Introduction
Ischemia-reperfusion (I/R) injury is a common
occurrence in liver surgery. Patients undergoing liver
transplantation may encounter severe postoperative complications,
namely primary liver dysfunction or non-function despite the long
waiting lists and high rate of mortality in those awaiting a
compatible donor (1). In addition,
patients subjected to major hepatectomy also have a high risk of
developing liver dysfunction, in which portal triad occlusion is
routinely utilized to reduce blood loss (2). Among the multiple pathogeneses
accounting for these complications, I/R induced injury is widely
accepted as an important contributor (1,2).
Sorbitol dehydrogenase (SDH), a key enzyme in the
polyol pathway, has traditionally been implicated in the
development of vascular and neurological complications in diabetes.
In this pathway, SDH converts sorbitol to fructose with the
concomitant reduction of coenzyme nictotinamide adenine
dinucleotide (NAD) to NADH (3).
Previously, accumulating evidence has demonstrated the beneficial
impact of a sorbitol dehydrogenase inhibitor (SDI) on ischemic
tissue (3–5). The potential mechanisms underlying
this effect have not been fully elucidated. In addition the role of
SDH in the ischemic liver remains unclear. Therefore, the present
study aimed to investigate how SDH may affect livers subjected to
I/R insult and the mechanism underling its effect.
Materials and methods
Ethical approval
The present study was approved by the Nanjing
Medical University Experimental Animal Department (no.
NJMU-AEARIA-4001-20120401; Nanjing, Jiangsu, China) and closely
followed the Guide for the Care and Use of Laboratory Animals
published by the US National Institutes of Health (NIH publication
no. 85-23, revised 1996).
Reagents
SDI (CP-470, 711) was obtained from Pfizer Global
Research and Development (Groton, CT, USA). The NAD(H) and
adenosine triphosphate (ATP) Quantification kits were purchased
from BioVision (Milpitas, CA, USA). The Annexin V-fluorescein
isothiocyanate (FITC)/propidium iodide (PI) apoptosis detection kit
was purchased from BD Biosciences (Bedford, MA, USA). Interleukin
(IL)-1β (catalogue no. MLB00C) and tumor necrosis factor (TNF)-α
ELISA assay kits (catalogue no. DY410-05) were obtained from
R&D Systems (Minneapolis, MN, USA). Primary monoclonal rabbit
anti-caspase 3 and secondary monoclonal mouse anti-rabbit
antibodies were purchased from Cell Signaling Technology, Inc.
(Danvers, MA, USA). Primary monoclonal rabbit antibodies against
SDH, SIRT1 and β-actin were purchased from Abcam (Cambridge, MA,
USA). The bicinchoninic acid (BCA) protein assay kit were purchased
from the Beyotime Institute of Biotechnology (Nanjing, China).
Animals and I/R protocol
Wild-type C57BL/6 male mice were obtained from the
Chinese Academy of Sciences (Shanghai, China), maintained in 25±2°C
under a 12 h dark/light cycle. The mice were randomized into three
groups containing ten animals each: (i) The SDI group, administered
CP-470,711 (5 mg/kg body weight/day) by gavage for five days and
subjected to 70% liver I/R; (ii) the control group, treated with
isoquant saline control and subjected to 70% liver I/R; (iii) the
sham group received isoquant CP-470,711 by gavage and sham
operation. The usage of CP-470,711 was determined with reference to
previous studies (6,7). A total of 24 h following the last
drug treatment, 50% of the animals from each group underwent 1 h of
70% hepatic ischemia as previously described (8) and ischemic liver lobes were harvested
to examine the real-time intrahepatic energy status. The other five
mice from each group were subjected to an additional 24 h of
reperfusion to detect hepatic tissue impairment and changes in
protein levels. The blood samples from mice that had undergone I/R
were collected to investigate the levels of inflammatory markers.
The animals were eventually sacrificed under deep anesthesia
induced by peritoneal chlorali hydras (0.6 g/kg body weight).
Liver function test
The blood samples were immediately centrifuged at
2,000 × g to obtain serum following collection. The serum alanine
aminotransferase (ALT) and aspartate aminotransferase (AST) were
then automatically measured on the Aeroset autoanalyzer (Abbott
Laboratories, Chicago, IL, USA) and reported in units per
liter.
Detecting serum inflammatory markers
The serum levels of IL-1β and TNF-α were
investigated in a 96-well microtitre plate using a commercial
enzyme-linked immunosorbent assay kit according to the
manufacturer’s instructions. The lower limits of IL-1β and TNF-α
detection were 2.31 and 1.88 pg/ml, respectively.
Morphological observations
The histological sections stained with hematoxylin
and eosin (H&E) were processed routinely. The degree of hepatic
necrosis and inflammation was evaluated by an experienced
pathologist using the Suzuki score, which evaluated the extent of
congestion, vacuolization and necrosis on a four-point scale for a
total score of 0–12 as previously described (9).
Flow cytometric analysis of the
proportion of hepatocytes undergoing necrosis and apoptosis
Single-hepatocyte suspensions were prepared from the
fresh liver using an enzymatic technique as previously described
(10). Cells were exposed to 0.04%
Trypan blue for 3 minutes and then observed under a light
microscope. Trypan Blue exclusion demonstrated that the cell
viability in all of the samples exceeded 89%. The degree of
apoptosis and necrosis in the hepatocytes was quantified by flow
cytometry using an Annexin V-FITC kit and FlowJo software (Tree
Star Inc., Ashland, OR, USA) as previously described (11). The cell percentage in the upper
right, upper left and lower right quadrants represented the
proportions of late apoptotic, necrotic and early apoptotic cells,
respectively. The total apoptosis ratio was then calculated by
addingthe proportions of late and early apoptotic cells.
Measuring cytosolic ATP and NAD(H)
contents
The hepatic contents of NAD(H) and the NAD/NADH
ratio were determined using NAD+/NADH Quantification Kit
(BioVision). Briefly, the liver samples were homogenized in
extraction buffer and centrifuged at 12,000 × g for 5 min. The
supernatant was filtered through 10-kDa Spin Columns (Catalogue no.
1997-25; BioVision) to remove the NADH-consuming enzyme NADase and
was then measured according to the manufacturer’s instructions.
The ATP content in the hepatocytes was detected by a
colorimetric method using a ATP Colorimetric/Fluorometric Assay kit
(BioVision), which consumes ATP to generate glycerol 3-phosphate,
according to the manufacturer’s instructions. To remove the innate
ATP-consuming enzymes, the homogenized samples (10 mg) were
filtered through the aforementioned spin column prior to being
inoculated into 96-well plates. Measurements were performed
according to the manufacturer’s instructions.
Immunoblotting assay
Total protein from the liver tissues following I/R
insult were analyzed by immunoblotting as previously described
(12). Prior to sample-loading,
the BCA Protein Assay kit was conventionally used to determine the
homogeneity of protein concentrations in different columns. The
primary antibodies were used at the following concentrations:
Anti-SDH (1:1,500), -caspase-3 (1:2,000), -SIRT1 (1:2,000) and
-β-actin (1:2,500). The relative protein levels were determined
using Image J software (NIH, Bethesda, MD, USA). β-actin served as
a reference protein
Statistical analysis
Data are expressed as the mean ± standard deviation.
Statistical analysis of the data was performed using the Duncan’s
test with SPSS 16.0 software (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference. All of the results were obtained from at least five
independent experiments.
Results
SDI protects mouse liver against
I/R-induced impairment
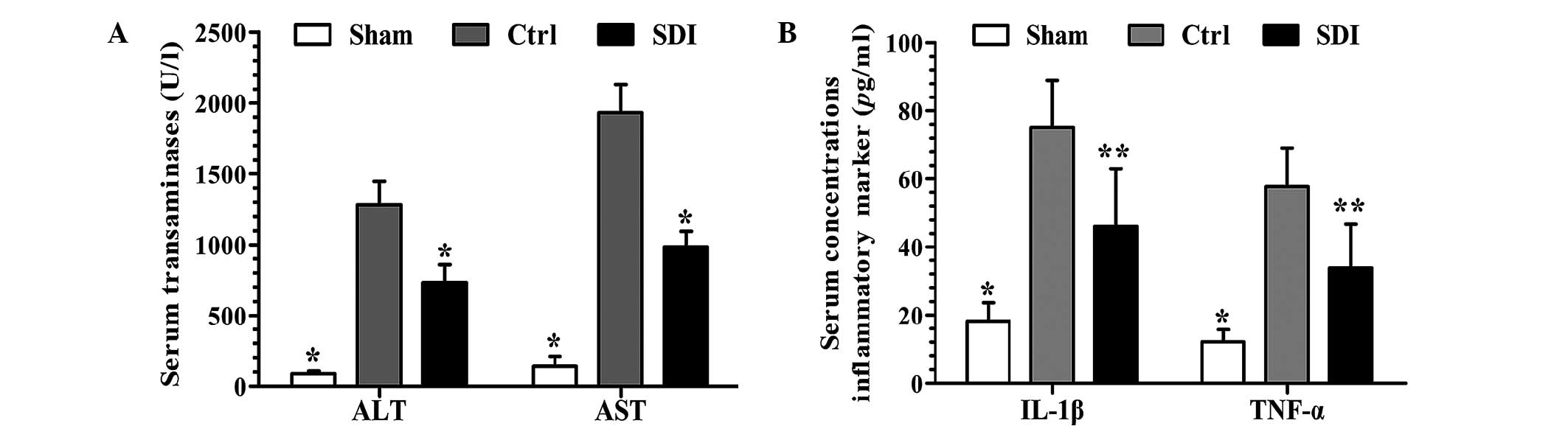
The transaminases ALT and AST are generated innately
within hepatocytes and marked elevations in serum levels denote
hepatocellular membrane leakage caused by inflammation and/or
necrosis. In the present study, I/R markedly increased the serum
transaminase levels, whereas SDI-pretreatment markedly ameliorated
these changes (Fig. 1A).
IL-1β and TNF-α are widely accepted to be
inflammatory markers and an increase in the serum level indicates a
severe inflammatory reaction and tissue injury (13). As demonstrated in Fig. 1B, SDI administration markedly
alleviated the I/R-mediated elevation in the serum level of IL-1β
and TNF-α as compared with the control group.
SDI-pretreatment prevents the I/R-induced
histological changes and improves hepatocyte fate
As direct causes of liver dysfunction following I/R,
hepatic inflammation, apoptosis and necrosis were analyzed in the
present study. As demonstrated in Fig.
2A, I/R insult significantly disrupted liver histology and
contributed to significant necrosis in contrast to the sham group,
where SDI markedly prevented the changes in hepatic histology.
Consistently, the quantitative Suzuki score in the control group
was also evidently higher than that in sham group and this trend
was significantly mitigated by SDI treatment (Fig. 2B). In the flow cytometric analysis,
the proportions of apoptotic and necrotic hepatocytes in the
control group were higher than the sham group, while SDI
significantly reverted these changes (Fig. 2C and D).
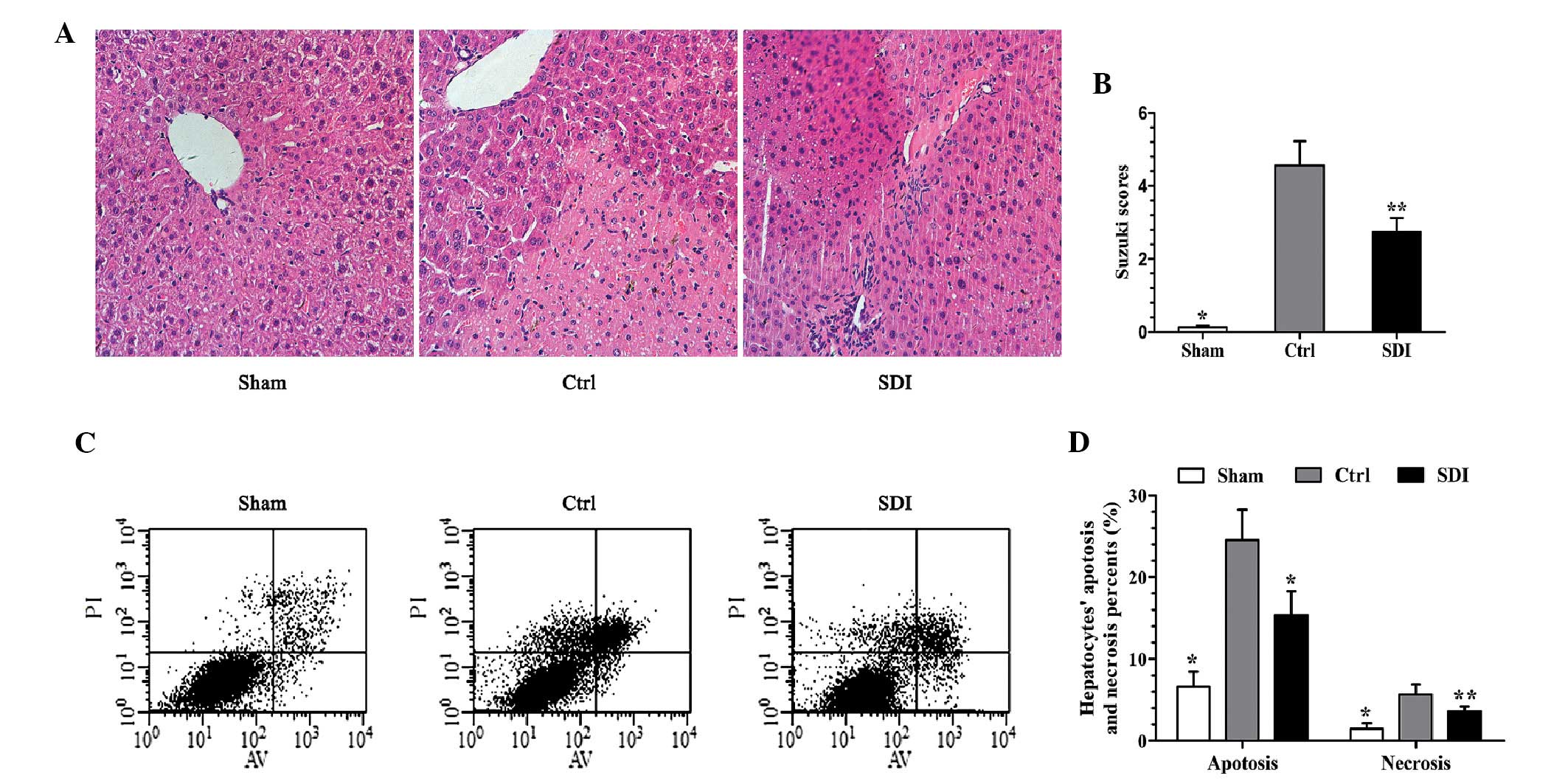 | Figure 2SDI-pretreatment normalized liver
histology and improved hepatocyte fate. (A and B) In hematoxylin
and eosin staining sections (magnification, ×400), the I/R insult
significantly disrupted liver histology and contributed to evident
necrosis, while SDI markedly prevented these changes. (B) The Ctrl
group demonstrated a higher Suzuki score than the sham group and
this trend was mitigated by SDI. (C and D) In flow cytometry, the
proportions of apoptotic and necrotic hepatocytes in the Ctrl group
were significantly higher than in the sham group, whereas SDI
inhibited these changes. n=5; *P<0.01 and
**P<0.05 as compared with the Ctrl group. SDI,
sorbitol dehydrogenase; I/R, ischemia/reperfusion; Ctrl, control;
AV, Annexin V; PI, propidium iodide. |
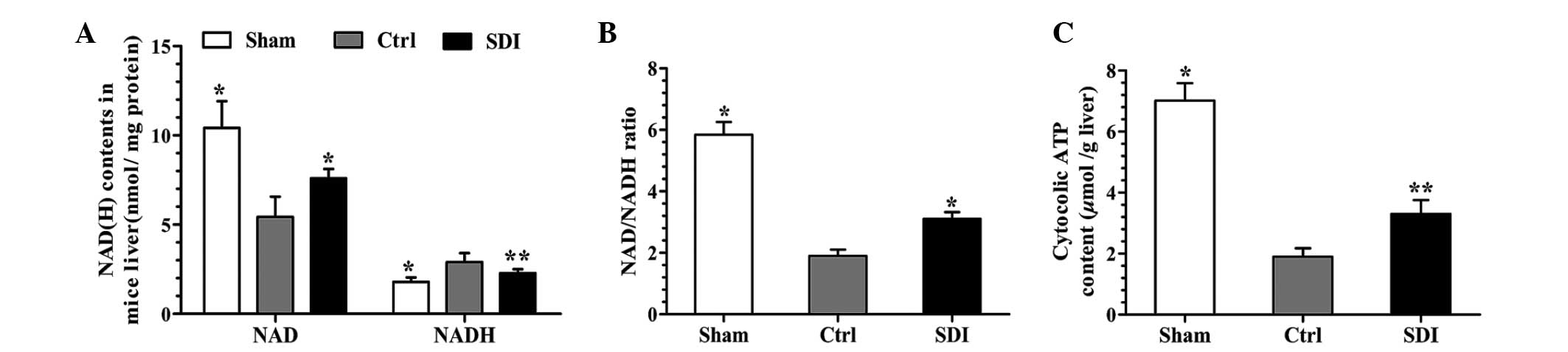
SDI reverses the I/R-mediated NAD/NADH
imbalance and energy deficit
The coenzyme NAD(H) participates in the polyol and
glycolysis pathways, the latter of which acts as a major ATP source
in the ischemic liver (3,14,15).
Therefore, the present study analyzed the NAD(H) and ATP content in
the ischemic liver, intending to examine the possible causal
association between SDI-administration, NAD(H) and ATP content. As
demonstrated in Fig. 3A–C, SDI
significantly alleviated the I/R-induced decrease in NAD and ATP
content and reversed the NAD/NADH imbalance compared with the
control group.
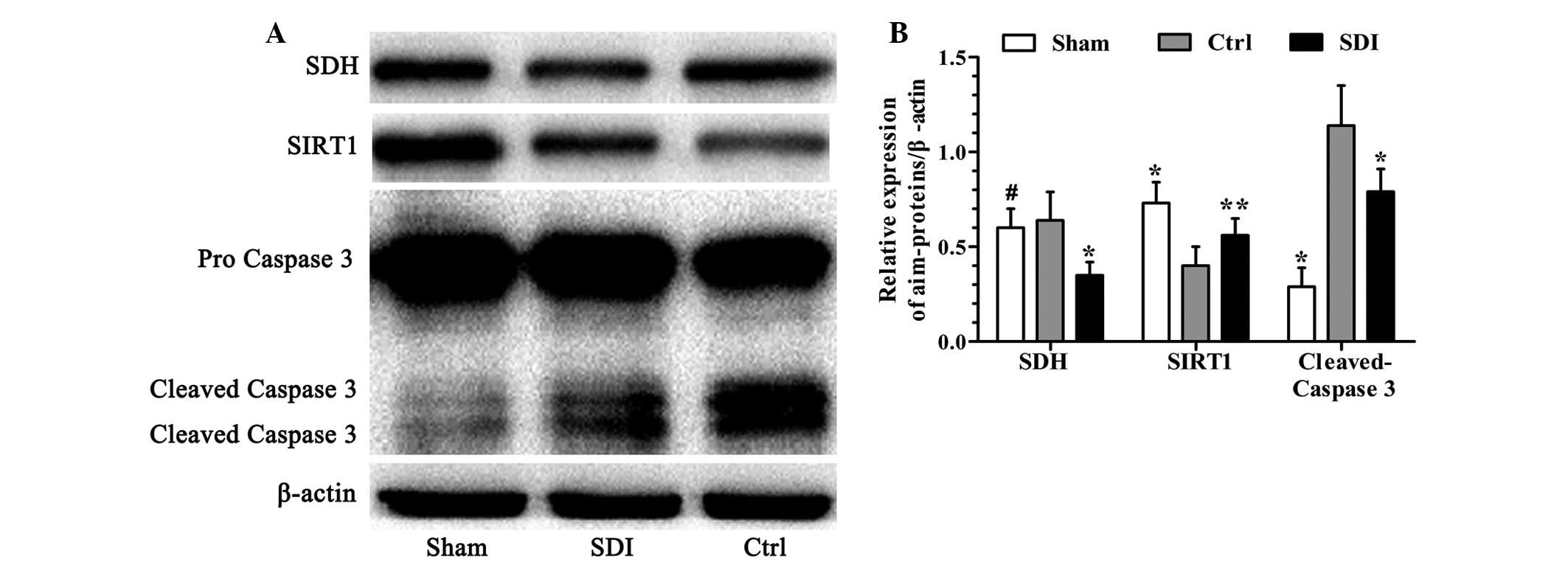
SDI enhances SIRT1 protein expression and
inhibits caspase 3 in the liver following I/R
As an important cellular function regulator and an
NAD-dependent enzyme, SIRT1 deacetylase was quantified by an
immunoblotting assay (16). As
demonstrated in Fig. 4A and B, I/R
markedly suppressed SIRT1 at the protein level while SDI alleviated
this change.
It is well-established that caspase 3 acts as a
major and common executor in multiple apoptotic pathways when cells
are exposed to cytotoxic stressors, including I/R (17,18).
The level of cleaved caspase 3 was then measured using western
blotting. As demonstrated in Fig. 4A
and B, SDI markedly suppressed the I/R-induced activation of
protein caspase 3 as compared with the control group.
Additionally, SDH protein was also quantified to
assess the inhibition efficiency of SDI. In the present study, the
sham and control groups exhibited a similar level of SDH protein,
which was notably higher than that in the SDI group (Fig. 4A and B).
Discussion
In the present study, it was firstly demonstrated
that SDI markedly alleviated liver I/R-induced impairment and
improved hepatocyte fate. These findings are consistent with those
of previous studies investigating other target organs (3–5). The
changes in NAD(H) content and proportion were markedly ameliorated
following SDI administration, providing evidence that these
beneficial effects may be associated with the improved co-enzyme
balance and resultant cellular metabolic and signaling
adaptations.
Under hypoxic conditions, low levels of ATP are
generated within the liver by glycolysis to sustain basic cellular
function (14,15,19).
In the glycolysis pathway, the key enzyme GAPDH exerts a catalytic
function dependent upon the synchronous reduction of NAD to NADH,
which denotes a competitive antagonistic correlation between SDH
and GAPDH due to the limited quantity of NAD (20,21).
Therefore, it is reasonable to hypothesize that inhibiting SDH may
prevent this competition for substrate and enhance the glycolysis
flux. This hypothesis has been widely validated by previous studies
based on models of ischemic heart, brain and kidney injury. In
these studies, the reduction in the levels of NAD and/or the
NAD/NADH ratio was caused by niacin deficiency or poly (ADP-ribose)
polymerase overactivation, which clearly aggravated tissue I/R
injury. The ARI/SDI pretreatment or NAD/niacin repletion markedly
reversed this phenomenon (3–5,22–25).
In the present study, pretreating ischemic mice with SDI also
contributed to an evident elevation in the NAD/NADH ratio and ATP
content. These results were consistent with the findings of
previous studies (3,4,22–24).
Hepatocyte inflammation, apoptosis and necrosis
represent different extremes in a continuum of cellular responses
to severe stressors (17,18). The major difference between them is
that inflammation refers mainly to the reaction of cells to
stimuli, while apoptosis and necrosis are cell death processes.
However, apoptosis is an active, ATP-consuming event, while
necrosis is passive, without ATP participation (13,18).
In the present study, intracellular ATP content was significantly
increased following SDI administration. This may explain why
histology and flow cytometry in the SDI group primarily
demonstrated inflammation and apoptosis, whereas the control group
demonstrated evidence of apoptosis and necrosis.
Aside from GAPDH, SIRT1 deacetylase represents
another NAD-dependent enzyme. SIRT1 is a member of the mammalian
sirtuin family and widely participates in cellular death modulating
activities, including mitochondrial function, poly (ADP-ribose)
polymerase-1 (PARP-1) cytotoxicity and apoptotic effects (20,22,26–29).
Therefore, due to its roles in delaying aging and anti-apoptosis,
SIRT1 is generally acknowledged to be a lifespan promoter,
preventing cell death. In the present study, SIRT1 protein
expression was suppressed by I/R injury, while SDI notably
inhibited this trend. Limited by the scale of the study, the
specific role of SIRT1 in the ischemic liver was not investigated.
However, it is hypothesized that the elevated SIRT1 flux may
contribute to reduced I/R-induced liver injury.
Accumulating evidence has suggested that NAD is a
fundamental mediator of various biological processes (20). Apart from the aforementioned
regulative activities, an appropriate proportion of NAD(H) is
required to maintain mitochondrial function and cellular calcium
homeostasis (3,20,30–32).
Furthermore, NAD also has a pivotal role in activating the
PARP-1-dependent death pathway (22,25,33).
Therefore, a paradoxical theme emerges: The SDI-induced NAD
elevation may protect the ischemic liver through improved
glycolysis and/or SIRT1 activities; conversely, increased cytosolic
NAD content may contribute to elevated PARP-1-mediated hepatocyte
apoptosis. Considering the variance in liver function, histology
and hepatocyte fate, it was hypothesized that SDI predominantly
exhibits a beneficial role in hepatic I/R injury. However, for
potential clinical consideration, further, more comprehensive
studies are required.
In conclusion, the present study demonstrated that
SDI protects the liver against I/R-induced injury in terms of
improved liver function, ameliorated histopathological changes and
reduced levels of hepatic necrosis and apoptosis. This effect may
be further associated with an elevated NAD/NADH ratio and resultant
elevation in glycolysis flux and SIRT1 expression. The present
study suggests that SDI may be used clinically to protect the liver
from severe tissue impairment following I/R injury.
Acknowledgements
The present was fully financed by the National
Natural Science Foundation of China (grant no. 81170415).
References
|
1
|
Elias-Miró M, Massip-Salcedo M, Raila J,
et al: Retinol binding protein 4 and retinol in steatotic and
nonsteatotic rat livers in the setting of partial hepatectomy under
ischemia/reperfusion. Liver Transpl. 18:1198–1208. 2012.PubMed/NCBI
|
|
2
|
Varona MA, Soriano A, Aguirre-Jaime A, et
al: Statistical quality control charts for liver transplant process
indicators: evaluation of a single-center experience. Transplant
Proc. 44:1517–1522. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tang WH, Wu S, Wong TM, Chung SK and Chung
SS: Polyol pathway mediates iron-induced oxidative injury in
ischemic-reperfused rat heart. Free Radic Biol Med. 45:602–610.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li Q, Hwang YC, Ananthakrishnan R, et al:
Polyol pathway and modulation of ischemia-reperfusion injury in
Type 2 diabetic BBZ rat hearts. Cardiovasc Diabetol. 7:332008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tang WH, Kravtsov GM, Sauert M, et al:
Polyol pathway impairs the function of SERCA and RyR in
ischemic-reperfused rat hearts by increasing oxidative
modifications of these proteins. J Mol Cell Cardiol. 49:58–69.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chu-Moyer MY, Ballinger WE, Beebe DA, et
al: SAR and species/stereo-selective metabolism of the sorbitol
dehydrogenase inhibitor, CP-470,711. Bioorg Med Chem Lett.
12:1477–1480. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schmidt RE, Dorsey DA, Beaudet LN, et al:
A potent sorbitol dehydrogenase inhibitor exacerbates sympathetic
autonomic neuropathy in rats with streptozotocin-induced diabetes.
Exp Neurol. 192:407–419. 2005. View Article : Google Scholar
|
|
8
|
Ramalho FS, Alfany-Fernandez I,
Casillas-Ramirez A, et al: Are angiotensin II receptor antagonists
useful strategies in steatotic and nonsteatotic livers in
conditions of partial hepatectomy under ischemia-reperfusion? J
Pharmacol Exp Ther. 329:130–140. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Suzuki S, Toledo-Pereyra LH, Rodriguez FJ
and Cejalvo D: Neutrophil infiltration as an important factor in
liver ischemia and reperfusion injury. Modulating effects of FK506
and cyclosporine. Transplantation. 55:1265–1272. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Berry MN and Friend DS: High-yield
preparation of isolated rat liver parenchymal cells: a biochemical
and fine structural study. J Cell Biol. 43:506–520. 1969.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang S, Chen T, Chen R, et al: Emodin
loaded solid lipid nanoparticles: preparation, characterization and
antitumor activity studies. Int J Pharm. 430:238–246. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee HR, Kim HJ, Ko JS, et al: Comparative
characteristics of porous bioceramics for an osteogenic response in
vitro and in vivo. PLoS One. 8:e842722013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nikoletopoulou V, Markaki M, Palikaras K
and Tavernarakis N: Crosstalk between apoptosis, necrosis and
autophagy. Biochim Biophys Acta. 1883:3448–3459. 2013. View Article : Google Scholar
|
|
14
|
So PW and Fuller BJ: Enhanced energy
metabolism during cold hypoxic organ preservation: studies on rat
liver after pyruvate supplementation. Cryobiology. 46:295–300.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Katz NR: Metabolic heterogeneity of
hepatocytes across the liver acinus. J Nutr. 122(Suppl): 843–849.
1992.PubMed/NCBI
|
|
16
|
Yu Q, Wang T, Zhou X, et al: Wld(S)
reduces paraquat-induced cytotoxicity via SIRT1 in non-neuronal
cells by attenuating the depletion of NAD. PLoS One. 6:e217702011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bantel H and Schulze-Osthoff K: Mechanisms
of cell death in acute liver failure. Front Physiol. 3:792012.
View Article : Google Scholar
|
|
18
|
Malhi H, Gores GJ and Lemasters JJ:
Apoptosis and necrosis in the liver: a tale of two deaths?
Hepatology. 43(Suppl 1): 31–44. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Brosnan JT, Krebs HA and Williamson DH:
Effects of ischaemia on metabolite concentrations in rat liver.
Biochem J. 117:91–96. 1970.PubMed/NCBI
|
|
20
|
Ying W: NAD+/NADH and NADP+/NADPH in
cellular functions and cell death: regulation and biological
consequences. Antioxid Redox Signal. 10:179–206. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dodson M, Darley-Usmar V and Zhang J:
Cellular metabolic and autophagic pathways: traffic control by
redox signaling. Free Radic Biol Med. 63:207–221. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ying W, Garnier P and Swanson RA: NAD+
repletion prevents PARP-1-induced glycolytic blockade and cell
death in cultured mouse astrocytes. Biochem Biophys Res Commun.
308:809–813. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hwang YC, Kaneko M, Bakr S, et al: Central
role for aldose reductase pathway in myocardial ischemic injury.
FASEB J. 18:1192–1199. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ramasamy R, Trueblood N and Schaefer S:
Metabolic effects of aldose reductase inhibition during low-flow
ischemia and reperfusion. Am J Physiol. 275:H195–H203.
1998.PubMed/NCBI
|
|
25
|
Benavente CA and Jacobson EL: Niacin
restriction upregulates NADPH oxidase and reactive oxygen species
(ROS) in human keratinocytes. Free Radic Biol Med. 44:527–537.
2008. View Article : Google Scholar
|
|
26
|
Dong W, Li F, Pan Z, et al: Resveratrol
ameliorates subacute intestinal ischemia-reperfusion injury. J Surg
Res. 185:182–189. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yan W, Fang Z, Yang Q, et al: SirT1
mediates hyperbaric oxygen preconditioning-induced ischemic
tolerance in rat brain. J Cereb Blood Flow Metab. 33:396–406. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lempiäinen J, Finckenberg P, Mervaala EE,
et al: Caloric restriction ameliorates kidney ischaemia/reperfusion
injury through PGC-1α-eNOS pathway and enhanced autophagy. Acta
Physiol (Oxf). 208:410–421. 2013.PubMed/NCBI
|
|
29
|
Pillai JB, Isbatan A, Imai S and Gupta MP:
Poly (ADP-ribose) polymerase-1-dependent cardiac myocyte cell death
during heart failure is mediated by NAD+ depletion and reduced
Sir2alpha deacetylase activity. J Biol Chem. 280:43121–43120. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Alano CC, Ying W and Swanson RA: Poly
(ADP-ribose) polymerase-1-mediated cell death in astrocytes
requires NAD+ depletion and mitochondrial permeability
transition. J Biol Chem. 279:18895–18902. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jang SY, Kang HT and Hwang ES:
Nicotinamide-induced mitophagy: event mediated by high
NAD+/NADH ratio and SIRT1 protein activation. J Biol
Chem. 287:19304–19314. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mouchiroud L, Houtkooper RH, Moullan N, et
al: The NAD+/sirtuin pathway modulates longevity through
activation of mitochondrial UPR and FOXO signaling. Cell.
154:430–441. 2013.
|
|
33
|
Siegel C and McCullough LD:
NAD+ depletion or PAR polymer formation: which plays the
role of executioner in ischaemic cell death? Acta Physiol (Oxf).
203:225–234. 2011.
|