Introduction
Prostate cancer (PCa) has accounted for the largest
number of estimated new cases of cancer among men (2018–2020) in
the United States, and the number of mortalities was the second
highest, accounting for 10% of all mortalities (1–3).
Globally, the number of diagnosed cases of PCa in men is the
highest among all cancer types in 109 countries (4). Although therapeutic strategies have
improved over time, the efficiency remains far from optimal.
Researchers have focused on the regulatory role of protein-coding
RNAs. Recently, numerous non-coding RNAs (ncRNAs), including long
ncRNAs (lncRNAs), have been revealed to be involved in PCa
progression (5,6). Therefore, there is an urgent need to
identify the underlying mechanisms of these regulatory transcripts
to fully understand PCa carcinogenesis in order to develop novel
diagnostic targets and treatment strategies.
lncRNAs are a subset of ncRNAs with a transcription
length of >200 nucleotides, and have limited or no protein
coding ability (7). Moreover,
lncRNAs exhibit both the highest abundance and complexity among
non-coding transcripts (8).
Numerous studies have reported that lncRNAs achieve their
regulatory role by regulating gene expression at the
transcriptional or post-transcriptional level (5,6,9).
Salmena et al (10) proposed
a novel regulatory mechanism, in which lncRNAs regulate mRNA
expression by competing with microRNAs (miRNAs/miRs), forming a
competing endogenous RNA (ceRNA) network. A large number of ceRNA
regulatory networks are involved in cancer development. For
example, lnc-TALC promotes O-6-methylguanine-DNA methyltransferase
expression and temozolomide resistance by regulating the
c-Met/STAT3/p300 axis via sponging of miR-20b-3p in glioblastoma
(11). lncRNA-PAGBC contributes to
gallbladder cancer development by regulating miR-511 as a ceRNA
(12), while LINC81507 acts as
tumor suppressor by competing with miR-199b-5p to regulate caveolin
1 expression and STAT3 activation in non-small cell lung cancer
(NSCLC) (13). In PCa, several
lncRNAs have been demonstrated to contribute to tumorigenesis as
ceRNAs. For instance, the highly specific lncRNA prostate cancer
associated 3 (PCA3) exerts an oncogenic effect by competitively
binding with miR-218-5p and promoting high mobility group box 1
expression in PCa (14). It has
also been revealed that forkhead box P4 (FOXP4)-antisense RNA 1
regulates FOXP4 at the post-transcriptional level by sponging
miR-3184-5p to promote tumorigenesis (15), while lncRNA small nucleolar RNA host
gene 12 promotes cancer cell proliferation and metastasis by
sponging miR-133b (16). Therefore,
it was hypothesized that other lncRNAs may also be involved in PCa
progression by acting as ceRNAs.
Our previous study identified that lncRNA AC245100.4
(ENSG00000231551 or RP11-495P10.1) was closely associated with PCa
(17). However, whether it had
clinical significance, the exact role it served in cancer
progression and its mechanism of action remain to be revealed in
PCa. Therefore, it is necessary to investigate the relationship
between AC245100.4 and PCa. In the present study, the expression of
AC245100.4 in patients with PCa was examined via bioinformatics
prediction. Cell proliferation assay, Transwell migration assay,
wound healing and subcutaneous models were used for assessing the
role of AC245100.4 in PCa. Moreover, the underlying mechanisms were
evaluated.
Materials and methods
Cell source and culture condition
The human normal prostate epithelium cell line
(RWPE1), PCa cell lines (DU145 and PC3) and 293T cells were all
purchased from The Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences. RWPE1 cells were cultured in
keratinocyte-serum-free medium containing 0.05 mg/ml bovine
pituitary extract and 5 ng/ml epidermal growth factor (all from
Gibco; Thermo Fisher Scientific, Inc.). DU145 cells were maintained
in RPMI 1640 medium (Invitrogen; Thermo Fisher Scientific, Inc.),
while PC3 and 293T cells were cultured in DMEM (Invitrogen; Thermo
Fisher Scientific, Inc.). All culture media were supplemented with
10% FBS (Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin (Sigma Aldrich; Merck KGaA). All cells were
maintained in a humidified atmosphere at 37°C with 5%
CO2.
Bioinformatics analysis
The different expression of AC245100.4 in prostate
cancer and healthy tissues were analyzed via Gene Expression
Profiling Interactive Analysis (GEPIA; version: 1.0; http://gepia.cancer-pku.cn/), using data from The
Cancer Genome Atlas (TCGA) database (18). The data included 492 PCa tissues
(tumor) and 52 adjacent para-cancerous tissues (healthy). Relative
analysis values were selected as follows: Gene=ENSG00000231551 or
RP11-495P10.1; |log2 FC| cutoff=0.05; P-value cutoff=0.05;
datasets=Prostate adenocarcinoma and match TCGA normal data.
Gene transfection in vitro
The overexpression pcDNA3.1- AC245100.4 and control
pcDNA3.1 recombinant plasmids were constructed by Shanghai
GenePharma Co., Ltd. Small interfering RNAs [si-AC245100.4 and
si-negative control (NC)] and miRNAs [(mimics-miR-145-5p, mimic-NC;
cat. no. miR10000437) and (inhibitors-miR-145-5p and inhibitor-NC;
cat. no. miR20000437)] were synthesized by Guangzhou RiboBio Co.,
Ltd. The siRNA sequences were as follows: si-NC,
5′-UUCUCCGAACGUGUCACGUTT-3′; si-AC245100.4-#1,
5′-UCAGAUCAGAACUAGCUAUTT-3′; si-AC245100.4-#2,
5′-GGAUUGUAUAUGUCCUCUUTT-3′; and si-AC245100.4-#3,
5′-GUCCAGCUGUAUAAUGAAATT-3′. Lipofectamine® 2000 (Thermo
Fisher Scientific, Inc.) was used for gene transfection following
the manufacturer's instructions. Cells were transfected with 100 nM
miRNAs (miR-145-5p mimics or inhibitors), 50 nM siRNA
(si-AC245100.4 or si-NC) or 2 µg plasmid for 24 h in 6-well plates.
Transfection procedures were conducted at room temperature. Cell
proliferation assay were performed 24 h post-transfection. Reverse
transcription-quantitative (RT-q) PCR, RNA immunoprecipitation
(RIP) assay, scratch wound healing assay and Transwell migration
assay were conducted 48 h post-transfection. Western blotting was
performed 72 h post-transfection.
RT-qPCR
Total RNA was isolated using RNAiso Plus reagent
(Takara Bio, Inc.) according to the manufacturer's protocol. RNA
concentration and purity were detected using a NanoDrop 2000
(Thermo Fisher Scientific, Inc.). Subsequently, 1 µg total RNA was
transcribed to cDNA using the PrimeScript™ RT reagent kit (Takara
Bio, Inc.). RT reaction were two steps: gDNA eraser at 42°C for 2
min, reverse transcription at 37°C for 15 min and 85°C for 5 sec.
RT-qPCR was performed according to the manufacturer's protocol
using SYBR Premix Ex Taq II (Takara Bio, Inc.). Reaction conditions
were set as: Initial denaturation at 95°C for 1 min, followed by 40
cycles of 95°C for 5 sec and 60°C for 10 sec. Relative RNA
expression was analyzed based on the 2−ΔΔCq method and
GAPDH was used as an internal control for the ΔCq of specific genes
(19). The primers used were as
follows: GAPDH forward, 5′-GAGTCAACGGATTTGGTCGT-3′ and reverse,
5′-GACAAGCTTCCCGTTCTCAG-3′; RBBP5 forward,
5′-AGACAAAAATTAGCTGGGCGTGG-3′ and reverse,
5′-GTGGTGCGATTCTCCACTCACT-3′; and AC245100.4 forward,
5′-AGCAATGCCTTCCTCTTTGA-3′ and reverse, 5′-AAGGGTCTCCTTCAGGTGCT-3′.
The primers for miR-145-5p were provided along with the miRNAs
synthesized by Guangzhou RiboBio Co., Ltd. (cat. no.
miR10000437).
Western blotting
Whole cell lysates were obtained by lysing cells
using RIPA buffer (Beyotime Institute of Biotechnology) with
proteinase inhibitor (Sigma-Aldrich; Merck KGaA) on ice. The
concentrations of isolated proteins were detected using a BCA assay
kit (Beyotime Institute of Biotechnology). Equal amounts of whole
cell lysate (50 µg) were subjected to 10% SDS-PAGE. Subsequently,
proteins in the gel were transferred to PVDF membranes (EMD
Millipore). Following blocking with 5% non-fat milk for 2 h at room
temperature, the membranes were incubated with specific primary
antibodies against RBBP5 (cat. no. 13171; 1:1,000) or GAPDH (cat.
no. 5174; 1:1,000) at 4°C overnight. Subsequently, the membranes
were incubated with horseradish peroxidase-linked secondary
antibodies (cat. no. 7074; 1:3,000) for 1 h at room temperature.
All antibodies were obtained from Cell Signaling Technology, Inc.
The blots were visualized using the substrate BeyoECL Plus
(Beyotime Institute of Biotechnology) and imaged using a Tanon 5200
Chemiluminescent Imaging system (Tanon Science and Technology Co.,
Ltd.). The densitometry was conducted using ImageJ software,
version 1.50 (National Institutes of Health).
RIP assay
RIP was performed according to the manufacturer's
protocol of the Magna Nuclear RIP™ (Cross-Linked) Nuclear
RNA-Binding Protein Immunoprecipitation kit (EMD Millipore). PC3
cells (2×107) transfected with indicated genes were
harvested and lysed on ice. Crosslinking of protein and RNAs were
obtained by adding 37% formaldehyde at room temperature for 10 min.
Excess formaldehyde was removed using glycine and cell pellets were
centrifugated at 800 × g for 5 min at 4°C. Subsequently, cell
pellets were incubated with the Nuclei Isolation Buffer on ice for
15 min. RIP Cross-linked Lysis Buffer was then added to release the
cross-linked protein/RNAs. The lysate was sonicated with 5 pulses
at 50% power with 30 sec intervals between pulses using SM-250D
(Nanjing Shunma Technology Co., Ltd.). The tubes were cooled on ice
at all times. Specific argonaute RNA-induced silencing complex
(RISC) catalytic component 2 (AGO2; cat. no. MA5-14861; 1:10;
Thermo Fisher Scientific, Inc.) or IgG antibodies (cat. no. PP64B;
1:10; EMD Millipore) were incubated with Protein A/G magnetic beads
(supplied in the kit) at 4°C for ~3 h to obtain conjugated beads.
Mix the conjugated beads with sonicated lysis and incubate in 4°C.
The beads were washed with ice cold low Salt Wash Buffer after 3 h.
Protein in beads was removed using Proteinase K. Finally, the
existence of AC245100.4 or miR-145-5p in immunoprecipitated RNA was
analyzed by qPCR.
Cell proliferation assay
A Cell Counting Kit 8 (CCK8; Beyotime Institute of
Biotechnology) was used to evaluate cell proliferation according to
the manufacturer's instructions. DU145 or PC3 cells after gene
transfection with AC245100.4 or miR-145-5p were collected and
counted separately. Cells were seeded at a density of 5,000
cells/well in 96-well plates. Following incubation for 1-4 days
CCK8 solution was added in the dark at a volume of 10% of the cell
suspension. Subsequently, the plate was incubated at 37°C for
another 1 h. The absorbance at a wavelength of 450 nm was measured
using a microplate reader.
Cell migration analysis
Both scratch wound healing and Transwell migration
assays were used to evaluate the cell migratory ability as
previously described (20). For the
scratch wound healing assay, DU145 and PC3 cells were scratched
using a 200 µl tip at 48 h after gene transfection. Cells were
washed and cultured in serum-free medium after scratching. Images
of wound were captured with light microscope (magnification, ×200)
at 0 and 24 h post scratching. The blank area between cell
boundaries at 0 h (Area 0) and 24 h (Area 24) was quantitated using
ImageJ software, version 1.50 (National Institutes of Health). The
migration rates were calculated by the formula: [(Area 0)-(Area
24)]/(Area 0) ×100%.
For the Transwell migration assay, cells
(5×105 cells/ml) in 200 µl serum-free medium were seeded
in the top chambers of 8-µm pore size Transwell inserts (Corning
Inc.). A total of 600 µl medium supplemented with 20% FBS was added
to the bottom Transwell chamber. After 24 h of incubation at 37°C,
the cells on the membrane of the top inserts were fixed with 100%
ethanol at room temperature for 15 min and stained with 0.5%
crystal violet for 20 min at room temperature. Cell numbers were
counted in ≥6 random fields with light microscope at ×20
magnification for the Transwell migration assays.
Luciferase reporter assay
The binding sites between AC245100.4 and miR-145-5p
were predicted using the online DIANA-LncBase version: v2.0
(https://diana.e-ce.uth.gr/home), while
TargetScan version 7.2 (http://www.targetscan.org/vert_72/) and miRDB version
1 (http://www.mirdb.org/) were used to predict
binding sites between RBBP5 and miR-145-5p (21–23).
The wild-type (WT) plasmids were produced by inserting the
predicted binding site in the AC245100.4 sequence and the
3′untranslated region (3′UTR) of RBBP5 into the psi-CHECK-2 vector
(Promega Corporation). Simultaneously, the predicted binding site
was deleted to generate mutant (MUT) plasmids. 293T cells
(1×106) were co-transfected with the WT or MUT plasmids
(2 µg in 6-well plates) along with miR-145-5p mimic or control
using Lipofectamine® 2000 (Thermo Fisher Scientific,
Inc.) at room temperature. Following 48 h of incubation after
transfection, luciferase activity was evaluated using a
Dual-luciferase Reporter assay system (Promega Corporation). The
activity of Renilla luciferase was used for the
normalization of firefly luciferase activity.
Nude mouse PCa subcutaneous
models
All animal experiment protocol and procedure were
approved by the Animal Care and Use Committee of Harbin Medical
University. The pGPH1-short hairpin RNA (shRNA) plasmids to
knockdown AC245100.4 (sh-AC245100.4 or sh-NC) were constructed by
Shanghai GenePharma Co. Ltd. A total of 2 µg sh-AC245100.4 or sh-NC
plasmids were transfected to PC3 cells using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.) at
room temperature. Cells were incubated for another 48 h. Then, 2
mg/ml neomycin was added to PC3 cells after transfection with
sh-AC245100.4 or sh-NC for 2-3 weeks to generate a stable
AC245100.4-knockdown PC3 cell line. PCa xenograft models were
established via subcutaneous injection of PC3 cells
(1×106 cells in 100 µl PBS) stably transfected with
control shRNA or shRNA-AC245100.4 into male BALB/c nude mice
(Beijing Vital River Laboratory Animal Technology Co., Ltd. age,
4-6 weeks; weight, 20-30 g). Cells were subcutaneously injected to
the left medial upper limb of mice. Animals were housed at constant
temperature (20-26°C) and humidity (40-70%) with a 12 h dark-light
cycle and unrestricted access to food and water. A total of 15 mice
were purchased, of which three were excluded from the experiment as
one animal died before subcutaneous injection (autopsy did not
determine the cause of mortality) and two animals died during
anesthesia. A total of 12 mice left were randomly divided into two
groups (shRNA-NC or shRNA-AC245100.4; n=6 per group). The mice
weight and condition were monitored every 2 days. The humane
endpoints used to determine when animals should be euthanized were
as follows: Weight loss of 20-25% of the animal's original body
weight; loss of appetite for 24 h; severe infection; tumors growth
>10% of the animal's original body weight or the average
diameter >20 mm; and animals that are weak or dying. After 28
days of subcutaneous injection, all 12 tumor-bearing mice lived and
were euthanized via excessive anesthetized with an intraperitoneal
injection of 0.7% pentobarbital sodium (150 mg/kg). After the mice
died, which was confirmed by cardiac arrest and respiratory arrest
for >2 min, tumor samples were excised and weighed. Tumor length
and width were measured. Tumor volume was calculated using the
formula: V=(length × width2)/2.
Statistical analysis
Each experiment was performed ≥3 times. Data are
presented as the mean ± SD of ≥3 separate experiments. Differences
between two groups were compared using an unpaired two-tailed
Student's t-test, and one-way ANOVA followed by Tukey's post hoc
test was used to compare differences among multiple groups. The
differences in AC245100.4 expression from TCGA database were
analyzed using GEPIA (version: 1.0; http://gepia.cancer-pku.cn/) using the method one-way
ANOVA for differential analysis. Disease state (Tumor or Normal)
was the variable for calculating differential expression.
Statistical analysis was performed using GraphPad Prism version 7
(GraphPad Software, Inc.). P<0.05 was considered to indicate a
statistically significant difference.
Results
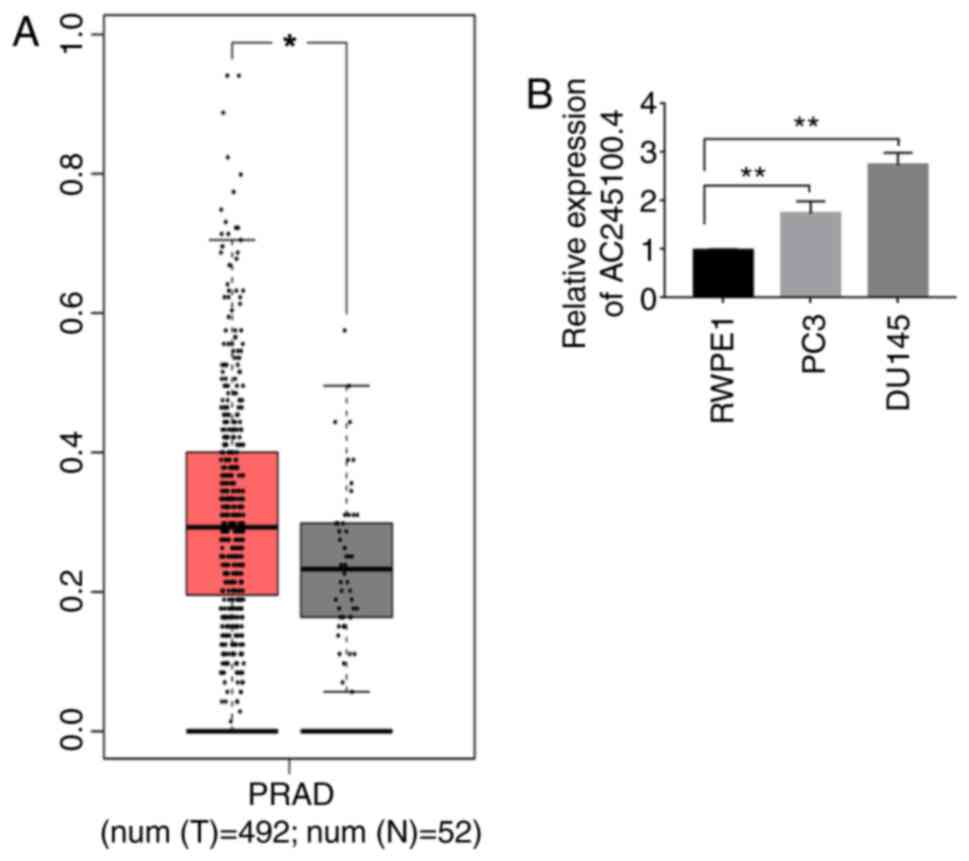
lncRNA AC245100.4 expression is
upregulated in PCa cells
Since AC245100.4 has rarely been investigated in
cancer, the present study first examined the association between
AC245100.4 and PCa. The differences in AC245100.4 expression were
analyzed by GEPIA using data from TCGA database (18). The data included 492 PCa tissues
(tumor) and 52 adjacent para-cancerous tissues (healthy). Relative
expression values of AC245100.4 were analyzed using |log2 fold
change| cut-off=0.05, P<0.05. The difference analysis (Fig. 1A) demonstrated that the expression
of AC245100.4 in PCa tissues was increased compared with those in
adjacent healthy tissues. Furthermore, the expression of AC245100.4
in PCa cell lines was examined using RT-qPCR. DU145 and PC3 cells
exhibited significantly higher expression levels of AC245100.4
compared with the normal prostate epithelium RWPE1 cell line
(Fig. 1B). The present results
suggested that AC245100.4 expression may be positively associated
with the development of PCa.
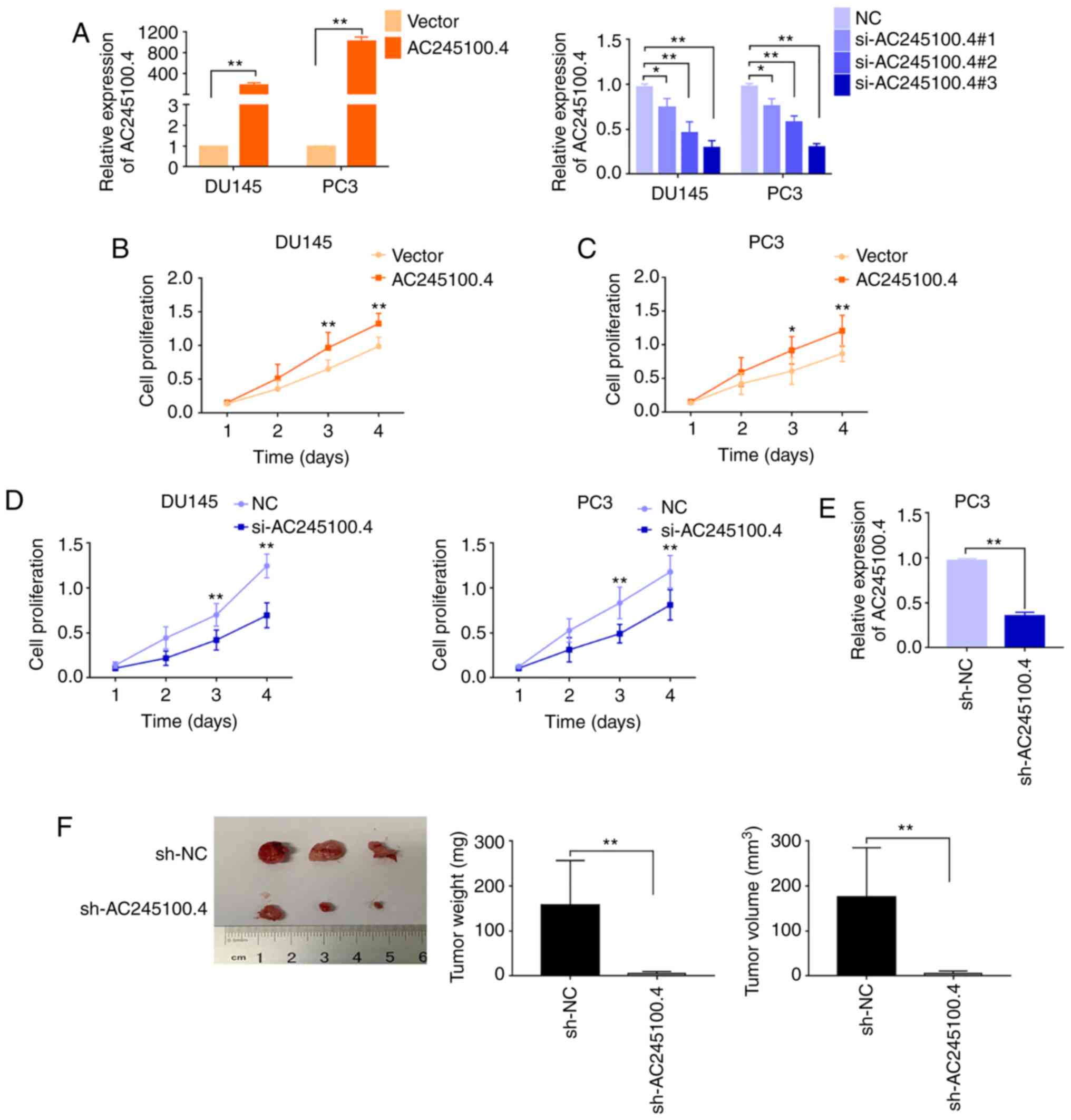
lncRNA AC245100.4 promotes PCa cell
proliferation
The present study investigated the effects of
AC245100.4 on cell proliferation in PCa. AC245100.4 expression in
DU145 and PC3 cells was altered by transfection with siRNA and
pcDNA3.1/AC245100.4 (Fig. 2A). The
RT-qPCR results demonstrated that AC245100.4 expression was
effectively regulated in DU145 and PC3 cells. siRNA#3 was selected
for the subsequent experiments due to its high efficiency. The CCK8
proliferation assay results demonstrated that AC245100.4 knockdown
decreased proliferation, whereas AC245100.4 overexpression
significantly increased proliferation in DU145 and PC3 cells
(Fig. 2B-D).
To further investigate the effects of AC245100.4
in vivo, a PC3 cell line with stable knockdown of AC245100.4
was constructed using shRNA (Fig.
2E). Animal models were established via subcutaneous injection
of control shRNA-treated or shRNA-AC245100.4-treated PC3 cells in
male nude mice. It was found that tumor volume and weight were
significantly decreased following AC245100.4 knockdown (Fig. 2F). These results suggested that
AC245100.4 may promote the proliferation of PCa cells.
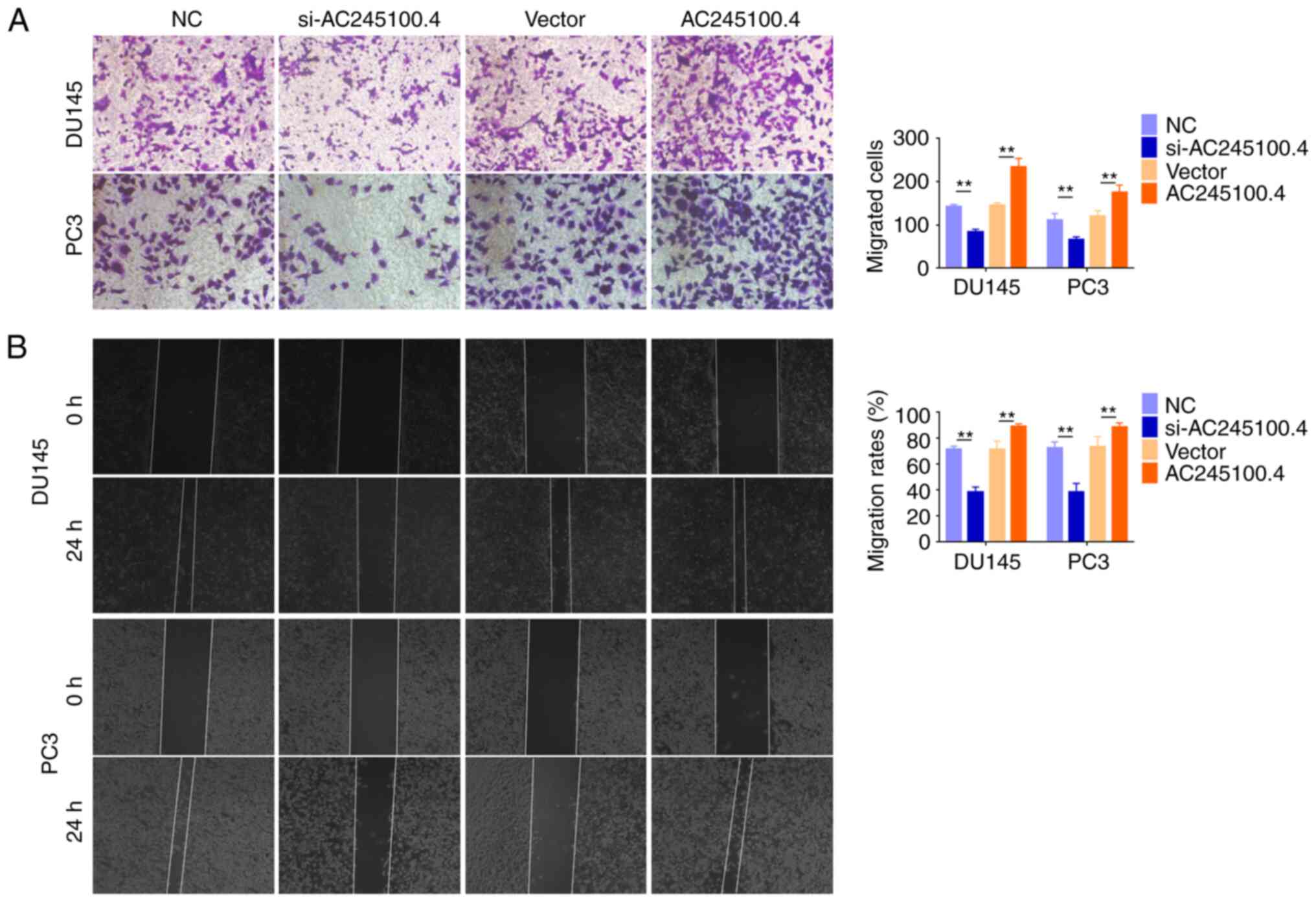
lncRNA AC245100.4 promotes PCa cell
migration in vitro
Cancer cell motility reflects the metastasis
potential of cancer (24).
Subsequently, the present study evaluated the migration abilities
of DU145 and PC3 cells following AC245100.4 knockdown or
overexpression. Transwell migration (Fig. 3A) and wound healing (Fig. 3B) assays indicated that AC245100.4
knockdown impaired the migration of PCa cells, while overexpression
of AC245100.4 significantly increased the migratory capacity of PCa
cells (Fig. 3). Based on its
promoting effects on proliferation and migration, it was concluded
that AC245100.4 served an oncogenic role in PCa.
lncRNA AC245100.4 targets miR-145-5p
in PCa cells
Since it was proposed that AC245100.4 may be an
oncogene, its mechanism of action was subsequently investigated. It
was identified that AC245100.4 was mostly located in the cytoplasm
(data not shown). It has been reported that lncRNAs may be involved
in post-transcriptional regulation of mRNAs by sponging miRNAs in
human cancer (6). Potential miRNA
binding sites in AC245100.4 were predicted using a human miRNA
target tool (DIANA Tools) (23).
Binding sites with miR-145-5p were observed (Fig. 4A). Therefore, the present study
targeted miR-145-5p for further investigation. Following AC245100.4
knockdown, miR-145-5p expression was significantly increased.
Conversely, overexpression of AC245100.4 decreased miR-145-5p
expression (Fig. 4B). Transfection
with miR-145-5p inhibitor could significantly decrease miR-145-5p
expression, and transfection with miR-145-5p mimic could
significantly increase miR-145-5p expression (Fig. 4C). Furthermore, AC245100.4
expression was downregulated by exogenous miR-145-5p mimic
transfection, but upregulated by miR-145-5p inhibitor transfection
(Fig. 4D).
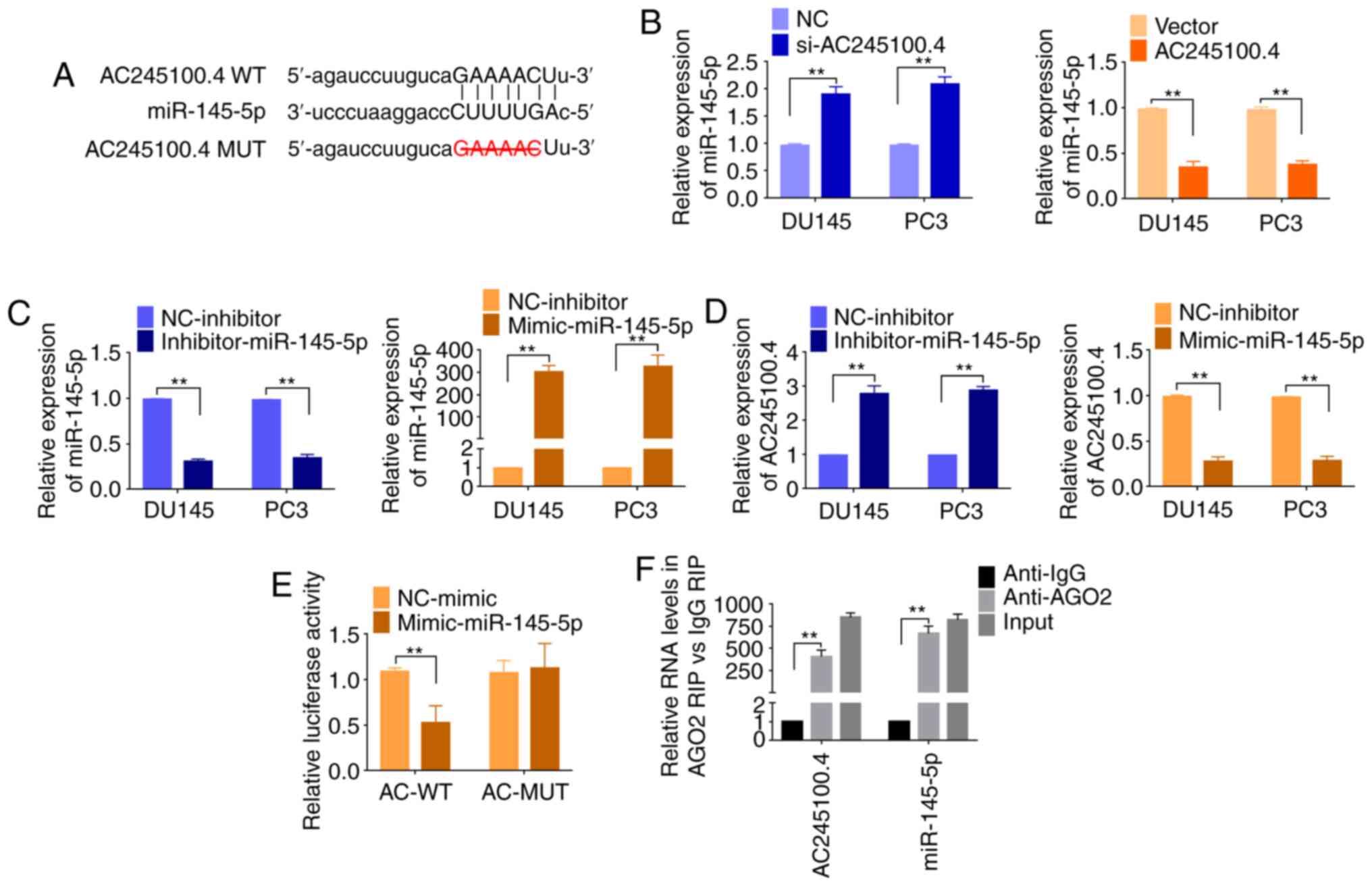 | Figure 4.miR-145-5p targets AC245100.4 in PCa.
(A) Bioinformatics analysis demonstrated that miR-145-5p directly
targeted the AC245100.4 sequence. (B) RT-qPCR analysis of
miR-145-5p expression in DU145 and PC3 cells transfected with
AC245100.4 si-RNA or pcDNA3.1-AC245100.4. (C) RT-qPCR analysis of
miR-145-5p expression in DU145 and PC3 cells transfected with
miR-145-5p mimic or inhibitor. (D) RT-qPCR analysis of AC245100.4
expression in DU145 and PC3 cells transfected with miR-145-5p mimic
or inhibitor. (E) Luciferase reporter plasmid containing AC-WT or
AC-MUT AC245100.4 was co-transfected into 293T cells with
miR-145-5p mimic or NC. (F) RIP experiments were performed in PC3
cells, and the coprecipitated RNA was subjected to RT-qPCR for
analysis of AC245100.4 and miR-145-5p expression. **P<0.01 vs.
IgG. Data are presented as the mean ± SD of three independent
experiments (n=3). **P<0.01. AC-WT, AC245100.4 wild-type;
AC-MUT, AC245100.4 mutant; miR-145-5p, microRNA-145-5p; PCa,
prostate cancer; RT-qPCR, reverse transcription-quantitative PCR;
NC, negative control; si, small interfering RNA; AGO2, catalytic
component; RIP, RNA immunoprecipitation. |
To confirm the direct binding, luciferase reporter
assays were conducted. A vector containing the AC245100.4/miR-
145-5p binding sequence (AC WT) and a vector with the deleted
binding site (AC MUT) were generated. Vectors were transfected
together with miR-145-5p mimics. The luciferase activity was
decreased following co-transfection of miR-145-5p mimic and AC WT
reporter vector, but remained unchanged for AC MUT vector (Fig. 4E), which indicated that AC245100.4
may be a target of miR-145-5p.
AC245100.4 and miR-145-5p both bind to
AGO2 in PCa cells
miRNAs exert their gene silencing function by
binding to AGO2, forming a functional RISC, and miRNAs degrade
mRNAs with the involvement of RISC (25). To further verify the present
hypothesis, anti-AGO2 RIP was performed using PC3 cell extracts.
RT-qPCR was used to determine whether AC245100.4 and miR-145-5p
were present in the immunoprecipitates. AC245100.4 was enriched in
anti-AGO2 conjugated immunoprecipitates compared with in the
anti-IgG control. Consistently, miR-145-5p was also present in the
immunoprecipitates (Fig. 4F).
Therefore, AC245100.4 and miR-145-5p were both pulled down by AGO2,
likely via direct binding. Overall, the present study suggested
that there was direct binding between AC245100.4 and miR-145-5p,
and demonstrated the reciprocal association between AC245100.4 and
miR-145-5p.
lncRNA AC245100.4 regulates RBBP5
expression indirectly by sponging miR-145-5p
To elucidate the ceRNA network among AC245100.4,
miR-145-5p and its targets in PCa, online prediction was conducted.
TargetScan predicted 907 targets and miRDB predicted 909 targets
(21,22). There were 100s of genes even after
the results were overlapped. RBBP5 was among the overlapping genes.
To validate the predicted relationships, luciferase assays were
performed. The WT reporter vector contained RBBP5 3′UTR binding
sites, but the binding sites were deleted in the MUT reporter
vector (Fig. 5A). The luciferase
activity was decreased after co-transfection of miR-145-5p mimic
and the RBBP5 WT reporter vector, but not the MUT vector,
indicating that miR-145-5p targeted RBBP5 by directly binding to
the 3′UTR of RBBP5 (Fig. 5B).
Additionally, miR-145-5p negatively regulated RBBP5 expression
(Fig. 5C-F). It was also identified
that knockdown or overexpression of AC245100.4 decreased or
increased RBBP5 protein expression, respectively, in DU145 and PC3
cells (Fig. 5E and F).
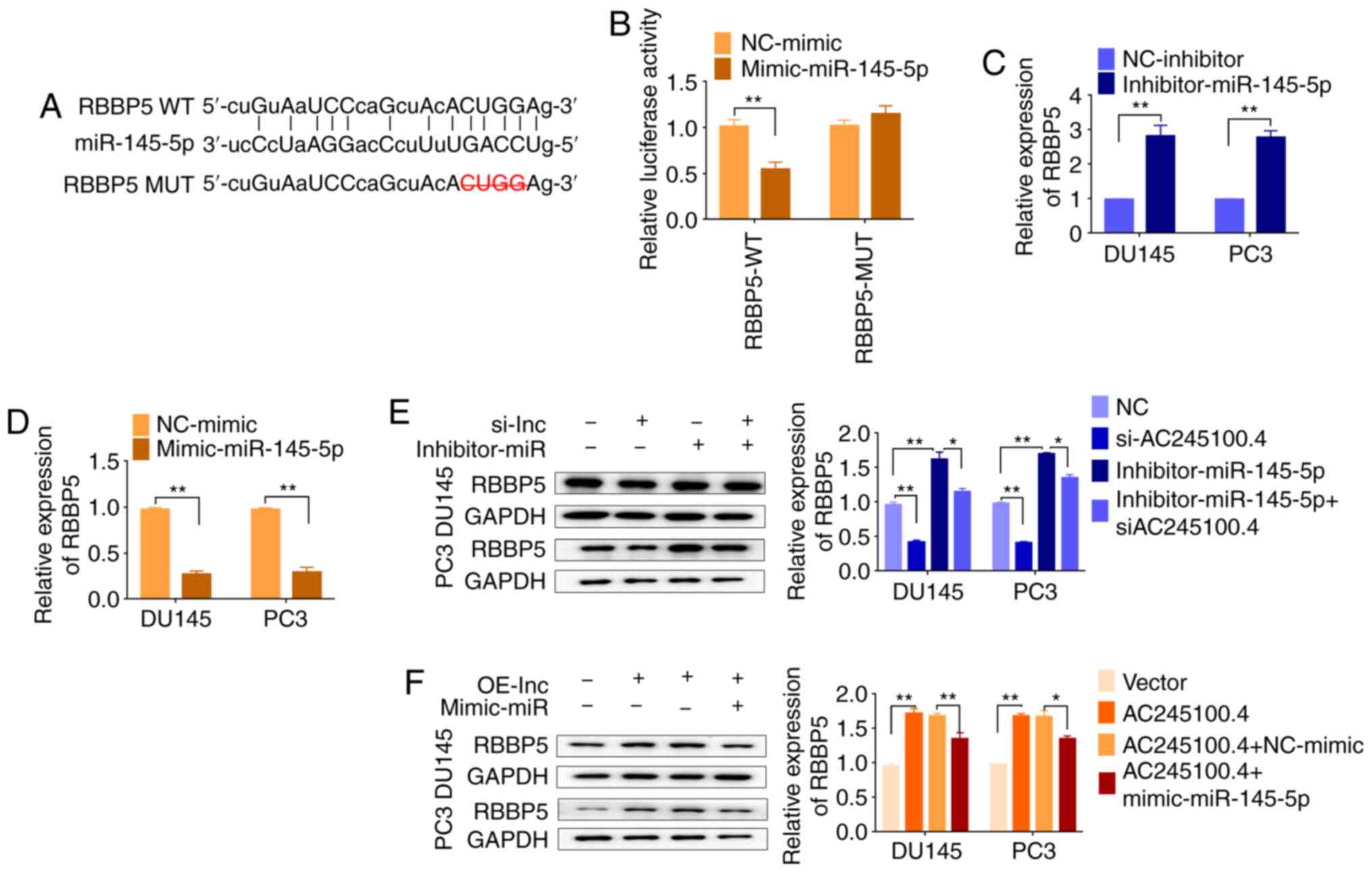 | Figure 5.RBBP5 is a miR-145-5p target gene and
is indirectly regulated by AC245100.4. (A) Bioinformatics analysis
demonstrated that RBBP5 is a target gene of miR-145-5p. (B)
Luciferase reporter plasmid containing RBBP5-WT or RBBP5-MUT
3′untranslated region of RBBP5 was co-transfected into 293T cells
with miR-145-5p mimic or NC. Reverse transcription-quantitative PCR
analysis of RBBP5 expression in DU145 and PC3 cells transfected
with miR-145-5p (C) inhibitor or (D) mimic. RBBP5 protein
expression was analyzed via western blotting in DU145 and PC3 cells
following (E) knockdown or (F) overexpression of AC245100.4 and
co-transfection with miR-145-5p inhibitor or mimic. GAPDH was used
as an internal control. Data are presented as the mean ± SD of
three independent experiments (n=3). *P<0.05 and **P<0.01.
OE-lnc, AC245100.4 overexpression; si-lnc, si-AC245100.4;
Inhibitor-miR, Inhibitor-miR-145-5p; Mimic-miR, mimic-miR-145-5p;
RBBP5, retinoblastoma binding protein 5; miR-145-5p,
microRNA-145-5p; WT, wild-type; MUT, mutant; NC, negative control;
si, small interfering RNA; lnc, long non-coding RNA. |
Subsequently, it was investigated whether miR-145-5p
was involved in the combination of AC245100.4 and RBBP5. As
presented in Fig. 5E and F, RBBP5
expression was restored by miR-145-5p inhibition in
AC245100.4-knockdown cells. Similarly, the increase in RBBP5
protein expression induced by AC245100.4 overexpression was
effectively reversed by miR-145-5p mimics. Collectively, these
findings suggested that AC245100.4 regulated the expression of
RBBP5 via regulation of miR-145-5p at the post-transcriptional
level.
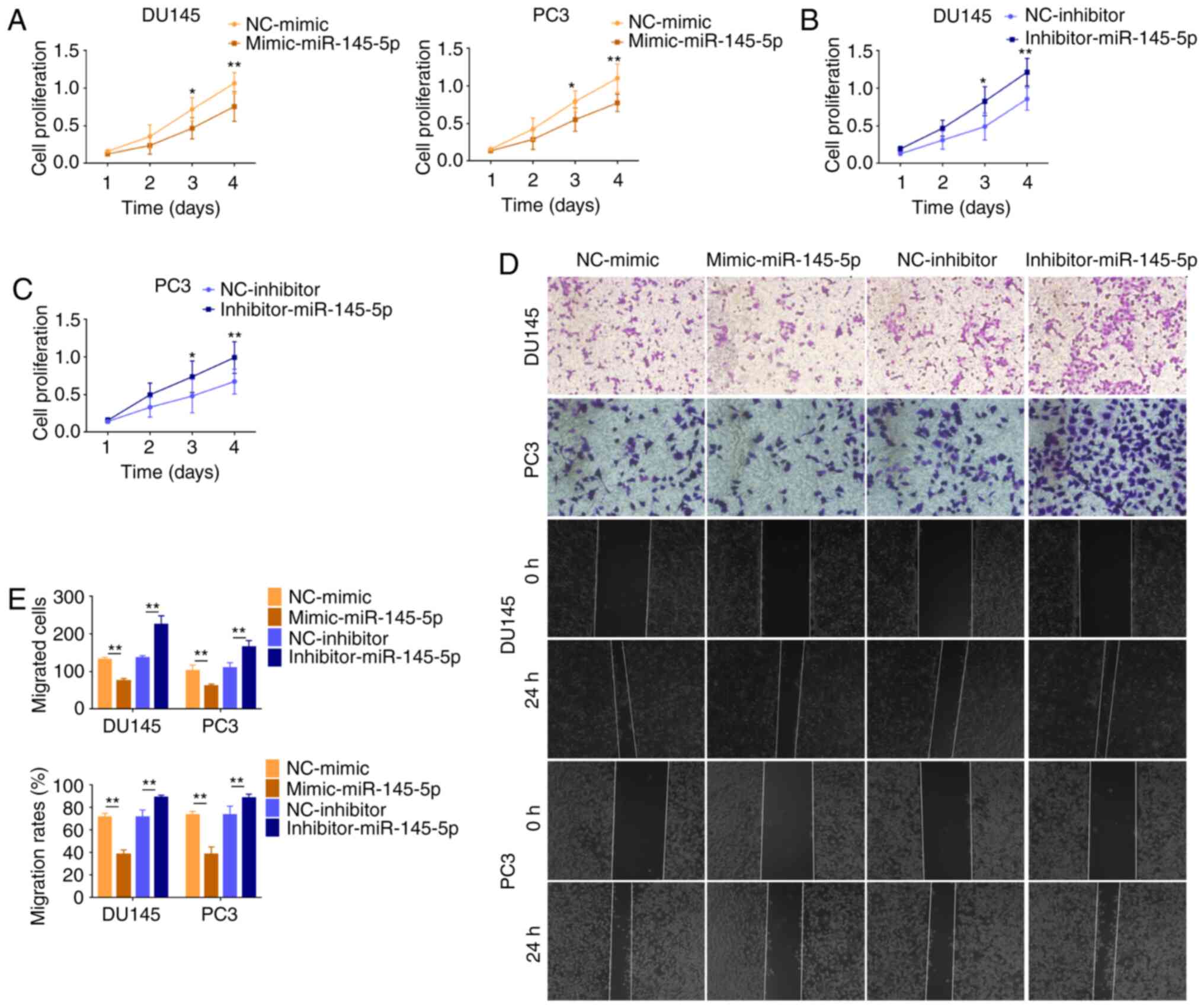
lncRNA AC245100.4 reverses the
suppressive role of miR-145-5p in PCa
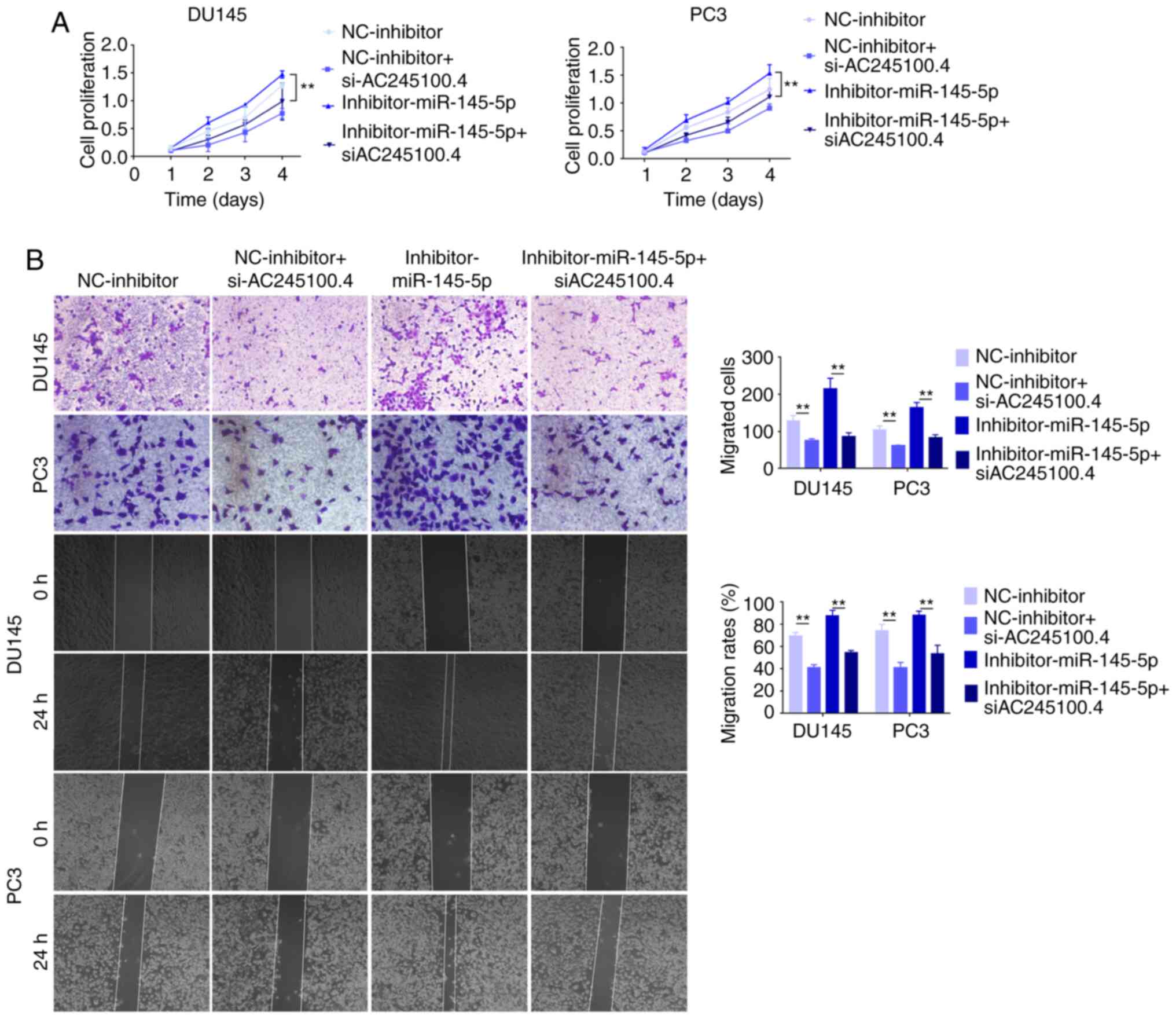
It was further investigated whether miR-145-5p
suppressed tumorigenesis in DU145 and PC3 cells. The proliferation
and migration of PCa cells were negatively associated with
miR-145-5p expression (Fig.
6A-E).
To examine the reciprocal effects of miR-145-5p on
AC245100.4, loss of function experiments were performed in PCa
cells. The results demonstrated that cell proliferation in the
inhibitor-miR-145-5p and si-AC245100.4 group was significantly
decreased compared with the inhibitor-miR-145-5p group, suggesting
that miR-145-5p silencing had promoting effect on proliferation,
and this was partially reversed by AC245100.4 knockdown (Fig. 7A). The cell migration in the
inhibitor-miR-145-5p and si-AC245100.4 group was significantly
decreased compared with the inhibitor-miR-145-5p group, suggesting
that the addition of the si-AC245100.4 altered the
inhibitor-miR-145-5p-mediated increase in cell migration (Fig. 7B). All data suggested that
miR-145-5p suppressed tumorigenesis in PCa and competed with
AC245100.4 to exert its inhibitory effect in PCa cells.
Discussion
PCa was the second most common newly diagnosed
cancer and fifth leading cause of cancer-associated mortality in
men worldwide in 2018 (4).
Therefore, a comprehensive understanding of the molecular events
responsible for PCa tumorigenesis is required. lncRNAs have gained
increasing attention as cryptic regulators in PCa (9). Thus, it is necessary to identify the
underlying mechanisms of these regulatory transcripts to fully
understand PCa carcinogenesis in order to develop novel diagnostic
targets and treatment strategies.
In the present study, a novel lncRNA, AC245100.4,
associated with PCa was identified. Upregulated expression levels
of AC245100.4 were observed in tumor tissues in TCGA data and cell
lines. The results demonstrated that AC245100.4 had an oncogenic
effect on PCa cell proliferation and migration. AC245100.4 exerted
its oncogenic role by sponging miR-145-5p, thereby repressing
degradation of RBBP5. The present findings provided evidence for
AC245100.4 as a candidate for overcoming therapeutic obstacles in
PCa.
Previous studies have reported that lncRNAs are
responsible for PCa carcinogenesis (9). For example, PCA3, metastasis
associated lung adenocarcinoma transcript 1 (MALAT1), HOX
transcript antisense RNA and prostate cancer associated transcript
(PCAT)19 serve oncogenic roles and promote growth in PCa, while
PCAT29 and Downregulated RNA in Androgen Independent Cells suppress
cellular migration and metastasis (8,26).
Certain characterized lncRNAs, such as PCA3, H19 and PCAT29, have
been considered to be more specific to PCa (8). Our previous study predicted that
AC245100.4 was closely associated with PCa using bioinformatics
methods (17). In the present
study, AC245100.4 was demonstrated to promote proliferation and
migration in PCa cell lines, and to induce tumor growth in an
animal model. Upregulated expression levels of AC245100.4 were
observed in TCGA tumor samples. To the best of our knowledge, this
was the first study to identify the oncogenic function of
AC245100.4 in PCa. However, the expression levels of AC245100.4
should be studied in additional patients in future studies.
A number of lncRNAs exert their function by acting
as ceRNAs in PCa (14,15,17).
In ceRNA networks, miRNAs are important mediators, and miRNAs could
serve roles in PCa development, and thus are potential targets for
novel therapies (27,28). There has been increased interested
in ncRNAs (miRNAs and lncRNAs) due to limitations of coding genes
in cancer diagnosis and treatment (29). lncRNA-miRNA ceRNA networks
represents the competitive association between lncRNAs and miRNAs,
which could explain some features of PCa. For example, lncRNA HLA
complex P5 expression is upregulated in PCa, and promotes cell
proliferation by inducing cell migration inducing hyaluronidase 1
expression by sponging miR-4656 (30).
In the present study, an online database of
biological information predicted that AC245100.4 had complementary
binding sites with miR-145-5p. The direct binding was further
confirmed using a luciferase reporter assay. Both AC245100.4 and
miR-145-5p could interact with AGO2 in PCa cells, which suggested
that AC245100.4 may function as an endogenous miRNA sponge.
Numerous studies have revealed that miR-145-5p suppresses tumor
progression in various types of cancer, including melanoma, NSCLC,
bladder cancer and gastric cancer (31–33).
In PCa, miR-145-5p inhibits docetaxel resistance by repressing
A-kinase anchoring protein 12, which could be reversed by MALAT1 in
a ceRNA manner (34). Furthermore,
the miR-145-5p/fascin actin-bundling protein 1 axis contributes to
tumor progression mediated by PCAT1, and PCAT1 competes with
miR-145-5p by direct binding (35).
The functions of miRNAs largely depend on the specific target, thus
the molecular mechanism of miR-145-5p was also a focus of the
present study. It was demonstrated that PCa cell proliferation and
migration were impaired following transfection of miR-145-5p
mimics. Moreover. AC245100.4 promoted the proliferation and
migration of DU145 and PC3 cells by targeting miR-145-5p. It is
important to identify real markers and develop novel specific
therapies to decrease the mortality of PCa (36). Therefore, further study of miRNAs is
important (37). The present
findings reinforced the tumor-suppressive role of miR-145-5p in
PCa, which suggests it as a promising biomarker.
The ceRNA network also requires a miRNA target gene
to complete the regulatory axis (10). According to bioinformatics analyses,
RBBP5 and AC245100.4 share the same response elements for
miR-145-5p. The association was verified using luciferase reporter
assays. Furthermore, miR-145-5p expression was negatively
associated with RBBP5 protein expression. It was identified that
knockdown or overexpression of AC245100.4 decreased or increased
RBBP5 protein expression. Therefore, it was demonstrated that RBBP5
was targeted by miR-145-5p. AC245100.4 may induce RBBP5 expression
by repressing its inhibitor, miR-145-5p. RBBP5 is a coding protein
in the nucleus (38), and can bind
to RB transcriptional corepressor 1, a known tumor suppressor
(39). However, the role of RBBP5
is ambiguous. Increased RBBP5 expression has been observed in
patients with hepatocellular carcinoma, and RBBP5 knockdown can
inhibit the proliferation of hepatocellular carcinoma cells
(40). Moreover, RBBP5 silencing
promotes cell adhesion and migration in cervical cancer cells
(41). These findings indicate that
RBBP5 has dual roles in cancer. The exact role and underlying
mechanism of RBBP5 in PCa should be investigated in future
studies.
In conclusion, the present study demonstrated that
the novel lncRNA AC245100.4 was upregulated in PCa cells and may be
involved in the development of PCa. High AC245100.4 expression may
be one of the factors affecting the prognosis of patients with PCa.
Mechanistically, AC245100.4 can regulate the expression of RBBP5,
acting as a ceRNA of miR-145-5p. The present findings provide
evidence that AC245100.4 may be a promising diagnostic or
therapeutic marker for PCa.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81572535), China
Postdoctoral Science Foundation (grant no. 2019M651297) and The
Fundamental Research Funds for the Provincial Universities of
Heilongjiang (grant no. 2018-KYYWF-0432).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HX and JBZ contributed equally. XGY and PL designed
and supervised the study. JNZ, QQW, XY and WYY performed the in
vitro experiments. HX and JBZ analyzed the data and wrote the
manuscript. JHW performed the bioinformatics analysis. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The animal experiments were approved by the Animal
Ethics Committee of Harbin Medical University (Harbin, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL and Miller KD: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL and Miller KD: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen QN, Wei CC, Wang ZX and Sun M: Long
non-coding RNAs in anti-cancer drug resistance. Oncotarget.
8:1925–1936. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tehrani SS, Karimian A, Parsian H,
Majidinia M and Yousefi B: Multiple functions of long non-coding
RNAs in oxidative stress, DNA damage response and cancer
progression. J Cell Biochem. 119:223–236. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Quinn JJ and Chang HY: Unique features of
long non-coding RNA biogenesis and function. Nat Rev Genet.
17:47–62. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ramnarine VR, Kobelev M, Gibb EA, Nouri M,
Lin D, Wang Y, Buttyan R, Davicioni E, Zoubeidi A and Collins CC:
The evolution of long noncoding RNA acceptance in prostate cancer
initiation, progression, and its clinical utility in disease
management. Eu Urol. 76:546–559. 2019. View Article : Google Scholar
|
|
9
|
Fatima R, Akhade VS, Pal D and Rao SM:
Long noncoding RNAs in development and cancer: Potential biomarkers
and therapeutic targets. Mol Cell Ther. 3:52015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The Rosetta Stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu P, Cai J, Chen Q, Han B, Meng X, Li Y,
Li Z, Wang R, Lin L, Duan C, et al: Lnc-TALC promotes
O6-methylguanine-DNA methyltransferase expression via
regulating the c-Met pathway by competitively binding with
miR-20b-3p. Nat Commun. 10:20452019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu XS, Wang F, Li HF, Hu YP, Jiang L,
Zhang F, Li ML, Wang XA, Jin YP, Zhang YJ, et al: LncRNA-PAGBC acts
as a microRNA sponge and promotes gallbladder tumorigenesis. EMBO
Rep. 18:1837–1853. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Peng W, He D, Shan B, Wang J, Shi W, Zhao
W, Peng Z, Luo Q, Duan M, Li B, et al: LINC81507 act as a competing
endogenous RNA of miR-199b-5p to facilitate NSCLC proliferation and
metastasis via regulating the CAV1/STAT3 pathway. Cell Death Dis.
10:5332019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang G, He X, Ren C, Lin J and Wang Q:
Long noncoding RNA PCA3 regulates prostate cancer through sponging
miR-218-5p and modulating high mobility group box 1. J Cell
Physiol. 234:13097–13109. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu X, Xiao Y, Zhou Y, Zhou Z and Yan W:
LncRNA FOXP4-AS1 is activated by PAX5 and promotes the growth of
prostate cancer by sequestering miR-3184-5p to upregulate FOXP4.
Cell Death Dis. 10:4722019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cheng G, Song Z, Liu Y, Xiao H, Ruan H,
Cao Q, Wang K, Xiao W, Xiong Z, Liu D, et al: Long noncoding RNA
SNHG12 indicates the prognosis of prostate cancer and accelerates
tumorigenesis via sponging miR-133b. J Cell Physiol. 235:1235–1246.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu Y, Zhang R, Zhao N, Zhang Q, Yan Z,
Chang Z, Wei Y, Wu C, Xu J and Xu Y: A comparative analysis reveals
the dosage sensitivity and regulatory patterns of lncRNA in
prostate cancer. Mol Biosyst. 12:3176–3185. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45:W98–W102.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang H, Jiang M, Liu Q, Han Z, Zhao Y and
Ji S: miR-145-5p inhibits the proliferation and migration of
bladder cancer cells by targeting TAGLN2. Oncol Lett. 16:6355–6360.
2018.PubMed/NCBI
|
|
21
|
Chen Y and Wang X: miRDB: An online
database for prediction of functional microRNA targets. Nucleic
Acids Res. 48:D127–D131. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
Elife. 4:e050052015. View Article : Google Scholar
|
|
23
|
Paraskevopoulou MD, Vlachos IS, Karagkouni
D, Georgakilas G, Kanellos I, Vergoulis T, Zagganas K, Tsanakas P,
Floros E, Dalamagas T and Hatzigeorgiou AG: DIANA-LncBase v2:
Indexing microRNA targets on non-coding transcripts. Nucleic Acids
Res. 44:D231–D238. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Aiello NM and Kang Y: Context-dependent
EMT programs in cancer metastasis. J Exp Med. 216:1016–1026. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Golden RJ, Chen B, Li T, Braun J,
Manjunath H, Chen X, Wu J, Schmid V, Chang TC, Kopp F, et al: An
Argonaute phosphorylation cycle promotes microRNA-mediated
silencing. Nature. 542:197–202. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sakurai K, Reon BJ, Anaya J and Dutta A:
The lncRNA DRAIC/PCAT29 locus constitutes a tumor-suppressive
nexus. Mol Cancer Res. 13:828–838. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cochetti G, Poli G, Guelfi G, Boni A,
Egidi MG and Mearini E: Different levels of serum microRNAs in
prostate cancer and benign prostatic hyperplasia: Evaluation of
potential diagnostic and prognostic role. OncoTargets Ther.
9:7545–7553. 2016. View Article : Google Scholar
|
|
28
|
Guelfi G, Cochetti G, Stefanetti V,
Zampini D, Diverio S, Boni A and Mearini E: Next generation
sequencing of urine exfoliated cells: An approach of prostate
cancer microRNAs research. Sci Rep. 8:71112018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Egidi MG, Cochetti G, Serva MR, Guelfi G,
Zampini D, Mechelli L and Mearini E: Circulating microRNAs and
kallikreins before and after radical prostatectomy: Are they really
prostate cancer markers? Biomed Res Int. 2013:2417802013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hu R and Lu Z: Long non-coding RNA HCP5
promotes prostate cancer cell proliferation by acting as the sponge
of miR-4656 to modulate CEMIP expression. Oncol Rep. 43:328–336.
2020.PubMed/NCBI
|
|
31
|
Jin C, Wang A, Liu L, Wang G, Li G and Han
Z: miR-145-5p inhibits tumor occurrence and metastasis through the
NF-κB signaling pathway by targeting TLR4 in malignant melanoma. J
Cell Biochem. Jan 30–2019.(Epub ahead of print). View Article : Google Scholar
|
|
32
|
Lu Q, Shan S, Li Y, Zhu D, Jin W and Ren
T: Long noncoding RNA SNHG1 promotes non-small cell lung cancer
progression by up-regulating MTDH via sponging miR-145-5p. FASEB J.
32:3957–3967. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sun M, Zhao W, Chen Z, Li M, Li S, Wu B
and Bu R: Circular RNA CEP128 promotes bladder cancer progression
by regulating Mir-145-5p/Myd88 via MAPK signaling pathway. Int J
Cancer. 145:2170–2181. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xue D, Lu H, Xu HY, Zhou CX and He XZ:
Long noncoding RNA MALAT1 enhances the docetaxel resistance of
prostate cancer cells via miR-145-5p-mediated regulation of AKAP12.
J Cell Mol Med. 22:3223–3237. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu W, Chang J, Du X and Hou J: Long
non-coding RNA PCAT-1 contributes to tumorigenesis by regulating
FSCN1 via miR-145-5p in prostate cancer. Biomed Pharmacother.
95:1112–1118. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Egidi MG and Cochetti G: Stability
assessment of candidate reference genes in urine sediment of
prostate cancer patients for miRNA applications. Dis Markers.
2015:9735972015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cochetti G, Rossi de Vermandois JA, Maulà
V, Giulietti M, Cecati M, Del Zingaro M, Cagnani R, Suvieri C,
Paladini A and Mearini E: Role of miRNAs in prostate cancer: Do we
really know everything? Urol Oncol. 38:623–635. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Saijo M, Sakai Y, Kishino T, Niikawa N,
Matsuura Y, Morino K, Tamai K and Taya Y: Molecular cloning of a
human protein that binds to the retinoblastoma protein and
chromosomal mapping. Genomics. 27:511–519. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gallie BL, Campbell C, Devlin H, Duckett A
and Squire JA: Developmental basis of retinal-specific induction of
cancer by RB mutation. Cancer Res. 59 (7 Suppl):1731S–1735S.
1999.PubMed/NCBI
|
|
40
|
Zhou H, Bao J, Zhu X, Dai G, Jiang X, Jiao
X, Sheng H, Huang J and Yu H: Retinoblastoma binding protein 5
correlates with the progression in hepatocellular carcinoma. BioMed
Res Int. 2018:10734322018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xia B, Joubert A, Groves B, Vo K, Ashraf
D, Djavaherian D, Awe J, Xiong Y, Cherfils J and Ma D: Modulation
of cell adhesion and migration by the histone methyltransferase
subunit mDpy-30 and its interacting proteins. PLoS One.
5:e117712010. View Article : Google Scholar : PubMed/NCBI
|