Introduction
Colorectal cancer (CRC) is a common malignancy
associated with high prevalence and mortality rates (1,2). In 2018,
1.8 million new CRC cases were diagnosed worldwide (3). Mortality occurs in >600,000 patients
with CRC annually (4), and thus CRC
is considered a major cause of cancer-related mortality (5), as well as a serious threat to public
health worldwide. Despite the substantial progress in early
screening and therapeutic strategies, the survival of patients with
CRC remains poor as a result of cancer metastasis. For instance,
the 5-year survival rate of patients with CRC with distal
metastasis is 12.5%, while it increases to ~90% in patients without
metastasis (6,7). Therefore, it is important to identify
the metastasis mechanism of CRC.
Circular RNAs (circRNAs), characterized by a
closed-loop structure (8), are
extensively expressed in human cells and serve post-transcriptional
regulatory roles in genes expression (9). Moreover, numerous circRNAs contain
microRNA (miRNA/miR) binding sites (10), and these circRNAs act as miRNA sponges
(11) and regulate gene expression
via competing endogenous RNA (ceRNAs) mechanisms (12–14).
circRNAs have been reported to participate in cancer progression.
For example, circZNF609 acts as a miR-134-5p sponge to modulate
B-cell translocation gene 2 expression, resulting in the repression
of glioma cell proliferation and migration capacity (15). In addition, the elevated expression of
circ-ASH2L in pancreatic cells and cancer tissues accelerates tumor
progression via sponging miR-34a to upregulate Notch1 (16). At present, it has been revealed that
circRNAs can regulate CRC cell aggressive features and cancer
progression. For example, circ_0001178 promotes CRC progression and
metastasis via sponging miR-382/587/616 (17). circ CBL.11 also inhibits CRC cell
proliferation ability via regulating YWHAE by sponging miR-6778-5p
(18).
As a vital circRNA, the regulatory action of
circ-FOXO3 in cancer progression is an interesting research topic.
For instance, Du et al (19)
observed that circ-FOXO3 was decreased in tumor tissues, and
overexpression of circ-FOXO3 resulted in tumor cell apoptosis.
Moreover, circ-FOXO3 is downregulated in esophageal squamous cell
cancer (ESCC) cells and tissues, and circ-FOXO3 overexpression
restrains ESCC development by modulating miR-23a and PTEN (20). However, the effect of circ-FOXO3 on
CRC remains unknown. Therefore, the aim of the present study was to
investigate the role of circ-FOXO3 in CRC progression and identify
the underlying mechanism.
Materials and methods
Patient section and cell culture
A total of 70 patients (age, 42–77 years; 24 males
and 46 females) with CRC undergoing surgical resection between
March 2017 and May 2018 in The Third Affiliated Hospital of
Southern Medical University were enrolled in the current study.
None of the patients underwent preoperative chemoradiotherapy.
After all patients signed informed consent forms, the tumor tissues
and healthy tissues adjacent to the tumor were collected and stored
at −80°C for subsequent experiments. The Ethics Committee of The
Third Affiliated Hospital of Southern Medical University approved
the study.
CRC cell lines, HT29 (cells were confirmed using
short tandem repeat profiling), HCT116, HCT8, LOVO, SW480 and
SW620, and wild-type colon epithelial cell line, FHC, were
purchased from the American Type Culture Collection. HT29, HCT116,
HCT8, LOVO, SW480 and SW620 cells were maintained in RPMI medium
(Gibco; Thermo Fisher Scientific, Inc.) containing 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.) in a 37°C incubator (Thermo Fisher
Scientific, Inc.) with 5% CO2. FHC cells were cultured
in DMEM/F12 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS, 10 mM HEPES, 5 µg/ml insulin, 10 ng/ml
cholera toxin, 5 µg/ml transferrin and 100 ng/ml hydrocortisone
(all Gibco; Thermo Fisher Scientific, Inc.) in a 37°C incubator
with 5% CO2.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from CRC tissues and cells
using TRIzol® reagent (Sigma-Aldrich; Merck KGaA) and
transcribed into the corresponding cDNA at 37°C for 15 min followed
by 10 sec at 85°C using PrimeScript RT reagent kit with gDNA Eraser
(Takara Biotechnology Co., Ltd.) and miR-X miRNA First-Strand
Synthesis kit (Takara Biotechnology Co., Ltd.), according to the
manufacturer's instructions. The qPCR reaction was conducted using
SYBR Green PCR Master mix (Takara Biotechnology Co., Ltd.) and
MiR-X miRNA RT-qPCR TB Green kit (Takara Biotechnology Co., Ltd.),
according to the manufacturer's instructions. The PCR conditions
were as follows: Initial denaturation at 95°C for 5 min, followed
by 40 cycles at 95°C for 10 sec, 60°C for 30 sec and 72°C for 10
sec, and a final extension at 72°C for 10 min. The sequences of
primers were: circ-FOXO3 forward (F),
5′-GTGGGGAACTTCACTGGTGCTAAG-3′ and reverse (R),
5′-GTCGTATCCAGTGCAGGGT-3′ (21);
miR-543 F, 5′-CTCCCTCCCGAATTTGAAG-3′ and R,
5′-GTCAGAGGGAGAGGTCAG-3′ (22); large
tumor suppressor kinase 1 (LATS1) F, 5′-CCACCCTACCCAAAACATCTG-3′
and R, 5′-CGCTGCTGATGAGATTTGAGTAC-3′ (23); GAPDH F, 5′-CATGAGAAGTATGACAACAGCCT-3′
and R, 5′-AGTCCTTCCACGATACCAAAGT-3′ (21); and U6 F, 5′-GCGCGTCGTGAAGCGTTC-3′ and
R, 5′-GTGCAGGGTCCGAGGT-3′ (22).
GAPDH and U6 were used as internal references. The relative RNA
expression was analyzed using the 2−ΔΔCq method
(24).
RNase R digestion assay
Total RNA was extracted using TRIzol®
reagent (Sigma-Aldrich; Merck KGaA), and then 5 µg RNA was
incubated with or without 3 U/µg RNase R (Epicentre
Biotechnologies; Illumina, Inc.) at 37°C for 15 min. The resulting
RNA was purified using an RNeasy MinElute cleaning kit (Qiagen
China Co., Ltd.) according to the manufacturer's instructions.
Then, the expression levels of circ-FOXO3 and FOXO3 were determined
using RT-qPCR assay. The primers of FOXO3 used in this study were:
F, 5′-CGGCTAGCTGCGCCTTGGCTTTATAACT-3′ and R,
5′-GGCTCGAGCCCTCCTTCACTGCTACTGG-3′ (25).
Nuclear-cytoplasmic fractionation
HT29 and HCT116 cells (1×107) were lysed
using cell fractionation buffer (Thermo Fisher Scientific, Inc.),
followed by centrifuged at 16,000 × g for 5 min at 4°C to separate
nuclear and cytoplasmic fractions. The obtained nuclear and
cytoplasm were used for extracting RNA using a PARIS kit (Thermo
Fisher Scientific, Inc.), following the manufacturer's protocol.
The extracted RNA was subjected to RT-qPCR to determine the
expression of circ-FOXO3.
Cell transfection
The pcDNA circ-FOXO3, miR-543 mimic
(5′-AAACAUUCGCGGUGCACUUCUU-3′), miR-543 inhibitor
(5′-UACUUAAUGAGAAGUUGCCCGUGUUUUUUUCGCUUUAUUUGUGACGAAACAUUCGCGGUGCACUUCUUUUUCAGUAU-3′),
small interfering (si)-LATS1 (5′-GCAATCAGTTAACCGCAAA-3′) and
indicated controls, including pcDNA vector, miRNA mimic negative
control (mimic NC) (5′-UUUGUACUACACAAAAGUACUG-3′), miRNA inhibitor
NC (inhibitor NC) (5′-ACUACUGAGUGACAGUAGA-3′), siRNA NC (si-NC)
(5′-GCACAGTTAACCGCATAAA-3′) were obtained from Guangzhou RiboBio
Co., Ltd. For cell transfection, cells (1×106) were
inoculated into 6-well plates and maintained for 24 h at 37°C,
followed by transfection with 100 nM pcDNA circ-FOXO3, miR-543
mimic, miR-543 inhibitor or si-LATS1 using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). After 48 h of transfection, HT29 and HCT116
cells were harvested for subsequent experiments. The transfection
efficiency was determined using RT-qPCR and cell fluorescence.
For cell fluorescence, cells were fixed with 4%
paraformaldehyde for 10 min at room temperature and incubated with
Cy3-labeled miR-543 probe in hybridization buffer at 37°C
overnight. The nuclei were stained with DAPI for 5 min at room
temperature. Cell fluorescence were captured under the confocal
microscope at ×40 magnification (Carl Zeiss AG).
Cell Counting Kit-8 (CCK-8) assay
HT29 and HCT116 cells were inoculated into 96-well
plates (1×103 cells/well) and cultured at 37°C. At
indicated time points (0, 24, 48 and 72 h), HT29 and HCT116 cells
in 96-well plates were further incubated with 10 µl CCK-8 reagent
(Sigma-Aldrich; Merck KGaA) according to the manufacturer's
instructions for 2 h at 37°C. Cell viability was calculated based
on the absorbance at 450 nm.
Colony formation assay
HT29 and HCT116 cells were made into a cell
suspension and inoculated into 6-well plates (200 cells/well).
After culturing for 14 days, HT29 and HCT116 cells were fixed using
4% paraformaldehyde (Sigma-Aldrich; Merck KGaA) for 15 min at room
temperature and stained with 0.1% crystal violet (Sigma-Aldrich;
Merck KGaA) for 15 min at room temperature. The number of colonies
(>50 cells) was assessed manually.
Transwell migration and invasion
assays
Transwell inserts with 8 µm pore size (Corning,
Inc.) were inserted into 6-well plates. HT29 and HCT116 cells
(1×105 cells/well) were inoculated into the upper
chamber of Transwell and cultured in serum-free medium (Gibco;
Thermo Fisher Scientific, Inc.). For cell invasion assays, the
upper chamber of Transwell was covered with 50 µl Matrigel (BD
Biosciences) at 37°C for 2 h, which was the only difference to the
migration assay. Complete medium supplemented with 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.) was added to the lower chamber of
Transwell. After 24 h of cultivation, cells were fixed with 4%
paraformaldehyde (Sigma-Aldrich; Merck KGaA) for 20 min at room
temperature and stained with 0.1% crystal violet (Sigma-Aldrich;
Merck KGaA) for 10 min at room temperature. The number of migrated
and invaded cells was determined under a light microscope (Zeiss
AG) in five random fields (magnification, ×100).
Bioinformatics analysis
The circInteractome database (26) (https://circinteractome.nia.nih.gov/) was screened to
identify miRNAs that were adsorbed by circ-FOXO3. miRanda (27) (http://www.microrna.org/) was used to search the
target genes of miR-543.
Luciferase assay
The wild-type (WT) and mutant type (MUT) fragments
of circ-FOXO3, and WT and MUT type 3′-untranslated region (3′-UTR)
of LATS1 were synthesized and constructed into the pGL3 vector
(Promega Corporation). Then, 100 nM circ-FOXO3 WT, circ-FOXO3 MUT,
LATS1 WT 3′-UTR, LATS1 MUT 3′-UTR, miR-NC and miR-543 mimic were
transfected into HT29 and HCT116 cells using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.).
After 48 h of transfection, the luciferase activity was determined
using a microplate reader (Thermo Fisher Scientific, Inc.) and
normalized to Renilla luciferase activity.
RNA pull-down assay
Biotinylated-miR-543 probe and biotinylated NC
(Bio-NC) probe were generated by Guangzhou RiboBio Co., Ltd. The
RNA pull-down was performed as previously described (28). To produce probe-coated beads, miR-543
(Guangzhou RiboBio Co., Ltd.) was incubated with C-1 magnetic beads
(Thermo Fisher Scientific, Inc.) for 2 h at 25°C. The cell lysates
were then harvested and treated with the 50 pmol miR-543 probe or
Bio-NC probe overnight at 4°C. After washing with wash/binding
buffer, the RNA complexes adsorbed in the beads were collected and
used to conduct RT-qPCR assay and northern blot analysis. For
northern blot analysis, 30 µg RNAs were separated in a 1%
agarose-formaldehyde gel and transferred to Hybond-N+ membrane
(Beyotime Institute of Biotechnology). Then, the membranes were
hybridized with digoxin-labeled DNA oligonucleotides specific to
circ-FOXO3 (Guangzhou RiboBio Co., Ltd.) at 37°C. The membranes
were exposed to phosphorimager screens and analyzed using Image Lab
V3.0 software (Bio-Rad Laboratories, Inc.).
Western blotting
Proteins were isolated from HT29 and HCT116 cells
using RIPA buffer (Thermo Fisher Scientific, Inc.) and quantified
using the BCA method. The extracted proteins (40 µg) were then
separated on 10% SDS-PAGE and transferred to a PVDF membrane
(Thermo Fisher Scientific, Inc.). After blocking with 4% skimmed
milk for 1 h at room temperature, the membrane was probed with
rabbit anti-human polyclonal LATS1 antibody (1:500; cat. no.
ab70562), mouse anti-human monoclonal E-cadherin antibody (1:500;
cat. no. ab76055), rabbit anti-human polyclonal N-cadherin antibody
(1:500; cat. no. ab76057), rabbit anti-human polyclonal Vimentin
antibody (1:500; cat. no. ab137321), mouse anti-human monoclonal
MMP9 antibody (1:500; cat. no. ab119906) and rabbit anti-human
polyclonal GAPDH antibody (1:1,000; cat. no. ab9485; all Abcam) at
4°C for 12 h. After the washing steps, the membrane was incubated
with HRP-conjugated rabbit polyclonal anti-human IgG H&L
(1:1,000; cat. no. ab6759) or HRP-conjugated mouse monoclonal
anti-human IgG H&L (1:1,000; cat. no. ab436; both Abcam) at
room temperature for 1 h. GAPDH was used as the loading control.
The blots were visualized using ECL western blotting substrate
(Thermo Fisher Scientific, Inc.).
Statistical analysis
SPSS Statistics 22.0 (IBM Corp.) was utilized to
conduct data analysis. Data are presented as the mean ± SD.
Student's t-test or one-way ANOVA with Tukey's post hoc test were
used for analyzing the significance between different groups. A
paired t-test was used to compare the difference between tumor
tissues and adjacent tissues. The relationship between circ-FOXO3
expression and the clinicopathological parameters of patients with
CRC was determined using a χ2 test. The overall survival
of patients with CRC was determined using Kaplan-Meier analysis
with the log-rank test. The correlations among the expression
levels of circ-FOXO3, miR-543 and LATS1 were determined via
correlation analysis with a Pearson test. All experiments were
repeated at least three times. P<0.05 was considered to indicate
a statistically significant difference.
Results
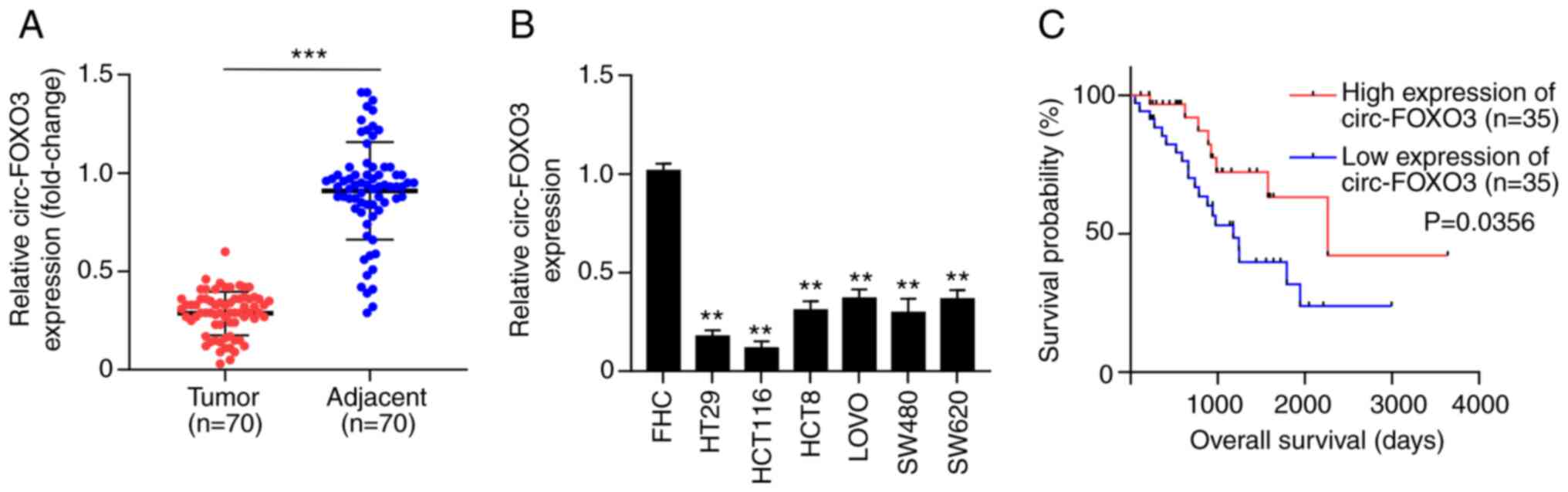
circ-FOXO3 is downregulated in CRC
tissues and cells
To assess the effects of circ-FOXO3 on CRC, the
aberrant expression of circ-FOXO3 in CRC tissues was investigated.
A significant downregulation of circ-FOXO3 expression was
identified in tumor tissues compared with healthy tissues of 70
patients with CRC (P<0.001; Fig.
1A). In addition, circ-FOXO3 expression in CRC cells, including
HT29, HCT116, HCT8, LOVO, SW480 and SW620, was significantly lower
compared with wild-type colon epithelial cell line (all P<0.01;
Fig. 1B).
To determine the association between circ-FOXO3 and
clinical characteristics of patients with CRC, 70 patients with CRC
were divided into the circ-FOXO3 high expression group and
circ-FOXO3 low expression group based on the median value of
circ-FOXO3 expression in CRC tissues as the cut-off value.
Kaplan-Meier analysis demonstrated that the low expression of
circ-FOXO3 was positively associated with poor overall survival of
patients with CRC (P=0.0356; Fig.
1C). In addition, circ-FOXO3 expression was significantly
associated with tumor size (P<0.01), distant metastasis
(P=0.0003), differentiation (P=0.0002), lymph node metastasis
(P<0.0001) and TMN stages (P=0.0081) (Table I). Therefore, these results suggested
that circ-FOXO3 was downregulated in CRC, and associated with the
overall survival, tumor size, distant metastasis, differentiation,
lymph node metastasis and TMN stages of patients with CRC.
 | Table I.Association between circ-FOXO3
expression and clinicopathological parameters of patients with
CRC. |
Table I.
Association between circ-FOXO3
expression and clinicopathological parameters of patients with
CRC.
| Clinicopathological
characteristics | Total | High
expression | Low expression | χ2 | P-value |
|---|
| Sex |
|
|
|
|
|
|
Male | 24 | 11 | 13 | 0.06341 | 0.8012 |
|
Female | 46 | 24 | 22 |
|
|
| Age, years |
|
|
|
|
|
|
≤60 | 37 | 20 | 17 | 0.2293 | 0.632 |
|
>60 | 33 | 15 | 18 |
|
|
| Tumor size |
|
|
|
|
|
| T1 | 17 | 14 | 3 | 16.33 | 0.001 |
| T2 | 15 | 10 | 5 |
|
|
| T3 | 16 | 6 | 10 |
|
|
| T4 | 22 | 5 | 17 |
|
|
| Distant
metastasis |
|
|
|
|
|
|
Positive | 33 | 13 | 20 | 12.87 | 0.0003 |
|
Negative | 37 | 22 | 15 |
|
|
|
Differentiation |
|
|
|
|
|
|
High | 26 | 6 | 20 | 17.37 | 0.0002 |
|
Moderate | 21 | 10 | 11 |
|
|
|
Poor | 23 | 19 | 4 |
|
|
| Lymph node
metastasis |
|
|
|
|
|
|
Positive | 38 | 6 | 26 | 20.78 | <0.0001 |
|
Negative | 32 | 29 | 9 |
|
|
| TNM stages |
|
|
|
|
|
| I | 19 | 15 | 4 | 11.8 | 0.0081 |
| II | 14 | 8 | 6 |
|
|
|
III | 15 | 6 | 9 |
|
|
| IV | 22 | 6 | 16 |
|
|
Overexpression of circ-FOXO3
suppresses CRC cell proliferation, migration and invasion
To investigate the effect of circ-FOXO3 on CRC, the
role of circ-FOXO3 in CRC cell features was determined. It was
identified that HT29 and HCT116 cells presented the lowest
circ-FOXO3 expression compared with that in HCT8, LOVO, SW480 and
SW620 cells (Fig. 1B). Thus, HT29 and
HCT116 cells were used for follow-up experiments. First, a RNase R
digestion assay was conducted in HT29 and HCT116 cells to assess
the circular characteristics of circ-FOXO3. circ-FOXO3 was
resistant to RNase R digestion, which indicated that circ-FOXO3 was
a circRNA (P>0.05; Fig. 2A).
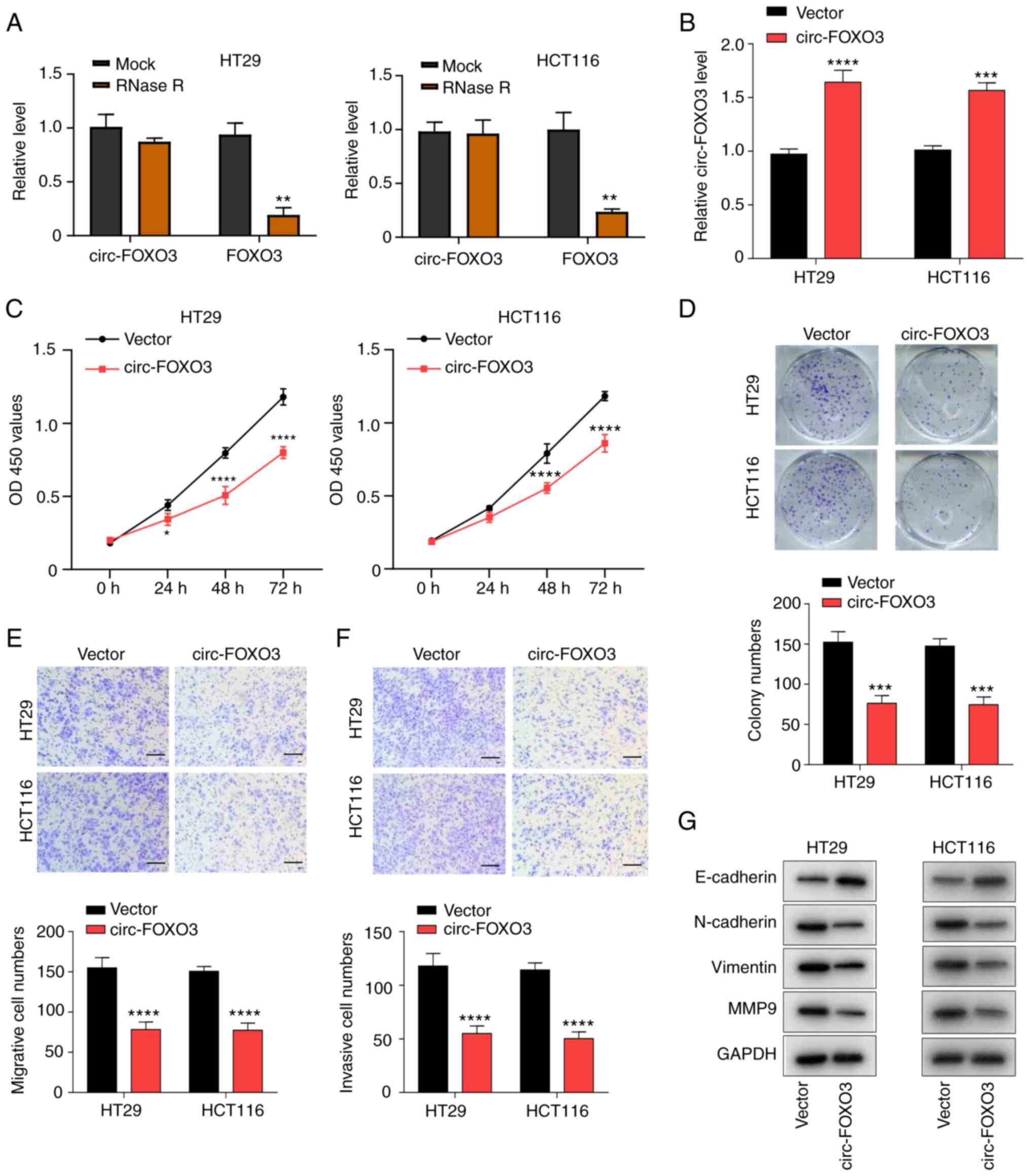 | Figure 2.Overexpression of circ-FOXO3 inhibits
colorectal cancer cell proliferation, migration and invasion. (A)
Circular characteristic of circ-FOXO3 was assessed using RNase R
digestion assay. (B) Reverse transcription-quantitative PCR was
used to detect the transfection efficiency of pcDNA circ-FOXO3 in
HT29 and HCT116 cells. (C) Cell Counting Kit-8 assay measured the
viability of HT29 and HCT116 cells after circ-FOXO3 overexpression.
(D) Colony formation assay determined the colony formation ability
of HT29 and HCT116 cells after overexpression of circ-FOXO3.
Transwell migration assay detected the (E) migratory and (F)
invasive abilities of cells after overexpression of circ-FOXO3.
Magnification, ×100; Scale bar, 100 µm. (G) Western blotting was
conducted to detect the protein expression levels of E-cadherin,
N-cadherin, Vimentin and MMP9 in HT29 and HCT116 cells after
overexpression of circ-FOXO3. All experiments were performed three
times. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001 vs.
Vector group. circ-FOXO3, circular RNA FOXO3; OD, optical
density. |
Subsequently, HT29 and HCT116 cells were transfected
with pcDNA circ-FOXO3 plasmid to overexpress circ-FOXO3 expression,
which were evaluated using RT-qPCR (P<0.0001 and P<0.001;
Fig. 2B). The effect of circ-FOXO3
overexpression on CRC cell aggressive features was investigated
after circ-FOXO3 overexpression. circ-FOXO3 overexpression
significantly suppressed the viability of HT29 and HCT116 cells
(all P<0.0001; Fig. 2C). Colony
formation results indicated that the colony numbers of HT29 and
HCT116 cells were significantly reduced after overexpression of
circ-FOXO3 (all P<0.001; Fig. 2D).
In addition, Transwell assays results found that circ-FOXO3
overexpression significantly inhibited cell migratory and invasive
abilities (all P<0.0001; Fig. 2E and
F). circ-FOXO3 overexpression also markedly increased the
expression of E-cadherin, and notably decreased the expression
levels of N-cadherin, Vimentin and MMP9 (Fig. 2G). Thus, these findings indicated that
overexpression of circ-FOXO3 suppressed CRC cell proliferation,
migration and invasion.
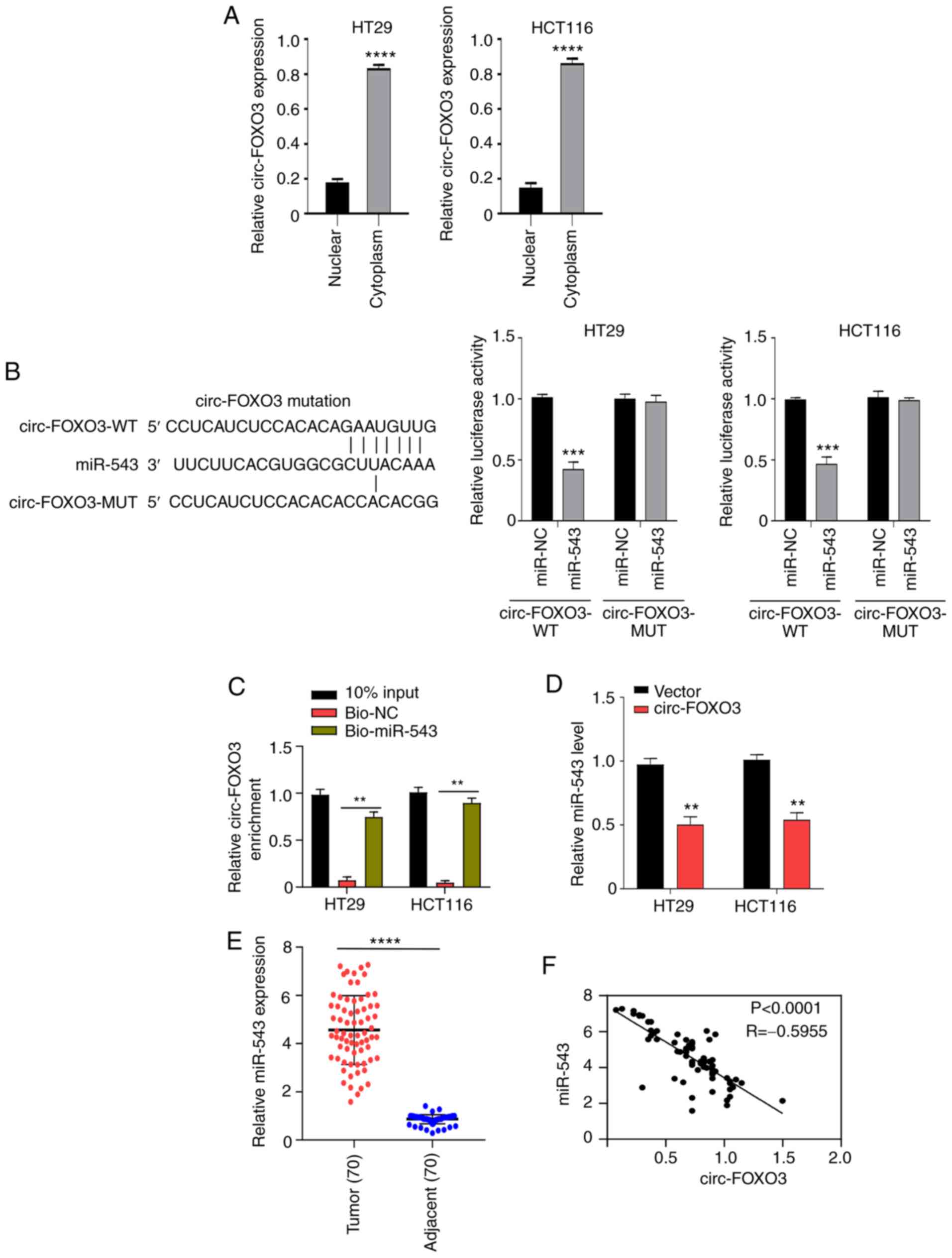
circ-FOXO3 functions as a miR-543
sponge
According to the aforementioned results, circ-FOXO3
presented aberrant expression in CRC and inhibited CRC cell
aggressive features, but the underlying mechanism of their action
remains to be determined. To identify the regulatory mechanism of
circ-FOXO3 on CRC cell features, the location of circ-FOXO3 in CRC
cells was first examined. A significantly higher expression of
circ-FOXO3 was found in the cytoplasm compared with the nucleus of
HT29 and HCT116 cells (all P<0.0001; Fig. 3A), which suggested that circ-FOXO3 was
mostly located in the cytoplasm. circRNA in cytoplasm usually
functions as a miRNA sponge (29).
Therefore, it was hypothesized that circ-FOXO3 may function as a
miRNA sponge in CRC cells to regulated CRC cell aggressive
features.
The circInteractome database (https://circinteractome.nia.nih.gov/) was screened to
identify miRNAs that are adsorbed by circ-FOXO3. A miR-543 binding
site was found in circ-FOXO3, and the predicted sequences of
circ-FOXO3 and miR-543 are presented in Fig. 3B. The transfection efficiency of
miR-543 mimic was demonstrated using RT-qPCR (all P<0.0001;
Fig. S1A). Luciferase assay results
demonstrated that miR-543 overexpression inhibited the relative
luciferase activity in cells transfected with WT circ-FOXO3 (all
P<0.001), while no significant effect was found in cells
transfected with MUT circ-FOXO3 (Fig.
3B). RNA pull-down results indicated a significantly higher
level of enrichment of circ-FOXO3 using the biotinylated-miR-543
probe compared with biotinylated-NC group in HT29 and HCT116 cells
(all P<0.01; Fig. 3C). Moreover,
overexpression of circ-FOXO3 decreased miR-543 expression (all
P<0.001; Fig. 3D).
To further examine the association between miR-543
and circ-FOXO3, miR-543 expression in patients with CRC was
detected, and it was found that miR-543 expression in tumor tissues
was significantly upregulated compared with healthy tissues
(P<0.0001; Fig. 3E). Pearson
correlation analysis demonstrated a moderate negative correlation
between the expression levels of miR-543 and circ-FOXO3 in CRC
(P<0.0001; R=−0.5955; Fig. 3F).
Thus, circ-FOXO3 may function as a miR-543 sponge.
circ-FOXO3 elevates LATS1 expression
via sponging miR-543
Next, miR-543 targets were screened using miRanda
(http://www.microrna.org/). LATS1 was
considered as a potential target of miR-543 and the putative
binding site sequences are presented in Fig. 4A. Luciferase assay was conducted to
identify whether miR-543 targeted LATS1. It was found that miR-543
overexpression suppressed relative luciferase activity in cells
transfected with WT LATS1 (all P<0.001), while no significant
effect was observed in cells transfected with mutant type LATS1
(Fig. 4A).
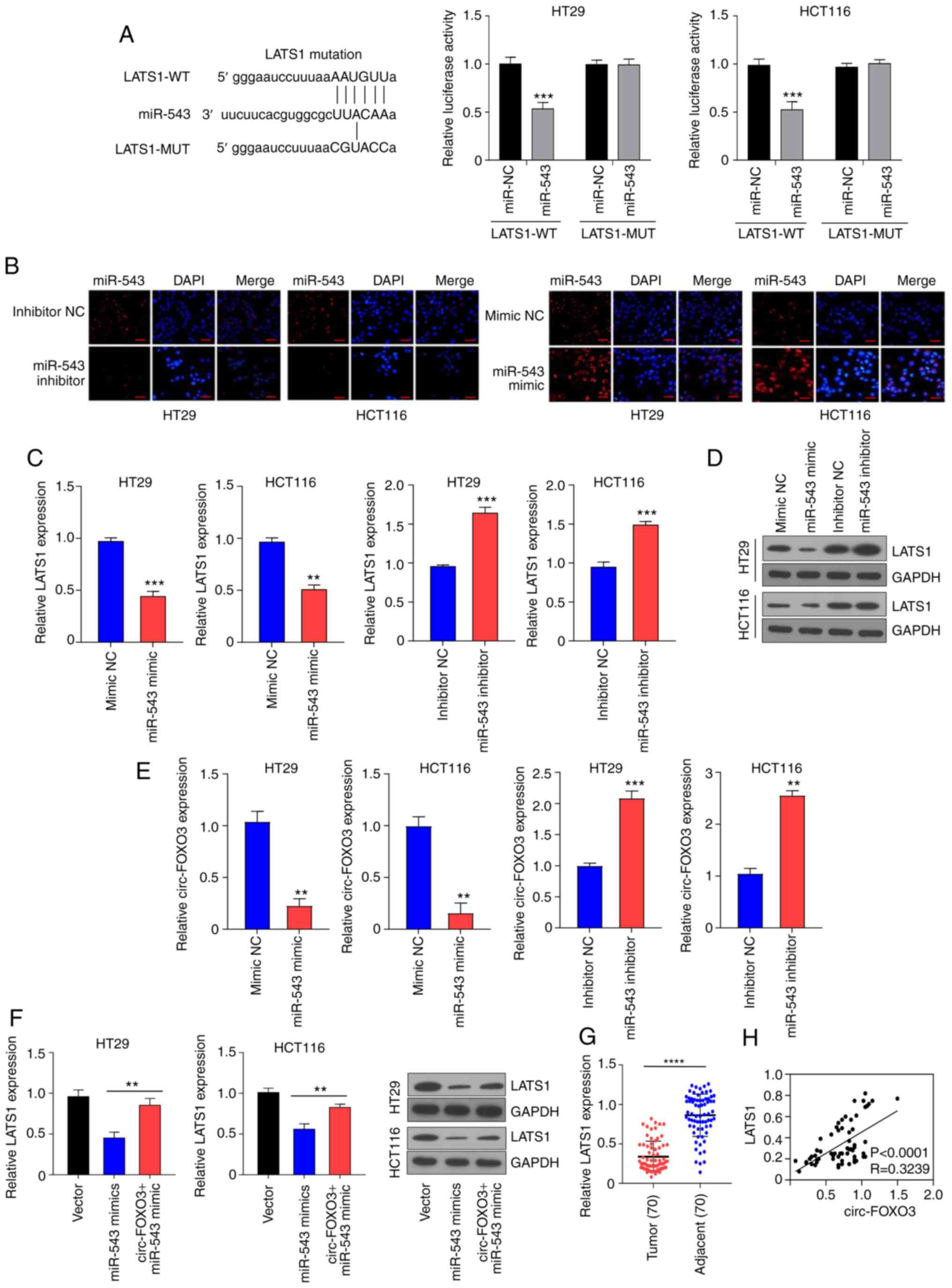 | Figure 4.circ-FOXO3 increases LATS1 expression
via sponging miR-543. (A) Luciferase assay was conducted to verify
whether miR-543 directly binds to LATS1 in HT29 and HCT116 cells.
(B) Transfection efficiency of miR-543 mimic and inhibitor in HT29
and HCT116 cells was determined using cell fluorescence.
Magnification, ×400; Scale bar, 50 µm. (C) mRNA and (D) protein
expression levels of LATS1 in cells transfected with miR-543 mimic
or inhibitor was determined using RT-qPCR and western blotting,
respectively. (E) Expression of circ-FOXO3 in HT29 and HCT116 cells
transfected with miR-543 mimic or inhibitor was determined using
RT-qPCR. (F) mRNA and protein expression levels of LATS1 in cells
transfected with pcDNA circ-FOXO3 or co-transfected with pcDNA
circ-FOXO3 and miR-543 mimic in HT29 and HCT116. (G) RT-qPCR assay
examined the mRNA expression of LATS1 in tumor tissues and adjacent
healthy tissues of 70 patients with CRC. (H) Pearson correlation
analysis determined the correlation between the expression levels
of LATS1 and circ-FOXO3. All experiments were performed three
times. **P<0.01, ***P<0.001, ****P<0.0001 vs. miR-NC
group. circ-FOXO3, circular RNA FOXO3; RT-qPCR, reverse
transcription-quantitative PCR; miR, microRNA; NC, negative
control; WT, wild-type; MUT, mutant; LATS1, Large tumor suppressor
kinase 1. |
The transfection efficiency of miR-543 mimic and
inhibitor in HT29 and HCT116 cells were determined via cell
fluorescence, which indicated that the miR-543 mimic notably
increased miR-543 expression, while miR-543 inhibitor decreased
miR-543 expression (Fig. 4B). In
addition, the transfection efficiency of the miR-543 inhibitor was
verified using RT-qPCR (all P<0.001; Fig. S1B). The miR-543 inhibitor elevated
the mRNA expression of LATS1 in HT29 (P<0.001) and HCT116
(P<0.001) cells, while miR-543 mimic inhibited the mRNA
expression of LATS1 in HT29 (P<0.001) and HCT116 (P<0.01)
cells (Fig. 4C). Similarly, the
protein expression of LATS1 oppositely modulated by miR-543
(Fig. 4D). The miR-543 mimic
decreased circ-FOXO3 mRNA expression (all P<0.01), while miR-543
inhibitor significantly increased the expression of circ-FOXO3
(P<0.01 and P<0.001; Fig. 4E).
Furthermore, the inhibitory effect of miR-543 mimic on LATS1 was
reversed by circ-FOXO3 overexpression in HT29 and HCT116 cells (all
P<0.01; Fig. 4F).
To further evaluate the association between LATS1
and circ-FOXO3, the expression of LATS1 was detected in patients
with CRC, and it was identified that LATS1 was significantly
downregulated in CRC tumor tissues compared with healthy tissues
(P<0.0001; Fig. 4G). In addition,
Pearson correlation analysis demonstrated a weak positive
correlation between the expression levels of circ-FOXO3 and LATS1
(P<0.0001; R=0.3239; Fig. 4H).
Collectively, these results suggested that circ-FOXO3 increased
LATS1 expression via sponging miR-543.
miR-543 overexpression or LATS1
knockdown inhibits circ-FOXO3-induced attenuated CRC aggressive
features
Based on the aforementioned results, it was
suggested that the effect of circ-FOXO3 on CRC aggressive features
may be mediated via miR-543 and LATS1. The transfection efficiency
of si-LATS1 was verified using RT-qPCR (all P<0.001; Fig. S1C). The inhibitory role of circ-FOXO3
overexpression on cell viability was abolished by both miR-543
overexpression and LATS1 knockdown in HT29 and HCT116 cells (all
P<0.001; Fig. 5A). Colony
formation assay results demonstrated that both miR-543
overexpression (all P<0.01) and LATS1 knockdown (P<0.01 and
P<0.05) reversed the inhibitory effect of circ-FOXO3
overexpression on colony formation ability in HT29 and HCT116 cells
(Fig. 5B). Furthermore, Transwell
assay results identified that the inhibited migratory and invasive
abilities exerted by circ-FOXO3 overexpression were blocked by
miR-543 overexpression (P<0.05-P<0.001) or LATS1 knockdown
(P<0.01 and P<0.05) in HT29 and HCT116 cells (Fig. 5C and D). The promotion effect of
circ-FOXO3 overexpression on E-cadherin expression, and its
inhibitory effects on N-cadherin expression were also markedly
reversed by miR-543 overexpression or LATS1 knockdown in HT29 and
HCT116 cells (Fig. 5E). Therefore,
these results indicated that both miR-543 overexpression and LATS1
knockdown inhibited circ-FOXO3-induced attenuated CRC aggressive
features.
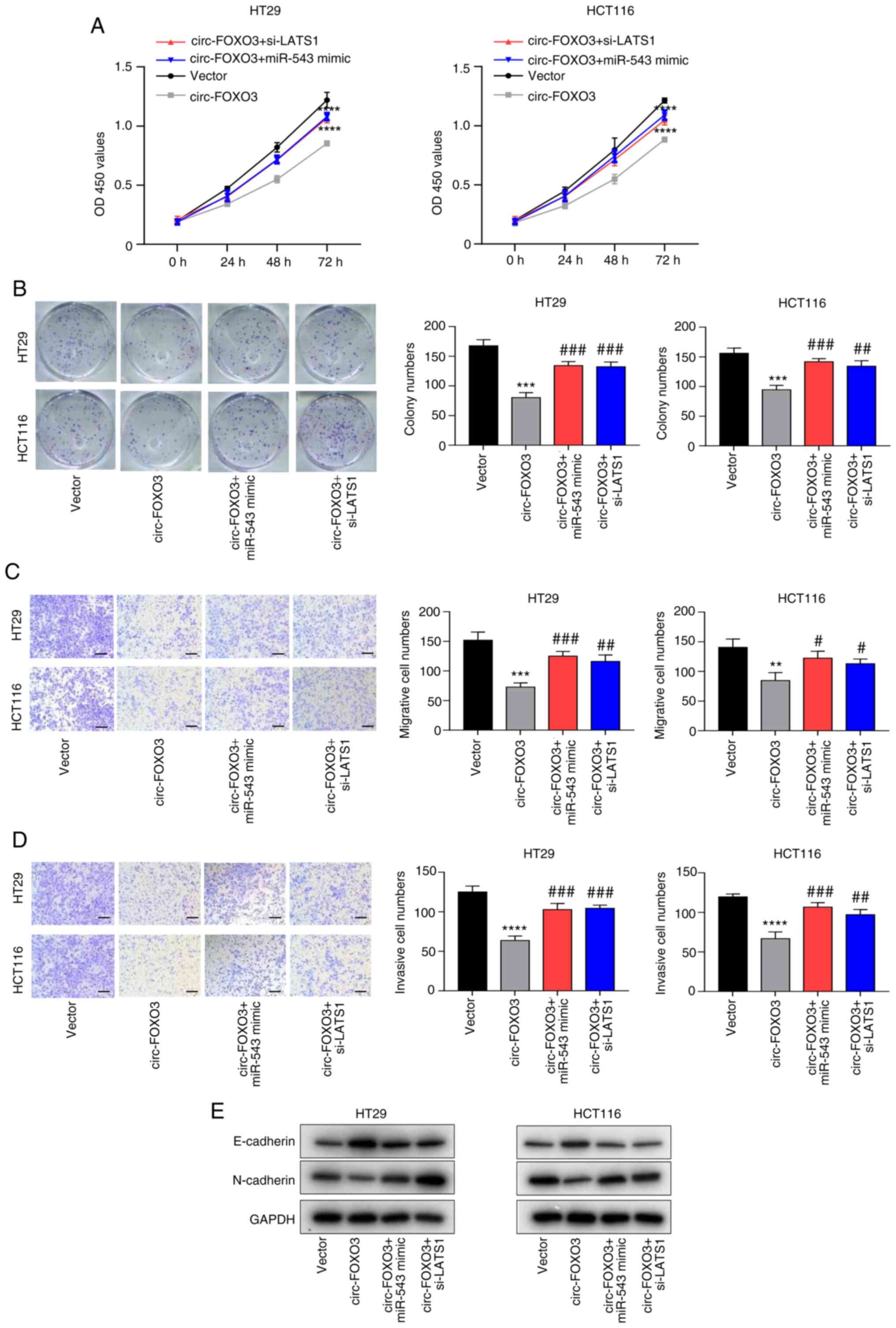 | Figure 5.miR-543 overexpression or LATS1
knockdown blocks circ-FOXO3-induced attenuated CRC aggressive
features. (A) Cell Counting Kit-8 assay was conducted to determine
the viability of HT29 and HCT116 cells after the transfection of
pcDNA circ-FOXO3, miR-543 mimic or si-LATS1. (B) Colony formation
assay determined the colony formation ability of cells after the
transfection of pcDNA circ-FOXO3, miR-543 mimic or si-LATS1.
Transwell migration assay was conducted to assess the (C) migratory
and (D) invasive abilities of HT29 and HCT116 cells after the
transfection of pcDNA circ-FOXO3, miR-543 mimic or si-LATS1.
Magnification, ×100; Scale bar, 100 µm. (E) Western blotting was
performed to detect the protein expression levels of E-cadherin and
N-cadherin in cells after transfection of pcDNA circ-FOXO3, miR-543
mimic or si-LATS1. All experiments were performed three times.
**P<0.01, ***P<0.001, ****P<0.0001 vs. Vector group;
#P<0.05, ##P<0.01,
###P<0.0001 vs. circ-FOXO3 group. circ-FOXO3,
circular RNA FOXO3; LATS1, Large tumor suppressor kinase 1; miR,
microRNA; siRNA, small interfering RNA; OD, optical density. |
Discussion
As a common malignancy, the morbidity and mortality
rates of patients with CRC remain high despite the progress of
early screening and therapeutic strategies (1,2). In 2018,
1.8 million new CRC cases were diagnosed worldwide (3). Mortality occurs in >600,000 patients
with CRC annually, worldwide (4). The
metastasis of cancer contributes to poor survival of patients with
CRC (6,7). Hence, it is important to identify the
metastasis mechanism of CRC. Recently, a previous study revealed
that circRNAs participated in cancer metastasis regulation
(30).
The present study investigated the effect of
circ-FOXO3 on CRC progression. circ-FOXO3 expression was
downregulated in CRC tissues and cells, which was in accordance
with previous studies (19,20). Du et al (19) reported that circ-FOXO3 was reduced in
tumor tissues of patients and cancer cells, while Xing et al
(20) revealed decreased circ-FOXO3
expression in ESCC. Moreover, downregulation of circ-FOXO3 was
associated with poor overall survival of patients with CRC, and was
also found to be associated with tumor size, distant metastasis,
differentiation, lymph node metastasis and TMN stages of patients
with CRC. In line with these findings, Zhou et al (31) observed that circ-FOXO3 expression was
downregulated in patients with de novo acute myeloid
leukemia, and patients with low-expression of circ-FOXO3 had poor
overall survival. Furthermore, the present results indicated that
circ-FOXO3 overexpression suppressed CRC cell proliferation,
migration and invasion in vitro, which were consistent with
previous studies (19,20). For instance, in ESCC, elevated
circ-FOXO3 was found to inhibit cell proliferation, migration and
invasion (20).
circRNAs are generally involved in cancer
progression by acting as miRNA sponges (11). For example, circ-FOXO3 could act as a
miR-23a sponge in ESCC (20). In the
current study, circ-FOXO3 was demonstrated to be a miR-543 sponge
and could negatively regulate the expression of miR-543. miR-543 is
a critical modulator in cancer progression (32–35), and
it can accelerated esophageal cancer metastasis by directly binding
to Phospholipase A2 Group IVA (32).
miR-543 also promotes gastric cancer migration and invasion ability
by targeting speckle type BTB/POZ protein (35).
miRNAs usually modulate cancer progression via
binding to the target genes (36).
Based on the present results, LATS1 was suggested to be a target of
miR-543. LATS1 is a member of the LATS family and it acts as a
tumor suppressor in different cancer types, such as gastric cancer
and breast cancer (37,38). Moreover, LATS1 suppresses cancer cell
proliferative and invasive abilities (39). circRNAs have been revealed to serve as
miRNA sponges and modulate gene expression via ceRNA mechanisms
(11). To determine whether
circ-FOXO3 regulated gene expression via the ceRNAs mechanism, the
relationship among circ-FOXO3, miR-543 and LATS1 was detected. The
present results suggested that circ-FOXO3 elevated LATS1 expression
via sponging miR-543. Furthermore, both miR-543 overexpression and
LATS1 knockdown blocked circ-FOXO3-induced attenuated CRC
aggressive cellular features. Thus, these findings indicated that
overexpression of circ-FOXO3 inhibited CRC progression via elevated
LATS1 expression by sponging miR-543. While the current study
demonstrated the role and mechanism of circ-FOXO3 FoxO3 in CRC, a
limitation of this study was the lack of in vivo
experiments. Therefore, in vivo experiments will be
conducted in a future study.
Collectively, the present study demonstrated that
circ-FOXO3 expression was downregulated in CRC, and its
overexpression inhibited CRC metastasis and progression via
elevated LATS1 by sponging miR-543. Thus, circ-FOXO3 may be a
promising target for CRC therapy.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YYD, YJM, KZ, XLZ and LXL designed the experiment.
YYD, YJM and KZ wrote the manuscript. QSY, MP, ZRC acquired the
data. LXL, HL, MW analyzed the data. XLZ and LXL approved the
manuscript. YYD and LXL are responsible for confirming the
authenticity of the raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
After all patients signed informed consent forms,
the tumor tissues and healthy tissues adjacent to the tumor were
collected for subsequent experiments. The Ethics Committee of The
Third Affiliated Hospital of Southern Medical University approved
the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kemper KE, Glaze BL, Eastman CL, Waldron
RC, Hoover S, Flagg T, Tangka FKL and Subramanian S: Effectiveness
and cost of multilayered colorectal cancer screening promotion
interventions at federally qualified health centers in Washington
State. Cancer. 124:4121–4129. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rawla P, Sunkara T and Barsouk A:
Epidemiology of colorectal cancer: Incidence, mortality, survival,
and risk factors. Prz Gastroenterol. 14:89–103. 2019.PubMed/NCBI
|
|
4
|
Han P, Li JW, Zhang BM, Lv JC, Li YM, Gu
XY, Yu ZW, Jia YH, Bai XF, Li L, et al: The lncRNA CRNDE promotes
colorectal cancer cell proliferation and chemoresistance via
miR-181a-5p-mediated regulation of Wnt/β-catenin signaling. Mol
Cancer. 16:92017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bian Z, Jin L, Zhang J, Yin Y, Quan C, Hu
Y, Feng Y, Liu H, Fei B, Mao Y, et al: LncRNA-UCA1 enhances cell
proliferation and 5-fluorouracil resistance in colorectal cancer by
inhibiting miR-204-5p. Sci Rep. 6:238922016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu X, Li R, Song Q, Zhang C, Jia R, Han Z,
Zhou L, Sui H, Liu X, Zhu H, et al: JMJD2C promotes colorectal
cancer metastasis via regulating histone methylation of MALAT1
promoter and enhancing beta-catenin signaling pathway. J Exp Clin
Cancer Res. 38:4352019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gao P, Wang Z, Hu Z, Jiao X and Yao Y:
Circular RNA circ_0074027 indicates a poor prognosis for NSCLC
patients and modulates cell proliferation, apoptosis, and invasion
via miR-185-3p mediated BRD4/MADD activation. J Cell Biochem.
121:2632–2642. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Feng Y, Yang Y, Zhao X, Fan Y, Zhou L,
Rong J and Yu Y: Circular RNA circ0005276 promotes the
proliferation and migration of prostate cancer cells by interacting
with FUS to transcriptionally activate XIAP. Cell Death Dis.
10:7922019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li J, Yang J, Zhou P, Le Y, Zhou C, Wang
S, Xu D, Lin HK and Gong Z: Circular RNAs in cancer: Novel insights
into origins, properties, functions and implications. Am J Cancer
Res. 5:472–480. 2015.PubMed/NCBI
|
|
11
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cheng Z, Yu C, Cui S, Wang H, Jin H, Wang
C, Li B, Qin M, Yang C, He J, et al: circTP63 functions as a ceRNA
to promote lung squamous cell carcinoma progression by upregulating
FOXM1. Nat Commun. 10:32002019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Song J, Wang HL, Song KH, Ding ZW, Wang
HL, Ma XS, Lu FZ, Xia XL, Wang YW, Fei-Zou and Jiang JY:
CircularRNA_104670 plays a critical role in intervertebral disc
degeneration by functioning as a ceRNA. Exp Mol Med. 50:942018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bai N, Peng E, Qiu X, Lyu N, Zhang Z, Tao
Y, Li X and Wang Z: circFBLIM1 act as a ceRNA to promote
hepatocellular cancer progression by sponging miR-346. J Exp Clin
Cancer Res. 37:1722018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tong H, Zhao K, Wang J, Xu H and Xiao J:
CircZNF609/miR-134-5p/BTG-2 axis regulates proliferation and
migration of glioma cell. J Pharm Pharmacol. 72:68–75. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen Y, Li Z, Zhang M, Wang B, Ye J, Zhang
Y, Tang D, Ma D, Jin W, Li X and Wang S: Circ-ASH2L promotes tumor
progression by sponging miR-34a to regulate Notch1 in pancreatic
ductal adenocarcinoma. J Exp Clin Cancer Res. 38:4662019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ren C, Zhang Z, Wang S, Zhu W, Zheng P and
Wang W: Circular RNA hsa_circ_0001178 facilitates the invasion and
metastasis of colorectal cancer through upregulating ZEB1 via
sponging multiple miRNAs. Biol Chem. 401:487–496. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li H, Jin X, Liu B, Zhang P, Chen W and Li
Q: CircRNA CBL.11 suppresses cell proliferation by sponging
miR-6778-5p in colorectal cancer. BMC Cancer. 19:8262019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Du WW, Fang L, Yang W, Wu N, Awan FM, Yang
Z and Yang BB: Induction of tumor apoptosis through a circular RNA
enhancing Foxo3 activity. Cell Death Differ. 24:357–370. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xing Y, Zha WJ, Li XM, Li H, Gao F, Ye T,
Du WQ and Liu YC: Circular RNA circ-Foxo3 inhibits esophageal
squamous cell cancer progression via the miR-23a/PTEN axis. J Cell
Biochem. 121:2595–2605. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xiao-Long M, Kun-Peng Z and Chun-Lin Z:
Circular RNA circ_HIPK3 is down-regulated and suppresses cell
proliferation, migration and invasion in osteosarcoma. J Cancer.
9:1856–1862. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang H, Guo X, Feng X, Wang T, Hu Z, Que
X, Tian Q, Zhu T, Guo G, Huang W and Li X: MiRNA-543 promotes
osteosarcoma cell proliferation and glycolysis by partially
suppressing PRMT9 and stabilizing HIF-1alpha protein. Oncotarget.
8:2342–2355. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu S, Song L, Zhang L, Zeng S and Gao F:
miR-21 modulates resistance of HR-HPV positive cervical cancer
cells to radiation through targeting LATS1. Biochem Biophys Res
Commun. 459:679–685. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kuo SJ, Liu SC, Huang YL, Tsai CH, Fong
YC, Hsu HC and Tang CH: TGF-β1 enhances FOXO3 expression in human
synovial fibroblasts by inhibiting miR-92a through AMPK and p38
pathways. Aging (Albany NY). 11:4075–4089. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dudekula DB, Panda AC, Grammatikakis I, De
S, Abdelmohsen K and Gorospe M: CircInteractome: A web tool for
exploring circular RNAs and their interacting proteins and
microRNAs. RNA Biol. 13:34–42. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Betel D, Wilson M, Gabow A, Marks DS and
Sander C: The microRNA.org resource: Targets and expression.
Nucleic Acids Res. 36((Database issue)): D149–D153. 2008.PubMed/NCBI
|
|
28
|
Wang K, Long B, Liu F, Wang JX, Liu CY,
Zhao B, Zhou LY, Sun T, Wang M, Yu T, et al: A circular RNA
protects the heart from pathological hypertrophy and heart failure
by targeting miR-223. Eur Heart J. 37:2602–2611. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jin L, Han C, Zhai T, Zhang X, Chen C and
Lian L: Circ_0030998 promotes tumor proliferation and angiogenesis
by sponging miR-567 to regulate VEGFA in colorectal cancer. Res
Square. Oct 20–2020.(Epub ahead of print). doi:
10.21203/rs.3.rs-92165/v1.
|
|
30
|
Meng S, Zhou H, Feng Z, Xu Z, Tang Y, Li P
and Wu M: CircRNA: Functions and properties of a novel potential
biomarker for cancer. Mol Cancer. 16:942017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou J, Zhou LY, Tang X, Zhang J, Zhai LL,
Yi YY, Yi J, Lin J, Qian J and Deng ZQ: Circ-Foxo3 is positively
associated with the Foxo3 gene and leads to better prognosis of
acute myeloid leukemia patients. BMC Cancer. 19:9302019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao H, Diao C, Wang X, Xie Y, Liu Y, Gao
X, Han J and Li S: MiR-543 promotes migration, invasion and
epithelial-mesenchymal transition of esophageal cancer cells by
targeting phospholipase A2 group IVA. Cell Physiol Biochem.
48:1595–1604. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu X, Gan L and Zhang J: miR-543
inhibites cervical cancer growth and metastasis by targeting TRPM7.
Chem Biol Interact. 302:83–92. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu G, Zhou J and Dong M: Down-regulation
of miR-543 expression increases the sensitivity of colorectal
cancer cells to 5-Fluorouracil through the PTEN/PI3K/AKT pathway.
Biosci Rep. 39:BSR201902492019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu J, Wang F, Wang X, He Z and Zhu X:
miRNA-543 promotes cell migration and invasion by targeting SPOP in
gastric cancer. OncoTargets Ther. 11:5075–5082. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang J, Wang G, Chu SJ, Zhu JS, Zhang R,
Lu WW, Xia LQ, Lu YM, Da W and Sun Q: Loss of large tumor
suppressor 1 promotes growth and metastasis of gastric cancer cells
through upregulation of the YAP signaling. Oncotarget.
7:16180–16193. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Furth N, Pateras IS, Rotkopf R, Vlachou V,
Rivkin I, Schmitt I, Bakaev D, Gershoni A, Ainbinder E, Leshkowitz
D, et al: LATS1 and LATS2 suppress breast cancer progression by
maintaining cell identity and metabolic state. Life Sci Alliance.
1:e2018001712018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Deng J, Zhang W, Liu S, An H, Tan L and Ma
L: LATS1 suppresses proliferation and invasion of cervical cancer.
Mol Med Rep. 15:1654–1660. 2017. View Article : Google Scholar : PubMed/NCBI
|