Introduction
Breast cancer is the most commonly diagnosed
malignant tumor in women worldwide. Despite the numerous and
advanced types of combined therapy, breast cancer remains among the
main causes of tumor-related deaths in women (1). Breast cancer is associated with
aggressive local invasion, metastasis in advanced stage and
multidrug resistance to chemotherapy (2). Various molecular markers have been
identified that contribute to the regulation of breast cancer
invasion and metastasis, such as cell cycle regulators and
cell-adhesion proteins (3).
The binding of chemokines to specific
G-protein-coupled receptors induce cytoskeletal rearrangement and
directional migration and then critically regulate tumor cell
migration and metastasis (4).
Chemokines were initially characterized due to their association
with inflammatory responses. These are members of a superfamily of
chemotactic cytokines (5).
Chemokine (C-C motif) ligand 18 (CCL18) is a type of chemokine,
which is predominantly derived from breast tumor-associated
macrophages (TAMs) (6). Previous
data indicate that the binding of CCL18 to its receptor, PYK2
N-terminal domain interacting receptor 1 (Nir1), promotes the
invasion, metastasis and EMT of breast cancer cells through the
PI3K/Akt/GSK3β/Snail signaling pathway (3,7).
However, the involvement of other factors in the CCL18-induced
migration, invasion and metastasis of breast cancer cells remains
unclear.
AnnexinA2 (AnxA2), a member of the calcium-dependent
phospholipid binding proteins, plays a crucial role in cancer
progression (8,9). AnxA2 is reported to be overexpressed
in a variety of cancers and contributes to invasion (10–12),
metastasis (11,13), angiogenesis (14), proliferation (13,15–17),
F-actin polymerization (16),
multidrug resistance to chemotherapy (18–20)
and epithelial-mesenchymal transition (EMT) (9).
EMT is crucial in embryonic development and cancer
progression and it is a process during which epithelial-like cancer
cells transform into mesenchymal-like tumor cells with upregulation
of mesenchymal markers and downregulation of epithelial markers.
Moreover, EMT is a vital contributor to the invasion and metastasis
of breast cancer cells (21,22).
In the present study, we detected the association of AnxA2
expression in breast tissue specimens with the clinicopathological
characteristics of patients and observed the role of AnxA2
expression in breast cancer migration and invasion under the
induction of rCCL18 to verify the hypothesis that AnxA2 promotes
CCL18-induced progression and metastasis of breast cancer cells. In
our reseach, AnxA2 was found to contribute to CCL18-induced
phosphorylation of integrin β1 in breast cancer cells, which is a
crucial step in cytoskeletal rearrangement and cell adhesion.
We hypothesized that the binding of CCL18 to Nir1
promotes tumor progression and metastasis by inducing EMT via the
PI3K/Akt/GSK3β/Snail signaling pathway though AnxA2 in vitro
and in vivo.
Materials and methods
Patients and tissue specimens
All tissue specimens were obtained from the
Department of Pathology, Affiliated Hospital of Weifang Medical
University from 1 January 2012 to 31 December 2016. These tissue
specimens consisted of samples from 120 cases of invasive ductal
carcinoma and their corresponding adjacent non-tumor tissues. The
distance between invasive ductal carcinoma and their corresponding
adjacent non-tumor tissues was >5 cm. The mean age of the
patients was 47.3 years (range, 35–76). All tissue specimens were
fixed, embedded in paraffin, serial sectioned and H&E stained
and finally stored at room temperature. The inclusion criteria for
the patients was a diagnosis of breast cancer and agreement to
participate in the present study. Patient samples were collected
according to a protocol approved by the Institutional Review Board,
and patients provided signed consent for use of their tissue
specimens in the present study. The study protocol was reviewed and
approved by the Weifang Medical University Ethics Committee
(approval no. 045, 26-Feb-2016). Clinical information of the
samples is documented in detail in Table I.
 | Table I.Association between AnxA2 expression
and clinical features of the invasive ductal carcinoma patients
enrolled in this study. |
Table I.
Association between AnxA2 expression
and clinical features of the invasive ductal carcinoma patients
enrolled in this study.
|
| AnxA2
expression |
|
|---|
|
|
|
|
|---|
| Variables | High | Low | P-value |
|---|
| Age (years) |
|
|
|
|
≤50 | 34 | 19 | 0.190 |
|
≥51 | 35 | 32 |
|
| Tumor size |
|
|
|
| ≤5
cm | 31 | 23 | 0.723 |
| >5
cm | 40 | 26 |
|
| Tumor
differentiation |
|
|
|
| I | 17 | 21 | 0.036 |
| II | 25 | 16 |
|
|
III | 30 | 11 |
|
| Lymph node
metastasis |
|
|
|
|
Yes | 48 | 21 | 0.007 |
| No | 23 | 28 |
|
| Distant
metastasis |
|
|
|
|
Yes | 46 | 20 | 0.009 |
| No | 25 | 29 |
|
| Nir1 |
|
|
|
|
Positive | 55 | 24 | 0.006 |
|
Negative | 18 | 23 |
|
Cell lines and reagents
The human breast cancer cell lines, MDA-MB-231, T47D
and MCF-7, were obtained from the American Type Culture Collection
(ATCC). Lipofectamine 2000 was obtained from Invitrogen; Thermo
Fisher Scientific, Inc. Plasmid construction was carried out by
Genescript (USA). The Pierce™ BCA Protein Assay Kit and the Pierce™
ECL kit were purchased from Thermo Fisher Scientific, Inc. Primary
antibodies including anti-AnxA2 (dilution 1:1,000; cat. no.
ab178677; Abcam), anti-phospho-AnxA2 (1:1,000; cat. no. sc-135753;
Santa Cruz Biotechnology), anti-Akt (dilution 1:1,000; cat. no.
2920; Cell Signaling Technology), anti-phospho-Akt-Ser473 (dilution
1:1,000; cat. no. 15116; Cell Signaling Technology),
anti-phospho-Akt-Thr308 (dilution 1:1,000; cat. no. 13038; Cell
Signaling Technology), anti-integrin β1 (dilution 1:1,000; cat. no.
34971; Cell Signaling Technology), anti-phospho-integrin β1
(dilution 1:1,000; cat. no. 44-873G; Thermo Fisher Scientific,
Inc.), anti-GSK3β (dilution 1:1,000; cat. no. 12456; Cell Signaling
Technology), anti-phospho-GSK3β (dilution 1:1,000; cat. no. 9323;
Cell Signaling Technology), anti-E-cadherin (dilution 1:1,000; cat.
no. 7687; Cell Signaling Technology), anti-N-cadherin (dilution
1:1,000; cat. no. 13116; Cell Signaling Technology), anti-vimentin
(dilution 1:1,000; cat. no. 3390; Cell Signaling Technology),
anti-Snail (1:1,000; cat. no. 3895; Cell Signaling Technology),
β-actin (dilution 1:1,000; cat. no. 3700; Santa Cruz
Biotechnology), anti-Nucleolin (dilution 1:1,000; cat. no. ab22758;
Abcam) and the anti-mouse, rabbit and goat secondary antibodies
were obtained from Santa Cruz Biotechnology, Inc.
Fluorochrome-conjugated secondary antibodies, Alexa Flour 594 and
TRITC, (Invitrogen; Thermo Fisher Scientific, Inc.) were diluted
1:600 in blocking buffer (Solarbio). All culture media and related
reagents were purchased from Hyclone/GE Healthcare, USA.
Animals
Forty 1-week-old female SCID mice were purchased
from Wei Tong Li Hua Experimental Animal Co. (initial mean weight
18.3 g; range 16.3–19.7 g). The mice were maintained in laminar
flow rooms under constant temperature and humidity. The food and
water of mice were sterilized, and the food and water were renewed
ad libitum.
Plasmid construction, shRNA and
plasmid transfection
Cells were cultured in a 6-well plate for 24 h in
complete medium before transfection. Transfection was performed
with Lipofectamine 2000 according to the manufacturer's
instructions. Sequences of AnxA2 siRNA were
5′-GGTCTGAATTCAAGAGAAA-3′ and 5′-GCCAAAGAAATGAACATTC-3′. Sequences
of Nir 1 siRNA were 5′-GGGAGAAGUGGCUUCGUAATT-3′ and
5′-UUACGAAGCCACUUCUCCCGG-3′. Sequences of Akt siRNA were
5′-GCUGGCUCCACAAACGUGGUGAAUA-3′ and
5′-UAUUCACCACGUUUGUGGAGCCAGC-3′. Culture medium with 600 ng/ml
hygromycin B was used to select stably transfected cells. The
MDA-MB-231 cells were stably transfected with the sequences of
AnxA2 siRNA, Nir 1 siRNA or Akt siRNA respectively marked as
SiAnxA2/MDA-231, SiNir1/MDA-231 and SiAkt/MDA-231. A scrambled
SiRNA was used as a control (Scr/MDA-231). Surviving cells were
evaluated for AnxA2 expression by western blot analysis. For
plasmid transfection, cells were plated in a 6-well plate for 24 h
before transfection. AnxA2 cDNA was cloned in the
KpnI-XbaI sites of pcDNA3.1, and was confirmed by DNA
sequencing. MCF-7 cells were transfected with the pcDNA3.1-AnxA2
plasmid, pcDNA3.1-Nir1 plasmid, pcDNA3.1-Akt plasmid or pcDNA3.1
vector using Lipofectamine 2000 and marked as AnxA2/MCF-7,
NIR1/MCF-7, Akt/MCF-7 or Con/MCF-7. Stable transfected cells were
selected and maintained in culture medium with G418.
Western blot analysis
Total cellular proteins were extracted with ice-cold
RIPA cell lysis buffer with Protease Inhibitor Cocktail. We
employed the BCA Protein Assay Kit (Pierce) to quantify the protein
concentration. The amount of protein loaded in each lane is 10 µl.
Lysates were separated by 10% SDS-PAGE and transferred onto a PVDF
membrane. The membrane was incubated in primary antibodies,
horseradish HRP-conjugated anti-rabbit IgG secondary antibody
successively. Western blots were visualized using an ECL kit
(Pierce; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. β-actin was served as an internal loading
control for each blot. The intensity of the detected bands was
analyzed using an ImageJ software (version 1.42; National
Institutes of Health).
Wound healing/scratch assay
Wound healing/scratch assays were carried out as
previously described (23). In
brief, cells were plated in 6-well plates and cultured to a
monolayer. Next, a scratch was created in the middle using a 10-µl
pipette tip. The medium was replaced by 1% bovine serum albumin
(BSA) in RPMI medium. The cells were incubated at 37°C in a sterile
incubator, and the wound trace was photographed at 0, 5, 10, 15 and
20 h. The distance of the scratch (in mm) was measured under a
light microscope at magnification ×200. All samples were examined
in triplicate, and the data are shown as the means.
Invasion assay
To evaluate the cell invasion ability, Transwell
assays were carried out as previously described (3,23). In
brief, the cell suspension at a final concentration of
4×105/ml was added into the upper Transwell inserts
precoated with Matrigel, and then the lower insert was loaded with
10% FBS in cell culture medium. After 24 h of incubation at 37°C in
a sterile incubator, the invaded cells were fixed. The invaded
cells were stained with 0.2% crystal violet at room temperature for
10 min. The number of invaded cells was counted under an inverted
microscope. All samples were examined in triplicate, and the data
are showed as the means.
Chemotaxis assay
Chemotaxis assay was performed as described
previously (3,23). Briefly, a polycarbonate filter
membrane was inserted between the upper and lower chambers. After
pretreatment with 10 µg/ml fibronectin overnight, it was dried in
air. CCL18, as chemoattractant, was added into the lower chamber
and the cell suspension at a final concentration of
5×105/ml was added to the upper chambers. Then, the
chamber was incubated at 37°C in a sterile incubator for 3 h. The
filter membrane was washed, fixed and stained. The number of
migrating cells was calculated using a microscope at a
magnification ×400 in three random fields. Chemotaxis index=the
number of migrating cells in the chemoattractant gradient/the
number of migrating cells in the medium control. All samples were
tested in triplicate and the data are shown as the means.
Adhesion assay
The adhesion assays was conducted as described
previously (23). The cells
suspension at a final concentration of 3×105/ml
incubated at 37°C in a sterile incubator for 20 min, and then,
cells were promptly added to a 24-well plate containing dried glass
coverslips with or without 10 ng/ml rCCL18. The coverslips were
pretreated with fibronectin and then dried for 30 min. After 5, 15
and 30 min of incubation, the cells were washed and fixed. The
number of cells attached to the coverslips were calculated using a
light microscope at magnification of ×400.
Cellular F-actin measurement
F-actin polymerization assay was performed as
previously described (4,24). In brief, cells were cultured for 24
h in complete medium and cultured in binding medium for 2 h. Cells
were fixed, permeabilized, and stained in the dark with
TRITC-labelled Phalloidin, which was diluted in F-buffer (10 mM
HEPES, 20 mM KH2PO4, 5 mM EGTA, 2 mM
MgCl2, Dulbecco's PBS, pH 6.8) for 30 min. Cells was
measured by fluorescence analysis. The F-actin content was
calculated by the following equation: F-actin Δt/F-actin
0=(fluorescence Δt/mg per ml/fluorescence 0/mg per ml). All
experiments were repeated at least three times.
Immunofluorescence
Immunofluorescence was performed as previously
described (3). Briefly, the cells
were cultured in 24-well plates containing sterile coverslips 24 h
before this experiment, and then starved overnight. After
stimulation with 10 ng/ml rCCL18, cells were washed, fixed and
permeabilized as previously described (3). Cells were incubated in primary
antibody overnight at 4°C. Negative control comprised rabbit IgG.
The cells were then directly incubated at room temperature for 2 h
in dark with Alexa Fluor 594-conjugated secondary antibodies and
4′,6-diamidino-2-phenylindole (DAPI) in the dark. The results were
analyzed using fluorescence microscopy. All experiments were
repeated at least three times.
In vivo spontaneous metastasis
assay
Metastasis assays were performed as described
previously (3). Forty SCID mice
were randomly divided into four groups. In total, the Scr/MDA-231
or SiAnxa2/MDA-231 cell suspension at a final concentration of
2×106/ml was injected into the mammary fat pads of the
mice. When the xenografts were 0.5 cm in diameter, an intratumoral
injection of 100 ng/kg rCCL18 or the same volume of vehicle was
administered biweekly for 4 continuous weeks. After 8 weeks, the
mice were sacrificed by cervical dislocation, and the lung surface
nodules were counted. The lung tissues were fixed, embedded in
paraffin, serial sectioned and H&E stained to detect lung
micrometastasis. The study protocol for the animal study was
reviewed and approved by Weifang Medical University Ethics
Committee (approval no. 045, 26-Feb-2016).
Statistical analyses
Data are presented as mean ± SEM from a
representative experiment; each assay was independently repeated at
least three times. All statistical analyses were performed using
SPSS v17.0 software (SPSS, Inc.). Student's t-test or one-way
analysis of variance (ANOVA) (SigmaPlot; SigmaStat; Jandel
Scientific Software) was used for comparisons between groups, and
the relationship between the clinicopathologic characteristics and
AnxA2 expression was determined using Chi-square test, with
P<0.05 considered statistically significant.
Results
Expression of AnxA2 in breast cancer
and the correlation between AnxA2 and clinicopathological
characteristics of the breast cancer cases
Comparative evaluation was carried out of the AnxA2
expression of the paired invasive ductal carcinoma tissues and
adjacent non-tumor tissues by western blot analysis. The coupled
tissues were obtained from the same patient. Our results showed
that the protein level of AnxA2 was overexpressed in invasive
ductal carcinoma tissues when compared with that in the coupled
adjacent non-tumor tissues (Fig.
1A, left image). To further investigate the correlation between
AnxA2 and the clinicopathological characteristics of the breast
cancer cases, we assessed the expression of AnxA2 in 120 archived
paraffin-embedded specimens of invasive ductal carcinoma and the
coupled adjacent non-tumor tissues by qRT-PCR. Our experimental
results indicated that the expression of AnxA2 in the invasive
ductal carcinoma tissues was significantly associated with tumor
differentiation, lymphatic metastasis, distant metastasis and Nir1
expression, but it was independent of age and tumor size (Table I).
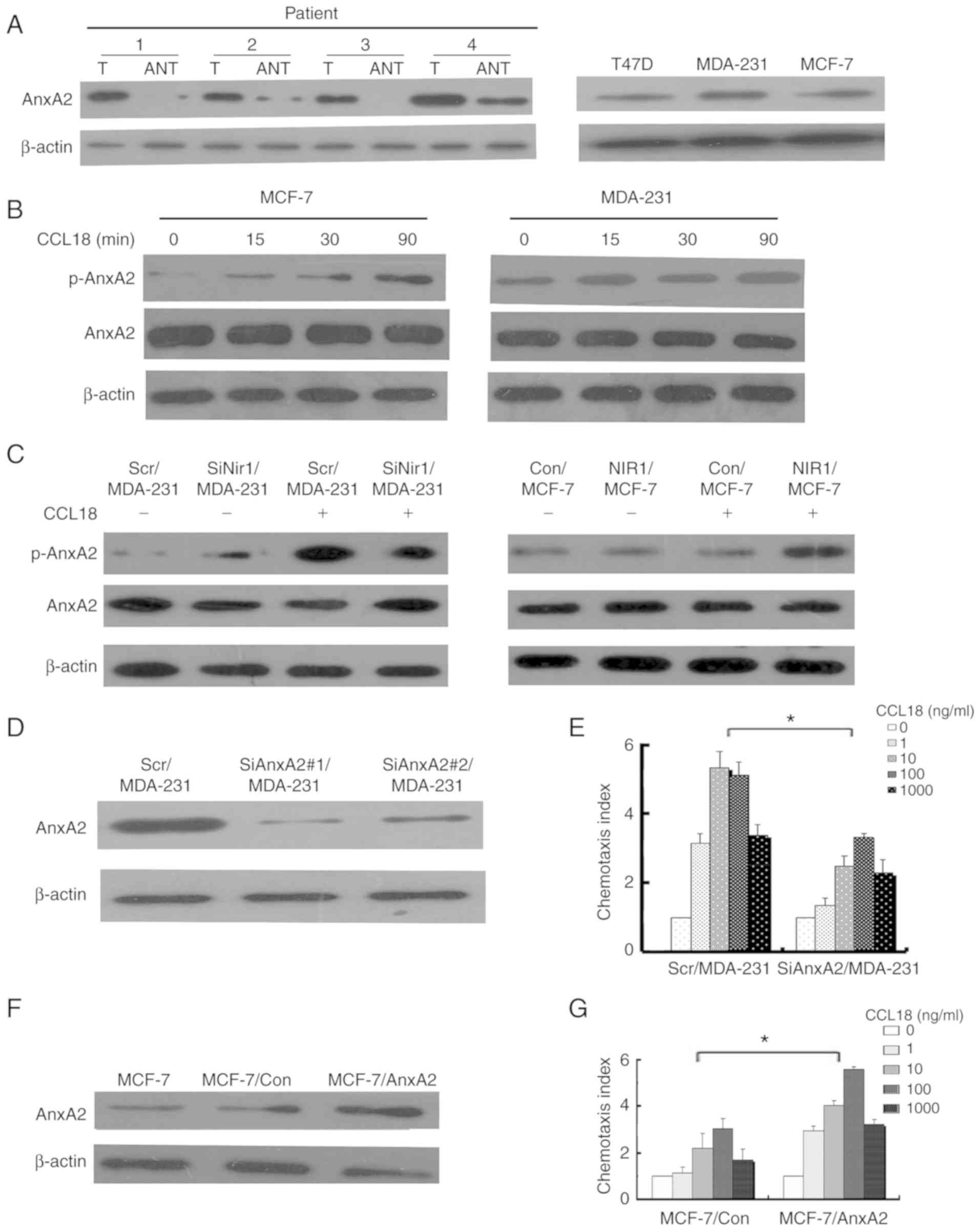 | Figure 1.AnxA2 expression in samples from
breast cancer patients and breast cancer cell lines. (A) Left
panels: Expression of AnxA2 protein in coupled breast cancer
tissues (T) and adjacent tissues (ANT), with each pair obtained
from the same patient. Right panels: Expression of AnxA2 protein in
breast cancer cell lines (T47D, MDA-MB-231, MCF-7). β-actin was
used as a loading control. The results are representative of at
least three indenpent experiments. (B) Expression levels of AnxA2
protein in MDA-MB-231 and MCF-7 cell lines upon 10 ng/ml rCCL18
stimulation for 0, 15, 30 and 90 min. β-actin was used as a loading
control. (C) Left panels: Expression levels of AnxA2 protein in
Scr/MDA-231 and SiNir1/MDA-231 cells with or without 10 ng/ml
rCCL18 stimulation were detected by western blot analysis. Right
panels: Expression levels of AnxA2 protein in Con/MCF-7 and
Nir1/MCF-7 cells with or without 10 ng/ml rCCL18 stimulation were
detected by western blot analysis. β-actin was used as a loading
control. (D) Expression of AnxA2 protein in MDA-MB-231 cells
transfected with a scrambled SiRNA as a control (Scr/MDA-231) and
with two sets of stable SiRNA-targeting AnxA2 (SiAnxA2#1/MDA-231
and SiAnxA2#2/MDA-231) was detected by western blot analysis.
β-actin was used as a loading control. (E) Comparison of
chemotactic responses with rCCL18 stimulation in Scr/MDA-231 and
SiAnxA2/MDA-231 (SiAnxA2#1/MDA-231) cells. Columns, mean of
triplicate measurements; Bars, standard deviation. *P<0.05
(two-way ANOVA). (F) Expression of AnxA2 protein in Con/MCF-7 and
AnxA2/MCF-7 (stable clone 2) cells was detected by western blot
analysis. β-actin was used as a loading control. (G) Comparison of
chemotactic responses with rCCL18 stimulation in Con/MCF-7 and
AnxA2/MCF-7 cells. The data were collected from a representative of
at least three independent experiments. Columns, mean of triplicate
measurements; bars, standard deviation. *P<0.05 (two-way ANOVA).
CCL18, chemokine (C-C motif) ligand 18; Nir1, PYK2 N-terminal
domain interacting receptor 1; AnxA2, Annexin A2. |
As our results showed that the protein level of
AnxA2 was overexpressed in invasive ductal carcinoma tissues
compared with that in the coupled adjacent non-tumor tissues, we
assessed the AnxA2 level in breast cancer cell lines. The protein
level of AnxA2 was diverse in the breast cancer cell lines. Highly
invasive cells (MDA-MB-231) expressed a higher level of AnxA2 than
the low-invasive cells (T47D and MCF-7) (Fig. 1A, right image). Furthermore, CCL18
affected the phosphorylation levels of AnxA2 in breast cancer cells
in a time-dependent manner, but not the expression of AnxA2 in
MCF-7 and MDA-MB-231 cells (Fig.
1B).
AnxA2 contributes to CCL18-induced
chemotaxis in breast cancer cells
We used siRNA to inhibit Nir1 or AnxA2 expression to
determine the specific function of AnxA2 breast cancer cells.
Tygromycin B was used for selecting the successfully transfected
cells that stably downregulated the expression of Nir1 or AnxA2.
SiRNAs (SiAnxA2#1 and SiAnxA2#2) were designed to target Nir1 or
AnxA2. Scr/MDA-231 cells were used as control cells that were
transfected with a scrambled sequence. We chose SiNir1/MDA-231 and
SiAnxA2/MDA-231 cells as the representatives.
Simultaneously, AnxA2 or Nir1 stably transfected
cell clones were generated by pcDNA3.1-AnxA2 or pcDNA3.1-Nir1
plasmid subsequent selection. Stable transfected cells were
obtained by using medium (culture medium with 600 ml G418). Hence,
we decided to present the results from clone 4, designated as
AnxA2/MCF-7 and Nir1/MCF-7 cells. Con/MCF-7 cells were used as
vector control cells that were transfected with a pcDNA3.1 vector.
We performed western blot analysis to identify the protein and
phosphorylation levels of AnxA2 in Scr/MDA-231, SiNir1/MDA-231,
Con/MCF-7, and Nir1/MCF-7 cells with or without rCCL18 stimulation.
Our results indicated that the expression of Nir1 affected
CCL18-induced AnxA2 phosphorylation. In other words, CCL18 binding
to Nir1 promoted the phosphorylation levels of AnxA2 in breast
cancer cells, but not the expression of AnxA2 (Fig. 1C). We performed western blot
analysis to identify AnxA2 in stably transfected cell clones. The
results are illustrated in Fig. 1D and
F. We performed chemotaxis assay to determine whether AnxA2
contributed to CCL18-induced breast cancer cell chemotaxis. The
CCL18-induced chemotaxis ability of the diverse breast cancer cell
lines followed a representative bell-shaped response curve
(Fig. 1E and G). These chemotaxis
assay results indicated that AnxA2 promotes CCL18-induced
chemotaxis in breast cancer cells.
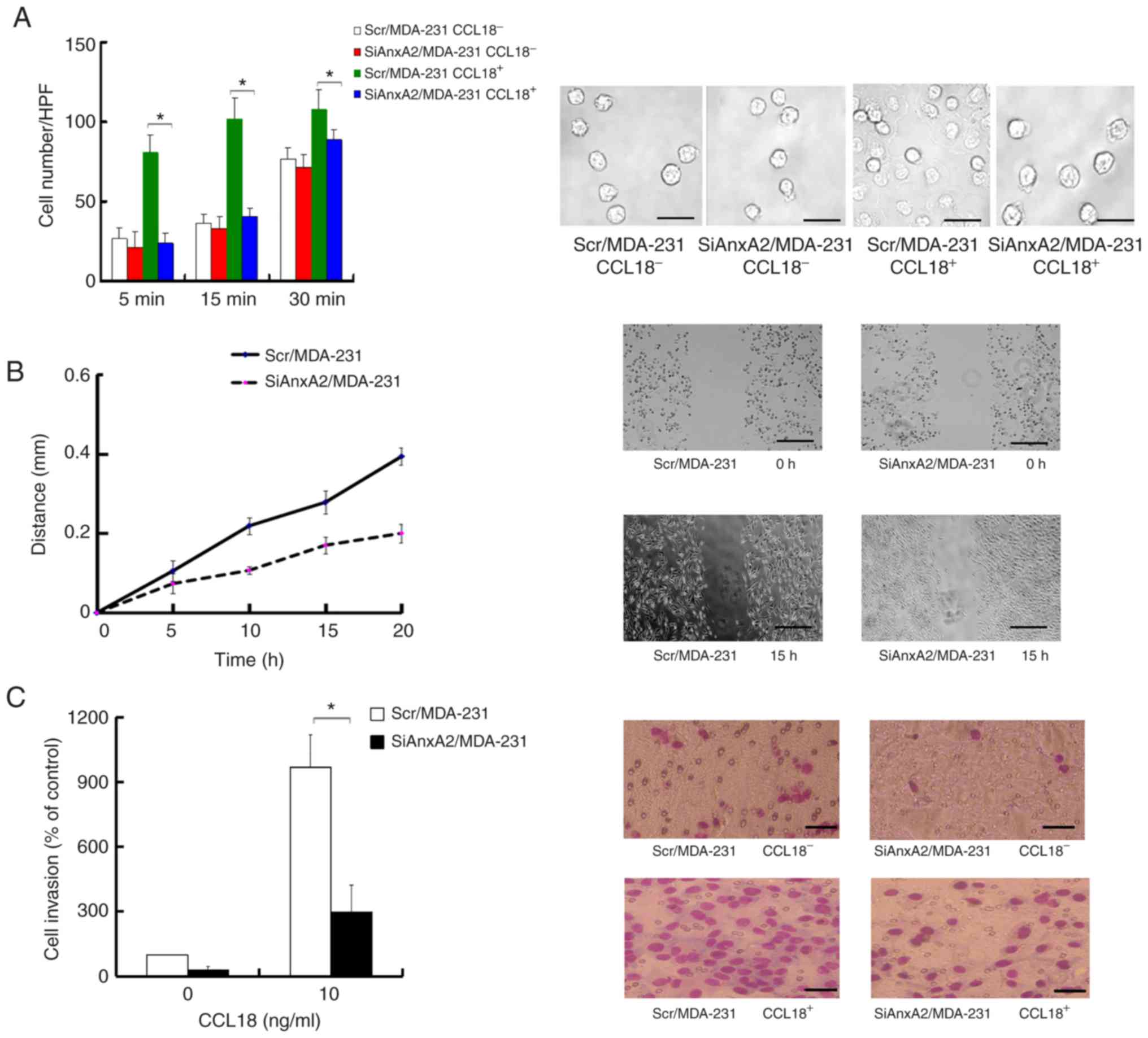
Decrease in AnxA2 suppresses adhesion,
migration and invasion in breast cancer cells
Migration, invasion and proliferation are correlated
with a highly aggressive phenotype in breast cancer. Previous
studies (9,17,18,25,26)
suggest that AnxA2 promotes invasion, migration, proliferation and
adhesion in cancer cells. Moreover, chemokine-induced adhesion
contributes to cell movement (26).
We aimed to determine the influence of AnxA2 on the adhesion in
breast cancer cells with or without CCL18 stimulation. Our results
revealed that treatment with 10 ng/ml of rCCL18 promoted the
adhesion, which was significantly impaired in the SiAnxA2/MDA-231
cells (Fig. 2A). The results
indicated that AnxA2 promoted CCL18-induced cell adhesion. To
identify whether AnxA2 affects CCL18-induced migration and invasion
in breast cancer cells, we carried out scratch assays and Transwell
invasion assays. When a wound was created in the cells, the
distance of the SiAnxA2/MDA-231 cell migration was more narrow than
the Scr/MDA-231 cells with rCCL18 stimulation (Fig. 2B). The number of SiAnxA2/MDA-231
cells that invaded the Matrigel were considerably fewer than the
Scr/MDA-231 cells with 10 ng/ml rCCL18 stimulation after 24 h, and
there was no obvious difference in the invasion between
SiAnxA2/MDA-231 and Scr/MDA-231 cells without rCCL18 stimulation at
24 h (Fig. 2C). These results
showed that suppression of AnxA2 inhibited CCL18-induced adhesion,
migration and invasion in breast cancer cells.
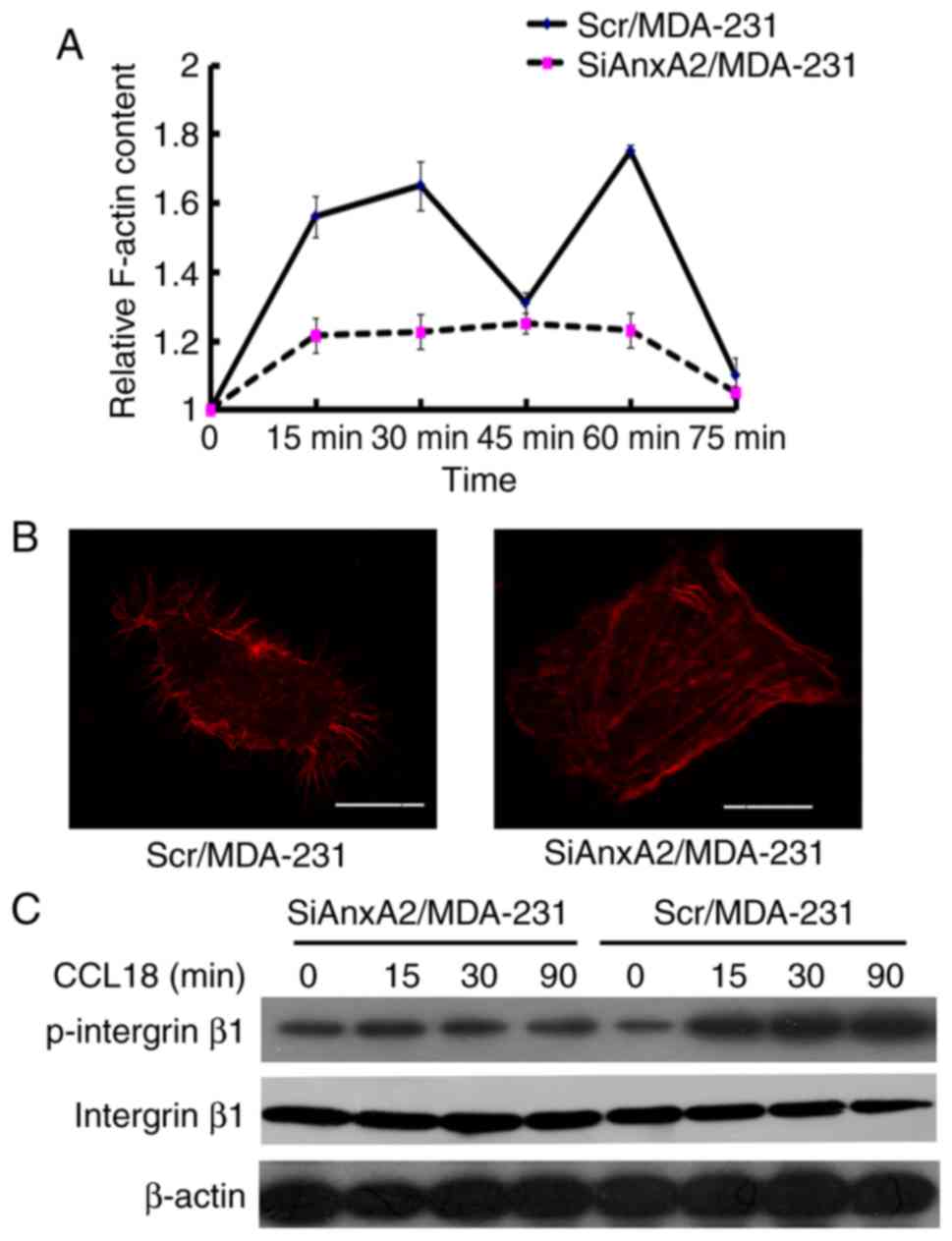
Reduction in AnxA2 suppresses
CCL18-induced F-actin polymerization in breast cancer cells
The migration and chemotaxis of cells depend on the
actin-based cytoskeleton (27,28).
The process of F-actin polymerization is quick and transient
(29). To testify the hypothesis
that the downregulation of AnxA2 could suppress CCL18-induced
chemotaxis of breast cancer cells by suppressing F-actin
polymerization, we performed F-actin polymerization assay. The
results indicated that CCL18 elicited transient actin
polymerization at 15 and 60 min in the Scr/MDA-231 cells. Actin
polymerization was significantly decreased in the SiAnxA2/MDA-231
cells (Fig. 3A), which suggested
that AnxA2 plays a vital role in regulating cytoskeleton
rearrangement with CCL18 stimulation. Immunofluorescent staining
was used to detect the F-actin content. rCCL18 increased the
F-actin content in the Scr/MDA-231 cells, but not in
SiAnxA2/MDA-231 cells (Fig. 3B).
Integrin β1 binding to fibronectin plays a crucial part in adhesion
of breast cancer cells (30). Thus,
western blot analysis was performed to detect the CCL18-induced
activation of integrin β1. As shown in Fig. 3C, the CCL18-induced phosphorylation
of integrin β1 was distinctly impaired in the SiAnxA2/MDA-231
cells, in accordance with a reduction in adhesion. The results
indicated that AnxA2 regulated migration and invasion directly by
modulating the CCL18-induced adhesion and F-actin polymerization in
the MDA-MB-231 cells.
AnxA2 promotes CCL18-induced EMT via
the Akt/GSK3β signaling pathway
EMT is a well-coordinated process during cancer
progression, which is related to the metastatic potential of tumor
cells. Our previous study (3)
indicated that CCL18 binding to Nir1 enhances the invasion ability
of breast cancer cells through the PI3K/Akt/GSK3β/Snail signaling
pathway. We assumed that AnxA2, as the downstream molecule of CCL18
that binds to Nir1, promotes progression and metastasis by EMT of
breast cancer cells through the PI3K/Akt/GSK3β/Snail signaling
pathway in breast cancer cells. To investigate the role of AnxA2 in
the promotion of migration and metastasis in breast cancer cells
through EMT, we performed western blot assays and
immunofluorescence to detect vimentin, N-cadherin (mesenchymal
marker), and E-cadherin (epithelial marker) in cells with or
without 10 ng/ml rCCL18 stimulation for 3 days. Western blot assays
showed that compared with the Scr/MDA-231 cells, the expression
levels of N-cadherin and vimentin were distinctly lower and the
expression of E-cadherin was higher in the SiAnxA2/MDA-231 cells
with 10 ng/ml rCCL18 for 3 days (Fig.
4A, left panels). In AnxA2/MCF-7 cells, the expression of
E-cadherin was downregulated, whereas levels of vimentin and
N-cadherin were upregulated (Fig.
4A, right panels). Immunofluorescence staining of cells
indicated the same results (Fig.
4B). Next, the nuclear expression of transcriptional factors
was detected to verify the downstream molecules that participate in
EMT, which were modulated by AnxA2 in cells. The nuclear expression
of Snail was distinctly upregulated in the Scr/MDA-231 cells with
10 ng/ml rCCL18 stimulation for 3 days when compared with that in
the SiAnxA2/MDA-231 cells (Fig. 4C,
left panels). The expression of Snail was upregulated in
AnxA2/MCF-7 cells with 10 ng/ml rCCL18 stimulation when compared
with the Con/MCF-7 cells (Fig. 4C,
right panels). To confirm that AnxA2, as an upstream regulator,
plays a role in the Akt/GSK3β signaling pathway during cell
chemotaxis, we investigated the influence of AnxA2 expression on
CCL18-induced activation of Akt and GSK3β. Phosphorylated Akt and
GSK3β were assessed after stimulation with 10 ng/ml rCCL18 for 3
days. The results showed that the phosphorylation of Akt at Ser473
and Thr308 and phosphorylation of GSK3β at Ser 9 were decreased in
the SiAnxA2/MDA-231 cells compared with the Scr/MDA-231 cells
(Fig. 4D, left panels). The
phosphorylated levels of Akt and GSK3β were increased in
AnxA2/MCF-7 cells when compared with the Con/MCF-7 cells (Fig. 4D, right panels). We treated the
cells with an inhibitor (LY294002) specific to the Akt/GSK3β
signaling pathway to detect the activation of AnxA2 with rCCL18
stimulation for 3 days. The results indicated that the
phosphorylation of AnxA2 was reduced in the SiAkt/MDA-231 cells and
MDA-MB-231 cells with LY294002 compared with the Scr/MDA-231 cells
(Fig. 4E). These results suggest
that the PI3K/Akt/GSK3β/Snail signaling pathway is pivotal for
AnxA2 to promote CCL18-induced EMT in breast cancer cells.
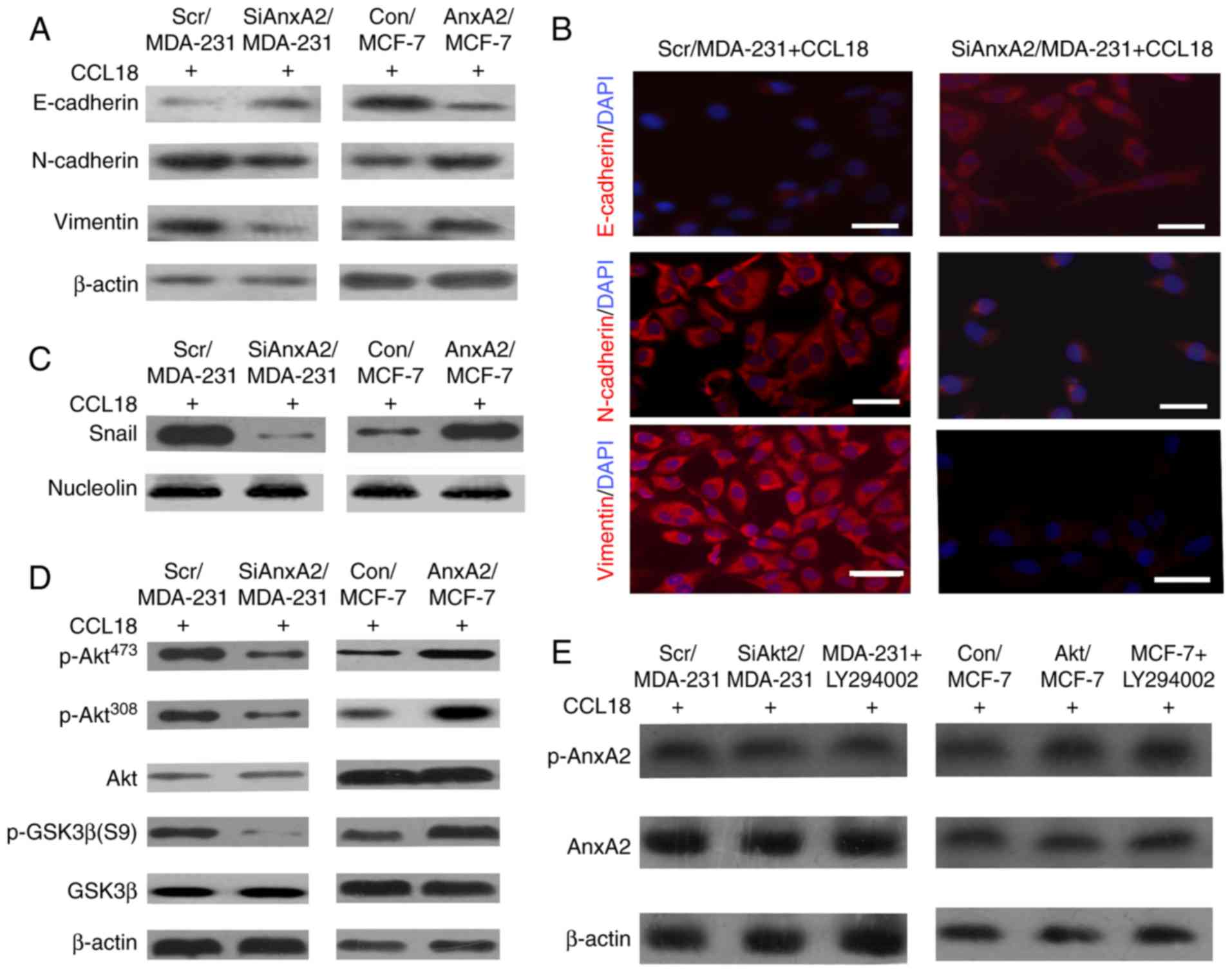 | Figure 4.AnxA2 promotes CCL18-induced EMT via
the Akt/GSK3β signaling pathway. (A) Expression levels of an
epithelial marker (E-cadherin), as well as mesenchymal markers
(N-cadherin and vimentin) were assessed by western blot analysis in
Scr/MDA-231, SiAnxA2/MDA-231, Con/MCF-7 and AnxA2/MCF-7 cells upon
10 ng/ml rCCL18 stimulation. β-actin was used as a loading control.
(B) Fluorescence microscopic staining of E-cadherin, N-cadherin and
vimentin (red) is indicated in the Scr/MDA-231, SiAnxA2/MDA-231,
Con/MCF-7 and AnxA2/MCF-7 cells upon rCCL18 stimulation. Nuclear
DNA was stained with DAPI (blue). Scale bar, 20 µm. (C) Nuclear
expression level of Snail in CCL18-induced Scr/MDA231 and
SiAnxA2/MDA-231 cells was examined by Western blot analysis.
Nucleolin was used as a loading control. rCCL18, 10 ng/ml. (D)
Expression levels of p-Akt (Ser473), p-Akt (Thr308), p-GSK3β and
GSK3β in the Scr/MDA-231, SiAnxA2/MDA-231, Con/MCF-7 and
AnxA2/MCF-7 cells upon rCCL18 stimulation was detected by western
blot analysis. Akt or β-actin was used as a loading control. (E)
CCL18-induced MDA-231 and MCF-7 cells were pretreated with LY294002
(20 µM) for 1 h. The expression levels of p-AnxA2 and AnxA2 in the
Scr/MDA-231, SiAkt/MDA-231, and MDA-231 + LY294002, Con/MCF-7,
Akt/MCF-7 and MCF-7 + LY294002 cells were detected by western blot
analysis. rCCL18, 10 ng/ml. Data were collected from a
representative of at least three independent experiments. CCL18,
chemokine (C-C motif) ligand 18; AnxA2, Annexin A2. |
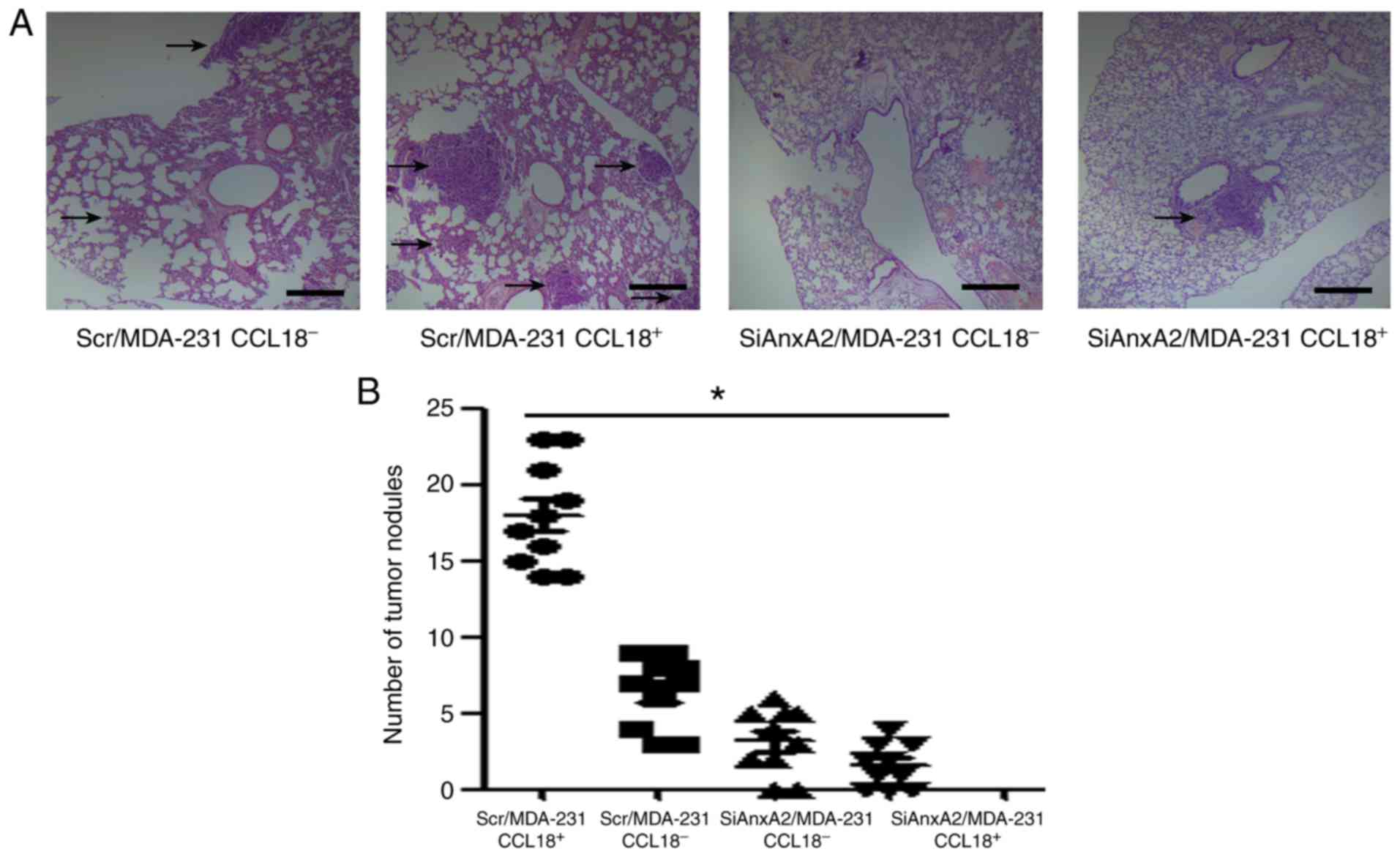
AnxA2 plays a role in breast cancer
metastasis with CCL18 stimulation in vivo
A xenograft transplant model in SCID mice was
performed to identify the metastatic properties of breast cancer
cells. Colonization in lungs was examined through H&E staining.
As showed in Fig. 5A, the results
showed that the number of metastatic tumor nodules was increased in
the lungs of mice whose Scr/MDA-231 cells were injected with rCCL18
compared with that in the lungs of mice whose SiAnxA2/MDA-231 cells
were injected with rCCL18. Quantitative analysis of the number of
metastatic tumor nodules indicated that compared with Scr/MDA-231
cells, the SiAnxA2/MDA-231 cells a 5-fold lower number of
metastatic tumor nodules in response to 10 ng/ml CCL18 (Fig. 5B). In our present study, we
demonstrated that AnxA2 contributes to CCL18-induced metastasis of
human breast cancer. Thus, our results revealed that AnxA2 was
vital for CCL18-induced metastasis of human breast cancer
cells.
Discussion
Various nonmalignant stromal cells in the tumor
microenvironment play a pivotal role in tumor progression and
metastasis (31). CCL18, which is
derived from TAMs, is correlated with the invasion, migration,
metastasis and poor prognosis of patients with breast cancer
(32,33). Decades of research have shown that
upregulation of Annexin A2 (AnxA2) is involved in many malignant
neoplasms, and is associated with the aggressiveness of breast
cancer cells (17,34–36).
Furthermore, AnxA2 may also be a significant biomarker for breast
cancer diagnosis and a potential target for breast cancer therapy
(37,38). In the present study, pathological
investigation of breast tissue specimens revealed that compared
with adjacent non-tumor tissues, AnxA2 was highly regulated in
breast cancer tissues, and AnxA2 was upregulated in 68% of patients
with lymph node metastases and in 32% of those with non-lymph node
metastases. AnxA2 was also highly expressed in 65% in patients with
distant metastases. These findings suggested that AnxA2 was highly
expressed in most breast cancer patient tissues and plays a vital
role in lymph node and distant metastases. In addition, AnxA2 was
upregulated in highly invasive human breast cancer cells. This
finding indicated that AnxA2 contributes to the invasion of breast
cancer cells. In animal experiments, AnxA2 enhanced the metastasis
of human breast cancer cells to lungs of SCID mice with rCCL18
stimulation, which provides molecular evidence that AnxA2 is
indispensable for CCL18-induced metastasis of human breast cancer
cells in vivo. For tumors to metastasize and invade,
neoplastic and endothelial cells must migrate into surrounding
tissues (39). In addition,
migration is highly associated with infiltration and chemotaxis
(40,41). In the present study, upregulation of
AnxA2 expression distinctly enhanced the migration ability of the
human breast cancer cells upon rCCL18 stimulation in the chemotaxis
and scratch assays. These results indicate that AnxA2 enhanced
CCL18-induced migration in breast cancer cells. Chemotaxis is the
ability of cells to migrate in response to a chemoattractant. Cell
migration can be viewed as a multistep of discrete processes that
result in net cell-body movement (42). Our study demonstrated that AnxA2
contributed to F-actin polymerization to mediate cytoskeletal
rearrangement, providing the driving force for migration and
invasion of breast cancer cells. Adhesion contributes to cell
movement, and the firm adhesion of tumor cells to the extracellular
matrix (ECM) is regulated by integrin β1 (26). Our results revealed that silencing
of AnxA2 in MDA-MB-231 cells inhibited CCL18-induced
phosphorylation of integrin β1. Although AnxA2 plays a pivotal role
in promoting invasion and migration in breast cancer cells, other
chemokines or proteins may also have undiscovered roles. Further
research should be performed to determine the undiscovered elements
associated with invasion, migration and metastasis in breast cancer
cells. EMT is indispensable for tumor cell migration, invasion and
metastasis in all types of malignant tumors (43). Our previous study (3) showed that MCF-7/Nir1 cells increased
invasion and migration compared with the MCF-7/Con cells, and the
binding of CCL18 to Nir1 enhanced EMT in breast cancer cells
through the PI3K/Akt/GSK3β/Snail signaling pathway. Moreover, the
expression levels of other transcriptional factors (Slug, Twist1,
Zeb1 and Zeb2) were stable in the Scr/MDA-231 and SiNir1/MDA-231
cells with or without CCL18 stimulation. Akt contributes to
invasion and metastasis, which is a predominant component of the
PI3K pathway. Our results showed that AnxA2 depletion in breast
cancer cells inhibited CCL18-induced Akt and GSK3β phosphorylation,
whereas the expression of Akt and GSK3β was not altered. In
addition, AnxA2 enhanced the stabilization of Snail, which is a
crucial transcriptional factor of the PI3K/Akt/GSK3β/Snail
signaling pathway. In this study, AnxA2 promoted CCL18-induced EMT
through the stabilization of Snail via the PI3K/Akt/GSK3β/Snail
signaling pathway.
In conclusion, our findings indicated that
upregulation of AnxA2 promotes breast cancer cell invasion and
metastasis. More importantly, we elucidated the molecular mechanism
by which AnxA2, as a downstream molecule of CCL18 binding to Nir 1,
promotes the progression of breast cancer cell lines. Thus, our
research suggests a novel mechanism for CCL18-induced breast cancer
cell invasion and metastasis and indicates a valuable prognostic
marker and novel therapeutic target for breast cancer.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Scientific Foundation of China (nos. 81672631, 81872163, 81072068,
81472365 and 81572578), The Young and Middle-Aged Scientists
Research Awards Foundation of Shandong Province (2010BSB14050), the
Foundation of Shandong Educational Committee (J14LK13), the
Scientific Foundation of Shandong Province (ZR2014HM003 and
ZR2015HM028) and the Shandong Province Outstanding Youth Scientist
Foundation Plan (BS2013YY020).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
BZ conceived and designed the study. CZ, SZ, ZY, ZD
and RW performed the experiments and collected and analyzed the
data. CZ, SZ and BZ wrote the manuscript. All authors read and
approved the manuscript and agree to be accountable for all aspects
of the research in ensuring that the accuracy or integrity of any
part of the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
Patient samples were collected according to a
protocol approved by the Institutional Review Board, and patients
provided signed consent for use of their tissue specimens in the
present study. The study protocol was reviewed and approved by the
Weifang Medical University Ethics Committee (approval no. 045,
26-Feb-2016). The study protocol for the animal study was reviewed
and also approved by Weifang Medical University Ethics Committee
(approval no. 045, 26-Feb-2016).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Liu Y, Ji R, Li J, Gu Q, Zhao X, Sun T,
Wang J, Li J, Du Q and Sun B: Correlation effect of EGFR and CXCR4
and CCR7 chemokine receptors in predicting breast cancer metastasis
and prognosis. J Exp Clin Cancer Res. 29:162010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang CZ, Yuan P and Li Y: MiR-126
regulated breast cancer cell invasion by targeting ADAM9. Int J
Clin Exp Pathol. 8:6547–6553. 2015.PubMed/NCBI
|
|
3
|
Zhang B, Yin C, Li H, Shi L, Liu N, Sun Y,
Lu S, Liu Y, Sun L, Li X, et al: Nir1 promotes invasion of breast
cancer cells by binding to chemokine (C-C motif) ligand 18 through
the PI3K/Akt/GSK3β/Snail signalling pathway. Eur J Cancer.
49:3900–3913. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Müller A, Homey B, Soto H, Ge N, Catron D,
Buchanan ME, McClanahan T, Murphy E, Yuan W, Wagner SN, et al:
Involvement of chemokine receptors in breast cancer metastasis.
Nature. 410:50–56. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Harvey JR, Mellor P, Eldaly H, Lennard TW,
Kirby JA and Ali S: Inhibition of CXCR4-mediated breast cancer
metastasis: A potential role for heparinoids? Clin Cancer Res.
13:1562–1570. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schraufstatter IU, Zhao M, Khaldoyanidi SK
and Discipio RG: The chemokine CCL18 causes maturation of cultured
monocytes to macrophages in the M2 spectrum. Immunology.
135:287–298. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen P, Li K, Liang Y, Li L and Zhu X:
High NUAK1 expression correlates with poor prognosis and involved
in NSCLC cells migration and invasion. Exp Lung Res. 39:9–17. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang F, Wang Z, Yuan J, Wei X, Tian R and
Niu R: RNAi-mediated silencing of Anxa2 inhibits breast cancer cell
proliferation by downregulating cyclin D1 in STAT3-dependent
pathway. Breast Cancer Res Treat. 153:263–275. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang T, Yuan J, Zhang J, Tian R, Ji W,
Zhou Y, Yang Y, Song W, Zhang F and Niu R: Anxa2 binds to STAT3 and
promotes epithelial to mesenchymal transition in breast cancer
cells. Oncotarget. 6:30975–30992. 2015.PubMed/NCBI
|
|
10
|
Kpetemey M, Dasgupta S, Rajendiran S, Das
S, Gibbs LD, Shetty P, Gryczynski Z and Vishwanatha JK: MIEN1, a
novel interactor of Annexin A2, promotes tumor cell migration by
enhancing AnxA2 cell surface expression. Mol Cancer. 14:1562015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zheng L, Foley K, Huang L, Leubner A, Mo
G, Olino K, Edil BH, Mizuma M, Sharma R, Le DT, et al: Tyrosine 23
phosphorylation-dependent cell-surface localization of annexin A2
is required for invasion and metastases of pancreatic cancer. PLoS
One. 6:e193902011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Keklikoglou I, Hosaka K, Bender C, Bott A,
Koerner C, Mitra D, Will R, Woerner A, Muenstermann E, Wilhelm H,
et al: MicroRNA-206 functions as a pleiotropic modulator of cell
proliferation, invasion and lymphangiogenesis in pancreatic
adenocarcinoma by targeting ANXA2 and KRAS genes. Oncogene.
34:4867–4878. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mittal P, Klingler-Hoffmann M, Arentz G,
Winderbaum L, Kaur G, Anderson L, Scurry J, Leung Y, Stewart CJ,
Carter J, et al: Annexin A2 and alpha actinin 4 expression
correlates with metastatic potential of primary endometrial cancer.
Biochim Biophys Acta Proteins Proteom. 1865:846–857. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu W and Hajjar KA: The annexin A2 system
and angiogenesis. Biol Chem. 397:1005–1016. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ren XL, He GY, Li XM, Men H, Yi LZ, Lu GF,
Xin SN, Wu PX, Li YL, Liao WT, et al: MicroRNA-206 functions as a
tumor suppressor in colorectal cancer by targeting FMNL2. J Cancer
Res Clin Oncol. 142:581–592. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shi H, Xiao L, Duan W, He H, Ma L, Da M,
Duan Y, Wang Q, Wu H, Song X and Hou Y: ANXA2 enhances the
progression of hepatocellular carcinoma via remodeling the cell
motility associated structures. Micron. 85:26–33. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang F, Liu Y, Wang Z, Sun X, Yuan J,
Wang T, Tian R, Ji W, Yu M, Zhao Y and Niu R: A novel
Anxa2-interacting protein Ebp1 inhibits cancer proliferation and
invasion by suppressing Anxa2 protein level. Mol Cell Endocrinol.
411:75–85. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang F, Zhang L, Zhang B, Wei X, Yang Y,
Qi RZ, Ying G, Zhang N and Niu R: Anxa2 plays a critical role in
enhanced invasiveness of the multidrug resistant human breast
cancer cells. J Proteome Res. 8:5041–5047. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang F, Zhang H, Wang Z, Yu M, Tian R, Ji
W, Yang Y and Niu R: P-glycoprotein associates with Anxa2 and
promotes invasion in multidrug resistant breast cancer cells.
Biochem Pharmacol. 87:292–302. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang HC, Zhang F, Wu B, Han JH, Ji W,
Zhou Y and Niu RF: Identification of the Interaction between
P-Glycoprotein and Anxa2 in multidrug-resistant human breast cancer
cells. Cancer Biol Med. 9:99–104. 2012.PubMed/NCBI
|
|
21
|
Sulaiman A, Yao Z and Wang L:
Re-evaluating the role of epithelial-mesenchymal-transition in
cancer progression. J Biomed Res. 32:81–90. 2018.PubMed/NCBI
|
|
22
|
Thomson S, Petti F, Sujka-Kwok I, Mercado
P, Bean J, Monaghan M, Seymour SL, Argast GM, Epstein DM and Haley
JD: A systems view of epithelial-mesenchymal transition signaling
states. Clin Exp Metastasis. 28:137–155. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang B, Ma Y, Guo H, Sun B, Niu R, Ying G
and Zhang N: Akt2 is required for macrophage chemotaxis. Eur J
Immunol. 39:894–901. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun R, Gao P, Chen L, Ma D, Wang J,
Oppenheim JJ and Zhang N: Protein kinase C zeta is required for
epidermal growth factor-induced chemotaxis of human breast cancer
cells. Cancer Res. 65:1433–1441. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang Y, Wu N, Wang Z, Zhang F, Tian R, Ji
W, Ren X and Niu R: Rack1 mediates the interaction of
P-glycoprotein with Anxa2 and regulates migration and invasion of
multidrug-resistant breast cancer cells. Int J Mol Sci. 17(pii):
E17182016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang B, Gu F, She C, Guo H, Li W, Niu R,
Fu L, Zhang N and Ma Y: Reduction of Akt2 inhibits migration and
invasion of glioma cells. Int J Cancer. 125:585–595. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Antón IM, Jones GE, Wandosell F, Geha R
and Ramesh N: WASP-interacting protein (WIP): Working in
polymerisation and much more. Trends Cell Biol. 17:555–562. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Etienne-Manneville S and Hall A: Rho
GTPases in cell biology. Nature. 420:629–635. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ananthakrishnan R and Ehrlicher A: The
forces behind cell movement. Int J Biol Sci. 3:303–317. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Doerr ME and Jones JI: The roles of
integrins and extracellular matrix proteins in the insulin-like
growth factor I-stimulated chemotaxis of human breast cancer cells.
J Biol Chem. 271:2443–2447. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu Y, Sun R, Wan W, Wang J, Oppenheim JJ,
Chen L and Zhang N: The involvement of lipid rafts in epidermal
growth factor-induced chemotaxis of breast cancer cells. Mol Membr
Biol. 24:91–101. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen J, Yao Y, Gong C, Yu F, Su S, Chen J,
Liu B, Deng H, Wang F, Lin L, et al: CCL18 from tumor-associated
macrophages promotes breast cancer metastasis via PITPNM3. Cancer
Cell. 19:541–555. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lin L, Chen YS, Yao YD, Chen JQ, Chen JN,
Huang SY, Zeng YJ, Yao HR, Zeng SH, Fu YS and Song EW: CCL18 from
tumor-associated macrophages promotes angiogenesis in breast
cancer. Oncotarget. 6:34758–34773. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wu B, Zhang F, Yu M, Zhao P, Ji W, Zhang
H, Han J and Niu R: Up-regulation of Anxa2 gene promotes
proliferation and invasion of breast cancer MCF-7 cells. Cell
Prolif. 45:189–198. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu X, Ma D, Jing X, Wang B, Yang W and
Qiu W: Overexpression of ANXA2 predicts adverse outcomes of
patients with malignant tumors: A systematic review and
meta-analysis. Med Oncol. 32:3922015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sharma MR, Koltowski L, Ownbey RT,
Tuszynski GP and Sharma MC: Angiogenesis-associated protein annexin
II in breast cancer: Selective expression in invasive breast cancer
and contribution to tumor invasion and progression. Exp Mol Pathol.
81:146–156. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sharma MC and Sharma M: The role of
annexin II in angiogenesis and tumor progression: A potential
therapeutic target. Curr Pharm Des. 13:3568–3575. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kesavan K, Nath G and Pandit JK: Sodium
alginate based mucoadhesive system for gatifloxacin and its in
vitro antibacterial activity. Sci Pharm. 78:941–957. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hood JD and Cheresh DA: Role of integrins
in cell invasion and migration. Nat Rev Cancer. 2:91–100. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Noritake J, Watanabe T, Sato K, Wang S and
Kaibuchi K: IQGAP1: A key regulator of adhesion and migration. J
Cell Sci. 118:2085–2092. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lu S, Niu N, Guo H, Tang J, Guo W, Liu Z,
Shi L, Sun T, Zhou F, Li H, et al: ARK5 promotes glioma cell
invasion, and its elevated expression is correlated with poor
clinical outcome. Eur J Cancer. 49:752–763. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Horwitz AR and Parsons JT: Cell
migration-movin' on. Science. 286:1102–1103. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial- mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|