Introduction
In 2020, colorectal cancer (CRC) accounted for 1.1
million new cancer cases and 570,000 cancer-associated deaths and
the second leading cause of cancer-associated death worldwide
(1). Despite the development of
diagnostic methods, surgical techniques and targeted treatment, the
5-year survival rate of patients with advanced stage CRC is 15%
(2). Therefore, the development of
effective chemopreventive agents against CRC, including for
metastatic cases, is important for CRC treatment. Medicinal plants
may be considered useful resources for identifying therapeutic
compounds as they may be safer and exert fewer side effects than
synthetic drugs (3). In addition,
phytochemicals have been used for antitumor therapy owing to their
protective and reparative mechanisms in the body (4–7).
Maple syrup is consumed as a sweetener due to its
unique taste and flavor; it also has a high nutritional value.
Maple syrup is prepared by boiling sap taken from sugar maple trees
(Acer saccharum) and is primarily produced in North America,
particularly in Canada and the United States (8–10).
Maple syrup is a better alternative to refined sugar in the human
diet owing to its low glycemic and insulinemic response properties
(11,12). In addition, it was previously
reported that maple syrup has an anti-tumor effect against
gastrointestinal cancer cells, including CRC (13–15).
Therefore, maple syrup may be a useful resource to investigate
phytochemicals for CRC treatment.
Several studies have examined the phytochemical
components in maple syrup regarding their antitumor effects
(16–18). Ethyl acetate extracts of maple syrup
exhibit antioxidant activity and antiproliferative effects against
lung and colon cancer cell lines (16). In addition, butanol and methanol
extracts of maple syrup exhibit an antiproliferative effect against
CRC cell lines (17). Several
studies on the effective components in maple syrup extracts have
also been performed: Gallic acid and syringaldehyde, present in the
butanol extract of maple syrup, have antitumor effects (17), and ginnalins A, B and C isolated
from maple syrup inhibit breast and colon cancer cell proliferation
(18). These ingredients are
isolated by organic solvent extraction and are all phenolic
compounds. Maple syrup contains not only phenolic compounds, but
also physiologically active substances, such as organic acids,
volatile sulfur compounds, pyrazines and proteins (9,19–23).
The antitumor effects of these non-phenolic components against CRC
cells are unclear. Thus, the aim of the present study was to
determine the antitumor effects of the non-phenolic components of
maple syrup using an extraction approach that does not rely on
organic solvents. The results suggested that the protein components
in maple syrup may have anti-tumor effects against CRC cells.
Materials and methods
Reagents
Ammonium sulfate was purchased from Fujifilm Wako
Pure Chemical Corporation. Maple Farms Japan, Inc (Osaka, Japan).
generously gifted grade A maple syrup made from sugar maple (A.
saccharum) by Bascom Maple Farms, Inc. (Acworth, New Hampshire,
USA).
CRC cell culture
DLD-1 colon adenocarcinoma cells were purchased from
the American Type Culture Collection and cultured in RPMI-1640
medium (Fujifilm Wako Pure Chemical Corporation) supplemented with
10% FBS (Gibco; Thermo Fisher Scientific, Inc.) in a humidified
incubator supplied with 5% CO2 maintained at 37°C.
T4056-C human primary normal colon epithelial cells (passage 2)
were cultured in Prigrow X Medium for T4056-C (both from Applied
Biological Materials Inc.) according to the manufacturer's
protocol.
Ultrafiltration
Maple syrup (100 ml) was diluted in 200 ml ultrapure
water and ultrafiltered through an Amicon Ultra-15, Ultracel-10K
centrifugal filter unit (MilliporeSigma) according to the
manufacturer's instructions. Filtrates and concentrates were
fractionated and collected as a low molecular weight (MW) fraction
of maple syrup (MW<10,000) and a high MW fraction
(MW≥10,000).
Preparation of protein fraction of
maple syrup (MSpf)
To obtain MSpf, ammonium sulfate was added to the
high MW fraction to a final concentration of 80% saturation. The
sample was centrifuged at 3,000 × g at 4°C for 15 min following
incubation at 4°C overnight and the supernatant was removed. These
steps were repeated once. The precipitate was dissolved in 5 ml PBS
(MilliporeSigma). Any ammonium sulfate that remained was removed by
ultrafiltration using an Amicon Ultra-15, Ultracel-10K. The total
protein concentration in solution was measured using a Bio-Rad
Protein Assay kit (Bio-Rad Laboratories, Inc.) according to the
manufacturer's instructions.
Cell viability assays
DLD-1 cells were cultured in a 6-well plate at a
density of 1×105 cells/well and grown in culture medium
as aforementioned. Following 24 h incubation at 37°C, the medium
was replaced and cells were grown in maple syrup, high or low MW
fraction, MSpf (2.5, 5.0 or 10 µg/ml) or PBS (untreated control) in
culture medium. DLD-1 cells were treated with maple syrup as
described in our previous study (13), and DLD-1 cells were also treated by
high or low MW fraction to be the same as treated with an equal
volume of maple syrup. The number of cells which were treated with
maple syrup or high or low MW fraction was counted after 72 h using
a Countess Automated Cell Counter (Thermo Fisher Scientific, Inc.).
The number of cells which were treated with MSpf or PBS was counted
after 24, 48, 72 and 96 h using a Countess Automated Cell Counter
(Thermo Fisher Scientific, Inc.).
DLD-1 cells were also plated at a density of
5×103 cells/well in a 96-well plate and grown in culture
medium as aforementioned. The following day, the medium was
replaced and cells were grown in MSpf (10 µg/ml) without ammonium
sulfate or in PBS control in culture medium, as aforementioned.
After 96 h, the cells were incubated with WST-8 cell counting
reagent (Fujifilm Wako Pure Chemical Corporation) at 2 h and the
optical density of the culture solution in the plate was measured
at 450 nm using an ELISA plate reader.
To examine the effect of the protein component of
maple syrup on the viability of normal colon cells, T4056-C cells
(passage no. 3) were plated at a density of 5×103
cells/well in a 96-well plate and grown in culture medium as
aforementioned. The 96-well plate was coated with Applied Cell
Extracellular Matrix (Applied Biological Materials Inc.) according
to the manufacturer's instructions. Following 24 h incubation at
37°C, the medium was replaced and cells were grown in MSpf (10
µg/ml)- or PBS (control)-containing culture medium, as
aforementioned. After 96 h, WST-8 assay was performed, as
aforementioned.
To examine the effect of the protein component of
maple syrup on cell autophagy, cells were plated at a density of
5×103 cells/well in a 96-well plate as aforementioned.
The following day, the medium was replaced and cells were grown in
either MSpf (10 µg/ml) or PBS (control) in the presence or absence
of bafilomycin A1 (10 nM; MilliporeSigma), as previously described
(24), in culture medium. After 72
h, the WST-8 assay was performed, as aforementioned.
SDS-PAGE
To determine the molecular weight distribution of
MSpf, SDS-PAGE was performed on a gradient SuperSep Ace (5–20%
gradient gel; Fujifilm Wako Pure Chemical Corporation) loaded 10 µg
MSpf/lane. Protein bands were stained with Silver Stain II kit
(Fujifilm Wako Pure Chemical Corporation) according to the
manufacturer's instructions.
Cell migration and invasion assay
In vitro migration assays were performed
using a Boyden chamber (BD Biosciences). DLD-1 cells were plated on
the inner surface of the inserts for 24-well plate with 8.0 µm pore
size at a density of 1×105 cells/insert, followed by
incubation at 37°C for 48 h in a humidified incubator. Culture
medium containing 10% FBS with MSpf (10 µg/ml) or PBS (control) was
added to the lower chamber as a chemoattractant. DLD-1 cells on the
outer surface of the inserts were counted after 48 h, as described
previously (25), and the cells
that moved to the outer surface of the inserts were fixed, stained
with a Diff-Quik staining kit (Sysmex, Corp.) according to the
manufacturer's instructions. All assays were performed in
triplicate and five fields of view (magnification, ×200) were
counted on each membrane in a blinded manner using a light
microscope, EVOS FLoid Cell Imaging Station (Thermo Fisher
Scientific, Inc.). The cell invasion assay was performed in the
same way, except a Matrigel Invasion Chamber 24-Well Plate 8.0 µm
was used as the insert (cat. no. 354480; Corning, Inc.).
Proteomic analysis
The MSpf (10 µg/lane) was separated by SDS-PAGE
using 5–20% gradient gel (Fujifilm Wako Pure Chemical Corporation).
Protein bands were stained with Silver Stain MS kit (Fujifilm Wako
Pure Chemical Corporation) according to the manufacturer's
instructions. The protein bands were excised manually and then
digested using In-gel Tryptic Digestion kit (Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
Then, the digests were purified using PepClean C-18 Spin Columns
(Thermo Fisher Scientific, Inc.) following the manufacturer's
instructions.
Peptide samples (~2 µg) were injected into a peptide
L-trap column (Chemicals Evaluation and Research Institute) with an
HTC PAL autosampler (CTC Analytics). They were further separated
through a Paradigm MS4 (AMR Inc.) with a reverse-phase C18-column
(L-column, 3-µm-diameter gel particles, 120 Å pore size, 0.2×50 mm;
Chemicals Evaluation and Research Institute). The column flow rate
was 1 µl/min, and the mobile phase consisted of 0.1% formic acid in
water (solution A) and acetonitrile (solution B), with a
concentration gradient of 5% solution B to 40% solution B over 45
min. Gradient-eluted peptides were introduced into the mass
spectrometer through the nanoelectrospray ionization (NSI)
interface that had a separation column outlet directly connected
with a NSI needle. We analyzed the peptides with an LTQ ion-trap
mass spectrometer (Thermo Fisher Scientific, Inc.), using no sheath
or auxiliary gas. The MS scan sequence was full-scan MS in the
normal/centroid mode and sequential MS/MS in the normal/centroid
mode. The positive ion mass spectra were acquired in a
data-dependent manner, with MS/MS fragmentation performed on the
two most intense peaks of every full MS scan with an isolation
width of m/z 1.0 and a collisional activation amplitude of 35% in
the m/z range of 300 to 2,000. All MS/MS spectral data were
searched against the SwissProt Acer L (Maple tree) database
using Mascot version 2.4.01 (Matrix Science). The search criteria
were enzyme as trypsin, with the following allowances: ≤2 missed
cleavage peptides; mass tolerance, ±2.0 Da; MS/MS tolerance, ±0.8
Da; cysteine carbamidomethylation for fixed modification; and
methionine oxidation modifications for variable modification.
ELISA
Advanced glycation end products (AGEs) in MSpf were
quantified by ELISA using the OxiSelect Advanced Glycation End
Product Competitive ELISA kit (cat. no. STA-817-T; Cell Biolabs,
Inc.) according to the manufacturer's instructions.
Cell morphology analysis
DLD-1 cells were plated at a density of
1×105 cells/dish in a 30-mm dish and grown in culture
medium as aforementioned. Following 24 h incubation at 37°C, the
medium was replaced and cells were cultured with MSpf (10 µg/ml) or
PBS (control) in culture medium as aforementioned. The cells were
imaged using an EVOS FLoid Cell Imaging Station (Thermo Fisher
Scientific, Inc.) after 72 h incubation.
Autophagy detection
To detect autophagic cells, DLD-1 cells were plated
at a density of 2×105 cells/slide in a Nunc Lab-Tek
Chamber Slide System (Thermo Fisher Scientific, Inc.) in the
aforementioned culture medium. The following day, the medium was
replaced and cells were grown in 0.15 µM DAPGreen (Fujifilm Wako
Pure Chemical Corporation) in culture medium for 30 min, according
to the manufacturer's protocol. Then, cells were grown in MSpf (10
µg/ml) or PBS (control) in the presence or absence of bafilomycin
A1 (10 nM) in culture medium after washing with culture medium.
After 72 h incubation at 37°C, the culture medium from all slides
was removed and the cells were washed three times with 1X PBS.
Next, cells were fixed with 4% paraformaldehyde at room temperature
for 10 min and washed again with 1X PBS. Images were captured using
an EVOS FLoid Cell Imaging Station.
Preparation of protein samples for
western blotting
DLD-1 cells were plated at a density of
5×105 cells in a 100-mm dish and grown in culture medium
as aforementioned. The following day, the medium was replaced and
cells were grown in culture medium containing MSpf (10 µg/ml) or
PBS (control) with or without bafilomycin A1 (10 nM) as
aforementioned. After 72 h, cells were solubilized in urea lysis
buffer (7 M urea, 2 M thiourea, 5% CHAPS and 1% Triton X-100). The
protein concentration was measured using a Bradford assay.
Western blot analysis
Total protein (10 µg/lane) was mixed with loading
buffer and boiled at 95°C for 10 min. The proteins were separated
by SDS-PAGE [8% gel E-cadherin and N-cadherin; 10% gel for MMP-2,
MMP-9, RAGE, phosphorylated (p)STAT3, pp44/42 MAPK,
p-stress-activated protein kinase 1c (SAPK)/JNK, pp38 MAPK and
pAKT; 15% gel for cleaved caspase-3, caspase-3 and Snail; 5–20%
gradient gel for LC3A] and transferred to PVDF membranes
(MilliporeSigma) for 30 min at 15 V. Following blocking with TBS
+0.1% Tween-20 buffer containing 5% skimmed milk for 2 h at room
temperature, the membranes were incubated with the following
antibodies at 4°C overnight: Anti-Caspase-3 [1:1,000; cat. no.
14220; Cell Signaling Technology, Inc. (CST)], anti-cleaved
Caspase-3 (1:1,000; cat. no. 9661; CST), anti-LC3A (1:1,000; cat.
no. 4599; CST), anti-E-cadherin (1:1,000; cat. no. 3195; CST),
anti-N-cadherin (1:1,000; cat. no. 13116; CST), anti-Snail
(1:1,000; cat. no. 3879; CST), anti-MMP-2 (1:1,000; cat. no. 87809;
CST), anti-MMP-9 (1:1,000; cat. no. 13667; CST), anti-RAGE (1:500;
cat. no. ab216329; Abcam, Inc.), anti-pSTAT3 (1:1,000; cat. no.
9134; CST), anti-pp44/42 MAPK (Erk1/2; 1:1,000; cat. no. 4370;
CST), anti-pSAPK/JNK (1:1,000; cat. no. 4668; CST), anti-pp38 MAPK
(1:1,000; cat. no. 4511; CST) or anti-pAKT (1:1,000; cat. no. 4051;
CST). The membranes were washed with 0.1% TBS-T) and incubated with
HRP-conjugated anti-rabbit or anti-mouse immunoglobulin G antibody
(both 1:4,000; cat. no. A102PU and A106PU; both American Qualex) at
room temperature for 1 h. The blots were washed with TBS-T and
signals were visualized using SuperSignal West Dura Extended
Duration substrate (Thermo Fisher Scientific, Inc.), and analyzed
using myECL Imager system (version 2.0; Thermo Fisher Scientific,
Inc.). The membranes were stripped using Restore Western Blot
Stripping buffer (Thermo Fisher Scientific, Inc.) and re-probed
with anti-β-actin (1:5,000; cat. no. sc-47778; Santa Cruz
Biotechnology, Inc.), anti-STAT3 (1:1,000; cat. no. 9139; CST),
anti-AKT (1:1,000; cat. no. 2920; CST), anti-Erk1/2 (1:1,000; cat.
no. 9102; CST), anti-SAPK/JNK (1:1,000; cat. no. 9252; CST), or
anti-p38 MAPK (1:1,000; cat. no. 8690; CST) antibodies at 4°C
overnight, which served as the protein loading or total protein
controls. All western blot analyses were performed as three
independent experiments.
Statistical analysis
Data are presented as the mean ± SEM of at least
three independent experiments. Statistical analyses were performed
using GraphPad Prism version 8.1.2 (GraphPad Software, Inc.;
Dotmatics). Differences between groups were evaluated using
unpaired Student's t test or one-way ANOVA followed by
Tukey-Kramer's post hoc test. P<0.05 was considered to indicate
a statistically significant difference.
Results
Active ingredients in maple syrup
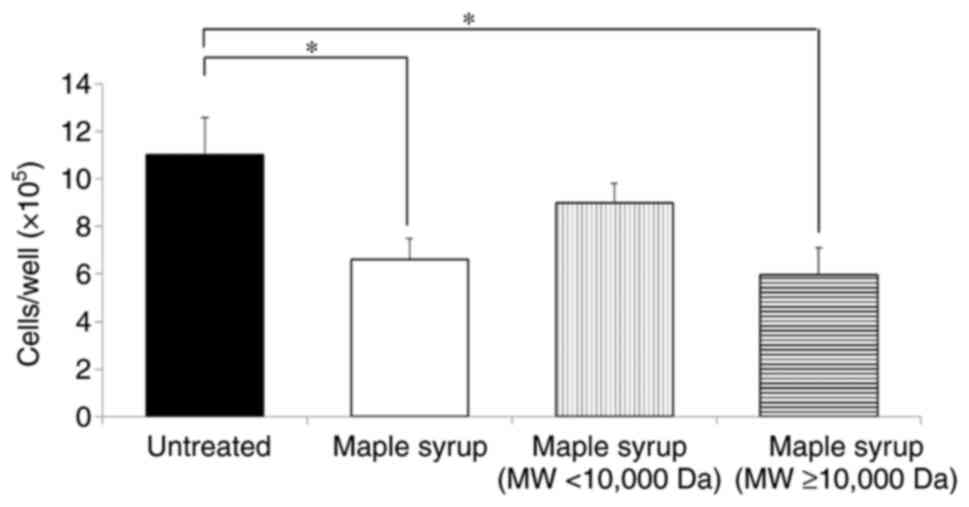
To investigate the active ingredients in maple syrup
that possessed putative antitumor effects against CRC cells, the
maple syrup was separated into high MW and low MW fractions with a
10 kDa cut-off, and the fraction that contained the active
ingredients was determined. Cell viability assays demonstrated that
treatment with the high MW fraction significantly inhibited cell
viability in DLD-1 cells, similar to treatment with the maple syrup
and our previous reports (Fig. 1)
(13,15). On the other hand, there was no
significant difference between high and low MW fraction treatment.
Proteins were considered to be the majority constituents in the
high MW component of the maple syrup. Therefore, the protein
components of maple syrup were suggested to be the active
ingredients in maple syrup.
Preparation and characterization of
the protein components in maple syrup
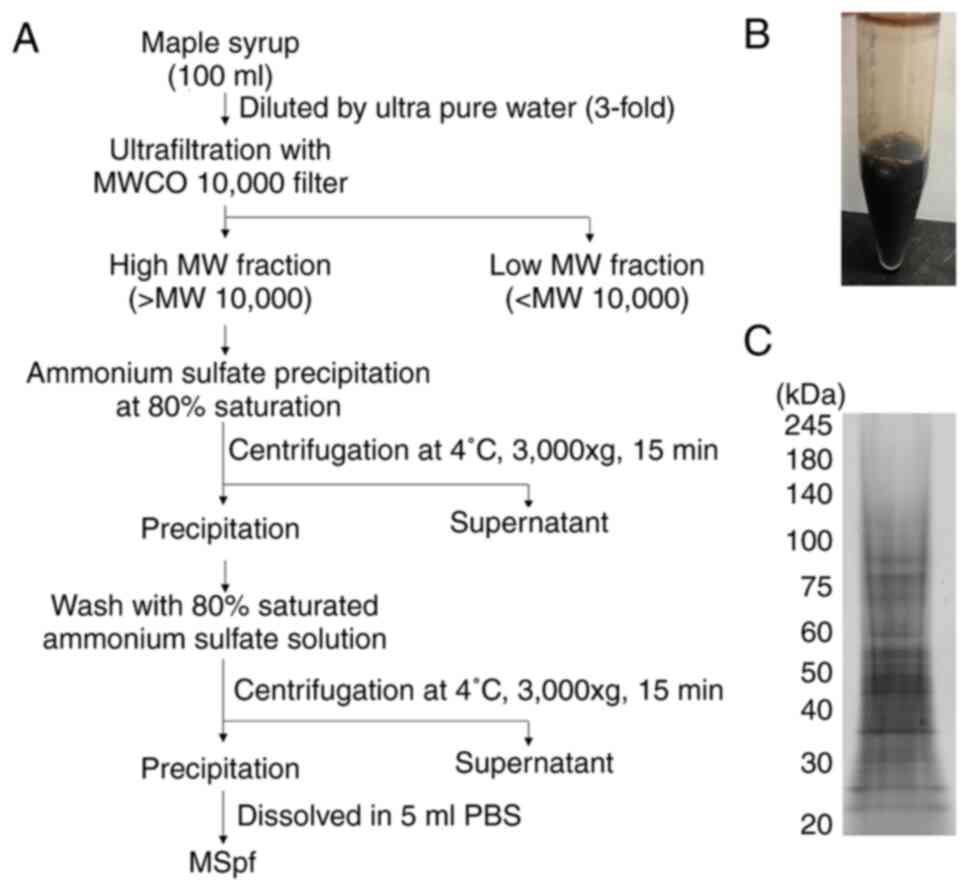
An overview of the preparation of MSpf is shown in
Fig. 2A. MSpf was dissolved in PBS
and used in subsequent experiments (Fig. 2B). Several bands between 20 and 100
kDa were observed following SDS-PAGE of MSpf (Fig. 2C). Based on the protein
concentration of MSpf (0.50±0.05 mg/ml), obtained 5 ml MSpf
solution contain 2.5 mg protein. Therefore, 100 ml maple syrup
contained ~2.5 mg protein.
Effect of MSpf on cell viability,
proliferation, migration and invasion
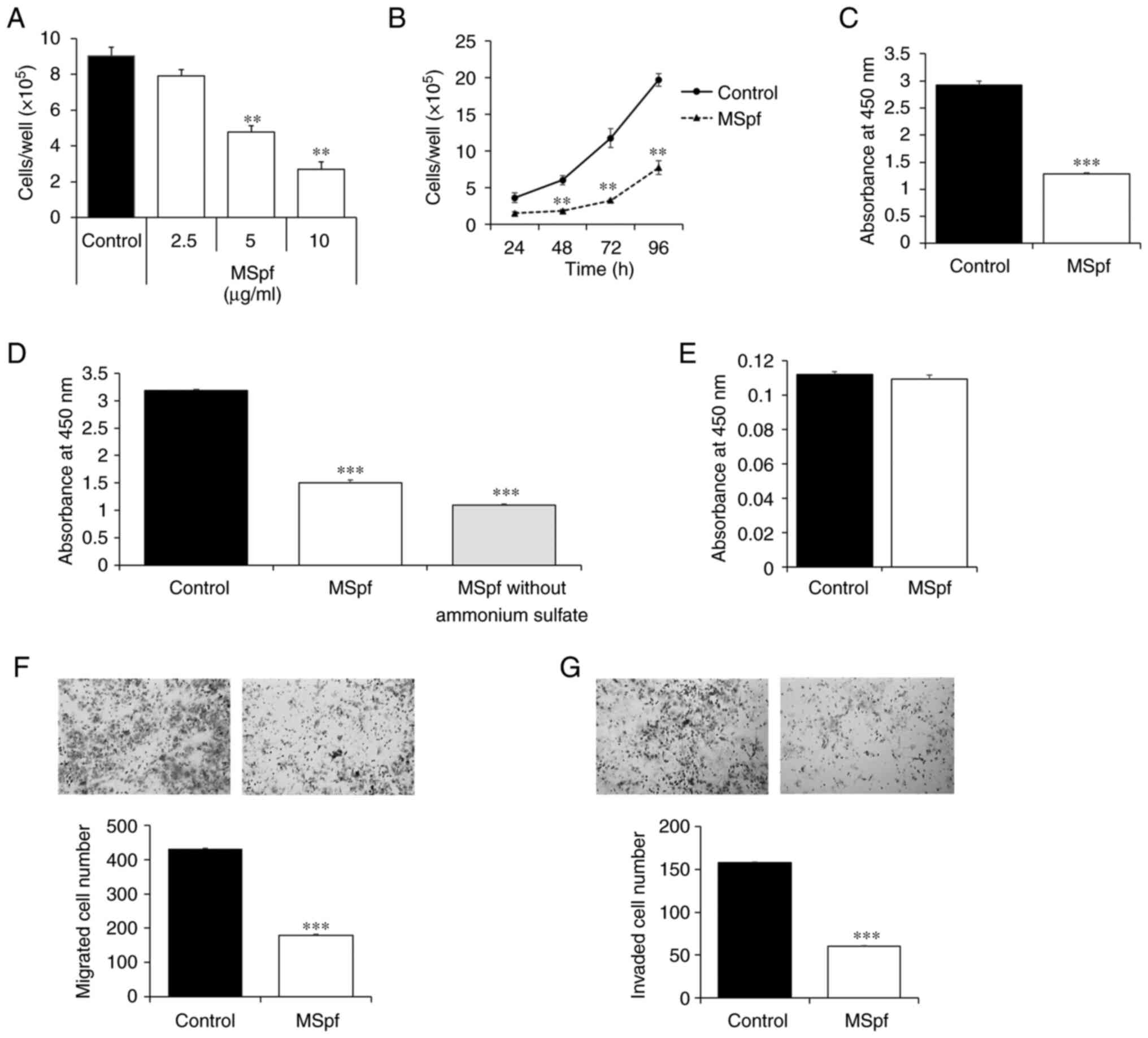
Compared with untreated control cells, 5 and 10
µg/ml MSpf significantly inhibited the viability of DLD-1 cells in
a dose-dependent manner (Fig. 3A);
10 µg/ml MSpf significantly inhibited proliferation of DLD-1 cells
compared with control cells at 48, 72 and 96 h (Fig. 3B). A WST-8 assay was used to confirm
the inhibitory effect of MSpf on viability of DLD-1 cells; 10 µg/ml
MSpf significantly inhibited the viability of DLD-1 cells compared
with the control cells at 96 h (Fig.
3C). MSpf was dissolved in PBS following ammonium sulfate
precipitation. To investigate whether the ammonium sulfate
remaining in the MSpf solution exhibited a cytotoxic effect,
solution was prepared to remove any residual ammonium sulfate by
ultrafiltration and a WST-8 assay was performed using this
solution. The results showed that 10 µg/ml MSpf without ammonium
sulfate significantly reduced viability of DLD-1 cells compared
with control (Fig. 3D). A WST-8
assay was also performed to determine whether MSpf treatment
affected the viability of T4056-C normal colon cells; 10 µg/ml MSpf
treatment did not affect proliferation of human primary colon cells
(Fig. 3E).
Furthermore, the migration and invasion of DLD-1
cells treated with MSpf were assessed, and the data show that the
number of migrating and invading cells was significantly decreased
by MSpf treatment compared with the untreated control group
(Fig. 3F and G, respectively).
Effects of MSpf on the expression of
proteins associated with cell proliferation, migration and
invasion
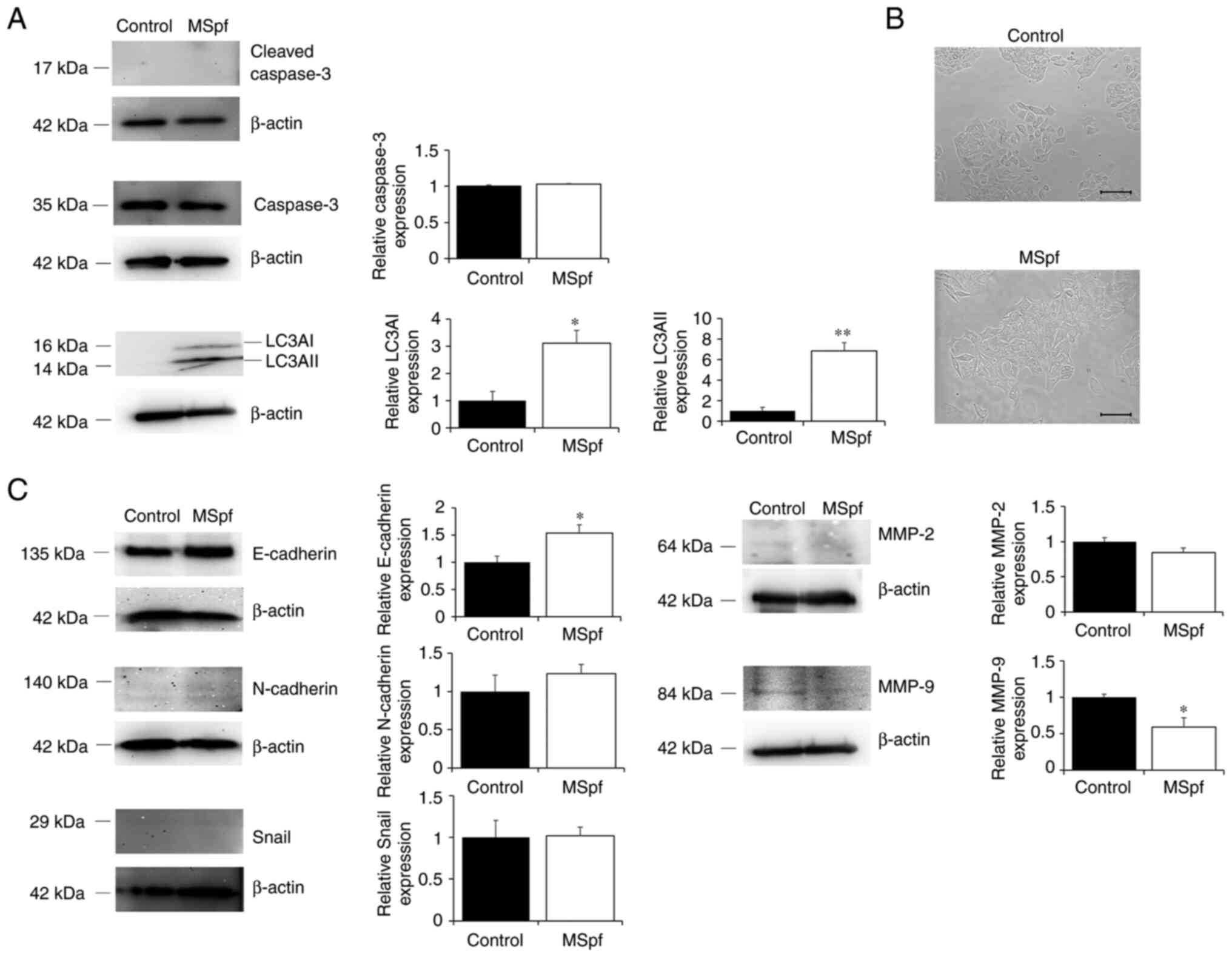
Since proliferation of DLD-1 cells was inhibited by
MSpf treatment, it was hypothesized that this suppression may be
involved in the induction of cell death. To clarify whether MSpf
treatment affected induction of apoptosis or autophagy of DLD-1
cells, the expression of cleaved caspase-3 (a marker of apoptosis)
(26) and LC3A (a marker of
autophagy; 27,28) in MSpf-treated DLD-1 cells was measured
(Fig. 4A). LC3AI and LC3AII
expression was notably increased in MSpf-treated DLD-1 cells
compared with control cells, whereas cleaved caspase-3 expression
was not detected.
In addition, it was determined whether MSpf
treatment was associated with cell morphology. MSpf-treated cells
exhibited a more uniform and adherent monolayer compared to the
controls (Fig. 4B). Therefore, it
was hypothesized that MSpf treatment may inhibit cell migration and
invasion by decreasing epithelial-mesenchymal transition (EMT). To
investigate this, the expression of EMT markers E-cadherin,
N-cadherin and Snail in MSpf-treated DLD-1 cells was determined.
E-cadherin expression was increased in MSpf-treated DLD-1 cells
compared with control cells, whereas N-cadherin and Snail
expression were not affected by MSpf treatment (Fig. 4C). The expression levels of MMP-2
and MMP-9 in MSpf-treated DLD-1 cells were examined to investigate
the potential association with the inhibition of cell invasion
following MSpf treatment. MMP-9 expression decreased in
MSpf-treated DLD-1 cells compared with control cells, whereas MMP-2
expression was not affected by MSpf treatment (Fig. 4C).
Determination of AGEs in MSpf
To clarify the molecular mechanism of the effects of
MSpf, proteins in MSpf were analyzed using proteomics (data not
shown). However, identification was not possible due to
insufficient registration of the proteins in the SwissProt database
against Maple tree) taxonomy. It has been reported that the free
amino groups of side chains of amino acid in proteins react with
sugars due to the Maillard reaction during the heat treatment, and
this led to formation of brown colored AGEs (29,30).
As maple syrup is made by boiling sap, the obtained protein
fraction has a dark brown color (Fig.
2B); therefore, MSpf might contain AGEs. In addition, maple
syrup contains various unique sugars rarely found in natural
products (31). Therefore, it was
hypothesized that the AGEs in MSpf generated by such unique
carbohydrate linkages had unique effect such an antitumor effect.
The quantity of AGEs in MSpf was examined by ELISA; the mean
concentration of AGEs in MSpf was 0.64±0.39 mg/ml. As protein
concentration in MSpf was (0.50±0.05 mg/ml), almost all protein
components in MSpf were predicted to be AGEs (127.07±45.33%).
Effects of MSpf on induction of
autophagy in DLD-1 cells
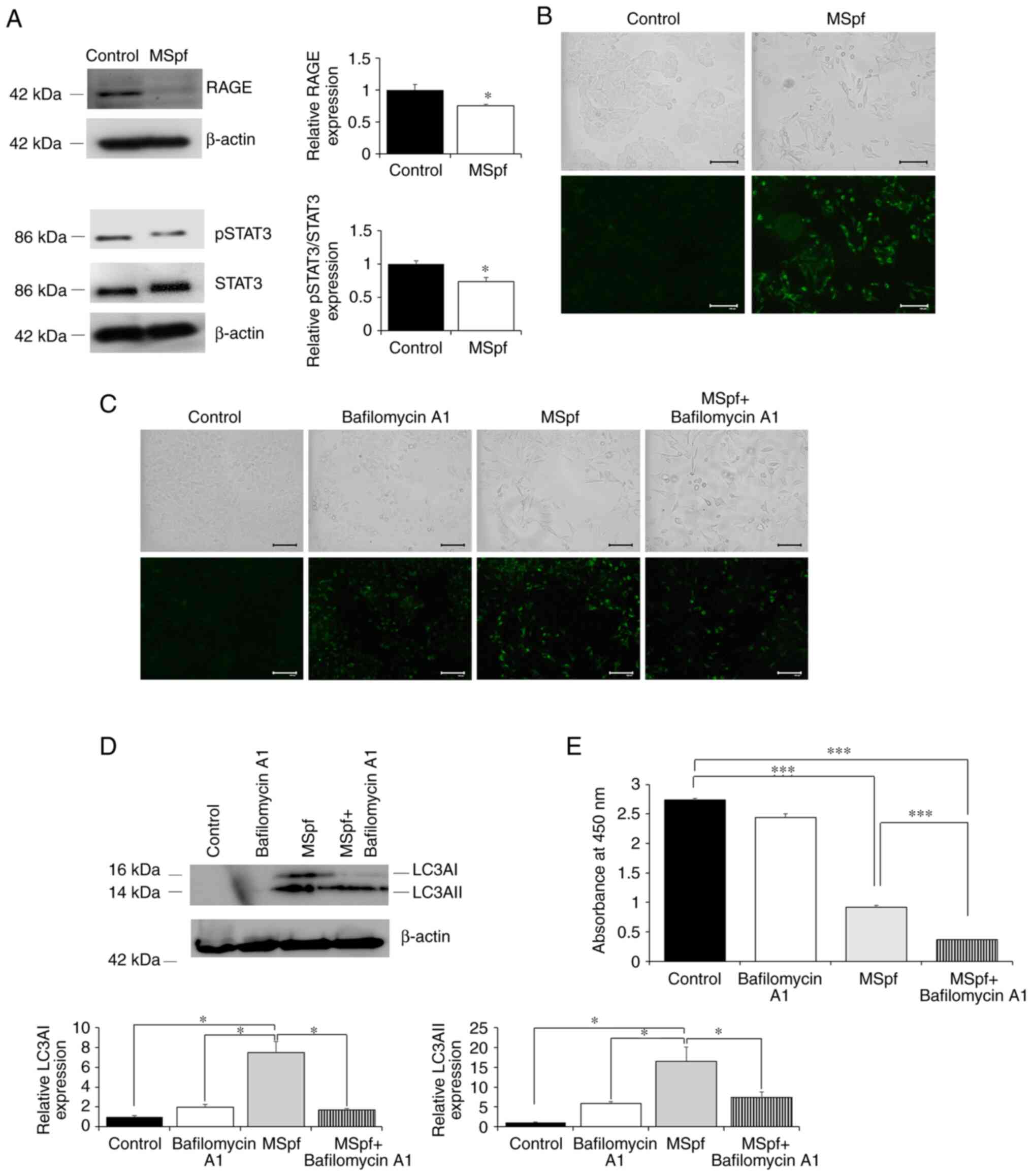
AGEs bind to their receptor RAGE and activate
intracellular transcription factors (32). Previous reports have shown that
autophagy is induced by regulation of the RAGE/STAT3 pathway
(33–36). To determine whether MSpf was
associated with the regulation of this pathway, the expression of
RAGE and pSTAT3 were determined. RAGE and pSTAT3 expression were
decreased in MSpf-treated DLD-1 cells compared with control cells
(Fig. 5A).
To investigate whether autophagy was induced by
MSpf, the effect of MSpf treatment on autophagosome formation was
assessed using DAPGreen staining. MSpf treatment increased the
DAPGreen-positive cells compared with control cells, which
suggested induction of autophagy by MSpf (Fig. 5B). To confirm the effect of MSpf
treatment on induction of autophagy, bafilomycin A1 (a specific
autophagy inhibitor of autophagosome-lysosome fusion; 24) was used.
Autophagy was successfully inhibited, as shown by an increase in
autophagosome formation following bafilomycin A1 treatment
(Fig. 5C; control and bafilomycin
A1). However, autophagosome formation induced by MSpf treatment was
not enhanced by bafilomycin A1 treatment (Fig. 5C; MSpf and MSpf+ bafilomycin A1).
This was confirmed by protein expression analysis of autophagic
marker LC3A (Fig. 5D). The
expression of LC3AI and II in MSpf-treated DLD-1 cells was
significantly induced compared with control, bafilomycin A1-treated
and MSpf+ bafilomycin A1-treated cells while there were no
significant differences among other treated cells. Conversely, the
inhibitory effect of MSpf treatment on cell viability was enhanced
by inhibition of autophagy when cells were treated with bafilomycin
A1 (Fig. 5E).
Additionally, MSpf treatment activated Akt, whereas
no effect on ERK, JNK and p38 phosphorylation was observed
(Fig. S1).
Discussion
The present study investigated the active
ingredients in maple syrup with putative antitumor effects against
CRC cells using an approach to isolate proteins that did not use
organic solvent-based extraction. Polysaccharides and/or proteins
were hypothesized to be the majority constituents in the high MW
component of the maple syrup. According to previously reported
saccharide profiles in maple syrup (31,37),
it consists primarily of neutral sugars. It was hypothesized that
polysaccharides that are polymerized to these neutral sugars are
insoluble. Therefore, the protein components of maple syrup were
suggested to be the active ingredients in maple syrup. MSpf was
obtained using ammonium sulfate precipitation, and MSpf treatment
suppressed proliferation of DLD-1 colon cancer cells, without
affecting viability of normal colon cells. Moreover, MSpf treatment
also suppressed migration and invasion of DLD-1 cells. As cell
migratory and invasive capacities are key for cancer metastasis,
which is an advanced stage of tumor progression, MSpf may be a
useful resource for developing a therapeutic drug for management of
CRC, including metastatic CRC.
The molecular mechanisms involved in cell
proliferation and metastasis underlying cancer progression are
multifactorial. The present study determined whether apoptosis or
autophagy was induced by MSpf treatment. The increase in LC3AI and
LC3AII protein expression, which are autophagy markers that serve a
key role in the formation of autophagosomes (27,28),
suggested that MSpf resulted in DLD-1 cell autophagy.
Conversely, EMT serves an important role in
conferring metastatic properties to cancer cells by facilitating
their migratory and invasive capabilities (38–41).
Thus, inhibition of cell migration and invasion by MSpf treatment
may be associated with inhibition of EMT. During EMT progression,
expression of epithelial marker proteins, such as E-cadherin, is
decreased in cancer cells (38,42,43).
By contrast, increased expression of mesenchymal markers, such as
N-cadherin and MMP-2 and MMP-9, is frequently observed in cancer
cells (44,45). Increased E-cadherin and decreased
MMP-9 expression were observed in the present study following MSpf
treatment. In addition, recent studies have suggested that
autophagy and EMT are associated with regulation of tumorigenesis
and tumor progression (46,47). Taken together, these results
suggested that MSpf treatment regulated induction of autophagy and
suppression of EMT in DLD-1 cells and that MSpf treatment may be a
valuable therapeutic strategy for CRC.
In the present study, almost all protein components
in the MSpf were predicted to be AGEs based on protein
concentration of obtained MSpf. AGEs are typically created by
non-enzymatic condensation between carbonyl groups of reducing
sugars and free amine groups from proteins, nucleic acids or
lipids, followed by further rearrangement (48,49).
Sato et al (31) reported
that maple syrup contains various unique sugars rarely found in
natural products. Therefore, AGEs generated by such unique
carbohydrate linkages may exhibit antitumor effects against the CRC
cells used in the present study. Further studies are necessary to
clarify what sugars are linked to AGEs and determine the effect of
the AGEs contained in or developed from MSpf.
AGEs exert various functions by binding to RAGE such
as aging-associated inflammation and oxidative stress (50,51).
The present study results suggested that MSpf inhibited the STAT3
signaling pathway through the decreased expression of RAGE. Recent
studies have reported that inhibition of STAT3 phosphorylation is
associated with induction of autophagy (34,35);
in the present study, autophagy was induced by MSpf. Autophagy is a
process that regulates cancer survival during stress (52). Several studies have suggested that
autophagy induction inhibits tumor initiation and progression
(53,54). Therefore, whether the inhibitory
effect of MSpf treatment on cell proliferation was due to induction
of autophagy was examined. The inhibitory effect of MSpf treatment
on cell viability was enhanced as a result of using a specific
autophagy inhibitor. This suggested that autophagy was induced to
protect cancer cells against MSpf treatment. Since inhibition of
the STAT3 signaling pathway is associated with not only induction
of autophagy but also induction of apoptosis and cell cycle arrest
(55–57), in addition to our previous study
showing that maple syrup treatment induces S-phase cell cycle
arrest (14), AGEs in MSpf may have
induced cell cycle arrest.
Conversely, inhibition of the STAT3 signaling
pathway is associated with increased expression of E-cadherin
(58–60), decreased expression of MMP-9
(61–63) and suppressed induction of EMT
(58). Therefore, AGEs in MSpf may
suppress EMT by decreased STAT3 signaling through the reduction of
RAGE. However, the mechanism underlying suppression of RAGE
expression by MSpf remains unclear. Further studies are needed to
elucidate the signaling pathways associated with regulation of RAGE
expression.
To the best of our knowledge, however, the role of
activated Akt in the antitumor effects of MSpf has not been
examined previously and thus further studies are required to
elucidate the effects of Akt activation following MSpf treatment.
In addition, MSpf treatment did not affect normal colon epithelial
cells in the present study. Moreover, it is possible to ingest
protein components equivalent to the effective concentration (10
µg/ml) by ingesting a small amount of maple syrup. Therefore, it is
necessary to clarify the effectiveness and safety of MSpf in
vivo, as well as metabolomics of maple syrup consumption.
In conclusion, the present study demonstrated that
MSpf inhibited DLD-1 cell proliferation, migration and invasion
through inhibition of the STAT3 signaling pathway. Furthermore,
AGEs in MSpf had antitumor effects. The results suggested that AGEs
in MSpf may serve as potential compounds for the development of
antitumor therapy, and MSpf may be useful in designing strategies
for improving CRC prognosis and therapy.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
All data generated and/or analyzed during the
present study are included in the published article.
Authors' contributions
AT conceived the study. KM and TY designed the
study. TY, RS, and YM performed data analysis. TY, RS, YM, KM and
AT performed the experiments. TY and AT collected the data. TY
drafted the manuscript. TY, KM, and AT reviewed and edited the
manuscript. TY and KM confirm the authenticity of all the raw data.
All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AGE
|
advanced glycation end product
|
|
CRC
|
colorectal cancer
|
|
EMT
|
epithelial-mesenchymal transition
|
|
MMP
|
matrix metalloproteinase
|
References
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD, Fuchs HE and Jemal
A: Cancer statistics, 2022. CA Cancer J Clin. 72:7–33. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dutta S, Mahalanobish S, Saha S, Ghosh S
and Sil PC: Natural products: An upcoming therapeutic approach to
cancer. Food Chem Toxicol. 128:240–255. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cheraghi O, Dehghan G, Mahdavi M,
Rahbarghazi R, Rezabakhsh A, Charoudeh HN, Iranshahi M and
Montazersaheb S: Potent anti-angiogenic and cytotoxic effect of
conferone on human colorectal adenocarcinoma HT-29 cells.
Phytomedicine. 23:398–405. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tundis R and Loizzo MR: A review of the
traditional uses, phytochemistry and biological activities of the
genus santolina. Planta Med. 84:627–637. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Castro-Puyana M, Pérez-Sánchez A, Valdés
A, Ibrahim OHM, Suarez-Álvarez S, Ferragut JA, Micol V, Cifuentes
A, Ibáñez E and García-Cañas V: Pressurized liquid extraction of
Neochloris oleoabundans for the recovery of bioactive carotenoids
with anti-proliferative activity against human colon cancer cells.
Food Res Int. 99:1048–1055. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nimalaratne C, Blackburn J and Lada RR: A
comparative physicochemical analysis of maple (Acer
saccharum Marsh.) syrup produced in North America with special
emphasis on seasonal changes in Nova Scotia maple syrup
composition. J Food Compos Anal. 92:1035732020. View Article : Google Scholar
|
|
9
|
Saraiva A, Carrascosa C, Ramos F, Raheem
D, Lopes M and Raposo A: Maple syrup: Chemical analysis and
nutritional profile, health impacts, safety and quality control,
and food industry applications. Int J Environ Res Public Health.
19:136842022. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhu K, Aykas DP and Rodriguez-Saona LE:
Pattern recognition approach for the screening of potential
adulteration of traditional and bourbon barrel-aged maple syrups by
spectral fingerprinting and classical methods. Foods. 11:22112022.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
St-Pierre P, Pilon G, Dumais V, Dion C,
Dubois MJ, Dubé P, Desjardins Y and Marette A: Comparative analysis
of maple syrup to other natural sweeteners and evaluation of their
metabolic responses in healthy rats. J Funct Foods. 11:460–471.
2014. View Article : Google Scholar
|
|
12
|
Nagai N, Yamamoto T, Tanabe W, Ito Y,
Kurabuchi S, Mitamura K and Taga A: Changes in plasma glucose in
Otsuka Long-Evans Tokushima Fatty rats after oral administration of
maple syrup. J Oleo Sci. 64:331–335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yamamoto T, Uemura K, Moriyama K, Mitamura
K and Taga A: Inhibitory effect of maple syrup on the cell growth
and invasion of human colorectal cancer cells. Oncol Rep.
33:1579–1584. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yamamoto T, Nishita T and Taga A:
Dark-colored maple syrup treatment induces S-phase cell cycle
arrest via reduced proliferating cell nuclear antigen expression in
colorectal cancer cells. Oncol Lett. 17:2713–2720. 2019.PubMed/NCBI
|
|
15
|
Yamamoto T, Sato K, Kubota Y, Mitamura K
and Taga A: Effect of dark-colored maple syrup on cell
proliferation of human gastrointestinal cancer cell. Biomed Rep.
7:6–10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Legault J, Girard-Lalancette K, Grenon C,
Dussault C and Pichette A: Antioxidant activity, inhibition of
nitric oxide overproduction, and in vitro antiproliferative effect
of maple sap and syrup from Acer saccharum. J Med Food.
13:460–468. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
González-Sarrías A, Li L and Seeram NP:
Anticancer effects of maple syrup phenolics and extracts on
proliferation, apoptosis, and cell cycle arrest of human colon
cells. J Funct Foods. 4:185–196. 2012. View Article : Google Scholar
|
|
18
|
González-Sarrías A, Ma H, Edmonds ME and
Seeram NP: Maple polyphenols, ginnalins A-C, induce S- and
G2/M-cell cycle arrest in colon and breast cancer cells mediated by
decreasing cyclins A and D1 levels. Food Chem. 136:636–642. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Perkins TD and van den Berg AK: Maple
syrup-production, composition, chemistry, and sensory
characteristics. Adv Food Nutr Res. 56:101–143. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Y, Yuan T, Li L, Nahar P, Slitt A
and Seeram NP: Chemical compositional, biological, and safety
studies of a novel maple syrup derived extract for nutraceutical
applications. J Agric Food Chem. 62:6687–6698. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Camara M, Cournoyer M, Sadiki M and Martin
N: Characterization and removal of buddy off-flavor in maple syrup.
J Food Sci. 84:1538–1546. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sabik H, Fortin J and Martin N:
Identification of pyrazine derivatives in a typical maple syrup
using headspace solid-phase microextraction with gas
chromatography-mass spectrometry. Food Chem. 133:1006–1010. 2012.
View Article : Google Scholar
|
|
23
|
Toufeili I, Itani M, Zeidan M, Al Yamani O
and Kharroubi S: Nutritional and functional potential of carob
syrup versus date and maple syrups. Food Technol Biotechnol.
60:266–278. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Luo X, Dan W, Luo X, Zhu X, Wang G, Ning
Z, Li Y, Ma X, Yang R, Jin S, et al: Caveolin 1-related autophagy
initiated by aldosterone-induced oxidation promotes liver
sinusoidal endothelial cells defenestration. Redox Biol.
13:508–521. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Siemens H, Jackstadt R, Hünten S, Kaller
M, Menssen A, Götz U and Hermeking H: miR-34 and SNAIL form a
double-negative feedback loop to regulate epithelial-mesenchymal
transitions. Cell Cycle. 10:4256–4271. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Silva FFVE, Padín-Iruegas ME, Caponio VCA,
Lorenzo-Pouso AI, Saavedra-Nieves P, Chamorro-Petronacci CM,
Suaréz-Peñaranda J and Pérez-Sayáns M: Caspase 3 and cleaved
caspase 3 expression in tumorogenesis and its correlations with
prognosis in head and neck cancer: A systematic review and
meta-analysis. Int J Mol Sci. 23:119372022. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kim YK, Shin JS and Nahm MH: NOD-like
receptors in infection, immunity, and diseases. Yonsei Med J.
57:5–14. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen RJ, Lee YH, Yeh YL, Wang YJ and Wang
BJ: The roles of autophagy and the inflammasome during
environmental stress-triggered skin inflammation. Int J Mol Sci.
17:20632016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Henning C and Glomb MA: Pathways of the
Maillard reaction under physiological conditions. Glycoconj J.
33:499–512. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Teodorowicz M, van Neerven J and Savelkoul
H: Food processing: The Influence of the Maillard reaction on
immunogenicity and allergenicity of food proteins. Nutrients.
9:8352017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sato K, Nagai N, Yamamoto T, Mitamura K
and Taga A: Identification of a novel oligosaccharide in maple
syrup as a potential alternative saccharide for diabetes mellitus
patients. Int J Mol Sci. 20:50412019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hollenbach M: The role of glyoxalase-I
(Glo-I), advanced glycation endproducts (AGEs), and their receptor
(RAGE) in chronic liver disease and hepatocellular carcinoma (HCC).
Int J Mol Sci. 18:24662017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen RJ, Lyu YJ, Chen YY, Lee YC, Pan MH,
Ho YS and Wang YJ: Chloroquine potentiates the anticancer effect of
pterostilbene on pancreatic cancer by inhibiting autophagy and
downregulating the RAGE/STAT3 pathway. Molecules. 26:67412021.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kang R, Loux T, Tang D, Schapiro NE,
Vernon P, Livesey KM, Krasinskas A, Lotze MT and Zeh HJ III: The
expression of the receptor for advanced glycation endproducts
(RAGE) is permissive for early pancreatic neoplasia. Proc Natl Acad
Sci USA. 109:7031–7036. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sheu ML, Pan LY, Hu HY, Su HL, Sheehan J,
Tsou HK and Pan HC: Potential therapeutic effects of
thiazolidinedione on malignant glioma. Int J Mol Sci. 23:135102022.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li JK, Zhu PL, Wang Y, Jiang XL, Zhang Z,
Zhang Z and Yung KK: Gracillin exerts anti-melanoma effects in
vitro and in vivo: Role of DNA damage, apoptosis and autophagy.
Phytomedicine. 108:1545262023. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sato K, Yamamoto T, Mitamura K and Taga A:
Separation of fructosyl oligosaccharides in maple syrup by using
charged aerosol detection. Foods. 10:31602021. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zeisberg M and Neilson EG: Biomarkers for
epithelial-mesenchymal transitions. J Clin Invest. 119:1429–1437.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mittal V: Epithelial mesenchymal
transition in tumor metastasis. Annu Rev Pathol. 13:395–412. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu J, Chen XF, Zhou XR, Yi RK, Yang ZN
and Zhao X: Lactobacillus fermentum ZS09 mediates
epithelial-mesenchymal transition (EMT) by regulating the
transcriptional activity of the Wnt/β-catenin signalling pathway to
inhibit colon cancer activity. J Inflamm Res. 14:7281–7293. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chae U, Kim B, Kim H, Park YH, Lee SH, Kim
SU and Lee DS: Peroxiredoxin-6 regulates p38-mediated
epithelial-mesenchymal transition in HCT116 colon cancer cells. J
Biol Res (Thessalon). 28:222021. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wheelock MJ, Shintani Y, Maeda M, Fukumoto
Y and Johnson KR: Cadherin switching. J Cell Sci. 121:727–735.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Mao L, Yang J, Yue J, Chen Y, Zhou H, Fan
D, Zhang Q, Buraschi S, Iozzo RV and Bi X: Decorin deficiency
promotes epithelial-mesenchymal transition and colon cancer
metastasis. Matrix Biol. 95:1–14. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Shay G, Lynch CC and Fingleton B: Moving
targets: Emerging roles for MMPs in cancer progression and
metastasis. Matrix Biol. 44–46. 200–206. 2015.
|
|
45
|
Lin TA, Lin WS, Chou YC, Nagabhushanam K,
Ho CT and Pan MH: Oxyresveratrol inhibits human colon cancer cell
migration through regulating epithelial-mesenchymal transition and
microRNA. Food Funct. 12:9658–9668. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lai MF, Liu LL, Zhu L, Feng W, Luo J, Liu
Y and Deng S: Triptolide reverses epithelial-mesenchymal transition
in glioma cells via inducing autophagy. Ann Transl Med. 9:13042021.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li Q, Liu S, Yang G, Li M, Qiao P and Xue
Q: Naringenin inhibits autophagy and epithelial-mesenchymal
transition of human lens epithelial cells by regulating the Smad2/3
pathway. Drug Dev Res. 83:389–396. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Twarda-Clapa A, Olczak A, Bialkowska AM
and Koziolkiewicz M: Advanced glycation end-products (AGEs):
Formation, chemistry, classification, receptors, and diseases
related to AGEs. Cells. 11:13122022. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Vistoli G, De Maddis D, Cipak A, Zarkovic
N, Carini M and Aldini G: Advanced glycoxidation and lipoxidation
end products (AGEs and ALEs): An overview of their mechanisms of
formation. Free Radical Res. 47 (Suppl 1):S3–S27. 2013. View Article : Google Scholar
|
|
50
|
Basta G, Schmidt AM and De Caterina R:
Advanced glycation end products and vascular inflammation:
Implications for accelerated atherosclerosis in diabetes.
Cardiovasc Res. 63:582–592. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Chen CY, Abell AM, Moon YS and Kim KH: An
advanced glycation end product (AGE)-receptor for AGEs (RAGE) axis
restores adipogenic potential of senescent preadipocytes through
modulation of p53 protein function. J Biol Chem. 287:44498–44507.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Galluzzi L, Pietrocola F, Bravo-San Pedro
JM, Amaravadi RK, Baehrecke EH, Cecconi F, Codogno P, Debnath J,
Gewirtz DA, Karantza V, et al: Autophagy in malignant
transformation and cancer progression. EMBO J. 34:856–880. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Liu Z, Chen P, Gao H, Gu Y, Yang J, Peng
H, Xu X, Wang H, Yang M, Liu X, et al: Ubiquitylation of autophagy
receptor Optineurin by HACE1 activates selective autophagy for
tumor suppression. Cancer Cell. 26:106–120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kong W, Zhu H, Zheng S, Yin G, Yu P, Shan
Y, Liu X, Ying R, Zhu H and Ma S: Larotrectinib induces autophagic
cell death through AMPK/mTOR signalling in colon cancer. J Cell Mol
Med. 26:5539–5550. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Huang B, Lang X and Li X: The role of
IL-6/JAK2/STAT3 signaling pathway in cancers. Front Oncol.
12:10231772022. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Hashemi M, Sabouni E, Rahmanian P,
Entezari M, Mojtabavi M, Raei B, Zandieh MA, Behroozaghdam M,
Mirzaei S, Hushmandi K, et al: Deciphering STAT3 signaling
potential in hepatocellular carcinoma: Tumorigenesis, treatment
resistance, and pharmacological significance. Cell Mol Biol Lett.
28:332023. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Al-Hetty HRAK, Abdulameer SJ, Alkubaisy
SA, Zaid SA, Jalil AT and Jasim IK: STAT3 signaling in pancreatic
ductal adenocarcinoma: A candidate therapeutic target. Pathol Res
Pract. 245:1544252023. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Czikora Á, Erdélyi K, Ditrói T, Szántó N,
Jurányi EP, Szanyi S, Tóvári J, Strausz T and Nagy P: Cystathionine
β-synthase overexpression drives metastatic dissemination in
pancreatic ductal adenocarcinoma via inducing
epithelial-to-mesenchymal transformation of cancer cells. Redox
Biol. 57:1025052022.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Guo H, Zheng L, Guo Y, Han L, Yu J and Lai
F: Curculigoside represses the proliferation and metastasis of
osteosarcoma via the JAK/STAT and NF-κB signaling pathways. Biol
Pharm Bull. 45:1466–1475. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Cui B, Yu C, Zhang S, Hou X, Wang Y, Wang
J, Zhuang S and Liu F: Delayed administration of nintedanib
ameliorates fibrosis progression in CG-induced peritoneal fibrosis
mouse model. Kidney Dis (Basel). 8:319–333. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Fang W, Huang J, Wang J, Huang T, Lin D
and Yin J: Blockade of interleukin-6 receptor attenuates apoptosis
and modulates the inflammatory response in Mycoplasma pneumoniae
infected A549 cells. Am J Transl Res. 14:6187–6195. 2022.PubMed/NCBI
|
|
62
|
Tossetta G, Fantone S, Busilacchi EM, Di
Simone N, Giannubilo SR, Scambia G, Giordano A and Marzioni D:
Modulation of matrix metalloproteases by ciliary neurotrophic
factor in human placental development. Cell Tissue Res.
390:113–129. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Wu K, Wu X, Liang Y, Wang T, Wu D, Li L
and Wang Z: Inhibitory effects of total triterpenoids isolated from
the Hedyotis diffusa willd on H1975 cells. Front Pharmacol.
13:9224772022. View Article : Google Scholar : PubMed/NCBI
|