsequently, the luciferase reporter plasmid
and different miRNA mimics were transfected into 293T cells.
Luciferase activity was significantly decreased by miR-6769b-5p
mimics transfection (Fig. 5B), but
not by the other miRNA mimics tested. To avoid unspecific binding,
the miR-6769b-5p binding site of linc01105 and VEGFA was mutated
from ACCCACC to TGGGTGG. Transfection of miR-6769b-5p mimics
significantly inhibited RLuc-Linc01105-WT activity, but had no
effect on RLuc-Linc01105-Mut activity (Fig. 5C). Similar results were observed for
the RLuc-VEGFA-WT and RLuc-VEGFA-MU activities (Fig. 5E). Next, the miRNA-6769b-5p expression
levels were detected in NB tissues and adjacent normal tissues
(Fig. 5F). To further confirm that
VEGFA was a target gene of miRNA-6769b-5p, miRNA-6769b-5p mimics
and inhibitor were transfected into SH-SY5Y cells (Fig. 5G). The results revealed that
miRNA-6769b-5p mimics decreased the expression of VEGFA, while
miRNA-6769b-5p inhibitor transfection increased VEGFA expression
(Fig. 5H).
HIF-1α activates the expression of
linc01105 by acting as a transcription factor
HIF-1α was predicted to bind with the linc01105
promoter region through analysis with the ALGGEN (http://alggen.lsi.upc.es) and JASPAR (http://jaspar.genereg.net) databases. Therefore, a
HIF-1α-expressing plasmid was transfected in SH-SY5Y cells
(Fig. 6A). The results demonstrated
that overexpression of HIF-1α upregulated linc01105 levels
(Fig. 6B). VEGFA expression levels
were also upregulated following HIF-1α overexpression (Fig. 6C). Subsequently, the linc01105
promoter (2 kb upstream the transcript start site) was cloned into
a luciferase gene reporter (RLuc-Linc01105-promoter-WT) and
transfected into 293T cells. Luciferase activity was significantly
increased following HIF-1α overexpression (Fig. 6D), confirming direct binding of HIF-1α
with the promoter of linc01105. Finally, a CHIP-PCR assay also
indicated that HIF-1α may bind directly with the promoter of
linc01105 (Fig. 6E) and promote
linc01105 transcription.
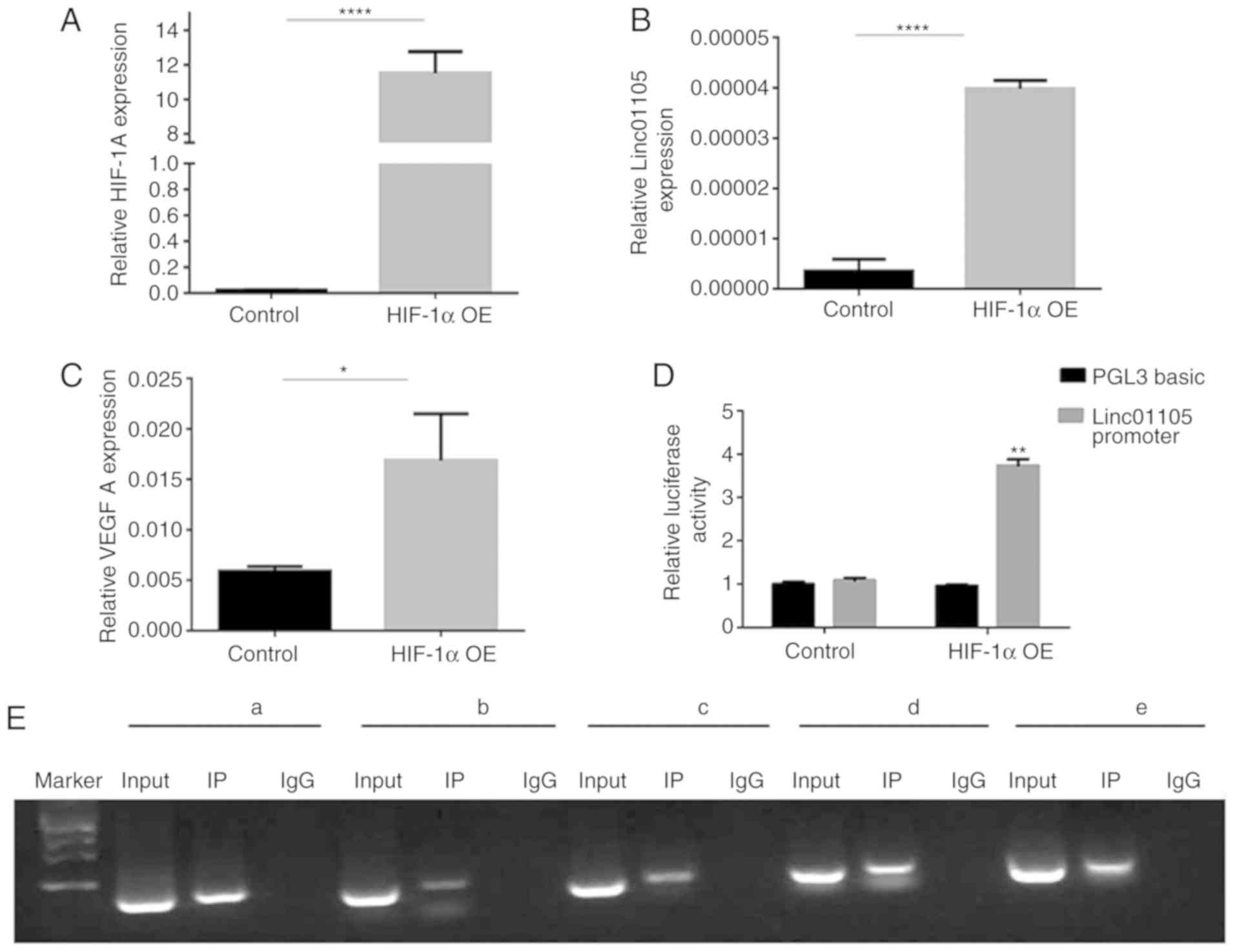 | Figure 6.HIF-1α activates the transcription of
linc01105. (A) Validation of HIF-1α overexpression in SH-SY5Y cells
by transient transfection with a HIF-1α-expressing plasmid. (B)
Linc01105 expression levels and (C) VEGFA mRNA expression levels
were upregulated following HIF-1α overexpression in SH-SY5Y cells.
(D) The relative luciferase activities following co-transfection
with a linc01105 promoter reporter plasmid and the HIF1α
overexpression plasmid. (E) Chromatin immunoprecipitation assay for
HIF-1α and the promoter region of Linc01105. The promoter region
fragments are shown as follows: A, 1–400 bp; b, 400–800 bp; c,
800–1,200 bp; d, 1,200-1,600 bp; e, 1,600-2,000 bp. *P<0.05,
**P<0.01 and ****P<0.0001. HIF-1α, hypoxia inducible
factor-1α; OE, overexpression; IP, immunoprecipitation; IgG,
immunoglobulin G. |
Linc01105 and miR-6769b-5p may
participate in angiogenesis
HUVECs were used for tube-formation assays, in order
to evaluate the angiogenesis potential. HUVECs were cultured with
HUVEC-specific medium or the CM of SH-SY5Y cells. Silencing of
linc01105 was demonstrated to inhibit endothelial cell tube
formation (Fig. 7A and B). The tube
formation ability of the CM was reduced by pretreatment of CM with
VEGFA antibody, by VEGFA siRNA transfection in the SH-SY5Y cells,
and by miR-6769b-5p overexpression by mimics transfection in the
SH-SY5Y cells (Fig. 7C and D). The
tube formation ability reduced by miR-6769b-5p overexpression was
restored by VEGFA overexpression (Fig. 7C
and D). Finally, silencing and overexpression of VEGFA were
confirmed by western blotting (Fig.
7E).
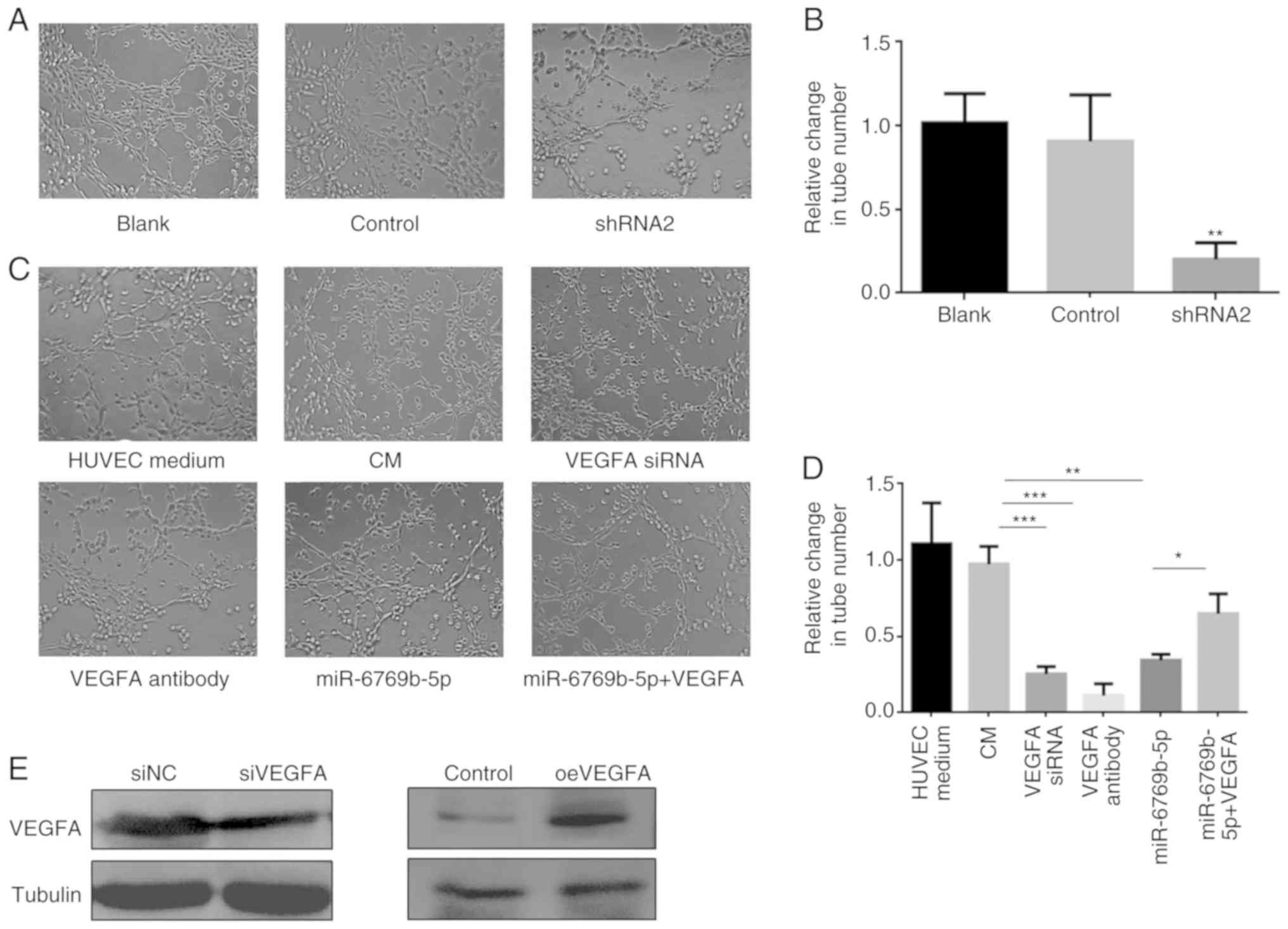 | Figure 7.Effects of linc01105, miR-6769b-5p
and VEGFA on angiogenesis. (A) HUVECs were cultured in CM from
SH-SY5Y cells that were either untreated (blank), infected with
control virus (control) or infected with linc01105-shRNA virus
(shRNA2). (B) Quantification of tube formation ability from panel
A. **P<0.01 compared with control. (C) HUVECs were cultured as
follows: With HUVEC-specific media or with SH-SY5Y CM as positive
control; with CM pre-treated with VEGFA antibody; with CM of
SH-SY5Y cells transfected with a VEGFA-targeting siRNA; with CM of
SH-SY5Y cells transfected with miR-6769b-5p mimics; and with CM of
SH-SY5Y cells transfected with miR-6769b-5p mimics and
VEGFA-overexpressing plasmid (n=3). Tube formation was observed
after 6 h of culture using phase-contrast light microscopy. (D)
Quantification of tube formation abilities from panel C.
*P<0.05, **P<0.01 and ***P<0.005, with comparisons
indicated by lines. (E) Validation of VEGFA knockdown and
overexpression by siRNA and plasmid transfection, respectively.
VEGFA, vascular endothelial growth factor A; HUVECs, human
umbilical vein endothelial cells; CM, conditioned media; siRNA,
small interfering RNA; shRNA, short hairpin RNA; NC, negative
control; OE, overexpression. |
Linc01105 gene targets
To explore more functions of linc01105, a CHIRP
assay was performed to identify gene locations that are directly
bound by linc01105. A protein interaction network was constructed
using STRING for the linc01105-specific binding gene targets (after
exclusion of targets identified by the negative control), revealing
significant interactions between 20 genes (Fig. S1 and Table
SI); among these genes were numerous ribosomal proteins and
heat shock protein 90, which have been previously reported to be
associated to tumorigenesis (27,28).
Indeed, GO analysis demonstrated that linc01105 may impact a
variety of biological processes. However, no KEGG pathway was found
to be enriched, most likely due to low number of the 20 genes used
as input in the analysis.
Discussion
Advanced NB patients are typically associated with a
poor prognosis and frequent relapses despite treatment with a
variety of therapies (29,30). Therefore, it is extremely important to
identify novel biomarkers in order to improve the prognostic
outcome of pediatric patients with NB (31). With the development of genomics
sequencing technologies, recent research has focused on lncRNAs
(32,33). Increasing evidence suggests that
lncRNAs participate in a number of biological processes and have
important roles in human diseases, such as cancer (34,35).
Our previous study found that linc01105 was
differentially expressed in NB and adjacent normal tissues through
genome-wide lncRNA analysis (14). In
addition, it was demonstrated that upregulation of linc01105 was
correlated with NB INSS stage. In that previous study, the
SK-N-BE(2)C cell line was used, which is a MYCN-amplification cell
line; in the present study, the cell line SH-SY5Y was used, which
is a non-MYCN amplification cell line, to further investigate the
mechanism of linc01105 in NB. The present study focused on the
molecular mechanism of linc01105. To explore the functions of
linc01105, a nuclear-cytoplasmic fractionation was performed and
the results revealed that linc01105 was mainly expressed in the
cytoplasmic fraction. Subsequently, a loss-of-function assay was
performed by linc01105 shRNA knockdown. Knockdown of linc01105
inhibited cell proliferation and promoted cell apoptosis. Notably,
knockdown of linc01105 resulted in inhibition of migration and
invasion. Furthermore, linc01105 silencing altered the expression
of Bcl-2 family proteins and activated the p53/Caspase signaling
pathway. These results indicated that linc01105 affected the
apoptosis process via regulating Bcl-2 proteins and activating the
p53/caspase signaling pathway.
One of the significant functions of lncRNAs,
especially those located in the cell cytoplasm, is to bind with
miRNAs, subsequently acting as a ‘sponge’ and inhibiting their
expression (36–38). For example, in gastric cancer,
linc01234 may have a role as a competing endogenous RNA to regulate
core-binding factor subunit β expression by sponging miR-204-5p
(36). Another study demonstrated
that miR-29b-3p was directly inhibited by linc00511, which resulted
in an increase in VEGFA expression in pancreatic ductal
adenocarcinoma (39). Angiogenesis is
crucial for tumor growth and is associated with tumor metastasis;
notably, VEGFA is a major regulator of NB angiogenesis (40,41).
Bioinformatics analysis was performed to identify
miRNAs that may bind with both linc01105 and VEGFA. The results
revealed that miR-6769b-5p shared complementary binding sites with
the VEGFA 3′UTR region and with linc01105 (924–948 nt region),
which was confirmed by luciferase assay. RT-qPCR results indicated
that miR-6769b-5p expression levels were lower in NB tissues
compared with adjacent normal tissues. In addition, transfection
with miR-6769b-5p mimics resulted in downregulation of VEGFA, while
miR-6769b-5p inhibition resulted in upregulation of VEGFA
expression. These findings suggested that linc01105, miR-6769b-5p
and VEGFA mRNA constituted a competing endogenous RNA regulatory
network. Silencing of linc01105 reduced the miR-6769b-5p competing
adsorption and increased the levels of free miR-6769b-5p, thereby
promoting the degradation of VEGFA by increasing binding to the
3′UTR region of VEGFA. Indeed, transfection with a miR-6769b-5p
inhibitor suppressed VEGFA expression. By contrast, upregulation of
linc01105 contributed to VEGFA upregulation via the competing
endogenous RNA network. Of note, VEGFA knockdown had no influence
on apoptosis but inhibited tumor migration (42,43).
Therefore, the present study suggested that miR-6769b-5p may
participate in NB tumorigenesis via interaction with VEGFA. The
VEGF-mediated mechanism most likely has no effect on the
p53/caspase signaling pathway, and the VEGFA network may only
affect the migration phenotype of the linc01105 knockdown and not
apoptosis.
The present study used the ALGGEN and JASPAR
databases to predict transcription factors which may bind with the
promoter of linc01105; among the predicted factors, HIF-1α was of
particular interest, as high expression of HIF-1α has been widely
reported to be associated with poor prognosis of NB (44,45).
Over-expression of HIF-1α significantly promoted linc01105
expression levels. Furthermore, a luciferase assay confirmed that
HIF-1α regulated the expression of linc01105 acting as a
transcription factor and directly binding to its promoter.
Additionally, the CHIRP assay identified 20 genes that directly
bind with Linc01105, and which are potential candidates for future
studies.
In conclusion, the present study demonstrated that
linc01105 was significantly upregulated in NB tissues compared with
normal tissues. Silencing of linc01105 resulted in the activation
of the p53/caspase signaling pathway and the inhibition of NB cell
proliferation, migration and invasion. In addition, silencing of
linc01105 suppressed angiogenesis via miR-6769b-5p targeting of
VEGFA. Taken together, these results suggested that the
linc01105/p53/caspase pathway or the linc01105/miR-6769b-5p/VEGFA
network may serve as candidate targets for future therapies for
NB.
Supplementary Material
Supporting Data
Acknowledgements
The authors wish to thank Ms Xia Wenjun and Mr Huang
Jianbo (Fudan University, Shanghai, China), who provided kind
assistance with all of the experiments.
Funding
This study received financial support from Shanghai
Key Disciplines (grant no. 2017ZZ02022), National Natural Science
Foundation of China (grant nos. 81771633 and 81572324) and Science
Foundation of Shanghai (grant nos. 17411960600 and
15ZR1404200).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
DK and YM designed the study. YM and MJ collected
the data and performed experiments. LB and LX analyzed and
interpreted the data. MD and DK were involved in critical reviewing
of the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
This study was approved by the Institute Research
Ethics Committee at the Children's Hospital of Fudan University.
Informed consent was acquired from every patient's legal
guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mondal T, Juvvuna PK, Kirkeby A, Mitra S,
Kosalai ST, Traxler L, Hertwig F, Wernig-Zorc S, Miranda C, Deland
L, et al: Sense-antisense lncRNA pair encoded by locus 6p22.3
determines neuroblastoma susceptibility via the USP36-CHD7-SOX9
regulatory axis. Cancer Cell. 33:417–434 e7. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Duan C, Wang H, Chen Y, Chu P, Xing T, Gao
C, Yue Z, Zheng J, Jin M, Gu W and Ma X: Whole exome sequencing
reveals novel somatic alterations in neuroblastoma patients with
chemotherapy. Cancer Cell Int. 18:212018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li Z, Takenobu H, Setyawati AN, Akita N,
Haruta M, Satoh S, Shinno Y, Chikaraishi K, Mukae K, Akter J, et
al: EZH2 regulates neuroblastoma cell differentiation via NTRK1
promoter epigenetic modifications. Oncogene. 37:2714–2727. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Roy J and Mallick B: Investigating
piwi-interacting RNA regulome in human Neuroblastoma. Genes
Chromosomes Cancer. 57:339–349. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Singh N, Liu X, Hulitt J, Jiang S, June
CH, Grupp SA, Barrett DM and Zhao Y: Nature of tumor control by
permanently and transiently modified GD2 chimeric antigen receptor
T cells in xenograft models of neuroblastoma. Cancer Immunol Res.
2:1059–1070. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang F, Yang H, Deng Z, Su Y, Fang Q and
Yin Z: HOX antisense lincRNA HOXA-AS2 promotes tumorigenesis of
hepatocellular carcinoma. Cell Physiol Biochem. 40:287–296. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cheetham SW, Gruhl F, Mattick JS and
Dinger ME: Long noncoding RNAs and the genetics of cancer. Brit J
Cancer. 108:2419–2425. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ding J, Xie M, Lian Y, Zhu Y, Peng P, Wang
J, Wang L and Wang K: Long noncoding RNA HOXA-AS2 represses P21 and
KLF2 expression transcription by binding with EZH2, LSD1 in
colorectal cancer. Oncogenesis. 6:e2882017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zheng H, Yang S, Yang Y, Yuan SX, Wu FQ,
Wang LL, Yan HL, Sun SH and Zhou WP: Epigenetically silenced long
noncoding-SRHC promotes proliferation of hepatocellular carcinoma.
J Cancer Res Clin Oncol. 141:1195–1203. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dong R, Liu GB, Liu BH, Chen G, Li K,
Zheng S and Dong KR: Targeting long non-coding RNA-TUG1 inhibits
tumor growth and angiogenesis in hepatoblastoma. Cell Death Dis.
7:e22782016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu L, Murat P, Matak-Vinkovic D, Murrell A
and Balasubramanian S: Binding interactions between long noncoding
RNA HOTAIR and PRC2 proteins. Biochemistry. 52:9519–9527. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tsai KW, Lo YH, Yeh CY, Chen YZ, Hsu CW,
Chen WS and Wang JH: Linc00659, a long noncoding RNA, acts as novel
oncogene in regulating cancer cell growth in colorectal cancer. Mol
Cancer. 17:722018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nallasamy P, Chava S, Verma SS, Mishra S,
Gorantla S, Coulter DW, Byrareddy SN, Batra SK, Gupta SC and
Challagundla KB: PD-L1, inflammation, non-coding RNAs, and
neuroblastoma: Immuno-oncology perspective. Semin Cancer Biol.
52:53–65. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tang W, Dong K, Li K, Dong R and Zheng S:
MEG3, HCN3 and linc01105 influence the proliferation and apoptosis
of neuroblastoma cells via the HIF-1α and p53 pathways. Sci Rep.
6:362682016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Russell MR, Penikis A, Oldridge DA,
Alvarez-Dominguez JR, McDaniel L, Diamond M, Padovan O, Raman P, Li
Y, Wei JS, et al: CASC15-S is a tumor suppressor lncRNA at the 6p22
neuroblastoma susceptibility locus. Cancer Res. 75:3155–3166. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu PY, Erriquez D, Marshall GM, Tee AE,
Polly P, Wong M, Liu B, Bell JL, Zhang XD, Milazzo G, et al:
Effects of a novel long noncoding RNA, lncUSMycN, on N-Myc
expression and neuroblastoma progression. J Natl Cancer Inst.
106(pii): dju1132014.PubMed/NCBI
|
|
17
|
Li D, Wang X, Mei H, Fang E, Ye L, Song H,
Yang F, Li H, Huang K, Zheng L and Tong Q: Long noncoding RNA
pancEts-1 promotes neuroblastoma progression through
hnRNPK-mediated β-catenin stabilization. Cancer Res. 78:1169–1183.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liang L, Xu J, Wang M, Xu G, Zhang N, Wang
G and Zhao Y: LncRNA HCP5 promotes follicular thyroid carcinoma
progression via miRNAs sponge. Cell Death Dis. 9:3722018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang ZK, Li J, Guan D, Liang C, Zhuo Z,
Liu J, Lu A, Zhang G and Zhang BT: A newly identified lncRNA MAR1
acts as a miR-487b sponge to promote skeletal muscle
differentiation and regeneration. J Cachexia Sarcopenia Muscle.
9:613–626. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhong J, Wang H, Chen W, Sun Z, Chen J, Xu
Y, Weng M, Shi Q, Ma D and Miao C: Ubiquitylation of MFHAS1 by the
ubiquitin ligase praja2 promotes M1 macrophage polarization by
activating JNK and p38 pathways. Cell Death Dis. 8:e27632017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Morelli C, Garofalo C, Sisci D, del Rincon
S, Cascio S, Tu X, Vecchione A, Sauter ER, Miller WH Jr and Surmacz
E: Nuclear insulin receptor substrate 1 interacts with estrogen
receptor alpha at ERE promoters. Oncogene. 23:7517–7526. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chu C, Quinn J and Chang HY: Chromatin
isolation by RNA purification (ChIRP). J Vis Exp. 25(pii):
39122012.
|
|
24
|
Gowda Saralamma VV, Lee HJ, Raha S, Lee
WS, Kim EH, Lee SJ, Heo JD, Won C, Kang CK and Kim GS: Inhibition
of IAP's and activation of p53 leads to caspase-dependent apoptosis
in gastric cancer cells treated with Scutellarein. Oncotarget.
9:5993–6006. 2017.PubMed/NCBI
|
|
25
|
Ehrnhoefer DE, Skotte NH, Ladha S, Nguyen
YT, Qiu X, Deng Y, Huynh KT, Engemann S, Nielsen SM, Becanovic K,
et al: p53 increases caspase-6 expression and activation in muscle
tissue expressing mutant huntingtin. Hum Mol Genet. 23:717–729.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guo XX, Li Y, Sun C, Jiang D, Lin YJ, Jin
FX, Lee SK and Jin YH: p53-dependent fas expression is critical for
ginsenoside Rh2 triggered caspase-8 activation in HeLa cells.
Protein Cell. 5:224–234. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rong B and Yang S: Molecular mechanism and
targeted therapy of Hsp90 involved in lung cancer: New discoveries
and developments (Review). Int J Oncol. 52:321–336. 2018.PubMed/NCBI
|
|
28
|
Gao C, Peng YN, Wang HZ, Fang SL, Zhang M,
Zhao Q and Liu J: Inhibition of heat shock protein 90 as a novel
platform for the treatment of cancer. Curr Pharm Des. 25:849–855.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen Q, Deng R, Zhao X, Yuan H, Zhang H,
Dou J, Chen R, Jin H, Wang Y, Huang J and Yu J: Sumoylation of
EphB1 suppresses neuroblastoma tumorigenesis via inhibiting PKCγ
activation. Cell Physiol Biochem. 45:2283–2292. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Batzke K, Büchel G, Hansen W and Schramm
A: TrkB-target galectin-1 impairs immune activation and radiation
responses in neuroblastoma: Implications for tumour therapy. Int J
Mol Sci. 19(pii): E7182018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Szemes M, Greenhough A, Melegh Z, Malik S,
Yuksel A, Catchpoole D, Gallacher K, Kollareddy M, Park JH and
Malik K: Wnt signalling drives context-dependent differentiation or
proliferation in neuroblastoma. Neoplasia. 20:335–350. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang S, Xu J and Zeng X: A six-long
non-coding RNA signature predicts prognosis in melanoma patients.
Int J Oncol. 52:1178–1188. 2018.PubMed/NCBI
|
|
33
|
Yamada A, Yu P, Lin W, Okugawa Y, Boland
CR and Goel A: A RNA-Sequencing approach for the identification of
novel long non-coding RNA biomarkers in colorectal cancer. Sci Rep.
8:5752018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang T, Wang M, Huang B, Chang A, Liu F,
Zhang Y and Jiang B: Long noncoding RNAs in the mTOR signaling
network: Biomarkers and therapeutic targets. Apoptosis. 23:255–264.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Janakiraman H, House RP, Gangaraju VK,
Diehl JA, Howe PH and Palanisamy V: The long (lncRNA) and short
(miRNA) of It: TGFβ-mediated control of RNA-binding proteins and
noncoding RNAs. Mol Cancer Res. 16:567–579. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen X, Chen Z, Yu S, Nie F, Yan S, Ma P,
Chen Q, Wei C, Fu H, Xu T, et al: Long noncoding RNA LINC01234
functions as a competing endogenous RNA to regulate CBFB expression
by sponging miR-204-5p in gastric cancer. Clin Cancer Res.
15:2002–2014. 2018. View Article : Google Scholar
|
|
37
|
Liu XH, Sun M, Nie FQ, Ge YB, Zhang EB,
Yin DD, Kong R, Xia R, Lu KH, Li JH, et al: Lnc RNA HOTAIR
functions as a competing endogenous RNA to regulate HER2 expression
by sponging miR-331-3p in gastric cancer. Mol Cancer. 13:922014.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xie CR, Wang F, Zhang S, Wang FQ, Zheng S,
Li Z, Lv J, Qi HQ, Fang QL, Wang XM and Yin ZY: Long noncoding RNA
HCAL facilitates the growth and metastasis of hepatocellular
carcinoma by acting as a ceRNA of LAPTM4B. Mol Ther Nucleic Acids.
9:440–451. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhao X, Liu Y, Li Z, Zheng S, Wang Z, Li
W, Bi Z, Li L, Jiang Y, Luo Y, et al: Linc00511 acts as a competing
endogenous RNA to regulate VEGFA expression through sponging
hsa-miR-29b-3p in pancreatic ductal adenocarcinoma. J Cell Mol Med.
22:655–667. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Weng WC, Lin KH, Wu PY, Ho YH, Liu YL,
Wang BJ, Chen CC, Lin YC, Liao YF, Lee WT, et al: VEGF expression
correlates with neuronal differentiation and predicts a favorable
prognosis in patients with neuroblastoma. Sci Rep. 7:112122017.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fakhari M, Pullirsch D, Paya K, Abraham D,
Hofbauer R and Aharinejad S: Upregulation of vascular endothelial
growth factor receptors is associated with advanced neuroblastoma.
J Pediatr Surg. 37:582–587. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Weng WC, Lin KH, Wu PY, Lu YC, Weng YC,
Wang BJ, Liao YF, Hsu WM, Lee WT and Lee H: Calreticulin regulates
VEGF-A in neuroblastoma cells. Mol Neurobiol. 52:758–770. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kaneko S, Ishibashi M and Kaneko M:
Vascular endothelial growth factor expression is closely related to
irinotecan-mediated inhibition of tumor growth and angiogenesis in
neuroblastoma xenografts. Cancer Sci. 99:1209–1217. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yin CP, Guan SH, Zhang B, Wang XX and Yue
SW: Upregulation of HIF-1α protects neuroblastoma cells from
hypoxia-induced apoptosis in a RhoA-dependent manner. Mol Med Rep.
12:7123–7131. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Westerlund I, Shi Y, Toskas K, Fell SM, Li
S, Surova O, Södersten E, Kogner P, Nyman U, Schlisio S and
Holmberg J: Combined epigenetic and differentiation-based treatment
inhibits neuroblastoma tumor growth and links HIF2α to tumor
suppression. Proc Natl Acad Sci USA. 114:E6137–E6146. 2017.
View Article : Google Scholar : PubMed/NCBI
|