Introduction
Colorectal cancer (CRC) is one of the most
frequently observed substantive malignancies in clinical
operations. CRC is the third most common cancer among male patients
and the second most common cancer among female patients worldwide.
The mortality rate among male patients is the fourth highest and
for female patients it is the third highest among all types of
cancer worldwide (1). The incidence
and mortality rates have been increasing annually over the past two
decades. Research on postoperative prevention and treatment of CRC
has resulted in improvements to treatments; however, the 5-year
survival rate has not improved significantly. One of the major
challenges in treating CRC is tumor neovascularization which
results in invasion and metastasis of CRC to other organ tissues
(2,3). Tumor angiogenesis is one of the
primary means by which CRC invades and metastasizes, and frequently
accompanies invasion and metastasis of CRC.
Hypoxia is a frequently observed pathological state
in solid tumors. Rapid proliferation of CRC cells results in local
tumor tissues becoming hypoxic, and activating the core hypoxia
response factor, upregulation of hypoxia inducible factor 1α
(HIF-1α). HIF-1α is a basic regulatory factor of tumor angiogenesis
during hypoxic conditions. It is composed of a heterodimer of α and
β subunits. The hypoxic microenvironment results in activation of
HIF-1α in tumor cells, which in turn upregulates the expression of
pro-angiogenic factors such as vascular endothelial growth factor
(VEGF) (4,5) and basic fibroblast growth factor
(bFGF) (6), which increase
vascularization in the solid tumor, and participate in multiple
aspects of tumor formation. Therefore, HIF-1α expression levels in
tissues and cells can be used as a key indicator when monitoring
tumor neovascularization (7) and
HIF-1α-based antineoplastic drug filtering (8). It is also a key indicator used to
evaluate the effectiveness of clinical tumor neovascularization
treatments (9).
Tanshinone IIA (Tan IIA) is the active ingredient of
Salvia, a traditional Chinese medicine, which inhibit
growths, induces apoptosis and reverses multidrug resistance in
various types of human cancer cells. However, there are relatively
fewer studies investigating the effects of Tan IIA on tumor
neovascularization and its underlying mechanisms. In our previous
study, it was demonstrated that Tan IIA inhibited angiogenesis in a
CRC mouse model (10). Tan IIA also
decreased VEGF expression levels in CRC cells by downregulating the
expression of COX-2 (11).
Hypoxia-induced tumor neovascularization is frequently observed in
CRC clinically and pre-empts CRC angiogenesis. Therefore, the aim
of the present study was to determine the effect of Tan IIA on
hypoxia and the underlying signaling pathways modulated by Tan
IIA.
The aim of the present study was to provide an
understanding of the means by which Tan IIA prevented angiogenesis.
A previously established HIF-1α overexpression vector and an HIF-1α
RNA interference plasmid were used to determine the effects of Tan
IIA on the expression of pro-angiogenic factors in CRC cells and
tube formation in human umbilical vein endothelial cells (HUVECs)
in normal and hypoxic conditions.
Materials and methods
Lentivirus plasmid
The lentiviral vector pGC-FU and pGC-FU-HIF-1α were
used for HIF-1α overexpression studies. A eukaryotic vector plasmid
containing small interfering (si)R-Mimic was used as a control and
three plasmids containing different siRNA (siR) sequences targeting
HIF-1α were used for knockdown experiments. All plasmids were
purchased from Shanghai GeneChem Co., Ltd.
Cell culture
Human CRC HCT-116 cells were purchased from The Cell
Bank of Type Culture Collection of the Chinese Academy of Sciences
and cultured in RPMI-1640 medium containing 10% fetal calf serum.
HUVECs, were obtained from The Experiment Center of Putuo Hospital
affiliated to Shanghai University of Chinese Medicine and were
cultured in endothelial cell medium containing 15% fetal calf
serum. Both cell lines were incubated at 5% CO2 at
37°C.
Reagents
Tan IIA (98% pure) was purchased from Xi'an Guanyu
Bio-Tech Co., Ltd. and cobalt chloride (CoCl2) was
obtained from Sigma-Aldrich; Merck KGaA.
Lentiviral infection of HCT-116
cells
HCT-116 cells were routinely cultured and during the
logarithmic phase were plated into a 24-well plate at a density of
1×105 cells/ml. Cells were divided into the following
four conditions: Blank group, CoCl2 group, pGC-FU vector
group and pGC-FU-HIF-1α group; with three wells per condition.
Cells were incubated until confluence had reached 30–50%. For
infection, 1×108TU/ml pGC-FU vector or pGC-FU-HIF-1α
lentivirus was added to 60 µl culture medium and this mix was added
to each well. After 8–12 h, the state of the cells was observed,
and the culture medium was replaced with fresh medium. A total of
48 h after infection, fluorescence was observed and subsequent
experiments were performed.
siR-HIF-1α transfection of HTC-116
cells
A total of 5×105 cells/ml HCT-116 cells
were plated per well in a 6-well plate. Cells were divided into the
following six conditions: Blank group, CoCl2 group,
siR-mimic group, siR-HIF-1α sequence 1 + CoCl2 group,
siR-HIF-1α sequence 2 + CoCl2 group and siR-HIF-1α
sequence 3 + CoCl2 group; with three wells per
condition. Cells were incubated until confluence had reached
40–60%. A mixture containing 4 µg of DNA and 12 µl of HilyMax was
added to 100 µl serum-free medium and sufficiently mixed. The
mixture was incubated at room temperature for 15 min and
subsequently added to the wells, which were cultured for 8–12 h.
The medium was replaced with fresh medium and incubated for a
further 48 h. Transfection was confirmed by observing fluorescence
expression under a fluorescent microscope.
Reverse transcription-quantitative
(RT-q)PCR
HCT-116 cells transfected with the various siRNAs
were used for RT-qPCR. The medium from cells treated with
CoCl2 was replaced with fresh medium containing 200 µM
CoCl2 and incubated for 48 h. Subsequently, a total RNA
extraction kit was used to extract RNA according to the
manufacturer's protocol. RNA purity and concentration were
determined. For reverse transcription, 4 µl 5Χ PrimeScript Buffer,
1 µl PrimeScript RT Enzyme Mix I, 1 µl Oligo dT Primer (50 µmol/l),
1 µl random hexamers (100 µmol/l) and 4 µl of total RNA were added
and brought to a final volume of 20 µl using Rnase-free water. The
reverse transcription temperature protocol was as follows: 37°C for
15 min and then 85°C for 5 sec. The primer sequences used were:
HIF-1α forward, 5′-CGAAGTAGTGCTGACCCTGC-3′ and reverse,
5′-AACTTTGTCTAGTGCTTCCATCG-3′; HIF-1α probe,
5′-AGGTGTCTGATCCTGAATCTGGGGCA-3′; GAPDH forward,
5′-CCACTCCTCCACCTTTGAC-3′ and reverse, 5′-ACCCTGTTGCTGTAGCCA-3′,
GAPDH probe; 5′-TTGCCCTCAACGACCACTTTGTC-3′. All primers were
synthesized by Sangon Biotech Co., Ltd. For qPCR, a solution
containing 0.4 µl each of forward and reverse primers, 0.8 µl
probe, 2 µl cDNA, 0.4 µl Rox Reference Dye and 10 µl Premix EX Taq
was prepared. The solution was brought to a final volume of 20 µl
using dH2O. The thermocycling conditions were as
follows: Pre-denaturation, 94°C for 10 sec; followed by 40 cycles
of denaturation, 95°C for 5 sec, annealing at 60°C and extension
for 34 sec. PCR was performed using an ABI 7300 SDS. GAPDH was used
as an internal reference. HIF-1α relative mRNA expression levels
were calculated using the 2−ΔΔCq method (DCq=HIF-1α
Cq-GAPDH Cq) (12).
Cell proliferation assay
A total of 5×103 HCT-116 cells/well were
plated in a 96 well plate (12 wells per condition) and cultured
overnight. Once the cells had adhered, 2, 4, 8, 16, 32, and 64 µM
of Tan IIA were added to the cells cultured under normal
conditions. Additionally, 1, 5, 10, 15 and 20 µM of Tan IIA were
added to the cells cultured under hypoxic conditions (simulated
using 200 µM of CoCl2). Cells were cultured for 24, 48
and 72 h, and HCT-116 medium was routinely collected. ECM
containing 15% FBS was used to produce 5 different volume
concentrations (6.25, 12.5, 25, 50 and 75% HCT-116 medium in the
cell culture). Subsequently, 1×103 cells/ml were plated
per well in a 96 well plate. Once the cells had adhered, the tumor
supernatant cultures were replaced with 5 gradient concentrations
of HCT-116 medium. The cells were cultured for 48 h after which 20
µl CCK-8 solution was added to each well, and cultured for a
further 4 h. Absorbance was measured using a microplate reader at
450 nm/630 nm. Growth inhibition rate (GIR) was calculated using
the following equation: GIR=[1-(ODn-OD0)/(ODc-OD0)]x100.
OD0 was the absorbance in the blank group.
ODc was the absorbance of the normal control group.
ODn was the absorbance of cells treated with the various
doses of Tan IIA. The IC50 of Tan IIA was calculated.
Experiments were repeated three times independently.
ELISA detection of VEGF and bFGF
A total 0.5 ml 1×105 cells/ml suspension
was plated per well. Cells were divided as follow: Blank group,
CoCl2 group, pGC-FU vector group, pGC-FU HIF-1α group,
siR-mimic group, siR-HIF-1α group, siR-HIF-1α + CoCl2
group, Tan IIA-L group (2.5 µM), Tan IIA-M group (5 µM), Tan IIA-H
group (10 µM) and Tan IIA-M + CoCl2 group. Cells were
treated 5 gradient concentrations of HCT-116 medium. To simulate
hypoxia, 200 µM of CoCl2 was added in the
CoCl2 group and cells were incubated. After 48 h, the
supernatant from each well was collected in a sterile area. The
supernatant was centrifuged at 626 × g for 10 min to remove the
cell debris. Specific ELISA kits were used measure the expression
levels of VEGF and bFGF according to the manufacturer's protocol
and experiments were repeated three times.
Western blotting
Total proteins were extracted from cells with
Nuclear and Cytoplasmic Protein Extraction kit (P0027; Beyotime
Institute of Biotechology) and protein detected with Enhanced BCA
Protein Assay kit (P0010; Beyotime Institute of Biotechology).
Lysates were diluted to a protein concentration of 5 µg/µl and 60
µg of protein samples were resolved on a 10% gel using SDS-page.
Proteins were transferred to PVDF membranes (EMD Millipore) using a
Bio-Rad Trans-Blot (Bio-Rad Laboratories, Inc.) at 100 mV and 135
mA. Membranes were blocked in 5% skimmed milk at room temperature
for 2 h or at 4°C overnight. Subsequently, the membranes were
incubated in 5% BSA solution with a HIF-1α primary antibody (cat.
no. 3716S; dilution 1:500; Cell Signaling Technology, Inc.) for 2 h
at room temperature or overnight at 4°C. Subsequently, the
membranes were washed with TBS-Tween four times, 10 min each, after
which the membranes were incubated with the secondary horseradish
peroxidase-conjugated anti-goat antibody (cat. no. HAF109; dilution
1:1,000; R&D Systems China Co., Ltd.) or anti-rabbit antibody
(cat. no. HAF008; dilution 1:1,000; R&D Systems China Co.,
Ltd.) at room temperature for 2 h. The membranes were washed six
times using TBS-Tween. Signals were visualized using by adding
enhanced chemiluminescent reagent (EMD Millipore) for 1–2 min, and
X-ray films were exposed to the membranes and developed.
Densitometry analysis was performed using ImageJ (National
Institutes of Health).
Flow cytometry
A total of 1.5×106 HCT-116 cells were
plated well in a 6-well plate and allowed to adhere. Medium
containing 200 µM of CoCl2 and 2.5, 5 or 10 µM Tan IIA
was added. Cells were further cultured for 24 h, after which the
medium was removed, the cells were trypisinized, resuspended and
fixed. A total of 200 µl of cells at a density of 5×105
were incubated with phycoerythrin-labeled anti-HIF-1α antibody
(cat. no. 79233; dilution 1:1,600; Cell Signaling Technology,
Inc.). As the blank control, 200 µl PBS was used. Cells were
incubated on ice for 40 min in the dark with the antibody and
subsequently fixed with 500 µl of 70% ethanol. Fluorescence was
measured using a flow cytometer (BD Biosciences).
Tube formation assay
HUVECs were divided into separate groups as follows:
HCT-116 group, pGC-FU vector group, pGC-FU-HIF-1α group, siR-mimic
group, siR-HIF-1α group, CoCl2 group, siR-HIF-1α +
CoCl2 group and 1.0, 2.5, 5.0 or 10 µM Tan IIA groups,
with three wells per condition. A suspension of HUVECs at a density
of 1×105 cells/ml single-cell suspension in serum-free
ECM, were plated in a 48-well plate pre-coated with 200 µl
Matrigel. After the cells had adhered, medium from one of the
groups above or 200 µl of tumor medium was added and cells were
cultured. Tube formation of HUVECs was observed and imaged under an
Olympus CKX41 inverted microscope (magnification ×100; Olympus
Corp.) and three images were obtained per well.
Migration assays
A total of 200 µl single-cell suspension of HUVECs
in ECM at a density of 6×105/ml were added to the upper
chamber of a Transwell insert. Tan IIA (1.0, 2.5, 5 or 10 µM) was
added to the wells. In the bottom chamber, 600 µl of ECM containing
20% FBS was added. Cells were incubated for 24 h, after which the
medium in the upper chamber was discarded. The insert was washed
twice with phosphate-buffered saline (PBS), and the cells on the
upper membrane were removed using a cotton swab. Cells which had
migrated were fixed at room temperature for 30 min, stained with
0.1% crystal violet for 20 min, washed by PBS three times and
observed and imaged under a Olympus CKX41 inverted microscope
(magnification ×100; Olympus Corp.). Cells were extracted using 100
µl 10% acetic acid per pore for 10 min. The optical density (OD)
values were measured at 590 nm, and 10% acetic acid in the blank
pores was used as the reference to zero the OD. The inhibition of
migration by Tan IIA was calculated as follows: Inhibition rate=(OD
in the positive control group-OD in the Tan IIA dosing group)/OD in
the positive control group ×100%.
Statistical analysis
Statistical analysis was performed using SPSS
version 22.0 (IBM, Corp.). Data are presented as the mean ±
standard deviation of three repeats. If the data passed tests for
normality and homogeneity of variance, the one-way ANOVA was
applied for statistical inference. If these conditions were not
met, a non-parametric test (Wilcoxon rank sum test) was selected.
The test used a standard α=0.05. P<0.05 was considered the
threshold for a statistically significant difference.
Results
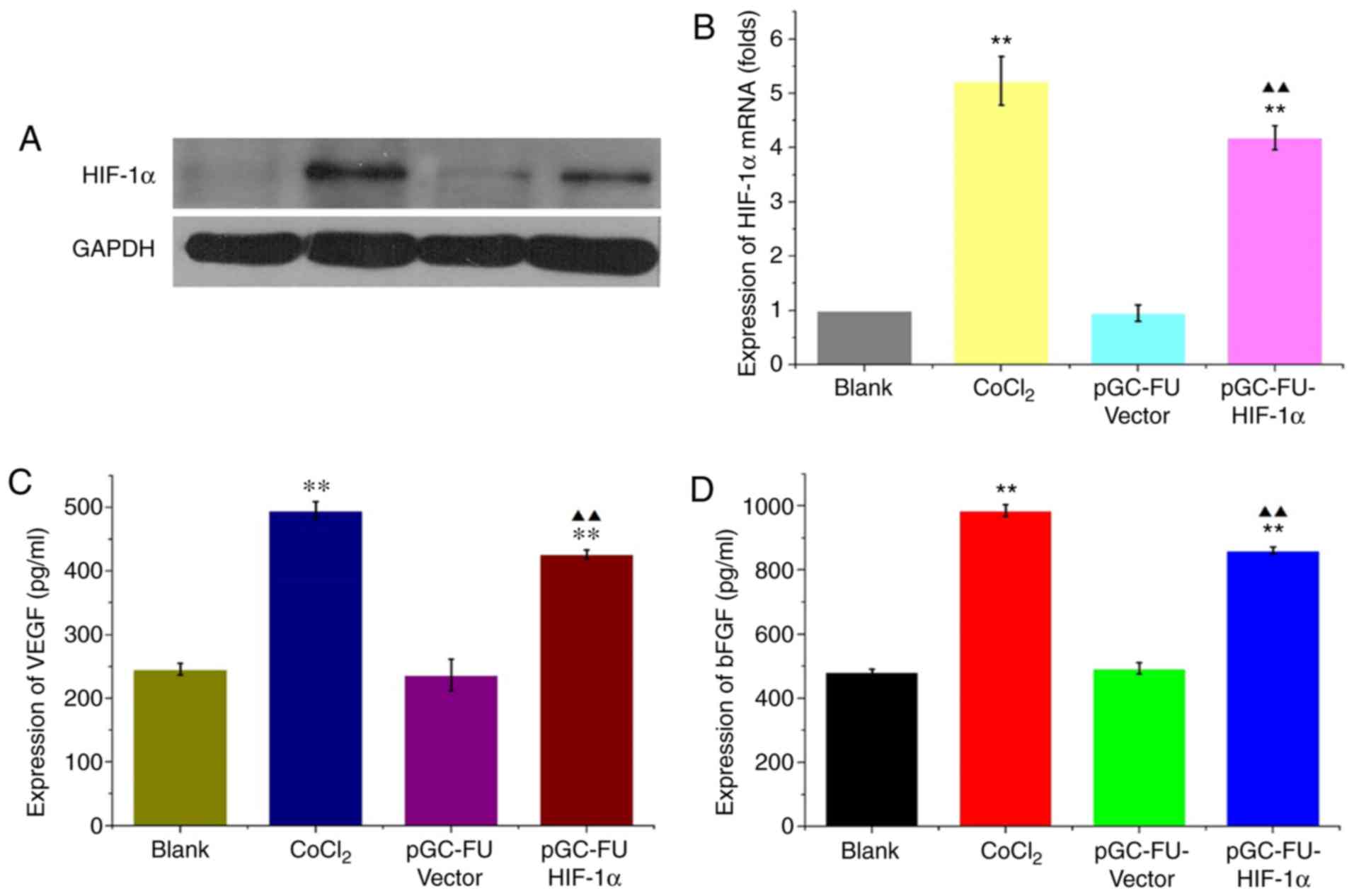
HIF-1α increases the expression of
pro-angiogenic factors in HCT-116 cells
In cells infected with the HIF-1α overexpression
lentiviral vector, pGC-FU-HIF-1α, HIF-1α mRNA expression levels
were 5.01±0.5 times higher compared with the control lentiviral
group, and 4.33±0.15 times higher compared with the blank group.
The protein expression levels of HIF-1α were also increased
significantly (P<0.05; Fig. 1A and
B). In cells infected with pGC-FU-HIF-1α, VEGF expression was
426.576±6.834 pg/ml compared with 246.129±8.948 pg/ml in the blank
group. bFGF expression was 861.39±11.106 pg/ml in cells infected
with pGC-FU-HIF-1α compared with 481.872±9.186 pg/ml in the blank
group. Expression of both VEGF and bFGF were significantly
increased compared to the blank group (both P<0.01; Fig. 1C and D) These results indicated that
pGC-FU-HIF-1α increased HIF-1α expression in HCT-116 cells which in
turn increased secretion of VEGF and bFGF.
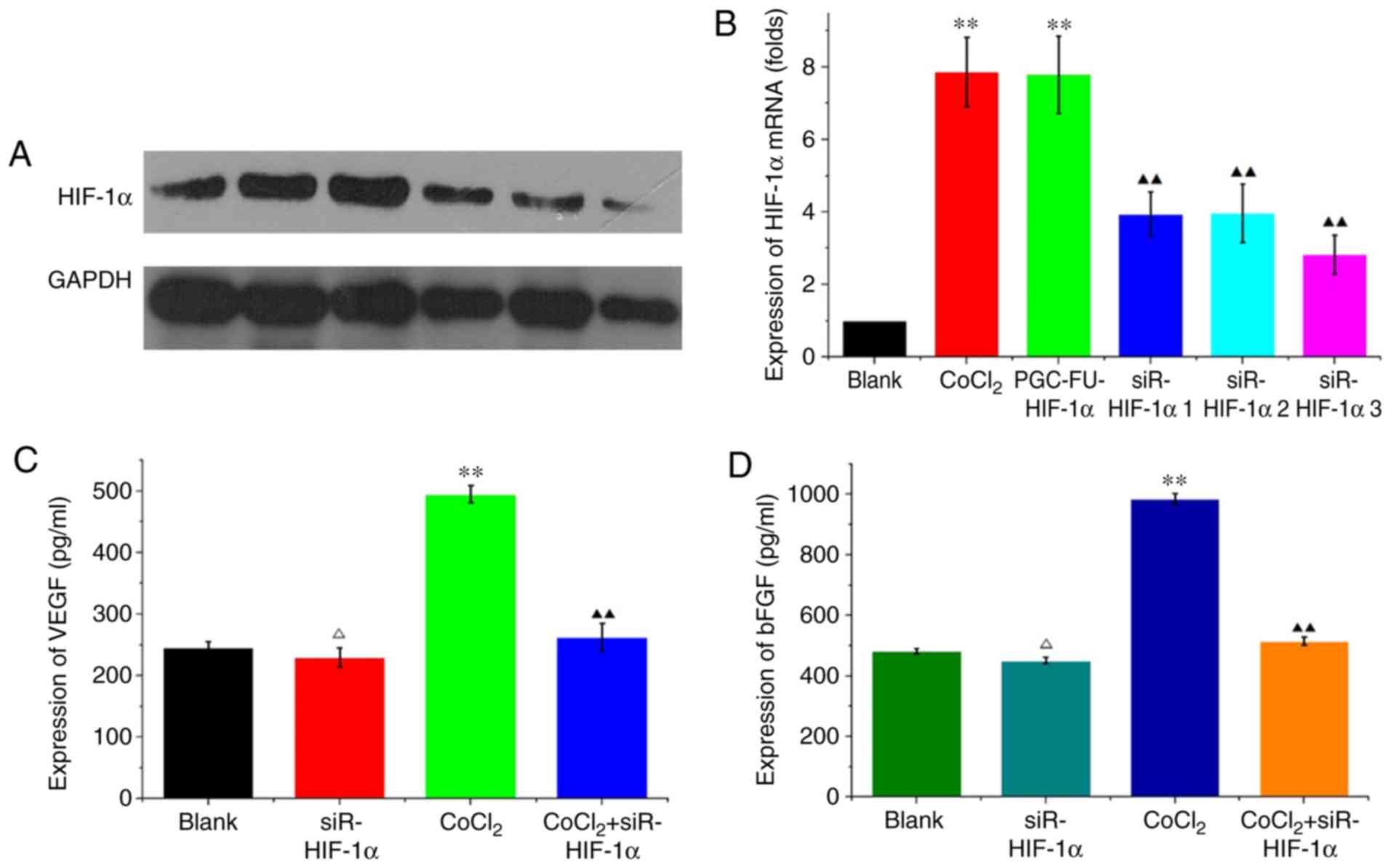
HIF-1α interference in HCT-116 cells
decreases the expression of VEGF and bFGF under hypoxic
conditions
HCT-116 cells were transfected with siR-mimic
control plasmid or one of the three HIF-1α interference plasmids.
Hypoxia was simulated using 200 µM CoCl2 for 48 h.
RT-qPCR and western blotting revealed that HIF-1α mRNA and protein
expression levels were decreased in siR-HIF-1α-transfected cells
compared with CoCl2-treated cells and the blank vector
group. siR-HIF1α sequence 3 demonstrated the greatest decrease on
expression (P<0.01; Fig. 2A and
B) and as such was used in all subsequent experiments. In
HCT-116 cells transfected with siRHIF-1α, VEGF expression was
229.725±15.712 pg/ml compared with 246.129±8.948 pg/ml in the blank
group under normal conditions, and this difference was not
significant (P=0.216; Fig. 2C).
Under hypoxic conditions, VEGF expression was significantly
decreased from 495.176±13.668 pg/ml in the CoCl2 group
compared with 262.533±22.069 pg/ml in cells transfected with
CoCl2 + siR-HIF1α (P<0.05; Fig. 2C). bFGF expression under normal
conditions was decreased from 478.93±11.676 pg/ml in the blank
group to 450.981±11.106 pg/ml in the siR-HIF1α group, although the
difference was not significant (P=0.216; Fig. 2D). However, under hypoxic
conditions, expression of bFGF significantly decreased from
984.954±18.372 pg/ml in the CoCl2 group compared with
514.234±13.239 pg/ml in cells transfected with CoCl2 +
siR-HIF1α (P<0.05; Fig. 2D).
These results indicated that knockdown of HIF-1α reduced VEGF and
bFGF expression under hypoxic conditions but not normal
conditions.
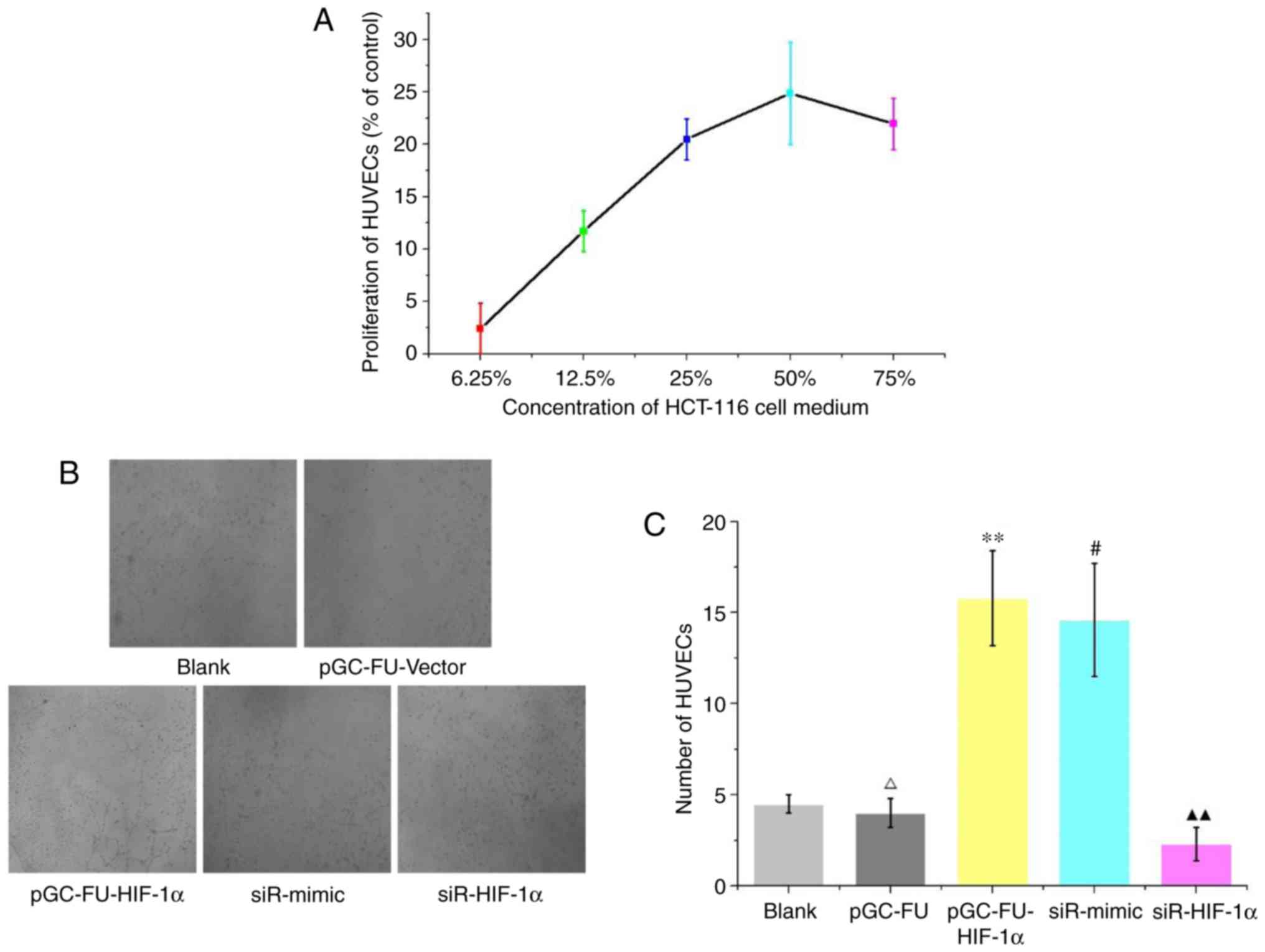
Effect of HIF-1α on tube formation in
HUVECs
Under normal conditions, HCT-116 cell culture medium
was used to prepare 6.25, 12.5, 25, 50 and 75% HCT-116 medium
conditioned ECM to simulate the growth environment of a tumor. The
media were used to culture HUVECs for 48 h. A CCK-8 assay was used
to determine the effect of ECM on the proliferation of HUVECs. The
results revealed that excluding the 6.25% medium, proliferation of
HUVECs was significantly increased when conditioned medium was
added. Among the various percentages of conditioned media used, the
50% media resulted in the largest increase in proliferation
(P<0.01; Fig. 3A). ECM gel was
used to assess the effects of HIF-1α on tube formation. The results
revealed that in the cells grown in medium from HIF-1α
overexpression cells (pGC-FU-HIF-1α), tube formation was
significantly increased compared with cells grown in medium from
either the blank group or pGC-FU-vector group (P<0.01).
Furthermore, medium from the siR-HIF-1α group significantly
decreased tube formation in HUVECs, and the difference was
statistically significant (P<0.05; Fig. 3B and C).
Tan IIA decreases proliferation of
HCT-116 cells
Under both normal and hypoxic conditions, Tan IIA at
different concentrations was added to the HCT-116 cells for 24, 48
and 72 h. The results revealed that Tan IIA decreased proliferation
of HCT-116 cells in a concentration and time-dependent manner
(Fig. 4A and B). Under normal
conditions, the IC50 of Tan IIA on HCT-116 cells for 24,
48 and 72 h was 30.7±1.52, 13.3±0.56 and 5.1±0.55 µM, respectively.
Under hypoxic conditions, the IC50 was 8.4±1.09,
2.1±0.18 and 0.99±0.17 µM, respectively (Fig. 4C and D). Therefore, Tan IIA
exhibited a more notable effect on proliferation under hypoxic
conditions.
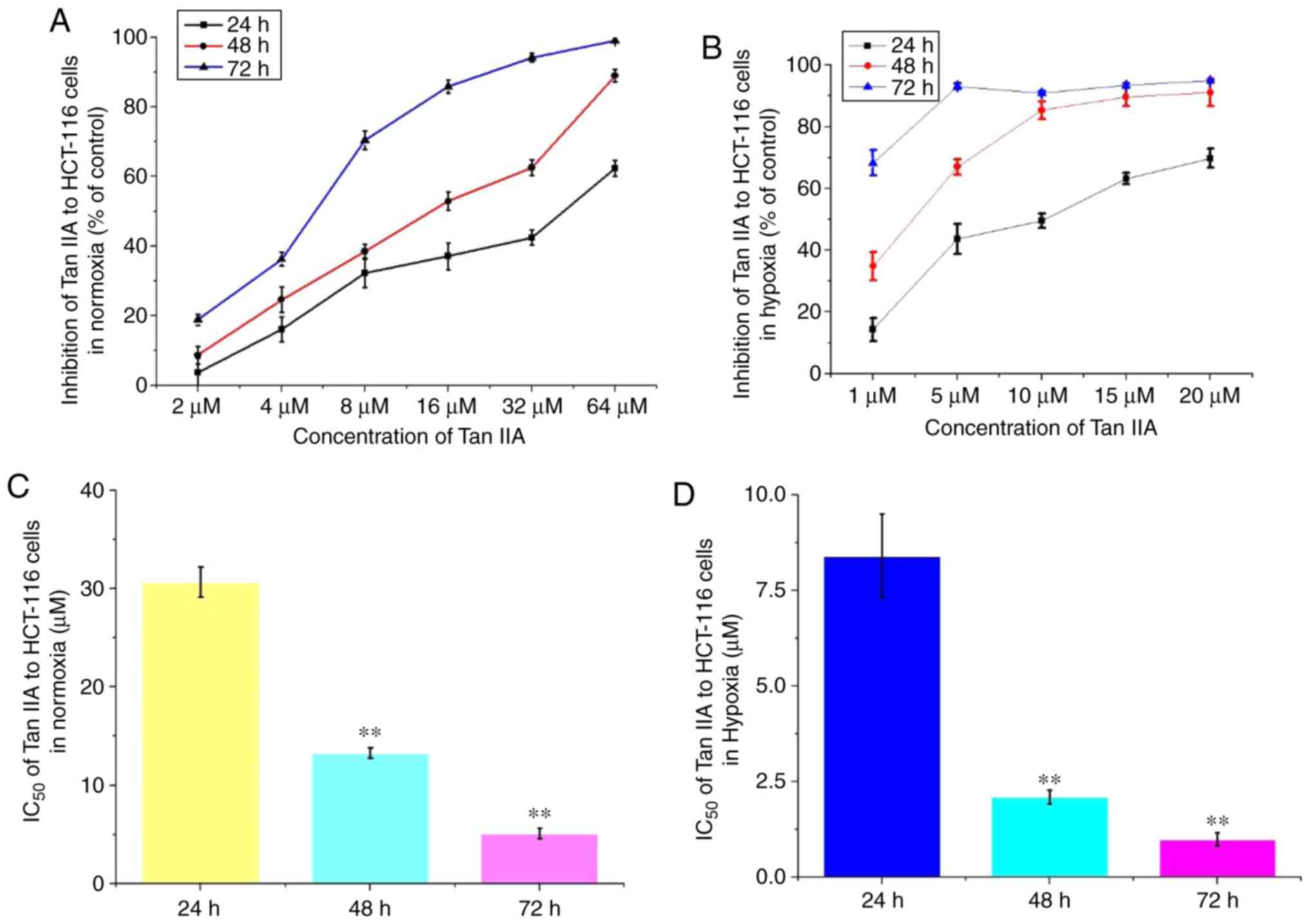 | Figure 4.Effects of Tan IIA on the
proliferation of HCT-116 under hypoxic conditions. The effects of
Tan IIA on proliferation of HCT-116 cell proliferation was analyzed
using a Cell Counting Kit-8 assay under (A) normal and (B) hypoxic
conditions. Tan IIA was used at a concentration of 2, 4, 8, 16, 32
and 64 µM under normal conditions, and 1, 5, 10, 15, 20 and 25 µM
under hypoxic. IC50 was analyzed in cells treated for
24, 48 and 72 h under (C) normal and (D) hypoxic conditions.
**P<0.01 vs. 24 h. Tan IIA, tanshinone IIA. |
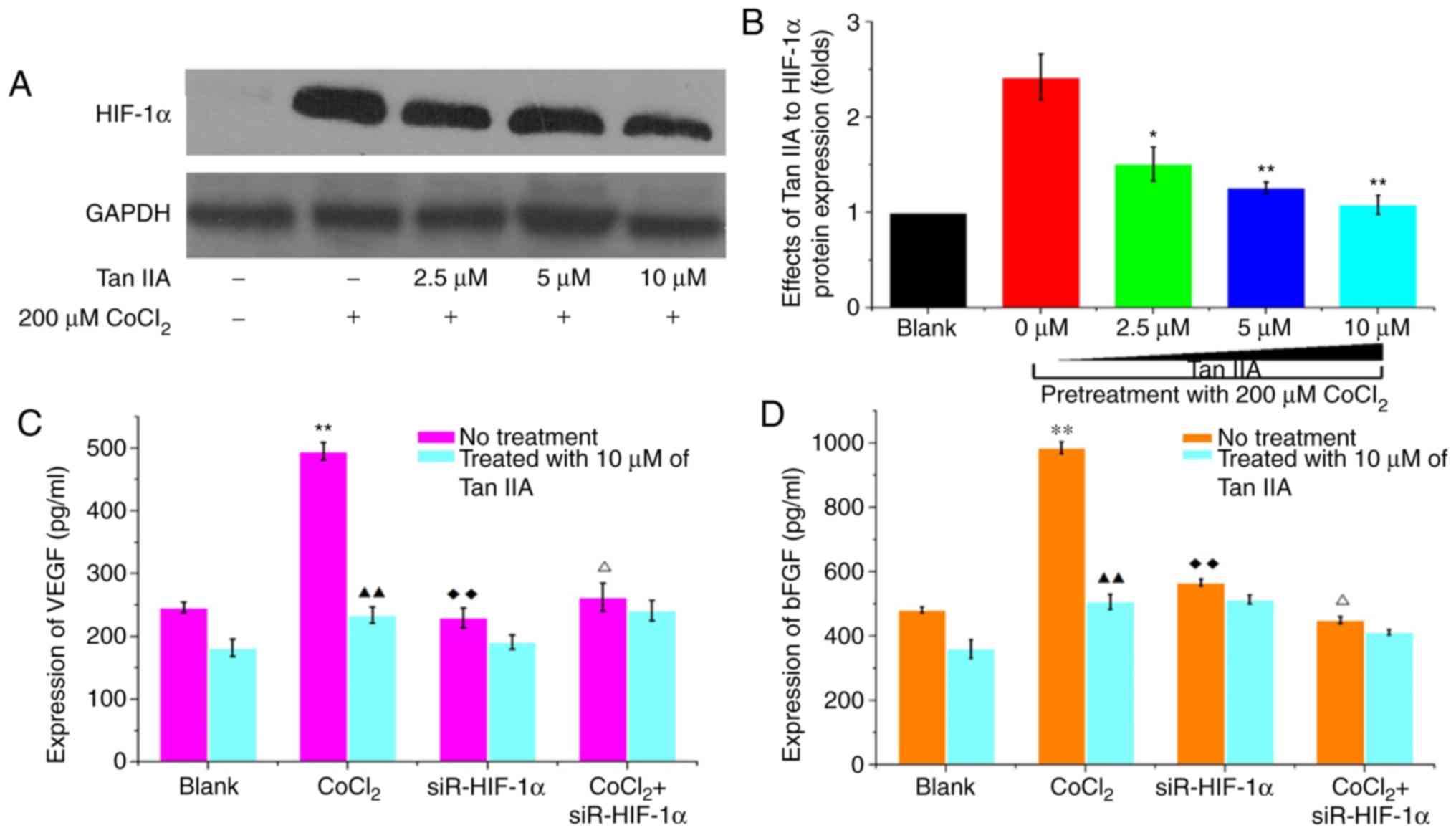
Tan IIA decreases the expression of
pro-angiogenic factors under hypoxic conditions
Western blotting and flow cytometry revealed that
Tan IIA can significantly decreased HIF-1α expression in HCT-116
under hypoxic conditions in a concentration dependent manner
(P<0.01). Under hypoxic conditions, 10 µM Tan IIA almost
completely abrogated CoCl2-induced HIF-1α upregulation
(P=0.489; Fig. 5A and B).
Similarly, 10 µM Tan IIA significantly inhibited secretion of VEGF
and bFGF in HCT-116 cells under normal conditions (P<0.05;
Fig. 5C and D). Furthermore, the
effects of Tan IIA on secretion of VEGF and bFGF were notably
greater under hypoxic conditions; 10 µM of Tan IIA could completely
abrogate the CoCl2-induced effects on HIF-1α expression
(P<0.01; Fig. 5C and D).
siR-HIF-1α decreased the expression of VEGF and bFGF in cells grown
under hypoxic conditions to levels similar to that observed under
normal conditions. However, siR-HIF1α did not affect the expression
of VEGF and bFGF under normal conditions (Fig. 5C and D). Tan IIA, significantly
decreased the expression levels of VEGF and bFGF in
HIF-1α-overexpressing and knockdown cells, (Fig. 5C and D). The results indicated that
Tan IIA decreased VEGF and bFGF expression in HCT-116 cells under
normal and hypoxic conditions.
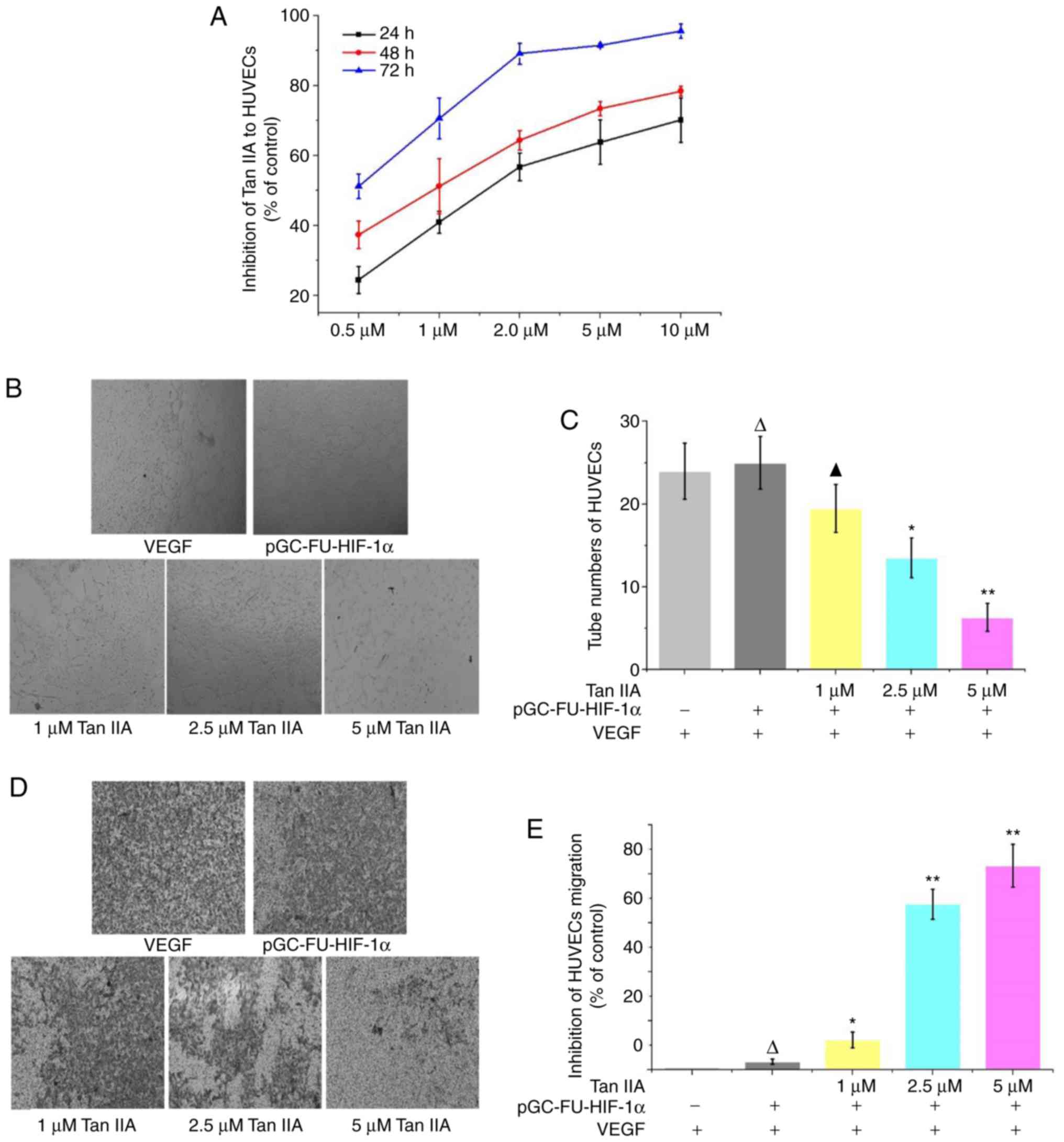
Inhibition of tube formation and
migration by Tan IIA under hypoxic conditions
A CCK-8 assay revealed that in HUVECs treated with
0.5, 1, 2, 5 and 10 µM Tan IIA HUVECs for 24, 48 and 72 h,
proliferation was decreased in a time and concentration dependent
manner compared with the control group (Fig. 6A). A tube formation assay revealed
that Tan IIA inhibited tube formation in HUVECs in a dose-dependent
manner, and 1 µM Tan IIA was sufficient to statistically reduce
tube formation (P<0.01; Fig. 6B and
C). A Transwell migration assay revealed that migration was
decreased in cells treated with Tan IIA, and 1 µM Tan IIA was
sufficient to statistically decrease migration. Tan IIA-mediated
inhibition of migration was determined to be
concentration-dependent (Fig. 6D and
E).
Discussion
Tumor angiogenesis serves an important role in the
growth, invasion, metastasis and recurrence of CRC. The growth of
tumors can be divided into two phases; an avascular phase and an
angiogenesis phase (13). Tumors in
the avascular phase are in a state of dormancy; whereas in the
angiogenesis phase, tumor cells secrete large quantities of
pro-angiogenic factors, resulting in rapid growth of tumors. Recent
studies demonstrated that a hypoxic microenvironment is the primary
factor facilitating the transition of tumors from the avascular
phase to the angiogenesis phase (14).
Hypoxia activates a series of pro-angiogenic factors
through the aberrant expression of HIF-1α such that tumors
transition to the angiogenesis phase (15,16).
Studies have identified numerous signaling molecules which
participate in the regulation of tumor angiogenesis (17,18).
Among these, VEGF and bFGF are deemed to be the most important
angiogenesis factors, and they can promote migration and
proliferation of endothelial cells. These factors are closely
associated with the growth of tumors and angiogenesis (18,19).
VEGF induces proliferation of endothelial cells in vitro,
and stimulates chemotaxis of endothelial cell angiogenesis
(20,21). bFGF promotes endothelial cell
proliferation, stimulates chemotaxis of endothelial cells towards
tumor tissues and upregulates production of a variety of proteases.
bFGF also induces collagen invasion, formation of vessel-like
structures and increases the levels of plasminogen activity factor
in tissues (22,23).
Hypoxia is a common pathological characteristic
observed in solid tumor tissues such as CRC. In poorly
differentiated solid tumors, rapid growth accompanied by a lack of
adequate vasculature, results in an insufficient blood supply and
thus potential lymphatic system disorders. In hypoxia, a number of
genes are upregulated in tumor cells in response to hypoxia. HIF-1α
associates with hypoxia response element (HRE), forming the
response element binding protein and activating the formation of
complexes of other transcription factors. HIF-1α has been revealed
to upregulate expression of VEGF and bFGF (24,25),
and upregulation of HIF-1α in CRC tissues during hypoxic conditions
enabled the survival of tumor cells and potentiated evolution of
tumors (26,27). Proliferation of endothelial cells
underlies the formation of blood vessels and thus is required for
angiogenesis, both physiologically and pathophysiologically in
tumor tissues. Hypoxia can induce the formation of a large number
of new blood vessels through proliferation of endothelial cells.
Specifically, hypoxia results in upregulation of HIF-1α, increasing
VEGF expression, which promotes angiogenesis.
In cell experiments, the use of CoCl2 to
simulate hypoxia and hypoxia incubators are both effective methods
to induce hypoxia, and cells treated with 100 µM (28), 200 µM (29) or 300 µM (30) CoCl2 are often used as
controls. Therefore, 200 µM CoCl2 was used in this
experiment to simulate hypoxia, and this method could effectively
induce hypoxia. A hypoxic cell incubator, as another method to
simulate hypoxia, cannot be the limitation of the use of
CoCl2 in this study.
The promoter region of VEGF and the regulating gene
region of bFGF contains an HRE, and HIF-1α can bind to the response
element, thus directly regulating the expression of VEGF and bFGF
at the genetic level (31,32). In the present study, ELISA was used
to determine the effects of regulating HIF-1α expression on the
secretion of VEGF and bFGF in the supernatant of CRC cells. HIF-1α
significantly increased the expression levels of VEGF and bFGF.
However, during normal conditions, targeted interference of HIF-1α
did not result in a decrease of VEGF expression. In
CoCl2-simulated hypoxia, VEGF expression was
significantly reduced. Additionally, bFGF expression was regulated
by HIF-1α. Both under normal conditions and
CoCl2-simulated hypoxia, silencing the HIF-1α gene
resulted in a decrease in bFGF expression. These findings indicated
that upregulation and interference of HIF-1α expression regulated
VEGF and bFGF expression in HCT-116 cells, and may therefore
regulate angiogenesis in CRC.
HUVEC tube formation, clonogenic assays and BrdU
assays are commonly used experimental methods, among which HUVEC
tube formation is a classical method for angiogenesis research
(33). In the present study, this
method was applied as a functional research method for Tan IIA to
inhibit angiogenesis in colorectal cancer. Interactions between
tumor cells and endothelial cells result in tumor angiogenesis,
tumor cells recruit endothelial cells and endothelial cells of the
blood vessels stimulate growth and proliferation of tumor cells
through paracrine effects (34).
The present study demonstrated that HIF-1α overexpression
significantly increased proliferation of HUVECs. Tube formation was
increased in HUVECs when HIF-1α was overexpressed compared to cells
where HIF-1α was knocked down. However, in cells where HIF-1α was
knocked down tube formation was further reduced in hypoxic
conditions.
Tan IIA exhibits antineoplastic activity in a
variety of malignancies, such as hepatic (35), gastric (36) and colorectal cancer (37), although the underlying mechanism has
not been determined. In our previous studies (10,11),
high-doses of Tan IIA significantly inhibited proliferation in CRC
cells and decreased VEGF expression through regulation of COX-2
expression. This suggests that Tan IIA may efficiently reduce
expression of angiogenesis-promoting factors, and this may underlie
its antineoplastic effects. In the present study, Tan IIA
significantly decreased proliferation of HCT-116 cells under
hypoxic conditions in a time- and concentration-dependent manner.
Tan IIA also significantly decreased expression of HIF-1α induced
by hypoxia. Therefore, based on these results it was hypothesized
that Tan IIA may decrease the expression of HIF-1α in HCT-116 cells
under hypoxic conditions. ELISA revealed that Tan IIA decreased the
expression levels of VEGF and bFGF in the HCT-116 cells.
Additionally, the effects of Tan IIA on VEGF and bFGF secretion
were more prominent in cells in which hypoxia was induced using 200
µM CoCl2. Treatment with 10 µM Tan IIA reversed
upregulation of VEGF and bFGF induced by CoCl2. In cells
where HIF-1α was silenced, the effects of Tan IIA on reduction of
VEGF and bFGF were increased.
The anti-proliferative effects of Tan IIA on HUVECs
revealed that Tan IIA effectively inhibited proliferation of
HUVECs. The median inhibitory concentration at 24, 48 and 72 h was
1.65, 0.99 and 0.36 µM, respectively. Tube formation and migration
assays with HUVECs revealed that 1 µM of Tan IIA inhibited tube
formation and metastasis in HUVECs. This suggests that Tan IIA
directly inhibited proliferation, tube formation and metastasis of
endothelial cells, and may prevent angiogenesis in CRC. The present
study demonstrated that Tan IIA inhibited secretion of
pro-angiogenic factors under hypoxic conditions through
downregulation of HIF-1α expression and also directly inhibited
proliferation, tube formation and metastasis of HUVECs. This
suggests that HIF-1α and its effects on angiogenesis are potential
targets for preventing progression and metastasis of CRC.
In summary, HIF-1α may serve as a potential target
in CRC and Tan IIA may be used to inhibit angiogenesis in CRC cells
under normal and hypoxic conditions by modulating the tumor hypoxia
microenvironment.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81673784, 81673783,
81874405, 81973651 and 81520108031), The Science Foundation for
Shanghai Committee of Science Project (grant nos. 18QA1404200 and
17QA1404100), The Shanghai Health Bureau Science Foundation [grant
no. ZY(2018–2020)-FWTX-4026], the Shang Youth Medical Talents
Training Program (grant no. 2017YQ077) and the Xinglin Young
Scholar, Shanghai University of Traditional Chinese Medicine.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the author on reasonable request.
Authors' contributions
YL, QL, and BZ designed the study. LZ and HS
performed the majority of the cell-based experiments with help from
TW, RJ, ZZ, YF, NL, JF, YW and QJ. BZ and NL performed the data
management and statistical analysis. LZ, HS, QL and YL wrote the
manuscript. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global Cancer Statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Deenen MJ, Meulendijks D, Cats A,
Sechterberger MK, Severens JL, Boot H, Smits PH, Rosing H,
Mandigers CM, Soesan M, et al: Upfront genotyping of DPYD*2A to
individualize fluoropyrimidine therapy: A safety and cost analysis.
J Clin Oncol. 34:227–234. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yamano T, Yoshimura M, Kobayashi M, Beppu
N, Hamanaka M, Babaya A, Tsukamoto K, Noda M, Matsubara N and
Tomita N: Malnutrition in rectal cancer patients receiving
preoperative chemoradiotherapy is common and associated with
treatment tolerability and anastomotic leakage. Int J Colorectal
Dis. 31:877–884. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ramjiawan RR, Griffioen AW and Duda DG:
Anti-angiogenesis for cancer revisited: Is there a role for
combinations with immunotherapy? Angiogenesis. 20:185–204. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Viallard C and Larrivée B: Tumor
angiogenesis and vascular normalization: Alternative therapeutic
targets. Angiogenesis. 20:409–426. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhu H, Li X, Yuan M, Wan W, Hu M, Wang X
and Jiang X: Intramyocardial delivery of bFGF with a biodegradable
and thermosensitive hydrogel improves angiogenesis and
cardio-protection in infarcted myocardium. Exp Ther Med.
14:3609–3615. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zeng M, Shen J, Liu Y, Lu LY, Ding K,
Fortmann SD, Khan M, Wang J, Hackett SF, Semenza GL and Campochiaro
PA: The HIF-1 antagonist acriflavine: Visualization in retina and
suppression of ocular neovascularization. J Mol Med (Berl).
95:417–429. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Oh ET, Kim CW, Kim HG, Lee JS and Park HJ:
Brusatol-mediated inhibition of c-Myc increases HIF-1α degradation
and causes cell death in colorectal cancer under hypoxia.
Theranostics. 7:3415–3431. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kasai M, Van Damme N, Berardi G, Geboes K,
Laurent S and Troisi RI: The inflammatory response to stress and
angiogenesis in liver resection for colorectal liver metastases: A
randomized controlled trial comparing open versus laparoscopic
approach. Acta Chir Belg. 118:172–180. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sui H, Zhao J, Zhou L, Wen H, Deng W, Li
C, Ji Q, Liu X, Feng Y, Chai N, et al: Tanshinone IIA inhibits
β-catenin/VEGF-mediated angiogenesis by targeting TGF-β1 in
normoxic and HIF-1α in hypoxic microenvironments in human
colorectal cancer. Cancer Lett. 403:86–97. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou LH, Hu Q, Sui H, Ci SJ, Wang Y, Liu
X, Liu NN, Yin PH, Qin JM and Li Q: Tanshinone II-a inhibits
angiogenesis through down regulation of COX-2 in human colorectal
cancer. Asian Pac J Cancer Prev. 13:4453–4458. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Salavati H, Soltani M and Amanpour S: The
pivotal role of angiogenesis in a multi-scale modeling of tumor
growth exhibiting the avascular and vascular phases. Microvasc Res.
119:105–116. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang WC, Chen SH, Chiang WH, Huang CW, Lo
CL, Chern CS and Chiu HC: Tumor microenvironment-responsive
nanoparticle delivery of chemotherapy for enhanced selective
cellular uptake and transportation within tumor. Biomacromolecules.
17:3883–3892. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lequeux A, Noman MZ, Xiao M, Sauvage D,
Van Moer K, Viry E, Bocci I, Hasmim M, Bosseler M, Berchem G and
Janji B: Impact of hypoxic tumor microenvironment and tumor cell
plasticity on the expression of immune checkpoints. Cancer Lett.
458:13–20. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Belluco C, Forlin M, Delrio P, Rega D,
Degiuli M, Sofia S, Olivieri M, Pucciarelli S, Zuin M, De Manzoni
G, et al: Elevated platelet count is a negative predictive and
prognostic marker in locally advanced rectal cancer undergoing
neoadjuvant chemoradiation: A retrospective multi-institutional
study on 965 patients. BMC Cancer. 18:10942018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pastorekova S and Gillies RJ: The role of
carbonic anhydrase IX in cancer development: Links to hypoxia,
acidosis, and beyond. Cancer Metastasis Rev. 38:65–77. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Broggini T, Wüstner M, Harms C, Stange L,
Blaes J, Thomé C, Harms U, Mueller S, Weiler M, Wick W, et al:
NDRG1 overexpressing gliomas are characterized by reduced tumor
vascularization and resistance to antiangiogenic treatment. Cancer
Lett. 380:568–576. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhu CC, Chen C, Xu ZQ, Zhao JK, Ou BC, Sun
J, Zheng MH, Zong YP and Lu AG: CCR6 promotes tumor angiogenesis
via the AKT/NF-κB/VEGF pathway in colorectal cancer. Biochim
Biophys Acta Mol Basis Dis. 1864:387–397. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Barravecchia I, Mariotti S, Pucci A,
Scebba F, De Cesari C, Bicciato S, Tagliafico E, Tenedini E,
Vindigni C, Cecchini M, et al: MICAL2 is expressed in cancer
associated neo-angiogenic capillary endothelia and it is required
for endothelial cell viability, motility and VEGF response. Biochim
Biophys Acta Mol Basis Dis. 1865:2111–2124. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu J, Liu M, Yu M, Shen J, Zhou J, Hu J,
Zhou Y and Zhang W: RasGRP1 is a target for VEGF to induce
angiogenesis and involved in the endothelial-protective effects of
metformin under high glucose in HUVECs. IUBMB Life. 71:1391–1400.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang HJ and Lo WY: Identification of basic
fibroblast growth factor as the dominant protector of laminar shear
medium from the modified shear device in tumor necrosis factor-α
induced endothelial dysfunction. Front Physiol. 8:10952017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Han JH, Hwang AR, Nam DH, Kim S, Choi HC,
Lim JH and Woo CH: ERK5 regulates basic fibroblast growth
factor-induced type 1 plasminogen activator inhibitor expression
and cell proliferation in lung fibroblasts. Life Sci. 135:1–8.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Meng L, Cheng Y, Tong X, Gan S, Ding Y,
Zhang Y, Wang C, Xu L, Zhu Y, Wu J, et al: Tumor oxygenation and
hypoxia inducible factor-1 functional inhibition via a reactive
oxygen species responsive nanoplatform for enhancing radiation
therapy and abscopal effects. ACS Nano. 12:8308–8322. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Deng Y, Huang G, Chen F, Testroet ED, Li
H, Li H, Nong T, Yang X, Cui J, Shi D and Yang S: Hypoxia enhances
buffalo adipose-derived mesenchymal stem cells proliferation,
stemness, and reprogramming into induced pluripotent stem cells. J
Cell Physiol. 234:17254–17268. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Han J, Jackson D, Holm J, Turner K,
Ashcraft P, Wang X, Cook B, Arning E, Genta RM, Venuprasad K, et
al: Elevated d-2-hydroxyglutarate during colitis drives progression
to colorectal cancer. Proc Natl Acad Sci USA. 115:1057–1062. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ni T, He Z, Dai Y, Yao J, Guo Q and Wei L:
Oroxylin A suppresses the development and growth of colorectal
cancer through reprogram of HIF1α-modulated fatty acid metabolism.
Cell Death Dis. 8:e28652017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bharti R, Dey G, Das AK and Mandal M:
Differential expression of IL-6/IL-6R and MAO-A regulates
invasion/angiogenesis in breast cancer. Br J Cancer. 118:1442–1452.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu Y, Xu S, Zu T, Li F, Sang S, Liu C, An
Y, Mi B, Orgill DP, Murphy GF and Lian CG: Reversal of TET-mediated
5-hmC loss in hypoxic fibroblasts by ascorbic acid. Lab Invest.
99:1193–1202. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Maliha AM, Kuehn S, Hurst J, Herms F, Fehr
M, Bartz-Schmidt KU, Dick HB, Joachim SC and Schnichels S:
Diminished apoptosis in hypoxic porcine retina explant cultures
through hypothermia. Sci Rep. 9:48982019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kobayashi M, Morinibu A, Koyasu S, Goto Y,
Hiraoka M and Harada H: A circadian clock gene, PER2, activates
HIF-1 as an effector molecule for recruitment of HIF-1α to promoter
regions of its downstream genes. FEBS J. 284:3804–3816. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fu X, Zhai S and Yuan J: Interleukin-6
(IL-6) triggers the malignancy of hemangioma cells via activation
of HIF-1α/VEGFA signals. Eur J Pharmacol. 841:82–89. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang M, Qiu L, Zhang Y, Xu D, Zheng JC
and Jiang L: CXCL12 enhances angiogenesis through CXCR7 activation
in human umbilical vein endothelial cells. Sci Rep. 7:82892017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Batlle R, Andrés E, Gonzalez L, Llonch E,
Igea A, Gutierrez-Prat N, Berenguer-Llergo A and Nebreda AR:
Regulation of tumor angiogenesis and mesenchymal-endothelial
transition by p38α through TGF-β and JNK signaling. Nat Commun.
10:30712019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ren X, Wang C, Xie B, Hu L, Chai H, Ding
L, Tang L, Xia Y and Dou X: Tanshinone IIA induced cell death via
miR30b-p53-PTPN11/SHP2 signaling pathway in human hepatocellular
carcinoma cells. Eur J Pharmacol. 796:233–241. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xu Z, Chen L, Xiao Z, Zhu Y, Jiang H, Jin
Y, Gu C, Wu Y, Wang L, Zhang W, et al: Potentiation of the
anticancer effect of doxorubicinin drug-resistant gastric cancer
cells by tanshinone IIA. Phytomedicine. 51:58–67. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Qian J, Fang D, Lu H, Cao Y, Zhang J, Ding
R, Li L and Huo J: Tanshinone IIA promotes IL2-mediated SW480
colorectal cancer cell apoptosis by triggering INF2-related
mitochondrial fission and activating the Mst1-Hippo pathway. Biomed
Pharmacother. 108:1658–1669. 2018. View Article : Google Scholar : PubMed/NCBI
|