Introduction
Renal cell carcinoma (RCC) is a common type of
kidney cancer, and >300,000 new cases are diagnosed worldwide
each year (1). Currently, surgical
resection followed by chemotherapy and radiotherapy are the primary
therapeutic treatments for RCC (2).
Although great improvement has been made in the diagnosis and
treatment of RCC in previous decades, RCC remains one of the most
drug-resistant malignancies and a frequent cause of cancer-related
deaths (2). Therefore, there is an
urgent need to identify novel and effective therapeutic strategies
for RCC.
In recent years, antitumour compounds from natural
herbs have received increasing attention due to their high
efficiency and low toxicity. It was estimated that >50% of the
small molecular antitumour agents that were developed between 1950
and 2015 originated from natural products or their derivates
(3). Ginseng, the root and rhizome of
Panax ginseng Meyer, has been widely used as a traditional
medicine in East Asia for thousands of years. Mounting evidence has
indicated that ginseng has different pharmacological effects on
various human diseases (4). In
addition, ginseng has also been found to possess anti-aging and
antioxidant effects (5).
Ginsenosides, a group of various steroidal saponins, are the major
bioactive compounds in ginseng, and >40 ginsenosides have been
identified thus far. Ginsenosides have been found to possess
various pharmacological activities, such as neuroprotection,
cardioprotection, anti-depressant, anti-inflammatory and antitumour
activities (5–7).
Compound K (CK)
[20-O-β-d-glucopyranosyl-20(S)-protopanaxadiol] is a major
metabolite of the ginsenosides Rb1, Rb2, Rc and Rd that are
generated by intestinal bacteria via the multistage cleavage of
sugar moieties after the oral administration of ginseng (8). To date, numerous investigations
regarding the antitumour activities of CK have been conducted. For
example, CK has been found to inhibit the tumorigenic activities of
glioma and glioma cells (9,10). CK can also inhibit lung cancer cells
via various mechanisms, such as endoplasmic reticulum stress and
hypoxia-inducible factor-1α (HIF-1α)-mediated glucose metabolism
pathways (11,12). In addition, CK also possesses
antitumour activities against leukaemia, nasopharyngeal carcinoma
and colorectal cancer (13–15). However, no investigation has been
conducted concerning the effects of CK on RCC cells.
In the present study, the antitumour activity of CK
was investigated in RCC cells. The results showed that CK could
inhibit the proliferation and metastatic ability of RCC cells. It
was also observed that CK could induce cell cycle arrest and
apoptosis in RCC cells. The mechanistic studies revealed that CK
exerted its antitumour effects at least partly via multiple
mechanisms. These data revealed that CK could have applications as
a novel, natural agent against RCC.
Materials and methods
Cell culture and chemicals
HK-2, Caki-1 and 786-O cells were purchased from the
American Type Culture Collection. All cells were cultured in
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.) with 10%
heat-inactivated foetal bovine serum (FBS; HyClone; Cytiva), 100
U/ml penicillin and 100 µg/ml streptomycin (Sigma-Aldrich; Merck
KGaA) at 37°C in a humidified 5% CO2 atmosphere. CK was
purchased from Sichuan WeiKeqi Biological Technology Co., Ltd.
Pan-caspase inhibitor z-VAD was obtained from Sigma-Aldrich (Merck
KGaA) and was used at a concentration of 10 µM at 37°C. z-VAD was
added into the culture medium 0.5 h prior to the treatment with CK.
N-Acetyl-L-cysteine (NAC) was purchased from Sigma-Aldrich (Merck
KGaA) and was used at 10 µM at 37°C. NAC was added into the culture
medium 0.5 h prior to the treatment with CK. All other chemicals
were obtained from Sigma-Aldrich (Merck KGaA).
Transfection
Small interfering RNA (siRNA) targeting testis
associated oncogenic (THOR) (si-THOR), negative control scramble
siRNA (si-NC), pcDNA3.1 expressing THOR and empty control were
synthesized and obtained from Suzhou GenePharma Co., Ltd.
Transfections were performed using Lipofectamine® 2000
(Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocols. Briefly, cells were seeded into a 6-well plate at a
density of 1×105 cells/well. Then, 12 h later, 20 mM
siRNAs and/or 10 ng pcDNA3.1 mixed with 5 µl Lipofectamine in 200
µl Opti-MEM® (Thermo Fisher Scientific, Inc.) were added
into each well. At 24 h after transfection, cells were collected
and assayed. The sequences used were as follows: si-THOR sense,
5′-GGUGAACACAAUCGAGCAATT-3′ and anti-sense,
5′-UUGCUCGAUUGUGUUCACCTT-3′; scrambled siRNA sense,
5′-ACGCGUAACGCGGGAAUUUdTdT-3′ and antisense
5′-AAAUUCCCGCGUUACGCGUdTdT-3′; and THOR sense,
5′-CTAATACGACTCACTATAGGGAGAAGCCGAGTTCGCGCCGCCGGTA-3′ and antisense,
5′-AAATATTTGGCTTTCCCCGGCC-3′.
Clonogenic assay
For the colony formation assay, cells were seeded in
6-well plates at a density of 500 cells/well and cultured in
complete medium (Thermo Fisher Scientific, Inc.). Two weeks later,
the cells were fixed and stained with 0.1% crystal violet (Beyotime
Institute of Biotechnology) for 30 min at room temperature. The
number of visible colonies (>50 cells/colony) was counted using
the CKX41-32 inverted microscope (magnification, ×100; Olympus
Corporation).
Cell viability assay
Cells were seeded into 96-well plates
(1×104 cells/well). A total of 24 h after culture, the
cells were treated with various doses of CK (10, 20 or 40 µM) for
different times (24, 48 or 72 h). Then, cells were collected, and
cell viability was measured as previously described (16).
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from cells using
TRIzol® reagent (Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. The quality and
quantity of RNA were measured using a NanoDrop™ 2000 (Thermo Fisher
Scientific, Inc.). Total RNA was reverse transcribed into cDNA
using the PrimeScript RT Reagent Kit (Takara Biotechnology Co.,
Ltd.) according to the manufacturer's instructions. RT-qPCR was
performed on an Applied Biosystems 7500 Detection System (Applied
Biosystems; Thermo Fisher Scientific, Inc.) using SYBR®
Premix Ex Taq™ (Takara Biotechnology Co., Ltd.) according to the
manufacturer's instructions. The following primers were used: HOX
antisense intergenic RNA forward, 5′-CAGTGGGGAACTCTGACTCG-3′ and
reverse, 5′-GTGCCTGGTGCTCTCTTACC-3′; double homeobox A pseudogene 8
forward, 5′-AGGATGGAGTCTCGCTGTATTGC-3′ and reverse,
5′-GGAGGTTTGTTTTCTTCTTTTTT-3′; small nucleolar RNA host gene 14
forward, 5′-GGGTGTTTACGTAGACCAGAACC-3′ and reverse,
5′-CTTCCAAAAGCCTTCTGCCTTAG-3′; colorectal neoplasia differentially
expressed forward, 5′-GAGGACGTGCTGGGGCT-3′ and reverse,
5′-CTGAGTCCATGTCCCGAATC-3′; growth arrest specific 5 forward,
5′-AGCTGGAAGTTGAAATGG-3′ and reverse, 5′-CAAGCCGACTCTCCATACC-3′;
THOR forward, 5′-CAAGGTGCTTCTCTCTGGATTT-3′ and reverse,
5′-GCCAAAGTCATTTGTTGGGTAT-3′; LOC653786 forward,
5′-CTCTCTGGGCTTGATAGCAT-3′ and reverse,
5′-ACCATTTAGCATTAAGGCAGTAG-3′; HEIRCC forward,
5′-ACCTCCAGAACTGTGATCCAAAATG-3′ and reverse,
5′-TCTTGCTTGATGCTTTGGTCTG-3′; long intergenic non-protein coding
RNA 460 forward, 5′-ACAGCATGAGCCAGGACATC-3′ and reverse,
5′-GAAAGCTGCAACATGCTCCC-3′; SPRY4 intronic transcript 1 forward,
5′-GCTGAGCTGGTGGTTGAAAGGAATC-3′ and reverse,
5′-GCTTGGCCCACGATGACTTGG-3′; PVT1 forward,
5′-GCCCCTTCTATGGGAATCACTA-3′ and reverse,
5′-GGGGCAGAGATGAAATCGTAAT-3′; urothelial cancer-associated 1
forward, 5′-CCGCTCGAGAGCGCGTGTGGCGGCCGAGCAC-3′ and reverse,
5′-CGCGGATCCAGACACGAGGCCGGCCACGCCACG-3′; metastatic renal cell
carcinoma-associated transcript 1 forward,
5′-CCTATCCCTTTCTCTAAGAA-3′ and reverse, 5′-ACTTCTGCAAAAACGTGCTG-3′;
metastasis associated lung adenocarcinoma transcript 1 forward,
5′-AAAGCAAGGTCTCCCCACAAG-3′ and reverse,
5′-GGTCTGTGCTAGATCAAAAGGCA-3′; TP73 antisense RNA 1 forward,
5′-CCGGTTTTCCAGTTCTTGCAC-3′ and reverse,
5′-GCCTCACAGGGAAACTTCATGC-3′; and GAPDH forward,
5′-TGAAGGTCGGAGTCAACGGATTTGGT-3′ and reverse,
5′-CATGTGGGCCATGAGGTCCACCAC-3′. The thermocycling conditions were
as follows: 95°C for 10 min; followed by 35 cycles of 15 sec at
94°C, 15 sec at 60°C and 15 sec at 72°C; and a final extension at
72°C for 10 min. The results were calculated with the
2−ΔΔCq method (17) and
normalized to the expression of GAPDH. All the assays were
performed in triplicate. The expression levels are shown as
fold-change relative to the corresponding controls, which were
defined as 1.0.
Long non-coding RNA (lncRNA) PCR
array
Cells were seeded into 6-well plates
(1×105 cells/well) and treated with or without CK (40
µM) for 24 h. Total lncRNA were extracted using TRIzol (Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. Next, the RNAs were sent to Shanghai Biotechnology
Corporation for human cancer lncPCR array. A total of 6 reference
genes and 97 lncRNAs were included in the array (RT2
lncRNA PCR Arrays; Qiagen, Inc.). lncRNAs were extracted from
various databases, including Ensembl (18), manually curated lncRNA literature
sources (search terms, ‘lncRNA’ and ‘cancer’), RefSeq (https://www.ncbi.nlm.nih.gov/refseq/)
and UCSC Genome Browser (https://genome.ucsc.edu). Briefly, cDNA was
synthesized using the RT2 First Strand Kit (Qiagen
GmbH). Then, qPCR was performed using the RT2
SYBR® Green Mastermix (Qiagen GmbH). Only one replicate
for each sample and one pair of primers for each lncRNA were used
for the Human Cancer LncRNA PCR Array. The parameters for qPCR
were: 95°C for 10 min; 95°C for 15 sec and 60°C for 1 min (40
cycles). lncRNA expression was compared using the 2−ΔΔCq
method (17). Differentially
expressed lncRNAs with statistical significance (as determined
using a two-tailed Student's t-test, P<0.05) were
identified.
Wound healing assay
Caki-1 and 786-O cells were seeded at a density of
1×106 cells/well in 6-well plates and cultured in medium
with a low concentration of FBS (5%). FBS was used in order to
reduce cell proliferation risk, cells were cultured in a medium
(19). After culture for 24 h when
cells reached ~90% confluency, they were scratched using a 200-µl
pipette tip and carefully washed with sterile PBS twice to remove
the debris. Then, cells were treated with various doses of CK (10,
20 or 40 µM) at 37°C. At different time points (0 and 24 h), the
images were captured using the CKX41-32 inverted microscope
(Olympus) and the results were analysed using ImageJ software
(v1.46; National Institutes of Health).
Invasion assay
For the cell invasion analysis, a 24-well Transwell
assay plate (Corning Life Sciences) was used. Matrigel
(Sigma-Aldrich; Merck KGaA) was diluted with serum-free RPMI-1640
medium (1:3) and used to coat the upper surface of the Transwell
chamber at room temperature for 1 h. RCC cells (2×104
cells/well) exposed to different treatments were added into the
upper chamber with 500 µl serum-free RPMI-1640. The lower chamber
was filled with 500 µl complete medium with 20% FBS, and cells were
cultured for 48 h at 37°C. Subsequently, the invasive cells were
fixed with methanol and stained with 0.5% crystal violet (Beyotime
Institute of Biotechnology) at room temperature for 1 h. Images of
cell invasion were captured under the CKX41-32 inverted microscope
(Olympus Corporation).
Cell cycle distribution and apoptosis
assay
For cell cycle analysis, cells were collected and
washed with 1X PBS after being treated with various doses of CK
(10, 20 or 40 µM) for 24 h at 37°C. The cell pellets were fixed in
70% cold ethanol for 30 min at 4°C. The fixed cells were
resuspended in 1X PBS containing 1 mg/ml RNase A (Sigma-Aldrich;
Merck Cayuga) and incubated for 1 h at 37°C, and the cells were
stained with 50 µg/ml PI (Sigma-Aldrich; Merck KGaA) for 30 min at
room temperature in the dark. To measure apoptosis, cells were
collected and washed with PBS. Then, the pellets were resuspended
in 1X Annexin V Binding Buffer (Sigma-Aldrich; Merck KGaA) followed
by incubation for 5 min in the dark with 5 µl Annexin V-FITC and PI
(Sigma-Aldrich; Merck KGaA). Both cell cycle distribution and
apoptosis were measured using the BD FACSCanto™ II flow cytometer
(BD Biosciences) and analysed with FlowJo software (v8.8.5; FlowJo
LLC).
Measurement of reactive oxygen species
(ROS)
The generation of ROS was measured by staining with
2′,7′-dichlorofluorescin diacetate (DCFHDA; Sigma-Aldrich; Merck
KGaA), which is converted into fluorescent 2′,7′-dichlorofluorescin
(DCF) in the presence of peroxides. Therefore, an increase in DCF
fluorescence is an indicator of the presence of ROS. A ROS
detection assay kit (cat. no. S0033S; Beyotime Institute of
Biotechnology) was used to measure intracellular oxidative stress
according to the manufacturer's instructions.
Caspase-3 activity assay
Caspase-3 activity was measured using a colorimetric
kit (cat. no. ab252897; Abcam), according to the manufacturer's
instructions. Briefly, cells were lysed in the lysis buffer
provided by the kit after treatment with various doses of CK (10,
20 or 40 µM) for 24 h at 37°C, and Ac-DEVD-pNA was used as the
substrate. Then, the absorbance was measured at OD405 with an
ELx800™ microplate reader (BioTek Instruments, Inc.).
Western blotting
After treatment, cells were collected and lysed in
RIPA buffer (Beyotime Institute of Biotechnology). The protein
concentrations were measured using a Bradford protein assay kit
(Beyotime Institute of Biotechnology). Equal amounts of protein (20
µg) were separated via SDS-PAGE on 12% gel, and separated proteins
were subsequently transferred to PVDF membranes (EMD Millipore).
After blocking with 5% skimmed milk for 1 h at room temperature,
the PVDF membranes were incubated with the primary antibodies (Cell
Signalling Technology, Inc.) overnight at 4°C. The following
primary antibodies were used: Anti-MMP-2 (cat. no. 40994; 1:1,000),
anti-MMP-9 (cat. no. 13667; 1:1,000), anti-N-cadherin (cat. no.
13116; 1:1,000), anti-E-cadherin (cat. no. 14472; 1:1,000),
anti-Vimentin (cat. no. 5741; 1:1,000), anti-Bcl-2 (cat. no. 15071;
1:1,000), anti-Bcl-xl (cat. no. 2762; 1:1,000), anti-Bax (cat. no.
2774; 1:1,000), anti-caspase-3 (cat. no. 14220; 1:1,000) and
anti-GAPDH (cat. no. 5174; 1:5,000). Then, the membranes were
incubated at room temperature for 1 h with anti-mouse (cat. no.
7076) and anti-rabbit HRP-conjugated secondary antibodies (1:5,000;
Cell Signalling Technology, Inc.) and visualized using ECL (Thermo
Fisher Scientific, Inc.).
Xenograft models
A total of 24 male BALB/c nude mice (age, 4–6 weeks
old; weight, 18–23 g) were randomly divided into the following four
groups (6 mice/group): i) Treatment with 0 mg/kg CK; ii) Treatment
with 25 mg/kg CK; iii) treatment with 50 mg/kg CK; and iv)
Treatment with 75 mg/kg CK. All mice were kept in housing
conditions of 40–70% humidity in a 12 h dark/light cycle with free
access to food and water at 21–25°C. 786-O and Caki-1 cells
(1×107) were suspended in 50 µl MEM media mixed with 50
µl Matrigel (BD Biosciences) and injected subcutaneously into the
right flank of mice under anesthetiza with 1.5% pentobarbital
sodium (60 mg/kg body weight; intraperitoneal injection). Weight
loss exceeding 20% was considered a humane endpoint. No animals
died during the experimental period. When the tumour size reached
~100 mm3, the mice were intravenously injected with
various concentrations of CK (0, 25, 50 or 75 mg/kg) for 30 days.
The tumour sizes were calculated using a calliper every 3 days, and
two perpendicular diameters of each tumour were recorded. At 30
days after implantation, mice were euthanized by CO2
inhalation (15% displacement) and the death of mice was confirmed
when there was a lack of pupil response to light. The tumour volume
was measured using the following formula: Volume=(width2
× length)/2. The protocol was approved by the Institutional Animal
Care and Use Committee of Zhongda Hospital, Southeast University
(Nanjing, China). All experiments involving animals were conducted
following the Guide for the Care and Use of Laboratory Animals
published by the Institute for Laboratory Animal Research (20). Necessary efforts were made to minimize
the number of animals and their suffering.
Haematoxylin and eosin (H&E)
staining and immuno-histochemistry (IHC)
Tumour tissues were fixed with 4% paraformaldehyde
(Sigma-Aldrich; Merck KGaA) overnight at room temperature, and then
paraffin embedded tissues were cut into 5-µm thick sections. For
H&E staining, the sections were stained with haematoxylin
(Sigma-Aldrich; Merck KGaA) for 40 sec and eosin (Sigma-Aldrich;
Merck KGaA) for 30 sec at room temperature. For IHC staining,
paraffin-embedded sections (5 µm) were blocked with 10% bovine
serum albumin (Sigma-Aldrich; Merck KGaA) for 1 h at room
temperature, post-hybridized slides were incubated with anti-Ki-67
antibody (cat. no. ab15580; 1:500; Abcam) overnight at 4°C. Then,
slides were washed three times with 0.01 M PBS for 10 min at room
temperature. The slides were incubated with HRP-conjugated rabbit
anti-mouse (cat. no. ab7076; 1:1,000; Abcam) secondary antibody for
1 h at room temperature, and then washed three times for 10 min
with 0.01 M PBS. Colour development was conducted using a DAB kit
(cat. no. ab64238; Abcam), according to the manufacturer's
instructions. The images were visualized using a light fluorescence
Olympus BX40 microscope (Olympus Corporation).
Statistical analysis
Statistical analysis was performed using SPSS
software v11.2 (SPSS, Inc.). All experiments were repeated at least
three times and the data are expressed as the mean ± SD.
Differences among groups were analysed using one-way ANOVA followed
by Tukey's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
CK inhibits the viability of RCC
cells
First, the effects of CK on the viability of RCC
cells were investigated. Both Caki-1 and 786-O cells are
well-established RCC cell lines (21). However, 786-O cells are defective in
von Hippel-Lindau disease tumour suppressor (VHL) expression and
have altered HIF and VEGF pathways (21). By contrast, Caki-1 cells harbour
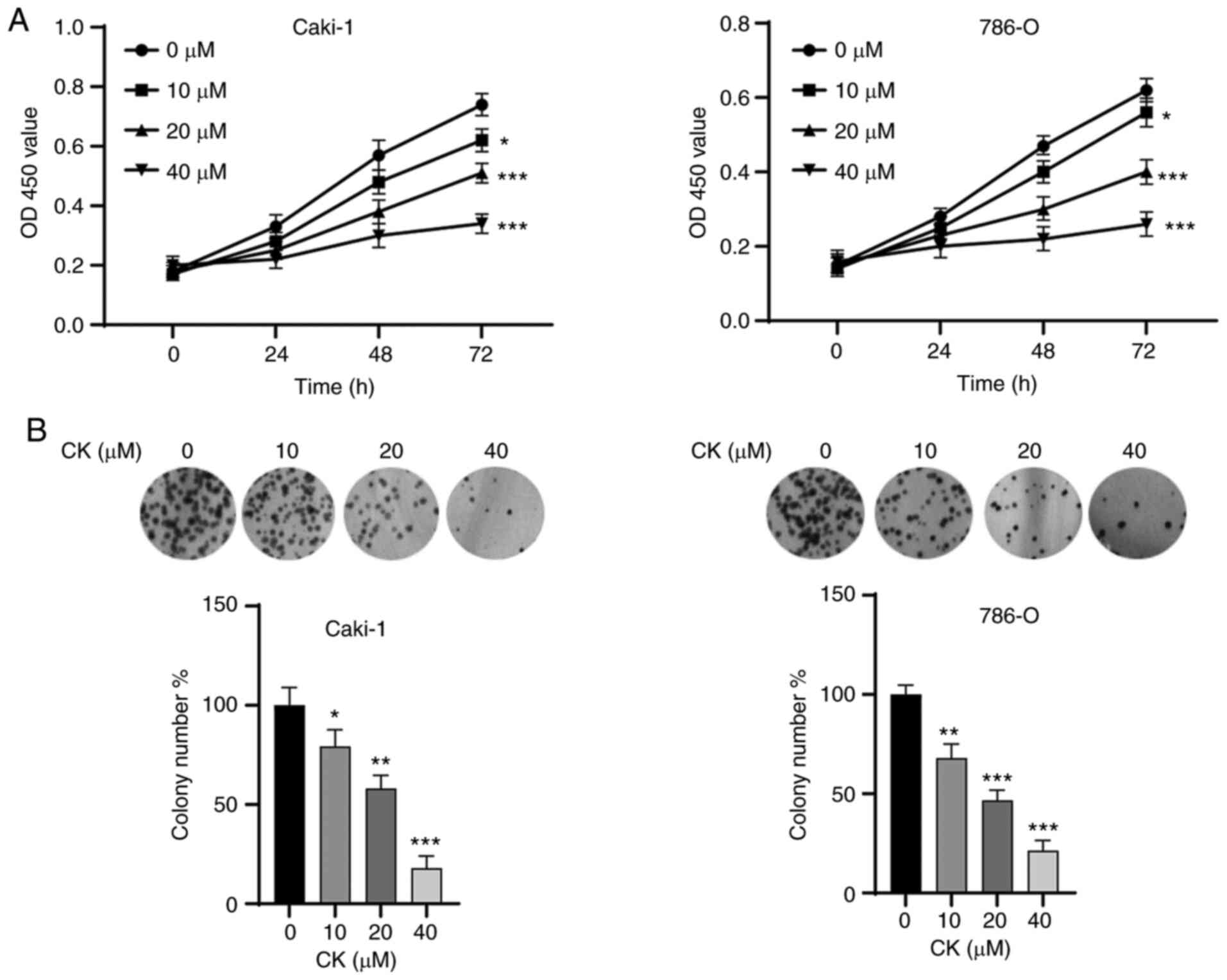
wild-type VHL and exhibit normal expression of VEGF (21). As shown in Fig. 1A, the MTT assay showed that the
viability of Caki-1 and 786-O cells was significantly inhibited by
CK in a time- and dose-dependent manner. Moreover, colony formation
assays also indicated that the viability of Caki-1 and 786-O cells
was significantly suppressed (Fig.
1B). These results suggested the antitumour potential of CK in
RCC cells.
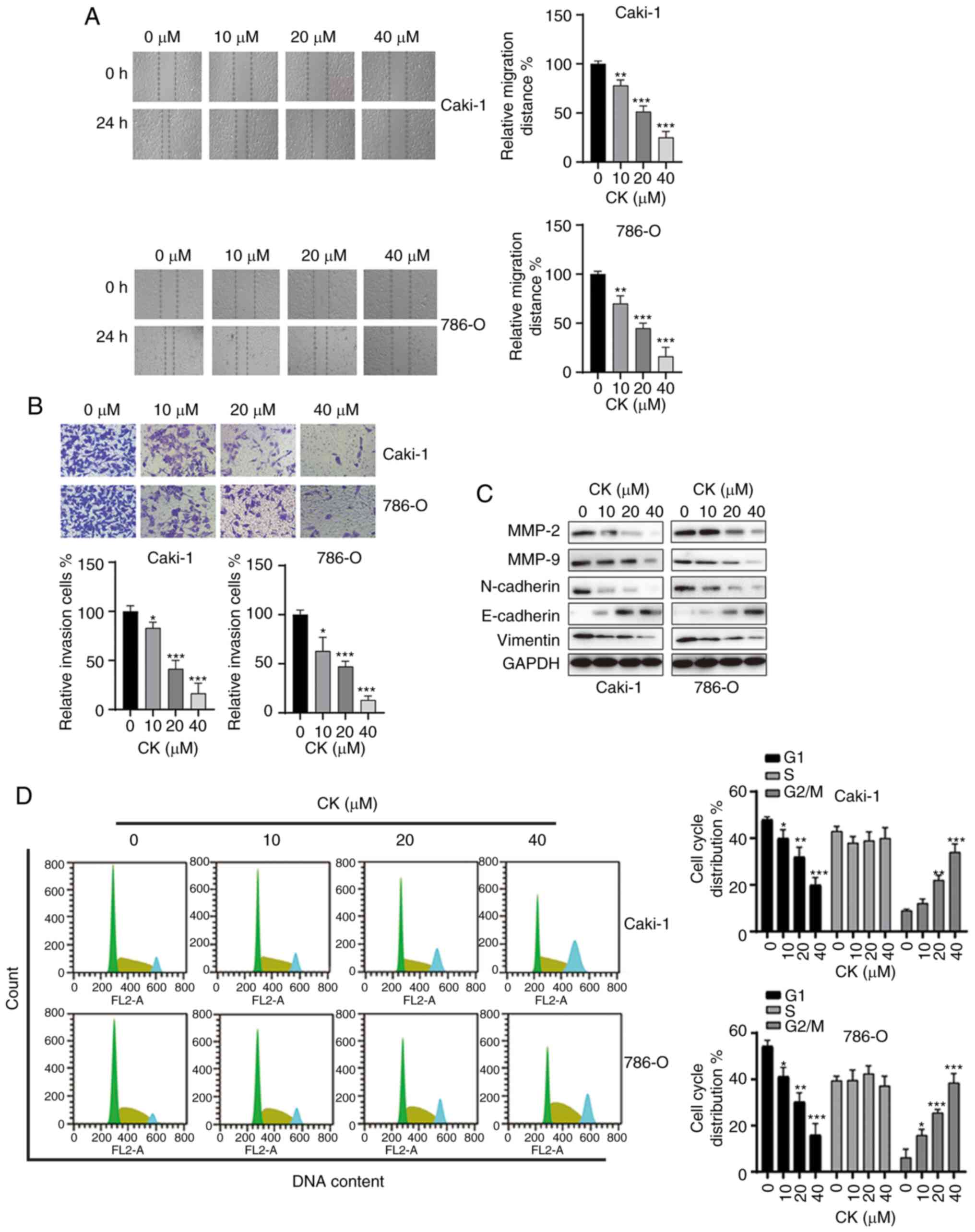
CK inhibits the migration and invasion
of RCC cells and induces cell cycle arrest
It is commonly known that migration and invasion are
essential steps in the metastatic behaviour of cancer cells. Thus,
it was next investigated whether CK could affect the mobility of
RCC cells, and wound healing and Transwell invasion assays were
performed after treatment with various doses of CK for 48 h. As
indicated in Fig. 2A and B, CK
treatment inhibited the migration and invasion of both Caki-1 and
786-O cells in a dose-dependent manner. To further analyse the
inhibitory effects of CK on the migration and invasion of RCC
cells, the expression of cell motility-related proteins were
measured via western blotting. As shown in Fig. 2C, treatment with CK led to the
downregulation of MMP-2, MMP-9, Vimentin and N-cadherin, and the
upregulation of E-cadherin in both RCC cell lines. Moreover, the
effect of CK on the cell cycle distribution of RCC cells was also
examined by PI staining. CK treatment led to cell cycle arrest at
the G2/M phase in RCC cells in a dose-dependent manner (Fig. 2D). Taken together, these data
suggested that CK repressed migration and invasion and induced cell
cycle arrest at the G2/M phase in RCC cells.
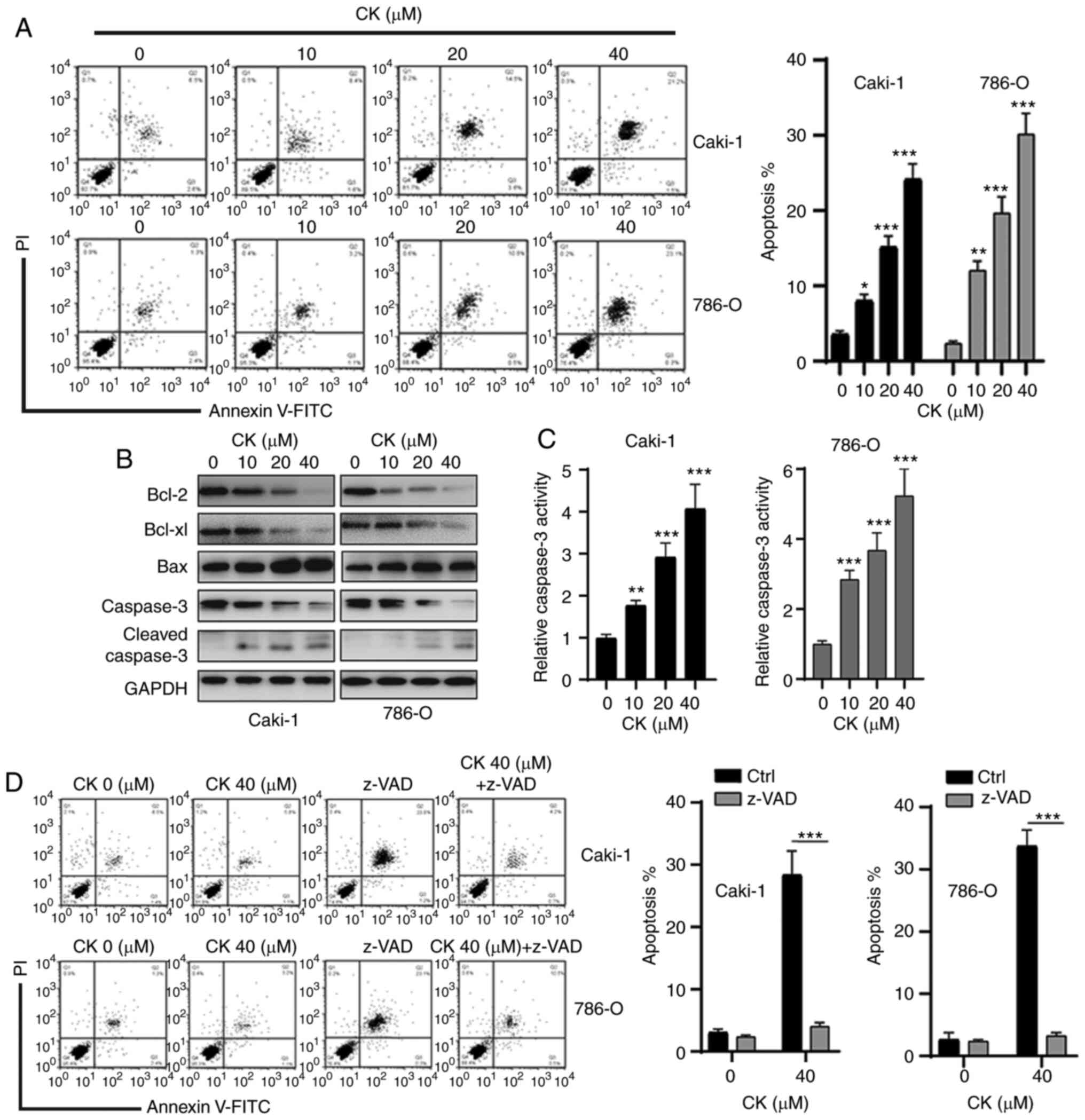
CK induces RCC cell apoptosis in a
caspase-dependent manner
Next, an Annexin V-FITC staining assay was performed
to examine whether CK could induce RCC cell apoptosis. As shown in
Fig. 3A, treatment of Caki-1 and
786-O cells with different doses of CK resulted in increased
apoptosis in a dose-dependent manner. To further confirm the
apoptosis-inducing effects of CK, the expression levels of
apoptosis-related proteins in RCC cells were measured via western
blotting after incubation with various doses of CK for 48 h. As
indicated in Fig. 3B, a
dose-dependent decrease in anti-apoptotic proteins, Bcl-2 and
Bcl-xl, and an increase in pro-apoptotic protein Bax were observed
in RCC cells after treatment with CK. CK treatment also led to an
increase in cleaved caspase-3 (Fig.
3B). In addition, the caspase-3 activity assay also revealed
that the activity of caspase-3 was increased in a dose-dependent
manner after treatment with CK (Fig.
3C). Furthermore, the pan-caspase inhibitor z-VAD could repress
the CK-induced apoptosis of RCC cells (Fig. 3D). Collectively, these data suggested
that CK induced apoptosis in a caspase-dependent manner in RCC
cells.
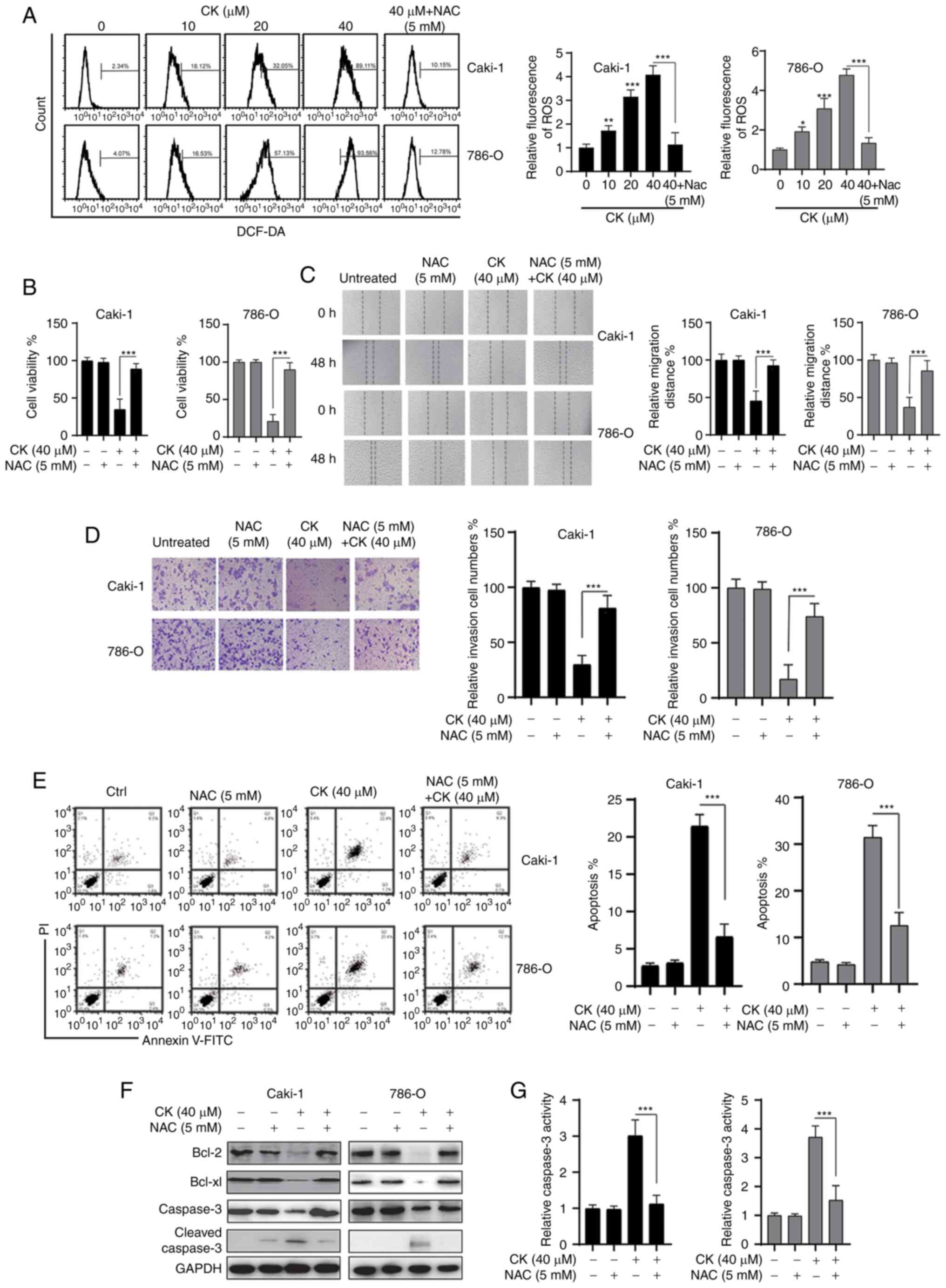
CK treatment led to the generation of
ROS in RCC cells
Previous studies have indicated that CK could
trigger oxidative stress in various cancer cells and tumour models
(22,23). Therefore, the present study
investigated whether CK treatment could lead to the accumulation of
ROS in RCC cells. As shown in Fig.
4A, a dose-dependent increase in ROS was detected in both RCC
cell lines after treatment with CK for 24 h. Notably, the ROS
scavenger NAC significantly prevented the accumulation of ROS after
treatment with CK in RCC cells (Fig.
4A). The MTT assay revealed that the inhibitory effects of CK
on the viability of RCC cells could be blocked by NAC (Fig. 4B). In addition, NAC also repressed the
CK-induced inhibition of migration and invasion in RCC cells
(Fig. 4C and D). Furthermore, the
CK-induced apoptosis could also be blocked by NAC in RCC cells
(Fig. 4E-G). Taken together, these
data suggested that the generation of ROS played an essential role
in the antitumour activity of CK in RCC cells.
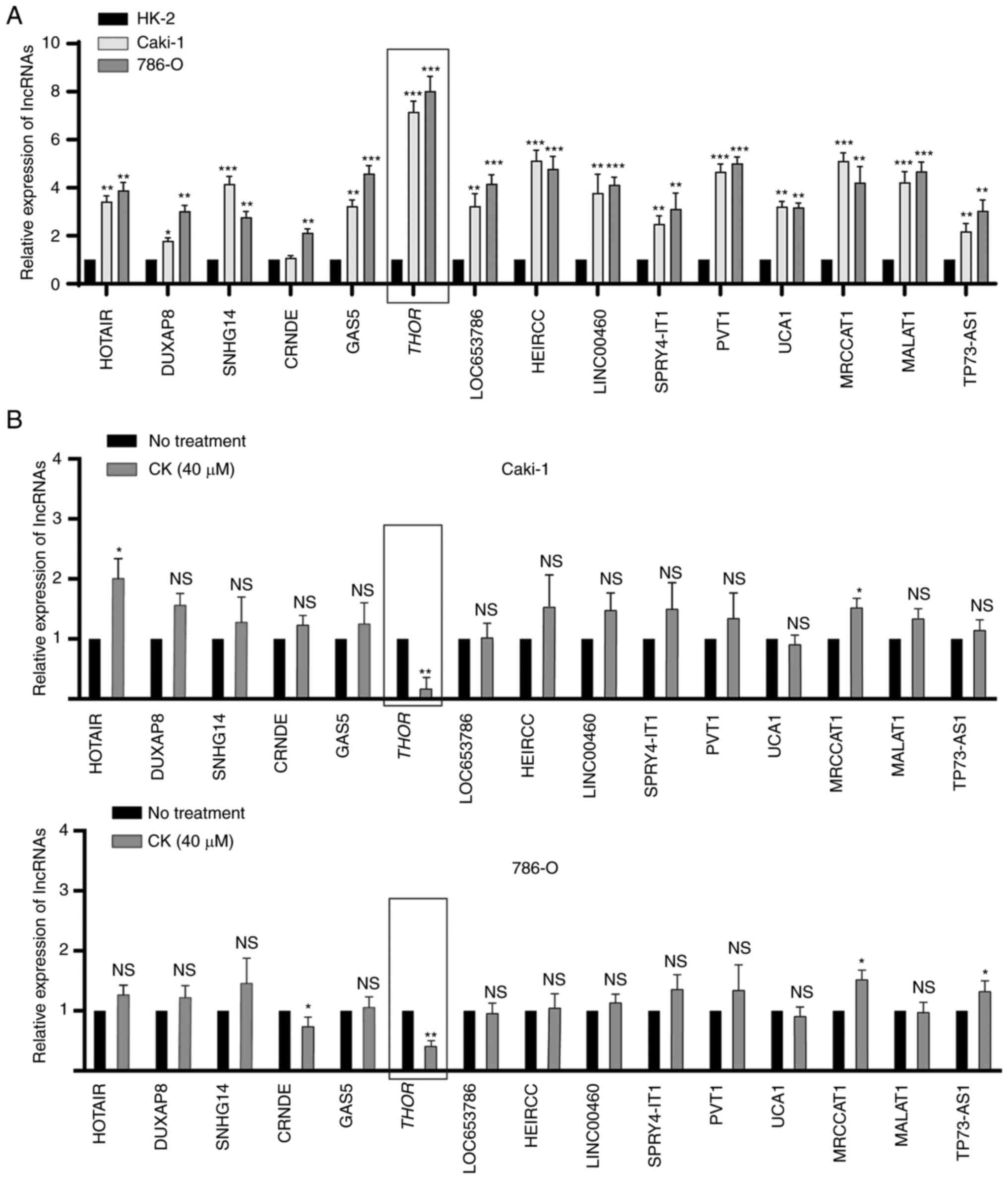
lncRNA THOR is downregulated in
CK-treated RCC cells
lncRNAs are commonly known to promote tumorigenesis
in various cancers, including RCC. To investigate whether any
lncRNAs participated in the antitumour activity of CK in RCC cells,
the RT2 lncRNA PCR Array system was used. The expression
profiles of 25 RCC-related lncRNAs were analysed in HK-2, Caki-1
and 786-O cells, and 15 lncRNAs were found to be significantly
upregulated in RCC cells compared with normal cells (Fig. 5A). Among these 15 lncRNAs, THOR was
the most significantly upregulated in both Caki-1 and 786-O cells
(Fig. 5A). To further confirm this
finding, the expression levels of 15 lncRNAs in the presence or
absence of CK were analysed in Caki-1 and 786-O cells. As indicated
in Fig. 5B, the expression of THOR
could be significantly inhibited by treatment with CK in both RCC
cell lines (Fig. 5B). Taken together,
these findings suggested that THOR, the lncRNA exhibiting the
greatest reduction after CK treatment, may participate in the
antitumour effects of CK. Hence, THOR was chosen for further study
in the following experiments.
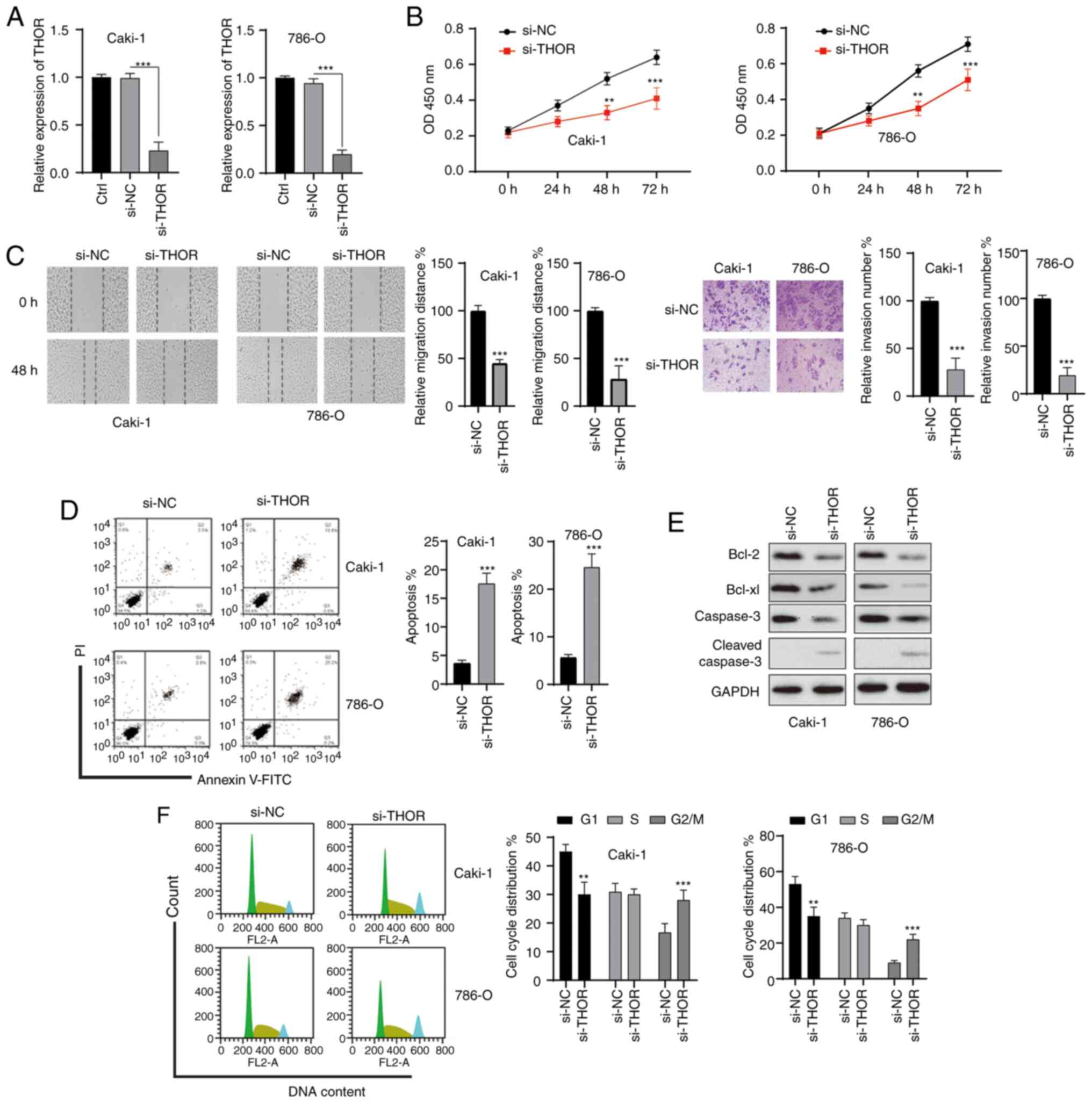
Silencing of THOR inhibits the
oncogenesis of RCC cells
To test whether the effect of CK on RCC cells might
be mediated by THOR, the functions of THOR in RCC cells were
studied. RCC cells were transfected with siRNAs, and satisfactory
transfection efficiency was achieved 48 h after transfection
(Fig. 6A). The MTT assay showed that
knockdown of THOR markedly significantly the viability of both RCC
cell lines (Fig. 6B). In addition,
wound healing and Transwell assays showed that the knockdown of
THOR significantly repressed the migration and invasion of RCC
cells (Fig. 6C). Furthermore, the
silencing of THOR also induced the apoptosis of RCC cells (Fig. 6D). The anti-apoptotic Bcl-2 protein
was downregulated and cleaved caspase-3 was increased after the
silencing of THOR in RCC cells (Fig.
6E). Furthermore, silencing of THOR also induced cell cycle
arrest at the G2/M phase in RCC cells (Fig. 6F). These data revealed that
downregulation of THOR triggered effects on RCC cells similar to
those of CK, suggesting that CK might exert its function by
targeting THOR.
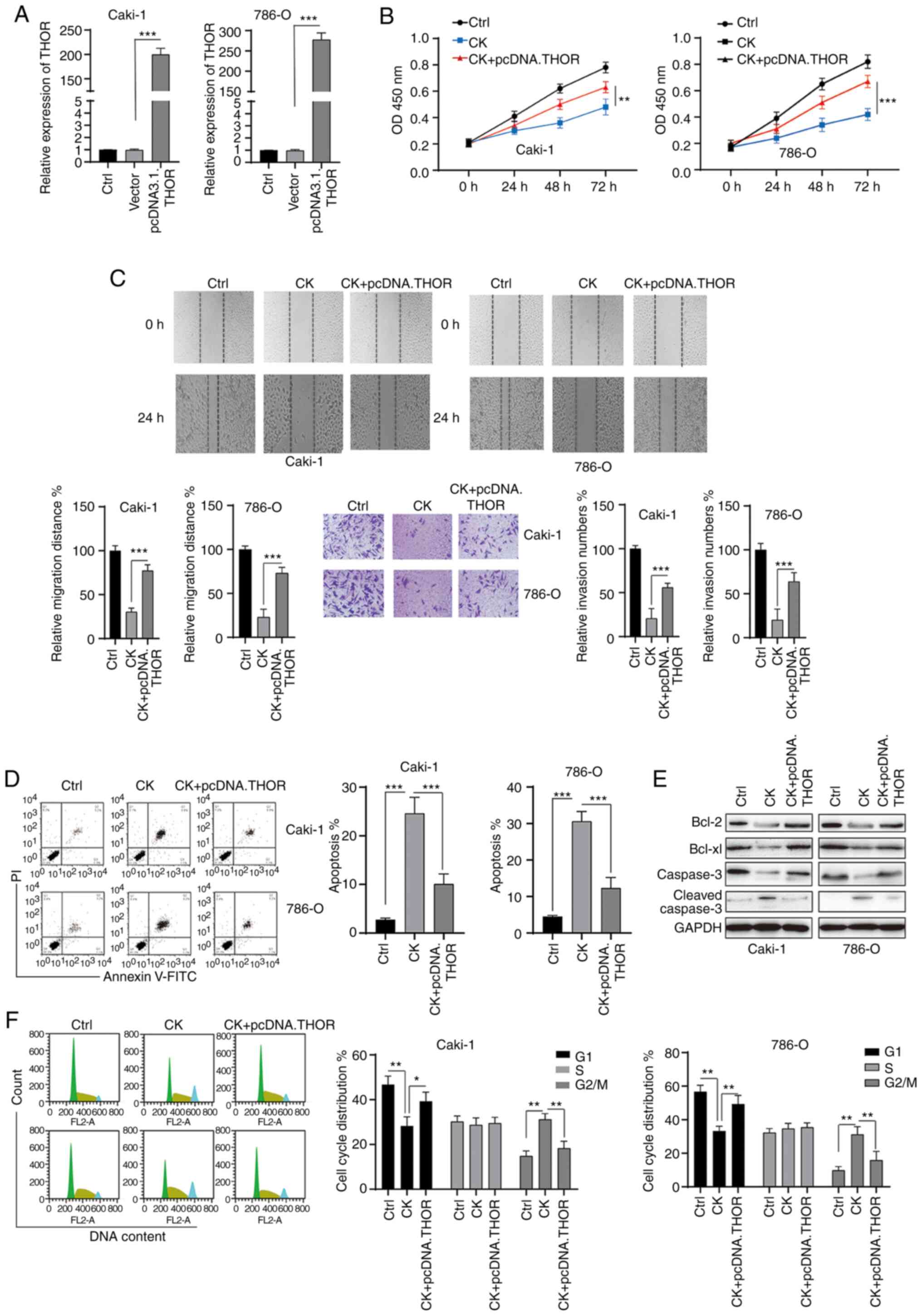
CK exerts its function in RCC cells at
least partially by targeting THOR
To further investigate the role of THOR in the
antitumour functions of CK, rescue experiments were conducted. RCC
cells were transfected with pcDNA3.1 THOR, which significantly
upregulated the expression of THOR (Fig.
7A). The MTT assay revealed that the inhibitory effects of CK
on the viability of RCC cells could be blocked by the
overexpression of THOR (Fig. 7B). In
addition, wound healing and Transwell invasion assays showed that
the effects of CK on RCC cell migration and invasion could be
reversed after the overexpression of THOR (Fig. 7C). The apoptosis-inducing effects of
CK on RCC cells were also blocked by the overexpression of THOR
(Fig. 7D and E). Furthermore, the
CK-mediated cell cycle arrest at the G2/M phase was also reversed
by increased THOR expression (Fig.
7F). Taken together, these data suggested that CK exerted its
antitumour effects at least partially by targeting THOR.
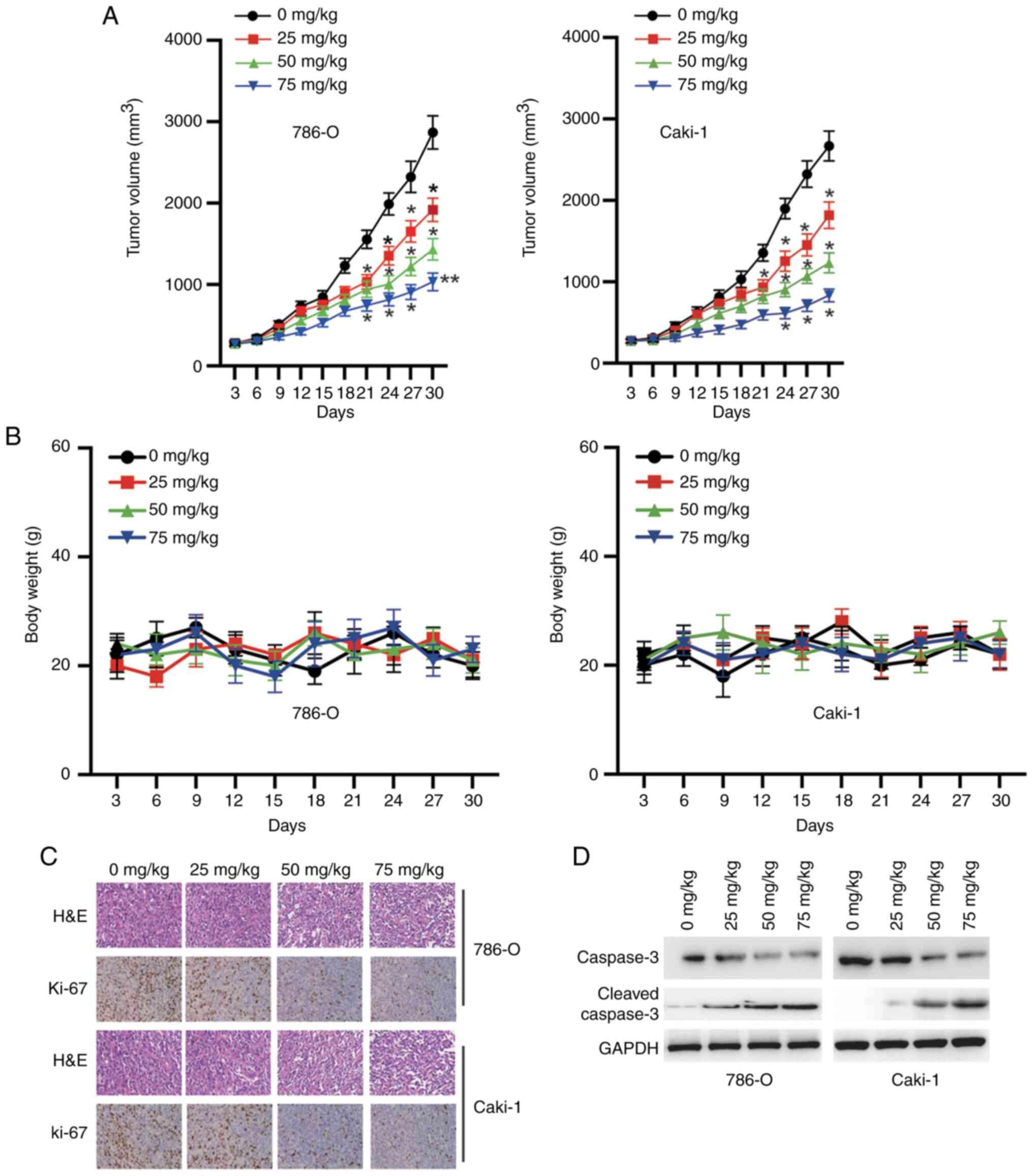
CK inhibits tumorigenesis in nude
mice
Finally, the antitumour activity of CK was evaluated
in male BALB/c mice harbouring established RCC xenografts. The mice
were randomized into four groups (0, 25, 50 or 75 mg/kg) and
treated with CK every 3 days. As shown in Fig. 8A, the mice treated with CK exhibited a
significant reduction in tumour size. Notably, the mice tolerated
all of the treatments, and no obvious differences in body weight
were observed, suggesting that CK was well tolerated (Fig. 8B). Further study showed that CK
treatment inhibited the expression of Ki-67 in a dose-dependent
manner in vivo (Fig. 8C). The
western blotting results also confirmed that treatment with CK
induced the cleavage of caspase-3 in a dose-dependent manner in
vivo (Fig. 8D). Taken together,
these data indicated that CK inhibited the growth of RCC cells
in vivo.
Discussion
In the current study, it was shown that CK, a
metabolite of protopanaxadiol-type ginsenosides produced by gut
microbiota, inhibited the oncogenesis of RCC cells via multiple
mechanisms in vitro. Ginseng, which is derived from the root
of P. ginseng Meyer, has been used for the treatment of
malignant diseases in Eastern Asia for thousands of years. However,
the effects of this herb are hindered by oral administration
because it is not easily absorbed by the body (24). By contrast, CK, a ginsenoside
metabolite of ginseng, was found to be more easily absorbed by the
metabolic system (25). A number of
reports have studied the inhibitory effects of CK on various
cancers, such as lung cancer, breast cancer, glioma, liver cancer
and colon cancer (10,11,26–28).
Consistent with previous studies, the present study demonstrated
that CK inhibited the growth of two RCC cell lines (Caki-1 and
786-O) in a dose- and time-dependent manner. Clonogenic, migration
and Transwell invasion assays also confirmed the inhibitory effect
of CK on RCC cells. These findings suggested that CK may have
potential as a potent therapeutic agent for RCC.
Next, the molecular mechanisms of the antitumour
effects of CK were systematically investigated. Mounting evidence
has indicated that tumour recurrence and metastasis are major
obstacles in the treatment of cancer. Epithelial to mesenchymal
transition (EMT) is a biological process that is often activated
during cancer invasion and metastasis (29). In the present study, CK treatment led
to the downregulation of the mesenchymal markers N-cadherin and
Vimentin and the upregulation of the epithelial marker E-cadherin.
These data suggested that CK treatment inhibited EMT in RCC cells.
This finding is consistent with a previous study that found that CK
inhibited EMT in breast cancer (28).
Targeting EMT also plays a critical role in the response of RCC to
chemo- and radiotherapies (30). It
would be useful to test the effects of CK combined with other
therapeutics on RCC.
Various agents mainly exert their antitumour effects
by inducing apoptosis. Similar to previous studies, CK was also
found to induce apoptosis in RCC cells (13,22).
Apoptosis can be triggered via two pathways, namely, the extrinsic
and intrinsic pathways (16). The
process of apoptosis is subject to regulation by various proteins,
such as Bcl-2 family members, mitochondrial proteins and caspases
(31). In the present study, it was
found that treatment with CK inhibited the expression levels of
Bcl-2 and Bcl-xl, and increased the expression levels of Bax and
cleaved caspase-3, which characterize the intrinsic pathway of
apoptosis. Hence, these data suggested that CK induced apoptosis in
RCC cells via the intrinsic pathway.
A number of studies have suggested that the
accumulation of ROS plays a critical role in the antitumour effects
of natural products, including ginsenosides (32). Consistent with a previous study, it
was also observed that CK-treated RCC cells have an increased level
of intracellular ROS compared with untreated cells (32). In addition, NAC, a ROS scavenger,
significantly inhibited ROS generation and prevented the cell death
caused by CK treatment. Moreover, the inhibitory effects of CK on
the viability, migration and invasion of RCC cells were also
reversed by NAC. These data further confirmed the role of ROS
generation in the antitumour effects of CK.
lncRNAs are a group of RNAs that lack the ability to
encode proteins and are >200 nucleotides in length. lncRNAs have
been implicated in the processes of various human diseases,
including cancer (33). To examine
the possible role of lncRNAs in the antitumour effects of CK, the
RT2 lncRNA PCR Array system was used in the present
study. After treatment with CK, the lncRNA that exhibited the
greatest reduction, namely, THOR, was chosen for further
investigation. THOR has been found to be dysregulated in various
cancers, including RCC (34). Similar
to a previous report, the present study also found that the
downregulation of THOR led to the repression of tumorigenesis in
RCC cells (34). Rescue experiments
showed that the antitumour effects of CK could be partially
reversed by the overexpression of THOR. These findings indicated
that CK exerted its effects by targeting THOR. To the best of our
knowledge, the present study is the first to show that lncRNAs are
involved in the pharmacological activities of CK.
There are, however, some limitations to this study.
The mechanism by which CK decreased the expression of THOR in RCC
cells remains unknown, and it would be of note to investigate the
potential underlying mechanisms. In addition, it would also be
useful to test the effects of CK alone or in combination with other
agents against RCC in clinical trials.
In summary, the antitumour activity of CK were
evaluated in RCC cells. The mechanistic investigations showed that
CK inhibited EMT and induced apoptosis in a caspase-dependent
manner in RCC cells. Furthermore, these findings showed that the
generation of ROS and downregulation of THOR were also involved in
the effects of CK on RCC cells. Collectively, these findings
provided some novel insights into the functions of CK, which may be
used as a potential antitumour agent for the treatment of renal
cancer.
Acknowledgements
The authors would like to acknowledge Dr Zhipeng Xu
(Nanjing University; Nanjing, China) for his helpful suggestions
during the present study.
Funding
The present study was supported by the Ningbo
Natural Science Foundation (grant no. 2019A610259) and K.C. Wong
Magna Fund of Ningbo University.
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SC, HY and FG made contributions to the acquisition
and analysis of data. SC, SM and CL made contributions to the
analysis of data. SC, CL and BX made contributions to the
interpretation of data. YR and RY designed the study, and made
contributions to the acquisition of funding and drafting of the
manuscript. SC, HY and FG confirm the authenticity of all the raw
data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The protocol was approved by the Institutional
Animal Care and Use Committee of Zhongda Hospital, Southeast
University (approval no. 2017043321; Nanjing, China). All
experiments involving animals were conducted following the Guide
for the Care and Use of Laboratory Animals published by the
National Institutes of Health.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Motzer RJ, Bander NH and Nanus DM:
Renal-Cell carcinoma. N Engl J Med. 335:865–875. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fontana F, Raimondi M, Marzagalli M, Di
Domizio A and Limonta P: The emerging role of paraptosis in tumor
cell biology: Perspectives for cancer prevention and therapy with
natural compounds. Biochim Biophys Acta Rev Cancer.
1873:1883382020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cho CW, Kim YC, Kang JH, Rhee YK, Choi SY,
Kim KT, Lee YC and Hong HD: Characteristic study on the chemical
components of Korean curved ginseng products. J Ginseng Res.
37:349–354. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang XD, Yang YY, Ouyang DS and Yang GP: A
review of biotransformation and pharmacology of ginsenoside
compound K. Fitoterapia. 100:208–220. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gan XT and Karmazyn M: Cardioprotection by
ginseng: Experimental and clinical evidence and underlying
mechanisms. Can J Physiol Pharmacol. 96:859–868. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Oh J and Kim JS: Compound K derived from
ginseng: Neuroprotection and cognitive improvement. Food Funct.
7:4506–4515. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hasegawa H, Sung JH, Matsumiya S and
Uchiyama M: Main ginseng saponin metabolites formed by intestinal
bacteria. Planta Med. 62:453–457. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim H, Roh HS, Kim JE, Park SD, Park WH
and Moon JY: Compound K attenuates stromal cell-derived growth
factor 1 (SDF-1)-induced migration of C6 glioma cells. Nutr Res
Pract. 10:259–264. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee S, Kwon MC, Jang JP, Sohng JK and Jung
HJ: The ginsenoside metabolite compound K inhibits growth,
migration and stemness of glioblastoma cells. Int J Oncol.
51:414–424. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shin DH, Leem DG, Shin JS, Kim JI, Kim KT,
Choi SY, Lee MH, Choi JH and Lee KT: Compound K induced apoptosis
via endoplasmic reticulum Ca2+ release through ryanodine
receptor in human lung cancer cells. J Ginseng Res. 42:165–174.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen HF, Wu LX, Li XF, Zhu YC, Wang WX, Xu
CW, Huang ZZ and Du KQ: Ginsenoside compound K inhibits growth of
lung cancer cells via HIF-1α-mediated glucose metabolism. Cell Mol
Biol (Noisy-le-grand). 65:48–52. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen Y, Xu Y, Zhu Y and Li X: Anti-Cancer
effects of ginsenoside compound k on pediatric acute myeloid
leukemia cells. Cancer Cell Int. 13:242013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Law CK, Kwok HH, Poon PY, Lau CC, Jiang
ZH, Tai WC, Hsiao WW, Mak NK, Yue PY and Wong RN: Ginsenoside
compound K induces apoptosis in nasopharyngeal carcinoma cells via
activation of apoptosis-inducing factor. Chin Med. 9:112014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang CZ, Du GJ, Zhang Z, Wen XD, Calway T,
Zhen Z, Musch MW, Bissonnette M, Chang EB and Yuan CS: Ginsenoside
compound K, not Rb1, possesses potential chemopreventive activities
in human colorectal cancer. Int J Oncol. 40:1970–1976.
2012.PubMed/NCBI
|
|
16
|
Yu R, Yu BX, Chen JF, Lv XY, Yan ZJ, Cheng
Y and Ma Q: Anti-Tumor effects of atractylenolide I on bladder
cancer cells. J Exp Clin Cancer Res. 35:402016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yates AD, Achuthan P, Akanni W, Allen J,
Allen J, Alvarez-Jarreta J, Amode MR, Armean IM, Azov AG, Bennett
R, et al: Ensemble 2020. Nucleic Acids Res. 48:D682–D688.
2020.PubMed/NCBI
|
|
19
|
Martinotti S and Ranzato E: Scratch wound
healing assay. Methods Mol Biol. 2109:225–229. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
American Association for Laboratory Animal
Science, . AALAS position statement on the humane care and use of
laboratory animals. Com Med. 4:4132007.
|
|
21
|
Brodaczewska KK, Szczylik C, Fiedorowicz
M, Porta C and Czarnecka AM: Choosing the right cell line for renal
cell cancer research. Mol Cancer. 15:832016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Oh JM, Kim E and Chun S: Ginsenoside
compound K induces ros-mediated apoptosis and autophagic inhibition
in human neuroblastoma cells in vitro and in vivo. Int J Mol Sci.
20:42792019. View Article : Google Scholar
|
|
23
|
Kim AD, Kang KA, Kim HS, Kim DH, Choi YH,
Lee SJ, Kim HS and Hyun JW: A ginseng metabolite, compound K,
induces autophagy and apoptosis via generation of reactive oxygen
species and activation of JNK in human colon cancer cells. Cell
Death Dis. 4:e7502013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qi LW, Wang CZ, Du GJ, Zhang ZY, Calway T
and Yuan CS: Metabolism of ginseng and its interactions with drugs.
Curr Drug Metab. 12:818–822. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Paek IB, Moon Y, Kim J, Ji HY, Kim SA,
Sohn DH, Kim JB and Lee HS: Pharmacokinetics of a ginseng saponin
metabolite compound K in rats. Biopharm Drug Dispos. 27:39–45.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang X, Zhang S, Sun Q, Jiao W, Yan Y and
Zhang X: Compound K induces endoplasmic reticulum stress and
apoptosis in human liver cancer cells by regulating STAT3.
Molecules. 19:14822018. View Article : Google Scholar
|
|
27
|
Yao H, Wan JY, Zeng J, Huang WH,
Sava-Segal C, Li L, Niu X, Wang Q, Wang CZ and Yuan CS: Effects of
compound K, an enteric microbiome metabolite of ginseng, in the
treatment of inflammation associated colon cancer. Oncol Lett.
15:8339–8348. 2018.PubMed/NCBI
|
|
28
|
Zhang K and Li Y: Effects of ginsenoside
compound K combined with cisplatin on the proliferation, apoptosis
and epithelial mesenchymal transition in MCF-7 cells of human
breast cancer. Pharm Biol. 54:561–568. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan
A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Piva F, Giulietti M, Santoni M, Occhipinti
G, Scarpelli M, Lopez-Beltran A, Cheng L, Principato G and
Montironi R: Epithelial to mesenchymal transition in renal cell
carcinoma: Implications for cancer therapy. Mol Diagn Ther.
20:111–117. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Boice A and Bouchier-Hayes L: Targeting
apoptotic caspases in cancer. Biochim Biophys Acta Mol Cell Res.
1867:1186882020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Vallejo MJ, Salazar L and Grijalva M:
Oxidative stress modulation and ROS-mediated toxicity in cancer: A
review on in vitro models for plant-derived compounds. Oxid Med
Cell Longev. 2017:45860682017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Huarte M: The emerging role of lncRNAs in
cancer. Nat Med. 21:1253–1261. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ye XT, Huang H, Huang WP and Hu WL: LncRNA
THOR promotes human renal cell carcinoma cell growth. Biochem
Biophys Res Commun. 501:661–667. 2018. View Article : Google Scholar : PubMed/NCBI
|