Introduction
Colon cancer remains one of the most prevalent forms
of cancer worldwide (1). Despite
the development of preventive and therapeutic strategies for
reducing morbidity and increasing survival, the 5-year survival
rate for patients with colon cancer varies from 92% for stage I to
11% for patients with metastatic cancers (2). The higher survival rate of colon
cancer in earlier stages offers the potential that prevention as
well as early detection and treatment for colon cancer can improve
patient overall survival. Chemoprevention is considered one of the
most promising approaches for preventing colon cancer and
diminishing its threat and is defined as the administration of
agents to prevent the induction of cancer and to inhibit or delay
the development of invasive cancer (3). Due to their chemical diversity, low
toxicity, and ability to modulate a variety of signal transduction
pathways and cell processes, natural products, particularly
plant-derived substances, have drawn increasing attention as new
chemopreventive candidates (4–6).
Buddlejasaponin IV (BS-IV) (Fig. 1A) is one of the active components of
the aerial part of Pleurospermum kamtschaticum (P.
kamtschaticum) (Hoffmann; Umbelliferae), which is a
perennial edible herb distributed in the fields and mountains of
Asian countries that has been traditionally used to treat cold,
arthritis, atherosclerosis, and impotence (7). BS-IV has been shown to exhibit a
remarkable protective effect against D-galactosamine-induced
hepatotoxicity but not carbon tetrachloride-induced hepatotoxicity
(8), and inhibit
hypercholesterolemia and hyperlipidemia through extrinsic and
intrinsic inducers by reducing oxidative stress in high cholesterol
diet-induced rats (9). BS-IV has
been revealed to inhibit the production of proinflammatory
mediators, including nitric oxide, prostaglandin E2,
tumor necrosis factor-α, interleukin (IL)-1β, and IL-6, and the
expression levels of cyclooxygenase-2, inducible nitric oxide
synthase, and nuclear factor-κB in lipopolysaccharide-stimulated
RAW 264.7 macrophages (10–12). The anti-inflammatory and analgesic
effects of BS-IV have also been observed in animal models (12). Regarding its antitumor activity, the
methanolic extract of P. kamtschaticum induced apoptosis in
colon cancer cells by activating mitochondrial-dependent apoptotic
signaling and upregulating nonsteroidal anti-inflammatory drug
activated gene-1 (NAG-1) apoptotic protein and inhibited the lung
metastasis of colon cancer cells in mice (13). BS-IV induced cell cycle arrest at
the G2/M phase and apoptosis via both mitochondrial-dependent and
death receptor-mediated pathways in immortalized human oral
keratinocytes and human oral squamous cell carcinoma cells
(14,15).
In the present study, the apoptosis-inducing
capability of BS-IV and the underlying molecular mechanism were
investigated and it was found that the inhibition of
integrin-mediated survival signaling, as well as the activation of
mitochondrial-dependent apoptotic signaling, are implicated in
BS-IV-induced colon cancer cell death.
Materials and methods
Reagents and antibodies
BS-IV was generously provided by Professor Hee-Jun
Park (Sangji University, Wonju, Korea) (10). Dulbecco's modified Eagle's medium
(DMEM), fetal bovine serum (FBS), 1% antibiotic-antimycotic
mixture, phosphate-buffered saline (PBS), and trypsin-EDTA were
purchased from Gibco; Thermo Fisher Scientific, Inc.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT),
dimethyl sulfoxide (DMSO), Triton X-100, bovine serum albumin
(BSA), and Tween-80 were purchased from Sigma-Aldrich; Merck KGaA
and protease inhibitor cocktail was purchased from Roche
Diagnostics GmbH. Z-VAD-FMK (cat. no. 627610), MG132 (cat. no.
474790), and calpeptin (cat. no. 03-34-0051) were purchased from
Calbiochem; Merck KGaA. Antibodies were obtained from the following
sources: Bax (cat. no. sc-7480) and Bcl-2 (cat. no. sc-7382)
antibodies and horseradish peroxidase (HRP)-conjugated secondary
antibodies from Santa Cruz Biotechnology, Inc.; caspase-3 (cat. no.
14220), caspase-9 (cat. no. 9508), poly (ADP-ribose) polymerase
(PARP) (cat. no. 9542), Akt (cat. no. 4685) and phosphorylated
(p)-Akt antibodies (cat. no. 4060) from Cell Signaling Technology,
Inc.; NAG-1/GDF15 antibody (cat. no. ab180929) from Abcam; integrin
α2β1 (cat. no. MAB1998), α2 (cat.
no. AB1936) and β1 (cat. no. AB1952) antibodies from
Chemicon International; Thermo Fisher Scientific, Inc.; focal
adhesion kinase (FAK; cat. no. 610088) and p-FAK (Tyr397; cat. no.
611807) antibodies from BD Transduction Laboratories; β-actin
antibody (cat. no. A1978) from Sigma-Aldrich; Merck KGaA; and
secondary fluorescein isothiocyanate (FITC)-conjugated antibody
(cat. no. 6400-08) from SouthernBiotech. All reagents used in the
present study were of analytical grade.
Cell culture
Human colorectal cancer HT-29 cells (cat. no. 30038)
and murine colorectal cancer CT-26 cells (cat. no. 80009; both from
Korea Cell Line Bank) were cultured in DMEM containing 10% FBS and
a 1% antibiotic-antimycotic mixture in a humidified 5%
CO2 incubator at 37°C. Both cell lines were
authenticated by STR profiling and screened periodically for
mycoplasma contamination using a Mycoplasma Detection kit
(MycoAlert™; Lonza Group, Ltd.).
MTT assay
HT-29 cells (5×103 cells/well) were
seeded into 96-well plates and incubated in DMEM with various
concentrations of BS-IV for 2 h at 37°C. Cell viability was
measured using an MTT assay as previously described (13). The purple formazan product was
dissolved with DMSO and absorbance was detected at 570 nm.
DNA fragmentation assay
HT-29 cells were treated with 4, 5 and 6 µM of
BS-IV. After 2 h, the cells were harvested, and DNA was extracted
as previously described (13).
Extracted DNA was separated on a 1.8% agarose gel containing
ethidium bromide at 50 V and then visualized at 300 nm by
ultraviolet transillumination.
Western blot analysis
HT-29 cells (1×106 cells) were treated
for 2 h with 4, 5 and 6 µM of BS-IV. HT-29 cells were also treated
for 2 h with proteasome inhibitor MG132 (5 µM), calpain inhibitor
calpeptin (10 µM), caspase-3 inhibitor Z-VAD-FMK (5 µM) in the
absence or presence of 6 µM BS-IV. Cells were lysed using RIPA
buffer containing protease inhibitor cocktail (Santa Cruz
Biotechnology, Inc.), and the protein concentration was determined
using a BCA protein assay kit (Pierce; Thermo Fisher Scientific,
Inc.) as previously described (13). Protein from the cell lysates (50 µg)
was separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and transferred onto a polyvinylidene difluoride
membrane (MilliporeSigma). Membranes were blocked in 5% skim milk
in Tris-buffered saline (10 mM Tris, pH 8.0 and 150 mM NaCl) with
0.1% Tween-20 (TBS-T) for 2 h at room temperature and then
incubated with primary antibodies (1:1,000) against target
molecules overnight at 4°C. After washing, the blots were incubated
with a 1:5,000 dilution of the respective HRP-conjugated secondary
antibodies for 1 h at room temperature. The targeted proteins were
visualized using an enhanced chemiluminescence detection kit
(Amersham; Cytiva) according to the manufacturer's protocol. The
Band intensities were quantified using an ImageJ analyzer (version
1.8.0; National Institutes of Health) and normalized to the
intensity of β-actin, a loading control.
Caspase-3 activity assay
HT-29 cells were treated with 4, 5 and 6 µM of BS-IV
for 2 h, and caspase-3 activity was measured using a commercially
available caspase activity assay kit (cat. no. E-CK-A311; Enzo Life
Sciences) according to the manufacturer's instructions. Absorbance
was measured at 405 nm using an ELISA plate reader.
Anoikis assay
HT-29 cells (3×106 cells/well) were
plated in culture dishes coated with solution of 1% polyHEMA
(Sigma-Aldrich; Merck KGaA) in 95% ethanol and treated with 4, 5
and 6 µM of BS-IV for 2 h. The attached cells and suspended cells
in culture media were harvested and stained with 0.4% trypan blue
solution. After 3 min incubation at room temperature, cells were
observed and counted under a light microscope at a magnification of
×10.
Cell adhesion assay
HT-29 cells (7.5×104 cells) were seeded
into CytoMatrixTM cell adhesion strips coated with human collagen
type I (cat. no. ECM104) or IV (cat. no. ECM105; both from
Sigma-Aldrich; Merck KGaA) and treated with 4, 5 and 6 µM of BS-IV
or anti-α2β1 integrin antibody. Cell adhesion
to BSA-coated strips served as a negative control. After 1 h of
incubation at 37°C, attached cells were stained with 0.2% crystal
violet in 10% ethanol for 5 min at room temperature. The stained
cells were gently washed with PBS, and the cell-bound stain was
completely solubilized using solubilization buffer (a 50/50 mixture
of 0.1 M NaH2PO4, pH 4.5 and 50% ethanol) for
5 min at room temperature. Absorbance was measured at 570 nm on a
Benchmark microplate reader (Bio-Rad Laboratories, Inc.).
Flow cytometric assay
HT-29 cells were treated with 4, 5 and 6 µM of BS-IV
for 2 h. The cells were collected, washed with PBS containing 1%
BSA, and incubated with anti-α2β1 integrin
antibody (10 µg/ml) for 1 h at room temperature. After washing, the
cells were resuspended in wash buffer (Mg2+ and
Ca2+-free PBS containing 1% BSA) and incubated with a
1:50 dilution of FITC-conjugated secondary antibody for 30 min at
4°C. The cells were suspended in 200 ml of wash buffer and finally
analyzed using a FACSCalibur equipped with CellQuest software
(version 7.5.3; Becton-Dickinson, Inc.).
Detection of deglycosylated
β1 integrin subunit
HT-29 cells (1×106 cells) were treated
with 5 µM BS-IV for 2 h. Cell lysates were treated with peptide:
N-glycosidase F (PNGase F; cat.no. P0704S; New England BioLabs,
Inc.) according to the manufacturer's instructions and then
incubated overnight at 37°C. The cell lysates were subjected to
western blotting using a monoclonal antibody against β1
integrin to assess the effects of deglycosylation.
Murine lung metastasis
Animal studies were approved (approval no.
2020-0193) by the animal ethics committee of Yonsei University
College of Dentistry (Seoul, Korea) and conducted in accordance
with the approved guidelines of the regional authorities according
to Yonsei University animal care regulations. Male BALB/c mice
(n=30, 5 weeks-old, 18±3 g; Orient Co. Ltd.) were provided a
standard laboratory diet and water ad libitum and were
maintained at a temperature of 22±2°C under a 12 h light/dark
cycle. Male BALB/c nude mice were randomly divided into 5 groups,
with 5 mice per group. Mouse colorectal cancer CT-26 cells
(1×105 cells/200 µl PBS) were injected into the tail
vein of Balb/c male mice (n=5) using a 27-gauge needle. Vehicle
(PBS containing 1% DMSO and 1% Tween-80) and BS-IV at 0.01, 0.1, 1,
and 2 mg/kg body weight (BW) were intraperitoneally administered to
mice 30 min before cancer cell injection and once daily for 14 days
after injection. Control mice received PBS and vehicle. The mice
were euthanized by rapid cervical dislocation, and the lobes of the
lungs were separated and fixed in Bouin's solution for 4 h at room
temperature (Sigma-Aldrich; Merck KGaA). The number of tumor
nodules on the lung surfaces was counted under an ocular
micrometer, and the lung weights were also recorded.
Statistical analysis
Data are expressed as the mean ± standard error (SE)
of three independent experiments. Student's t-test was used to
compare the differences between two groups and one-way ANOVA
followed by a Turkey's post hoc test was used to compare
differences between multiple groups using SPSS version 19.0 (IBM
Corp.). P<0.05 was considered to indicate a statistically
significant difference.
Results
BS-IV induces apoptosis in HT-29
cells
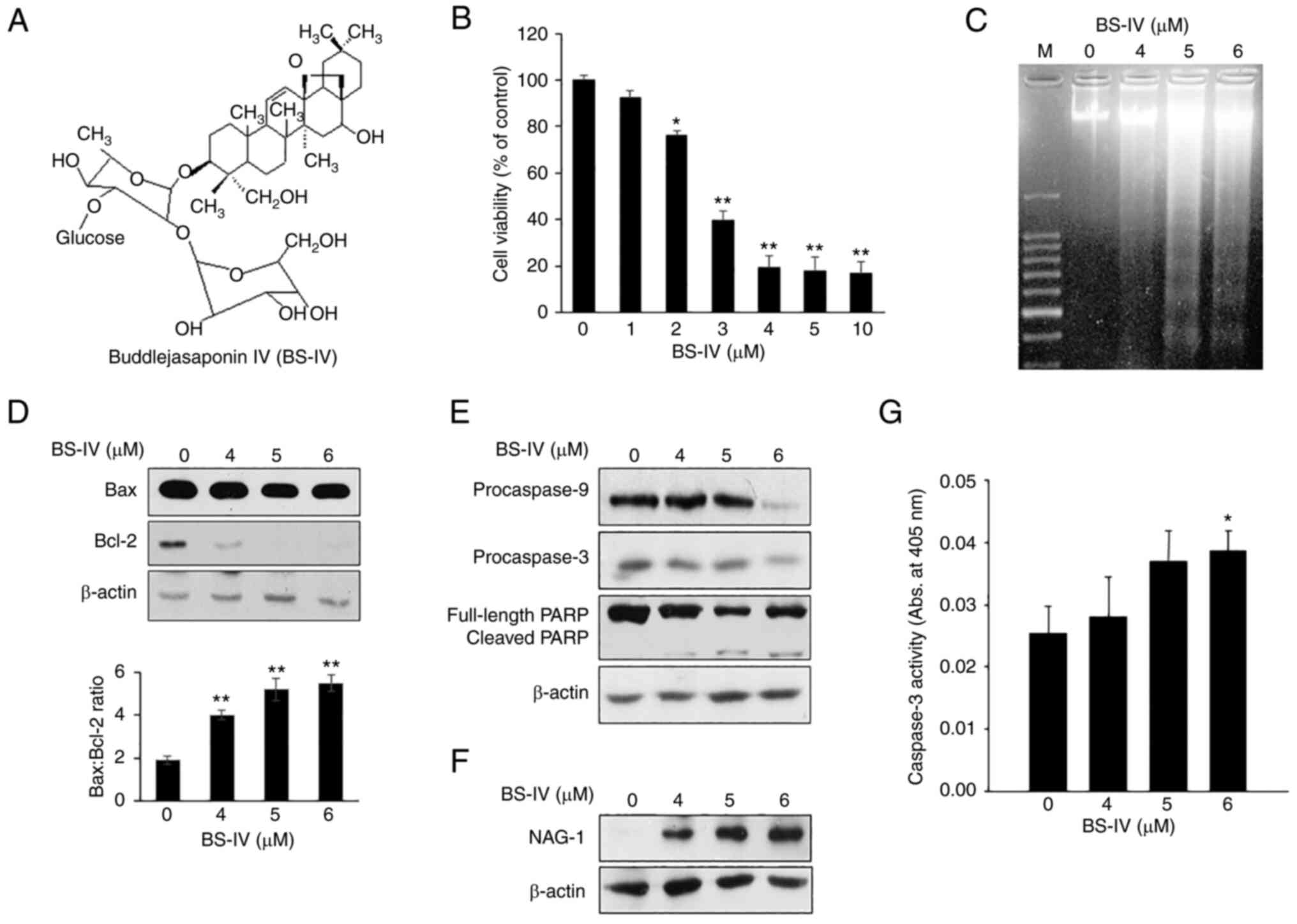
Treatment with BS-IV for 2 h inhibited the viability
of HT-29 colorectal cancer cells by 23.6% at 2 µM, 60.3% at 3 µM,
and 80.5% at 4 µM (Fig. 1B).
Treatment with BS-IV for 2 h displayed a characteristic ladder
pattern of discontinuous DNA fragmentation, a distinct biochemical
hallmark of apoptosis (Fig. 1C).
Western blot analysis showed that treatment with BS-IV for 2 h
weakly reduced pro-apoptotic Bax expression but significantly
suppressed antiapoptotic Bcl-2 expression, resulting in an
increased ratio of Bax to Bcl-2 in HT-29 cells in a dose-dependent
manner (Fig. 1D). Expression levels
of procaspase-9, procaspase-3, and full-length PARP as a substrate
of caspase-3 were reduced, and cleaved PARP level was increased in
BS-IV-treated HT-29 cells (Fig.
1E). In addition, BS-IV treatment induced dose-dependent
increase in NAG-1 expression (Fig.
1F) and increased caspase-3 activity (Fig. 1G). These results indicated that
BS-IV induces apoptosis via a mitochondrial-dependent pathway and
expression of NAG-1 in HT-29 human colorectal cancer cells.
BS-IV decreases cell adhesion and
expression levels of α2β1 integrin in HT-29
cells
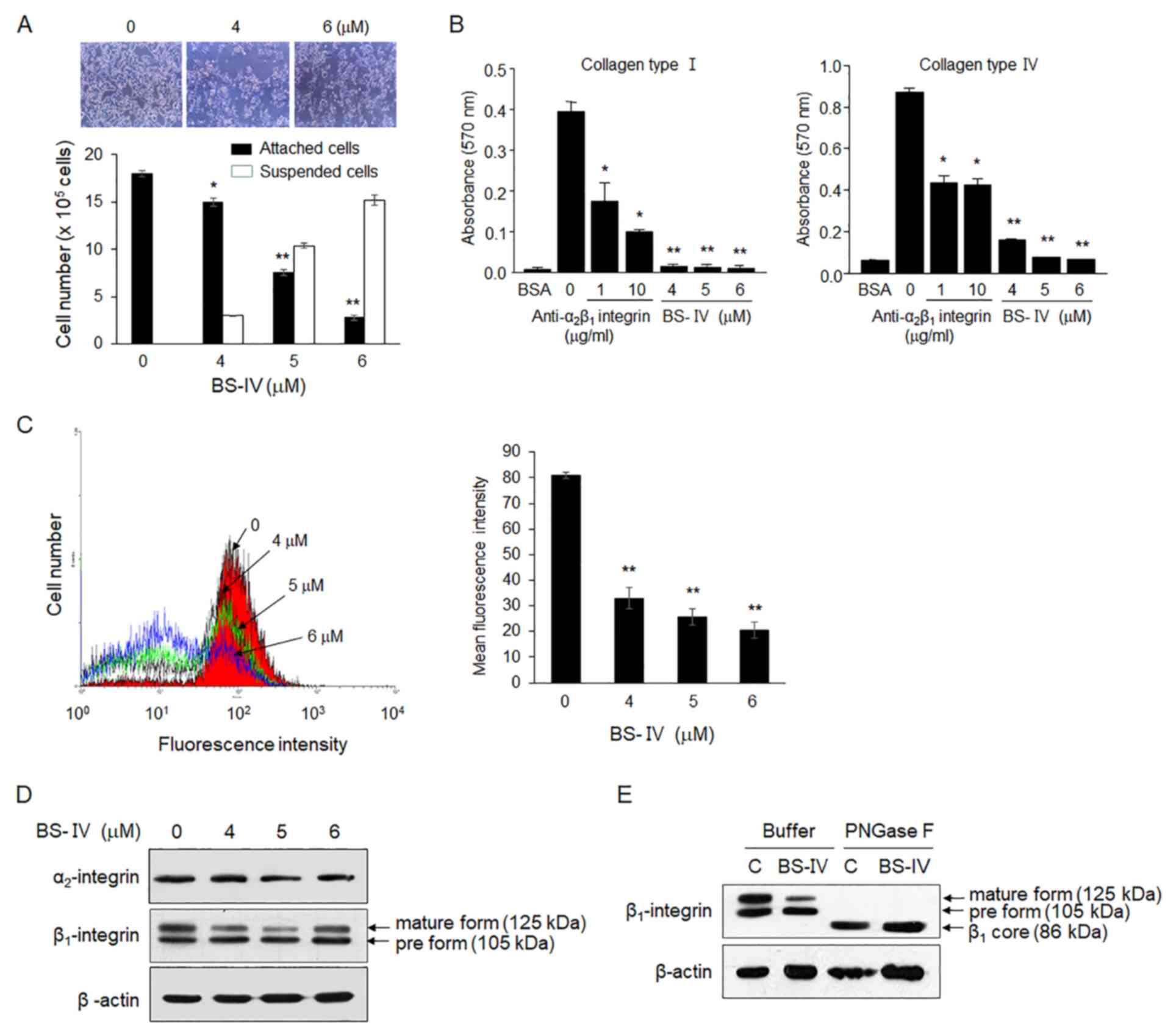
It was also detected that the number of attached
cells decreased and the number of floating cells increased in
BS-IV-treated HT-29 cells (Fig.
2A). It was next investigated whether BS-IV affected the
attachment of HT-29 cells to the extracellular matrix (ECM). The
attachment of HT-29 cells was significantly increased in collagen
type I- or type IV-coated plates compared with BSA-coated plates.
BS-IV treatment reduced the attachment of HT-29 cells to collagen
types I and IV to the level of cell attachment to BSA, whereas the
addition of anti-α2β1 integrin antibody at 10
µg/ml blocked cell attachment to collagen types I and IV by 75 and
51%, respectively (Fig. 2B). The
molecular mechanism by which BS-IV inhibits the attachment of HT-29
cells to collagen types I and IV was further explored. Flow
cytometric analysis demonstrated that treatment with BS-IV
inhibited cell surface expression of α2β1
integrin in HT-29 cells in a dose-dependent manner (Fig. 2C). Western blot analysis indicated
that treatment with BS-IV reduced cellular levels of the
α2 subunit and β1 subunit mature form
(Fig. 2D). The β1
integrin subunit is synthesized as an 86 kDa polypeptide and
sequentially glycosylated to a precursor form of 105 kDa followed
by a mature form of 125 kDa in the endoplasmic reticulum and Golgi
apparatus. The maturation of β1 integrin is important for its
transport to the cell surface and/or binding to ECM ligands. Cell
surface α integrins are transported to the cell surface after
binding to β1 integrin in cells (16). Treatment with BS-IV and then PNGase
F, which completely removes oligosaccharide chains from
glycoproteins, converted both the precursor and mature forms of
β1 integrin to the β1 integrin core of an 86
kDa polypeptide. β1 integrin core levels were not
reduced by BS-IV treatment (Fig.
2E). These results indicated that BS-IV inhibits the adhesion
of HT-29 cells on ECM components and that the reduced cell adhesion
to collagen types I and IV in response to BS-IV treatment may be
attributed to reduced levels of α2β1, the
cellular receptor of collagen types I and IV, which appears to
occur by reducing expression of the α2 subunit and
blocking the maturation of β1 integrin precursors via
glycosylation.
BS-IV decreases the expression and
phosphorylation of FAK and Akt
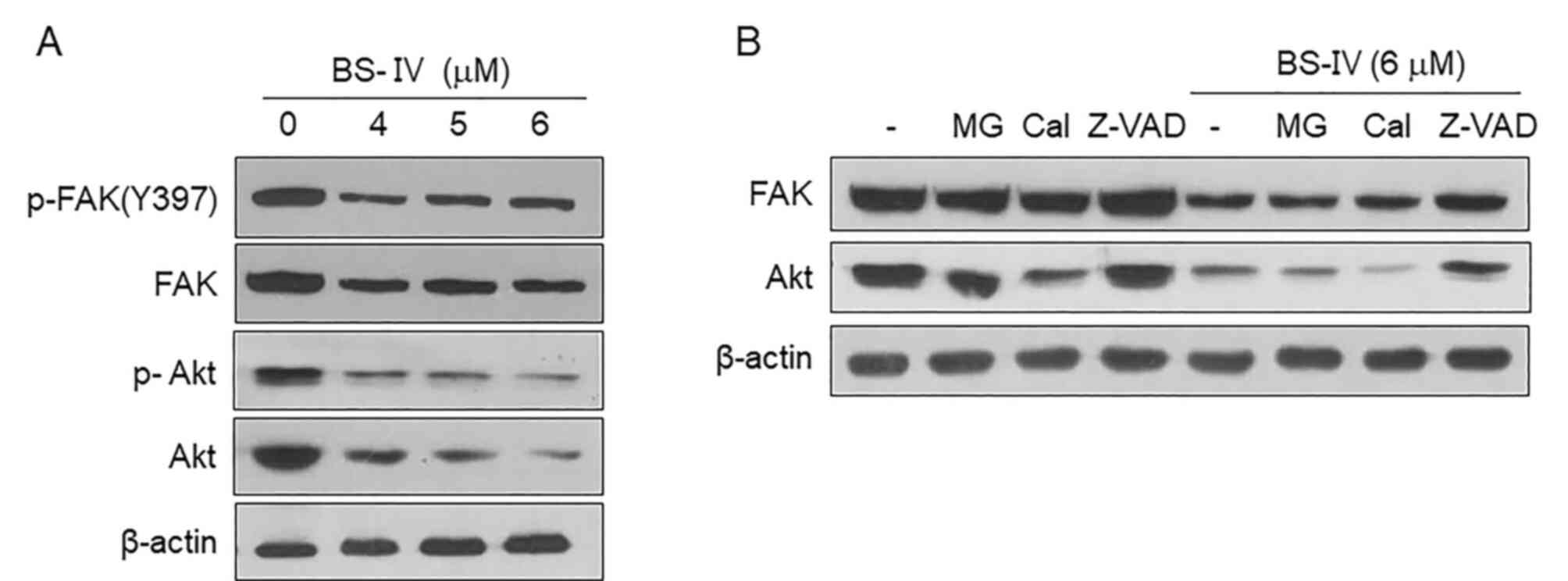
It was further examined whether BS-IV affected the
activation of FAK and Akt, major signaling molecules that play
important roles in numerous fundamental cellular functions,
including cell adhesion (17).
Treatment with BS-IV reduced the expression and phosphorylation of
Akt, as well as the expression and Tyr397
phosphorylation of FAK (Fig. 3A).
The reduced FAK and Akt expression levels were recovered in the
presence of the caspase-3 inhibitor Z-VAD-FMK but not the
proteasome inhibitor MG132 or the calpain inhibitor calpeptin in
HT-29 cells treated with 6 µM BS-IV (Fig. 3B). These results indicated that
caspase-3 activated in response to BS-IV treatment may reduce the
expression and phosphorylation levels of Akt and FAK by degrading
them.
BS-IV administration inhibits in vivo
lung metastasis
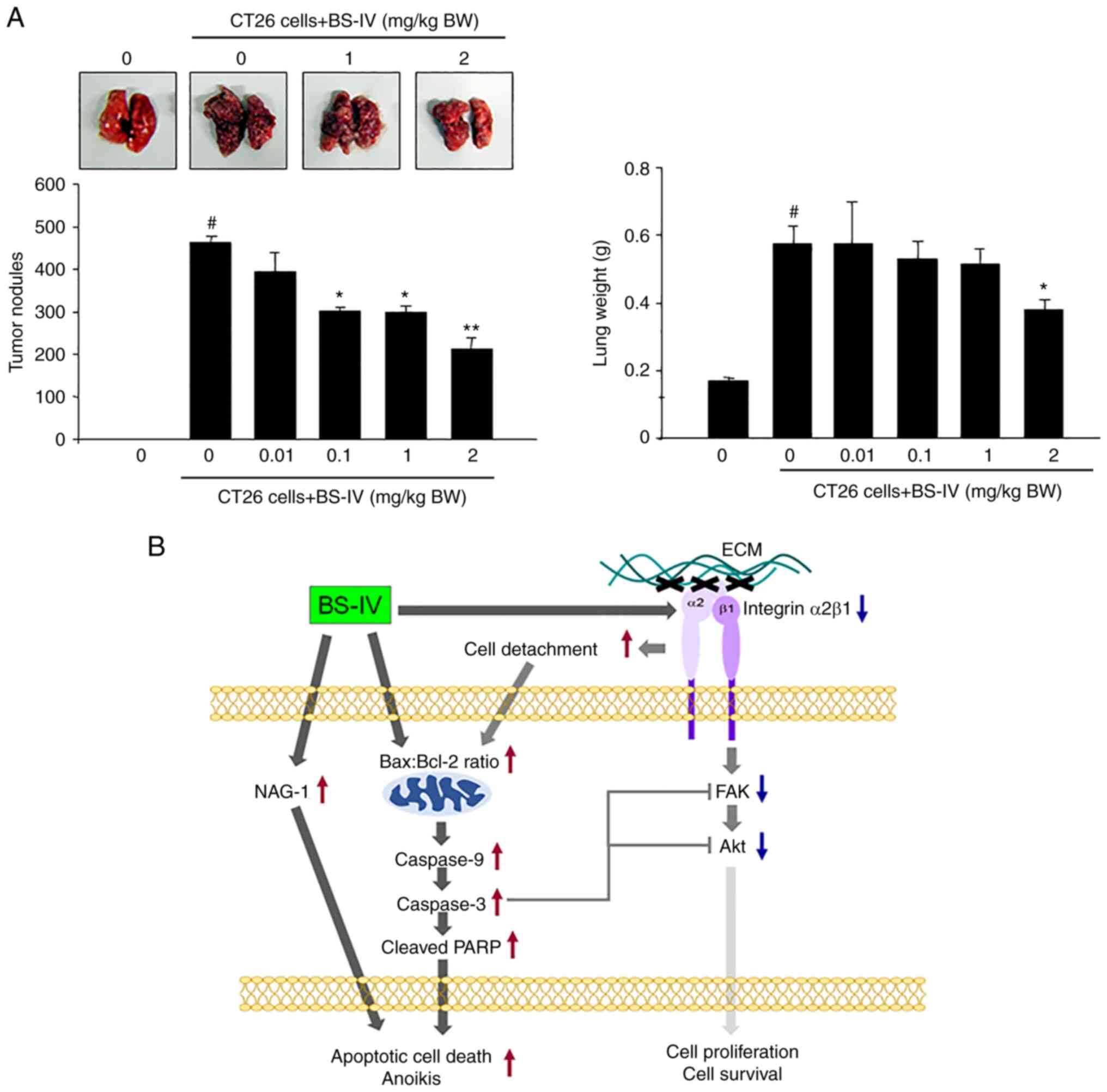
Intravenous injection of CT-26 murine colorectal
cancer cells into mice caused the formation of tumor nodules in the
lung, resulting in a significant increase in lung weight.
Intraperitoneally injected BS-IV significantly reduced the number
of lung tumor nodules and lung weight in a dose-dependent manner
(Fig. 4A). Administration of BS-IV
at 2 mg/kg BW reduced the formation of lung tumor nodules by 54%
and lung weight by 33%. These results indicated that BS-IV
suppresses lung metastasis of murine colon carcinoma CT-26
cells.
Discussion
Apoptosis is a critical process for organism
development and the maintenance of tissue homeostasis and is
tightly regulated by mitochondrial-dependent and death
receptor-mediated apoptotic signaling pathways. Cancer cells lose
the ability to undergo apoptosis and lead to uncontrolled
proliferation (18). The induction
of apoptosis in cancer cells can be an attractive target for cancer
chemoprevention and treatment. In a previous study, it was reported
that the methanol extract of P. kamtschaticum induced
apoptosis via mitochondrial-dependent apoptotic signaling and the
induction of the NAG-1 apoptotic protein in colon cancer cells
(13). In the present study, it was
found that BS-IV is an apoptosis-inducing compound of P.
kamtschaticum by confirming DNA fragmentation, increased
Bax/Bcl-2 ratio, activation of caspase-3 and caspase-9, cleavage of
PARP, and induction of the NAG-1 apoptotic protein in HT-29
colorectal cancer cells. NAG-1, a member of the transforming growth
factor-β superfamily, can be induced either in a p53-dependent or
p53-independent manner, and overexpression of NAG-1 is correlated
with growth inhibition and apoptosis induction in cancer cells
(19). Furthermore, it was revealed
that BS-IV induces anoikis, a special type of apoptotic cell death,
and the molecular mechanism by which BS-IV reduces cell adhesion
and induces anoikis in HT-29 colorectal cancer cells was
investigated.
Anoikis is a crucial cell death program induced in
the absence of cell attachment to the ECM or by cell adhesion to
inappropriate locations. Anoikis resistance endows cancer cells to
survive under suspension conditions or to proliferate at ectopic
sites. This dysregulation in anoikis execution is considered a
hallmark of cancer cells and contributes to distant metastasis
(20). Thereby, anoikis-inducing
phytochemicals may be promising candidates for anticancer agents.
Cell attachment to the ECM is mediated via integrins, cell surface
receptors for ECM ligands. Integrin binding of ECM promotes cancer
cell proliferation, migration, and metastasis and protects cancer
cells from apoptosis and anoikis by eliciting diverse signaling
pathways (21).
α2β1 integrin has been implicated in the
adhesion, proliferation, and metastasis of colon cancer cells
(22–24). The present study indicated that
BS-IV significantly inhibited the attachment of HT-29 cells to
collagen types I and IV, and this inhibition resulted from the
downregulated cell surface expression of α2β1
integrin by reducing protein levels of the α2 integrin
subunit and maturation of the β1 integrin subunit via
glycosylation. These results suggested that disruption of the
α2β1 integrin-ECM interaction in HT-29 cells
is at least a partial inducer of BS-IV-mediated death
signaling.
Cell adhesion to the ECM triggers pro-survival
pathways through the activation of downstream molecules and
suppresses anoikis through the intrinsic and extrinsic apoptotic
pathways of cell death (19). FAK
is one of the most important integrin signaling molecules recruited
into focal adhesions upon cell-ECM contact and promotes cell
survival, proliferation, and motility. Upon integrin ligation and
clustering, FAK auto phosphorylates at Tyr 397, enhancing
intermolecular FAK kinase activity, and association with FAK and
phosphoinositide 3-kinase leads to the phosphorylation of Akt
(25,26). Akt activation promotes cell survival
by releasing Bcl-2 and directly inhibiting the caspase cascade
(27). In the present study, BS-IV
reduced protein expression levels of FAK and Akt, resulting in
their decreased phosphorylation levels. FAK and Akt protein levels
were rescued by treatment with a caspase-3 inhibitor. These
findings suggested that the degradation of FAK and Akt by
BS-IV-activated caspase-3 can also contribute to inhibiting
integrin-mediated signaling in HT-29 colorectal cancer cells.
Metastasis is a sequential process in which tumor
cells detach from their site of primary growth, invade through the
surrounding host tissue into the circulation, disseminate to
distant organs, and extravasate and proliferate to form metastatic
foci. Intraperitoneal administration of BS-IV, which induced
anoikis in HT-29 colorectal cancer cells, inhibited the lung
metastasis of CT-26 murine colorectal cancer cells.
In conclusion, BS-IV induces apoptotic cell death,
including anoikis, in colon cancer cells by activating the
mitochondrial-dependent pathway, inducing NAG-1 expression, and
inhibiting integrin-mediated antiapoptotic signaling (Fig. 4B). Therefore, BS-IV may serve as a
beneficial chemopreventive and anti-metastatic agent with potent
proapoptotic and anoikis-inducing potential. BS-IV is a triterpene
saponin and has been reported to show a poor bioavailability due to
the poor permeability and significant hepatic first-pass effect
(28). For clinical application of
BS-IV, further studies shall be conducted by the authors to improve
its limitations, including water solubility, absorption, and
bioavailability.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Research
Foundation of Korea (NRF) grant funded by the Korea government
(MSIT) (grant no. NRF-2021R1A2C2011252).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JEK, SKL and WYC conceived the study, designed and
performed experiments, analyzed the data, interpreted the results
and wrote the manuscript. SKL and WYC confirm the authenticity of
all the raw data. JP, MJJ, SEA and HJY contributed to the
conception of the study, analyzed the data and reviewed the
results. All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
Animal studies were approved (approval no.
2020-0193) by the animal ethics committee of Yonsei University
College of Dentistry (Seoul, Korea) and conducted in accordance
with the approved guidelines of the regional authorities according
to Yonsei University animal care regulations.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Douglass EC: Development of ZD1839 in
colorectal cancer. Semin Oncol. 30 (3 Suppl 6):S17–S22. 2003.
View Article : Google Scholar
|
|
2
|
Vermeer NCA, Snijders HS, Holman FA,
Liefers GJ, Bastiaannet E, van de Velde CJH and Peeters KCMJ:
Colorectal cancer screening: Systematic review of screen-related
morbidity and mortality. Cancer Treat Rev. 54:87–98. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gustin DM and Brenner DE: Chemoprevention
of colon cancer: Current status and future prospects. Cancer
Metastasis Rev. 21:323–348. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Madka V and Rao CV: Anti-inflammatory
phytochemicals for chemoprevention of colon cancer. Curr Cancer
Drug Targets. 13:542–557. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhao Y, Hu X, Zuo X and Wang M:
Chemopreventive effects of some popular phytochemicals on human
colon cancer: A review. Food Funct. 9:4548–4568. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ranjan A, Ramachandran S, Gupta N, Kaushik
I, Wright S, Srivastava S, Das H, Srivastava S, Prasad S and
Srivastava SK: Role of phytochemicals in cancer prevention. Int J
Mol Sci. 20:49812019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee IK, Choi SU and Lee KR: Triterpene
saponins from Pleurospermum kamtschaticum and their
biological activity. Chem Pharm Bull (Tokyo). 60:1011–1018. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guinea MC, Parellada J, Lacaille-Dubois MA
and Wagner H: Biologically active triterpene saponins from
Bupleurum fruticosum. Planta Med. 60:163–167. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jung HJ, Nam JH, Park HJ, Lee KT, Park KK,
Kim WB and Choi J: The MeOH extract of Pleurospermum
kamtschaticum and its active component buddlejasaponin (IV)
inhibits intrinsic and extrinsic hyperlipidemia and
hypercholesterolemia in the rat. J Ethnopharmcol. 112:255–261.
2007. View Article : Google Scholar
|
|
10
|
Jung HJ, Kim SG, Nam JH, Park KK, Chung
WY, Kim WB, Lee KT, Won JH, Choi JW and Park HJ: Isolation of
saponins with the inhibitory effect on nitric oxide, prostaglandin
E2 and tumor necrosis factor-alpha production from Pleurospermum
kamtschaticum. Biol Pharm Bull. 28:1668–1671. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Won JH, Im HT, Kim YH, Yun KJ, Park HJ,
Choi JW and Lee KT: Anti-inflammatory effect of buddlejasaponin IV
through the inhibition of iNOS and COX-2 expression in RAW 264.7
macrophages via the NF-kappaB inactivation. Br J Pharmacol.
148:216–225. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tundis R, Bonesi M, Deguin B, Loizzo MR
and Menichini F, Conforti F, Tillequin F and Menichini F: Cytotoxic
activity and inhibitory effect on nitric oxide production of
triterpene saponins from the roots of physospermum verticillatum
(Waldst & Kit) (Apiaceae). Bioorg Med Chem. 17:4542–4547. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim JE, Chung WY, Chun KS, Lee CK, Park
HJ, Kim WB and Park KK: Pleurospermum kamtschaticum extract
induces apoptosis via mitochondrial pathway and NAG-1 expression in
colon cancer cells. Biosci Biotechnol Biochem. 74:788–792. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hwang YS, Chung WY, Kim J, Park HJ, Kim EC
and Park KK: Buddlejasaponin IV induces cell cycle arrest at G2/M
phase and apoptosis in immortalized human oral keratinocytes.
Phytother Res. 25:1503–1510. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim KR, Park KK, Chung WY and Hwang YS:
The inhibitory effect of buddlejasaponin IV on the growth of YD-10B
human oral squamous cell carcinoma cells. J Cancer Prev.
18:330–336. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hu W, Xu R, Zhang G, Jin J, Szulc ZM,
Bielawski J, Hannun YA, Obeid LM and Mao C: Golgi fragmentation is
associated with ceramide-induced cellular effects. Mol Biol Cell.
16:1555–1567. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Desiniotis A and Kyprianou N: Significance
of talin in cancer progression and metastasis. Int Rev Cell Mol
Biol. 289:117–147. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mohammad RM, Muqbil I, Lowe L, Yedjou C,
Hsu HY, Lin LT, Siegelin MD, Fimognari C, Kumar NB, Dou QP, et al:
Broad targeting of resistance to apoptosis in cancer. Semin Cancer
Biol. 35 (Suppl):S78–S103. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang X, Baek SJ and Eling TE: The diverse
roles of nonsteroidal anti-inflammatory drug activated gene
(NAG-1/GDF15) in cancer. Biochem Pharmacol. 85:597–606. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Paoli P, Giannoni E and Chiarugi P:
Anoikis molecular pathways and its role in cancer progression.
Biochim Biophys Acta. 1833:3481–3498. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hynes RO: Integrins: Versatility,
modulation, and signaling in cell adhesion. Cell. 69:11–25. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Buda A, Qualtrough D, Jepson M, Martines
D, Paraskeva C and Pignatelli M: Butyrate downregulates alpha2beta1
integrin: A possible role in the induction of apoptosis in
colorectal cancer cell lines. Gut. 52:729–734. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Massoumi R, Nielsen CK, Azemovic D and
Sjölander A: Leukotriene D4-induced adhesion of Caco-2 cells is
mediated by prostaglandin E2 and upregulation of
alpha2beta1-integrin. Exp Cell Res. 289:342–351. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bartolomé RA, Barderas R, Torres S,
Fernandez-Aceñero MJ, Mendes M, García-Foncillas J, Lopez-Lucendo M
and Casal JI: Cadherin-17 interacts with α2β1 integrin to regulate
cell proliferation and adhesion in colorectal cancer cells causing
liver metastasis. Oncogene. 33:1658–1669. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun H, Calle E, Chen X, Mathur A, Zhu Y,
Mendez J, Zhao L, Niklason L, Peng X, Peng H and Herzog EL:
Fibroblast engraftment in the decellularized mouse lung occurs via
a β1-integrin-dependent, FAK-dependent pathway that is mediated by
ERK and opposed by AKT. Am J Physiol Lung Cell Mol Physiol.
306:L463–L475. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang Z, Wang Z, Li G, Wu H, Sun K, Chen J,
Feng Y, Chen C, Cai S, Xu J and He Y: CXCL1 from tumor-associated
lymphatic endothelial cells drives gastric cancer cell into
lymphatic system via activating integrin β1/FAK/AKT signaling.
Cancer Lett. 385:28–38. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cardone MH, Roy N, Stennicke HR, Franke
TF, Stanbridge E, Frisch S and Reed JC: Regulation of cell death
protease caspase-9 by phosphorylation. Science. 282:1318–1321.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li Y, Xu H, Chen L and Tan L: A simple and
sensitive UHPLC-MS/MS method for quantification of buddlejasaponin
IV in rat plasma and its application to a pharmacokinetic study. J
Pharm Biomed Anal. 120:374–382. 2016. View Article : Google Scholar : PubMed/NCBI
|