Introduction
Artesunate (Art), a water-soluble hemisuccinate
derivative of dihydroartemisinin, is the most widely used member of
the family of artemisinin drugs. Artemisinin compounds are in
widespread use for the treatment of severe and complicated malaria
in humans. Art has been the most commonly used derivative for
>15 years, and according to a large number of clinicians,
intravenous administration of Art is the most effective treatment
for severe malaria (1–3). The effectiveness of Art in
vivo has been attributed to its rapid and extensive conversion
to dihydroartemisinin (DHA) (4).
Studies have reported that Art may have anti-cancer effects, as Art
inhibited the growth of cancer cells via cell cycle arrest and
induction of apoptosis (5,6).
Deregulated cell cycle progression is a hallmark of
cancer. Regulatory components of the cell cycle machinery often
become targets of genetic alterations during carcinogenesis.
Cyclin-dependent kinases (CDK) have central roles in promoting cell
cycle progression and uncontrolled activation of CDKs is the
driving force of cancer cell proliferation (7). Overexpression of CDKs and cyclins, as
well as downregulation of CDK inhibitors, is found in the majority
of human cancers, and components of the CDK system appear to be
functionally redundant. CDC25 genes are cell cycle-activating
phosphatases that remove the inhibitory phosphates of threonine and
tyrosine residues at the ATP-binding sites of CDK (8,9).
CDC25A is member of the CDC25 gene family. CDC25A regulates G1/S-
and G2/M-associated CDK activities (10,11).
In late G1, S and G2 phase, CDC25A protein has a short half-life as
a consequence of checkpoint kinase 1 (CHK1)-mediated
phosphorylation during a normal cell cycle. DNA damage or
inhibition of DNA replication activates CHK1 and CHK2 kinases that
then lead to rapid degradation of CDC25A, thereby preventing cell
cycle progression (12,13). Furthermore, concordant in
vitro and in vivo findings showed that CDC25A is
overexpressed in various types of human neoplastic malignancy
(14–17).
Cell apoptosis and cell growth are regulated through
complex signaling systems in the human body, and their disorder or
imbalance may induce the development of tumors. The efficiency of
chemotherapy drugs is often evaluated by their ability to induce
cell apoptosis. Mitochondrial membrane potential, B-cell lymphoma-2
(BCL-2), BCL-2-associated X protein (Bax) and caspase-3 regulate
cell apoptosis, where upregulation of bax and caspase-3, and
downregulation of mitochondrial membrane potential and bcl-2 may
result in cell apoptosis (18,19).
In the present study, the anti-cancer activity of
Art was examined by assessing its effect on the expression of
CDC25A, mitochondrial membrane potential, BCL-2, Bax and caspase-3
in Eca109 and Ec9706 human esophageal carcinoma cells as well as in
a nude mouse xenograft model.
Materials and methods
Chemicals and reagents
Art was purchased from Guiling Pharmaceutical Co.
(Guangxi, China). RT-PCR kit was obtained from Promega Corporation
(Madison, WI, USA). Mouse anti-human CDC25A (cat. no. sc-7389),
BCL-2 (cat. no. sc-7382), Bax (cat. no. sc-20067) and Caspase-3
(cat. no. sc-7272) monoclonal antibodies were purchased from Santa
Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). TRIzol was obtained
from Bio Basic Inc. (Markham, ON, Canada).
Cells and cell culture
The human esophageal cancer cell line Eca109 was
obtained from the Cancer Institute of the Fourth Hospital of Hebei
Medical University (Shijiazhuang, China). Human esophageal cancer
cells Ec9706 were obtained from the Molecular Oncology State Key
Laboratory, Cancer Institute and Hospital, Chinese Academy of
Medical Sciences (Beijing, China).
Cells were cultured in RPMI 1640 medium
(Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% fetal
bovine serum (Sigma-Aldrich), 100 units/ml penicillin (North China
Pharmaceutical Co., Ltd., Shijiazhuang, China) and 100 μg/ml
streptomycin (North China Pharmaceutical Co., Ltd., Shijiazhuang,
China) at 37°C in a humidified atmosphere of 5% CO2.
Animals
BALB/c nu/nu mice were purchased from The Institute
of Laboratory Animal Science, Chinese Academy of Medical Sciences
(Beijijng, China). The mice were maintained under specific
pathogen-free conditions at room temperature (23±1°C) and relative
humidity (40–60%), and were supplied with sterilized food and
water. The animals were maintained in 12 h light/dark cycles and
were provided access to food and water ad libitum. Animal
experiments were conducted according to the Institutional Animal
Care and Use Committee guidelines, and were approved by the ethics
committee of the Fourth Hospital of Hebei Medical University
(Shijiazhuang, China). 4–6 week-old female/male nu/nu mice weighing
18–20 g were inoculated with 200 μl Eca109 cells
(6×106 cells) in the left forelimb. This model has been
used for more than two decades to study the efficacy of
chemotherapeutic drugs against cancer. A total of 24 inoculated
Eca109-transplanted tumor mice were divided into four groups: The
negative control group was intraperitoneally injected physiological
saline, and treatment groups were injected with 100 mg/kg Art, 200
mg/kg Art or 3 mg/kg cisplatin. The mice were treated once a day
for 14 days. Following 14 days of treatment, the mice were
sacrified by cervical dislocation and tumor volume was measured at
the shortest and the longest diameters of the tumor using a vernier
caliper. The tumor volume was calculated using the following
standard formula: axb2/2, where a is the longest
diameter and b is the shortest diameter.
Flow cytometric analysis
Analysis of cell apoptosis
Cultured tumor cells (Eca109 and Ec9706 cells)
treated with various concentrations of Art (0, 30, 60 or 120
μmol/l) for 24 h were harvested using pancreatin
(Sigma-Aldrich). The cells were stained with propidium iodide (PI;
Sigma-Aldrich) and annexin V-fluorescein isothiocyanate (FITC;
Beckman Coulter, Miami, FL, USA), and analyzed using a Beckman
Coulter Epics-XL type flow cytometer (Beckman Coulter). Early
apoptotic cells were positive for annexin V and negative for PI
staining, whereas late apoptotic cells undergoing secondary
necrosis were positive for annexin V and PI staining.
Cell cycle analysis
Cultured tumor cells (Eca109 and Ec9706 cells;
1×106 cells) were harvested routinely following
treatment with various concentrations of Art (0, 30, 60 or 120
μmol/l) for 24 h. Cells were fixed in 70% (w/v) ice-cold
ethanol (Zeneca Co., Ltd., Handan, China). Following two washes
with ice-cold phosphate-buffered saline (PBS; Sigma-Aldrich), the
fixed cells were stained with 1 ml PI solution (50 μg/ml).
Following incubation for 30 min in the dark at 4°C and two washes
with ice-cold PBS, the stained cells were re-suspended in 1 ml PBS.
The stained cells were analyzed using a Beckman Coulter Epics-XL
type flow cytometer (Beckman Coulter).
Analysis of CDC25A, BCL-2, Bax and
caspase-3 protein
Cultured tumor cells (Eca109 and Ec9706 cells) were
harvested routinely following treatment with various concentrations
of Art (0, 30, 60 or 120 μmol/l) for 24 h. Cells were fixed
overnight with 70% ice-cold ethanol. Following two washes with
ice-cold PBS, the fixed cells were re-suspended in 1 ml PBS
containing CDC25A, BCL-2, Bax and caspase-3 antibody (1:100),
respectively, following incubation for 30 min in the dark at room
temperature. Following two washes with PBS, cells were re-suspended
in 1 ml PBS containing antibody II (immunoglobulin G-FITC; Jackson
ImmunoResearch Laboratories, Inc., West Grove, PA, USA) with
incubation for 30 min in the dark at room temperature. Following
two washes with PBS, cells were re-suspended in 1 ml PBS. The
stained cells were analyzed using a Beckman Coulter Epics-XL type
flow cytometer (Beckman Coulter).
Analysis of mitochondrial membrane
potential in Eca109 and Ec9706 cells
Cultured tumor cells (Eca109 and Ec9706 cells) were
harvested routinely following treatment with various concentrations
of Art (0, 30, 60 or 120 μmol/l) for 24 h. Following two
washes with ice-cold PBS, the cells were stained using 1 ml
rhodamine 123 solution (10 μg/ml; Sigma-Aldrich). Following
incubation for 30 min in the dark at 37°C and two washes with
ice-cold PBS, the stained cells were re-suspended in 1 ml PBS. The
stained cells were analyzed using a Beckman Coulter Epics-XL type
flow cytometer (Beckman Coulter).
Statistical analysis
Values are expressed as the mean ± standard
deviation and were statistically analyzed using multiple analysis
of variance followed by the Newman-Keuls method of post-hoc
comparison (SPSS software; SPSS, Inc., Chicago, IL, USA). P<0.05
was considered to indicate a significant difference.
Results
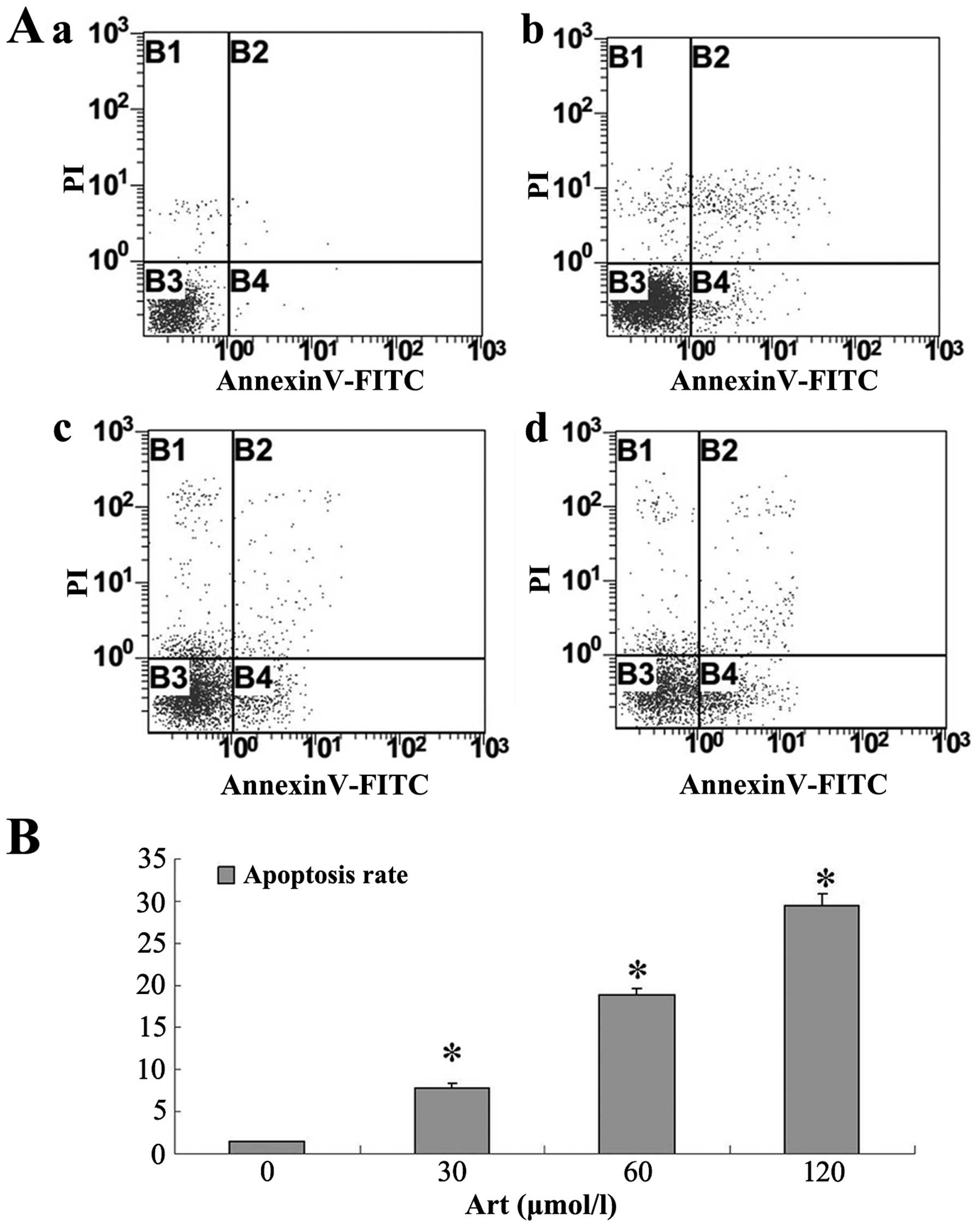
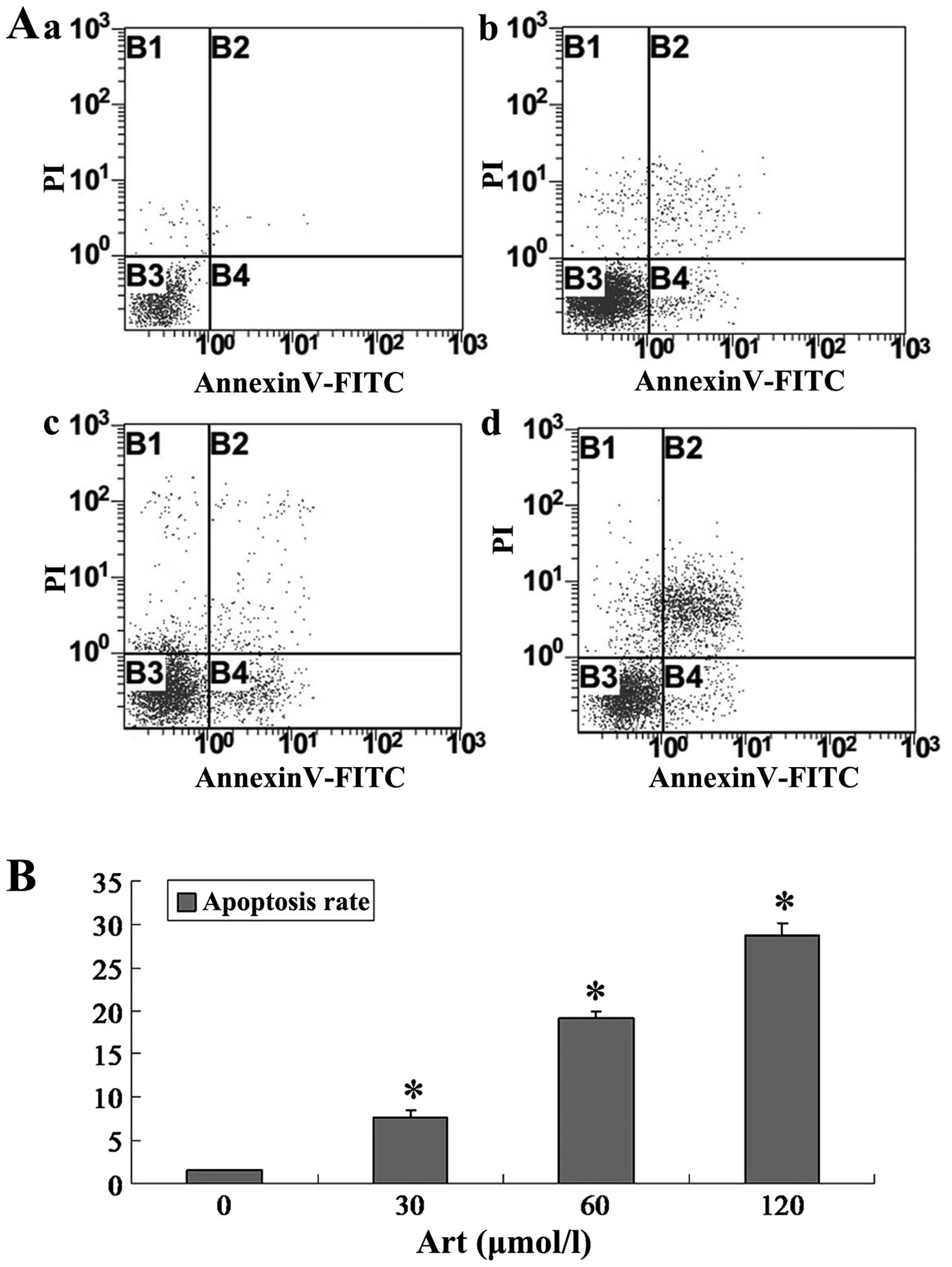
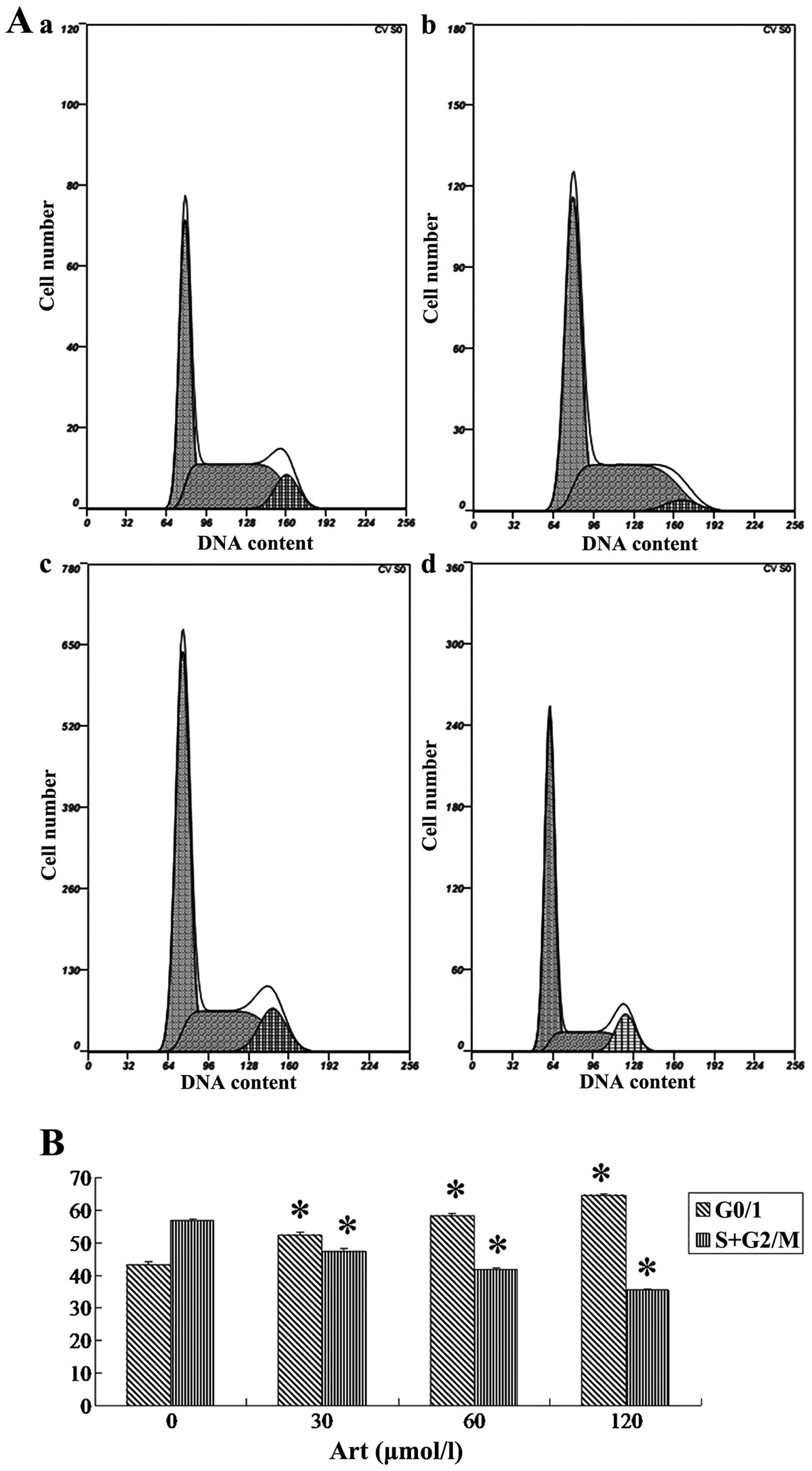
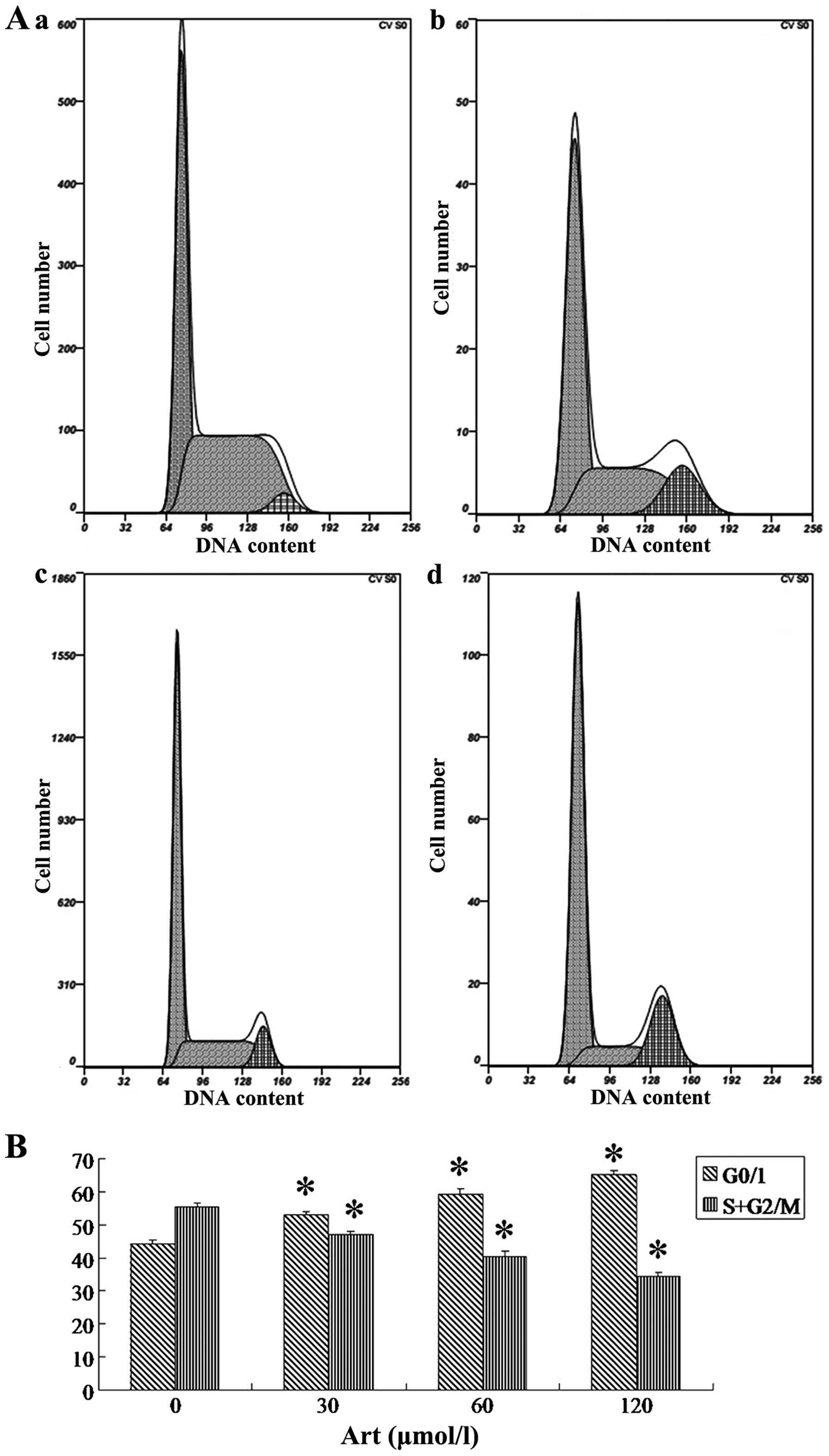
Art induces apoptosis and cell cycle
arrest in human esophageal cancer cells
Following Art exposure, Eca109 and Ec9706 cells
underwent apoptosis and cell cycle arrest in a dose-dependent
manner. In the range of 30–120 μmol/l, Art induced
apoptosis, as determined by Annexin V/PI staining (Figs. 1 and 2). The percentages of apoptotic Eca109
and Ec9706 cells increased with increasing Art concentration
(Figs. 1 and 2). Treatment of Eca109 and Ec9706 cells
with Art resulted in pronounced G0/1 phase arrest in a
dose-dependent manner (Figs. 3 and
4). The results for Ec9706 cells
were consistent with those for Eca109 cells.
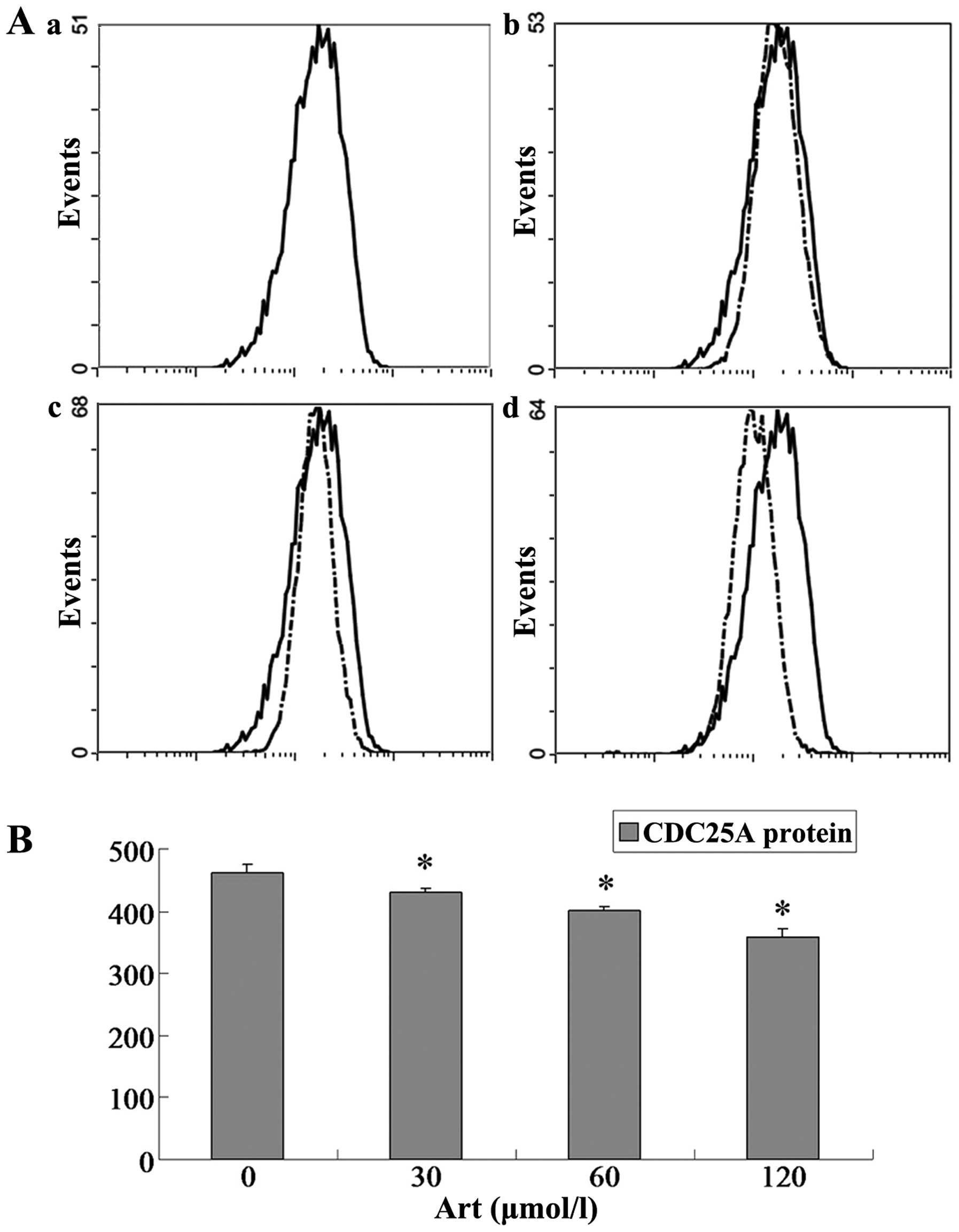
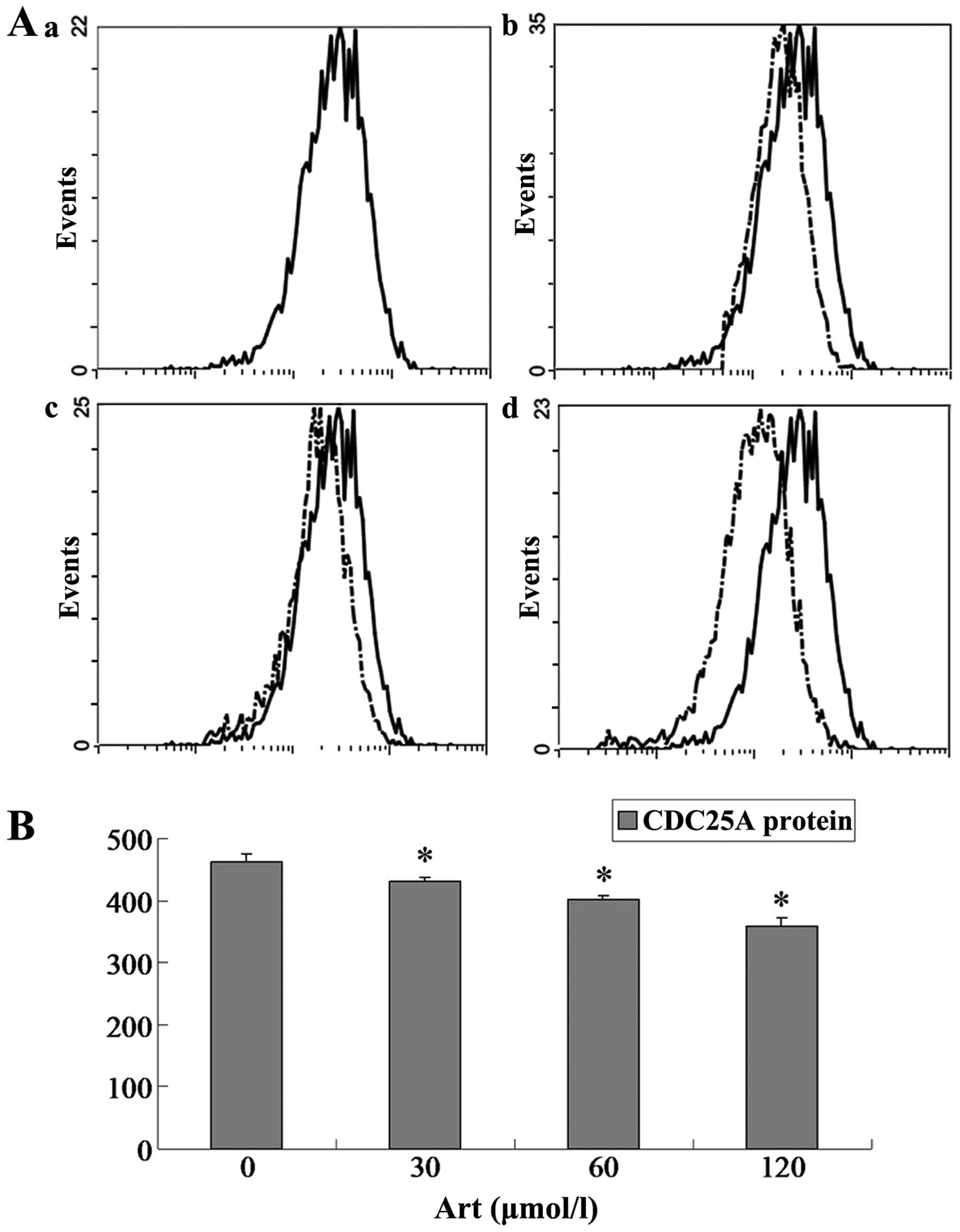
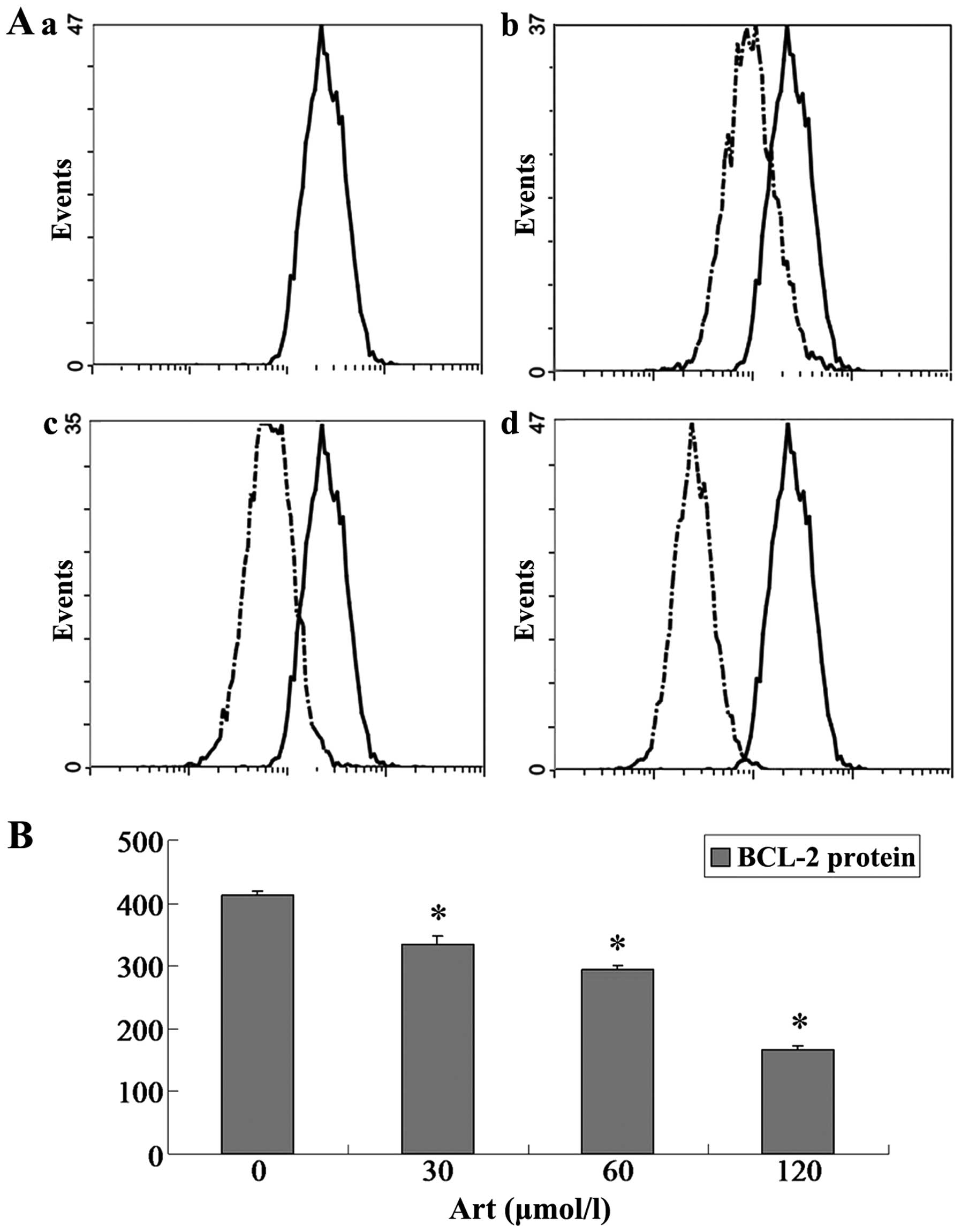
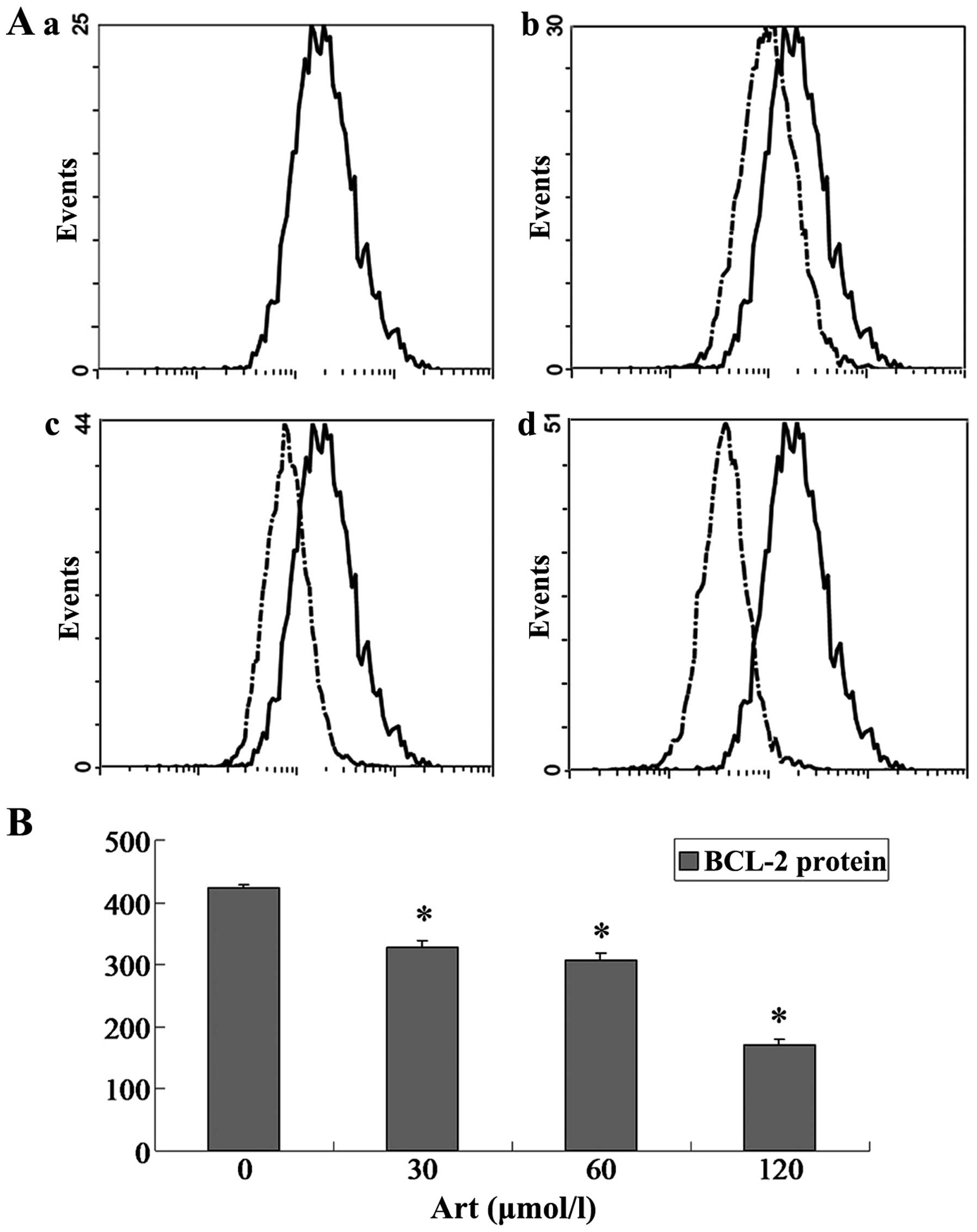
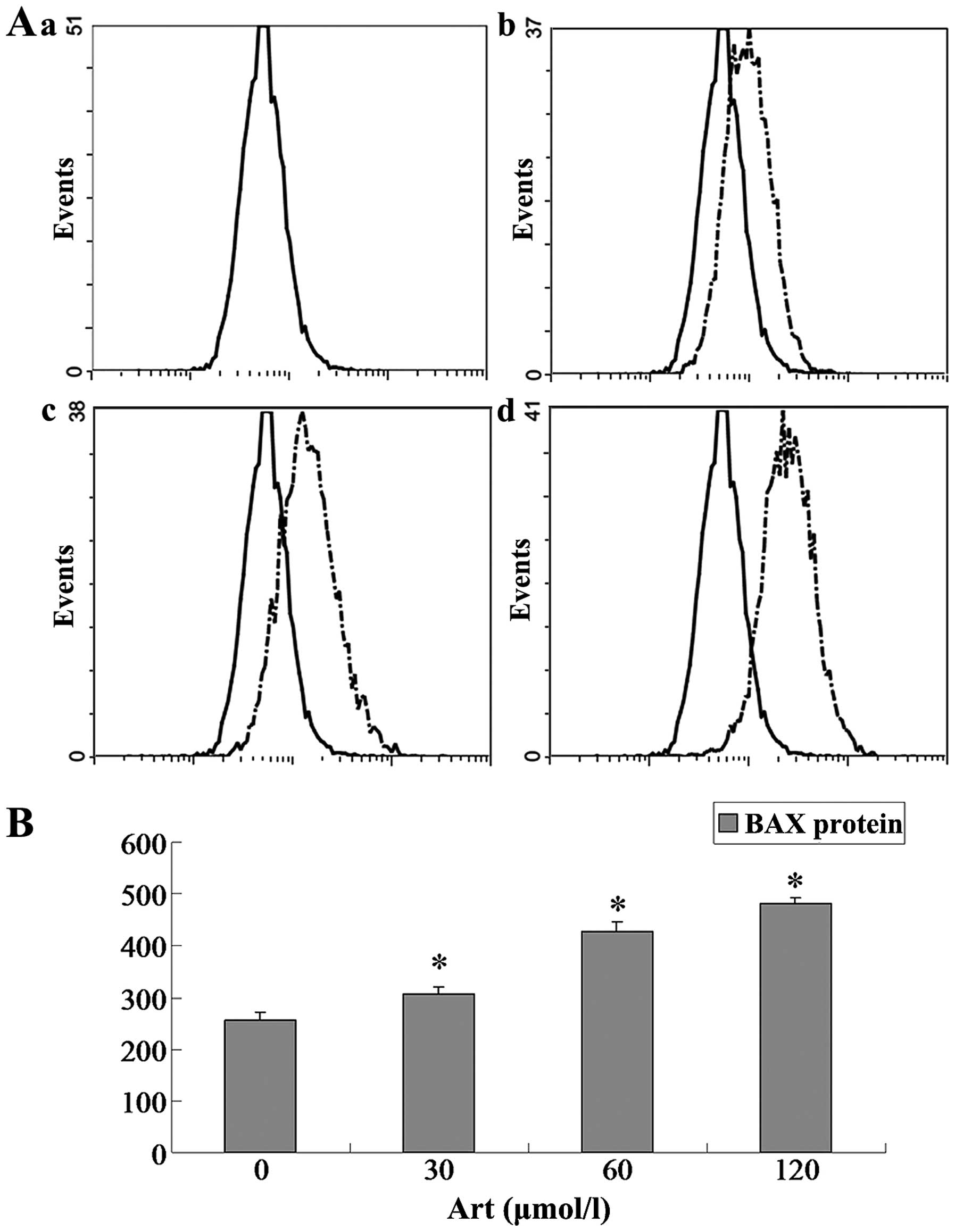
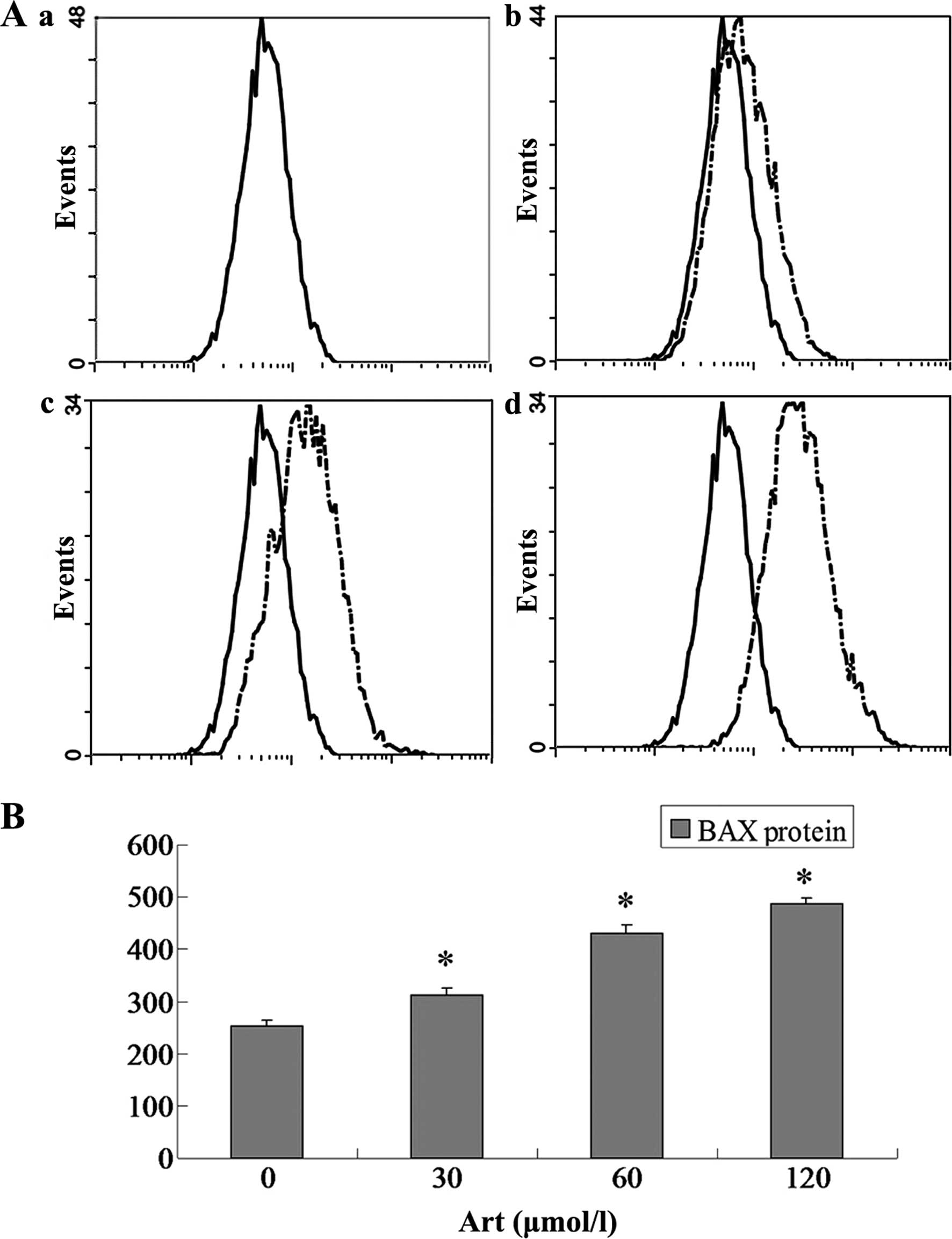
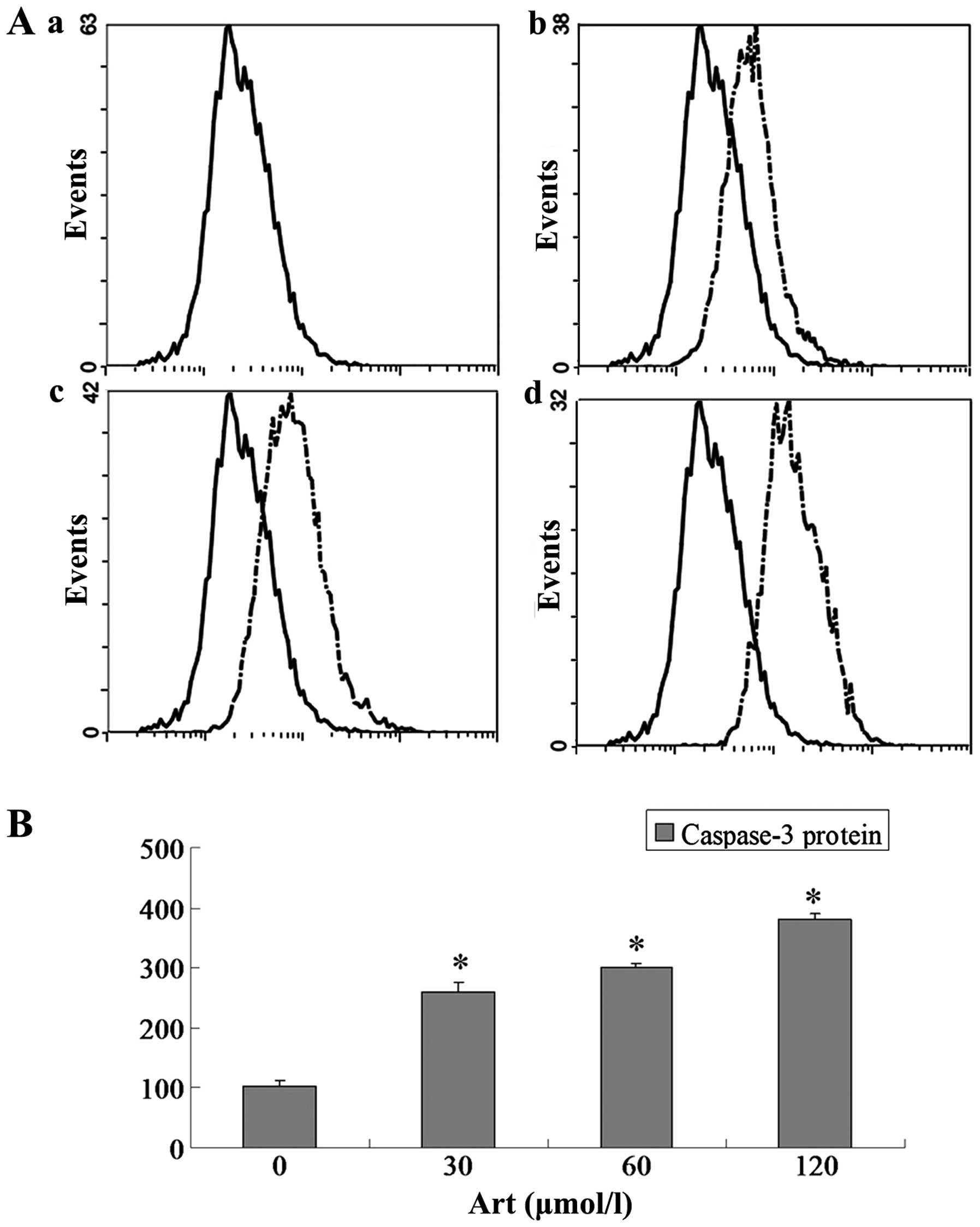
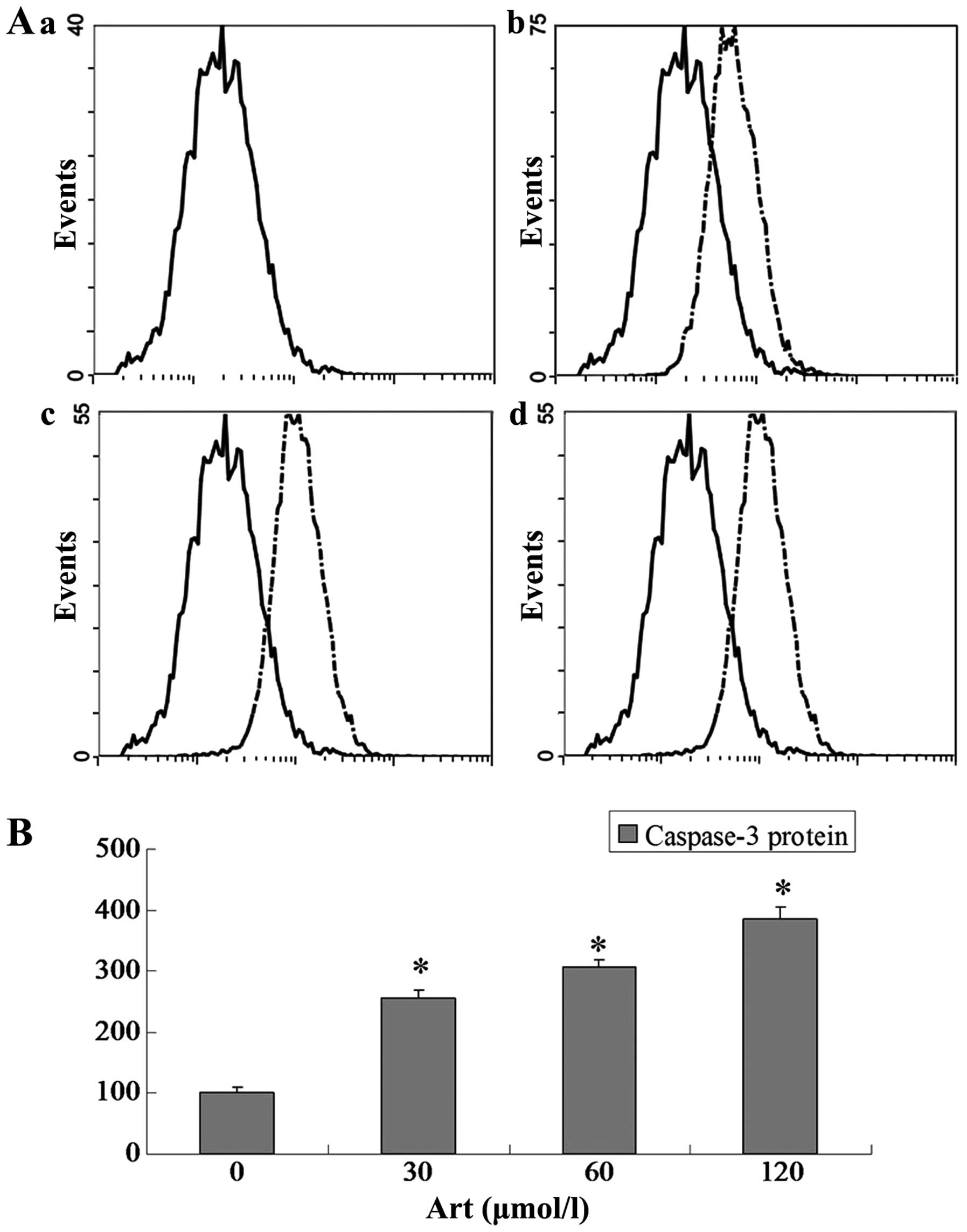
Effects of Art on CDC25A, BCL-2, Bax and
caspase-3 expression levels in Eca109 and Ec9706 cells
Eca109 and Ec9706 cells were treated with 0, 30, 60
and 120 μmol/l Art for 24 h, washed with cold PBS, and
analyzed for protein expression of CDC25A, BCL-2, Bax and caspase-3
using flow cytometry. The protein expression of CDC25A and BCL-2
was significantly downregulated compared with that of the controls
(P<0.05) when the Eca109 and Ec9706 cells were treated for 24 h
with various concentrations of Art (30–120 μmol/l) (Figs. 5Figure 6Figure 7–8). By contrast, the Bax and caspase-3
protein expression was significantly upregulated compared with that
in the control cells (P<0.05) following treatment with various
concentrations of Art for 24 h (Figs.
9Figure 10Figure 11–12).
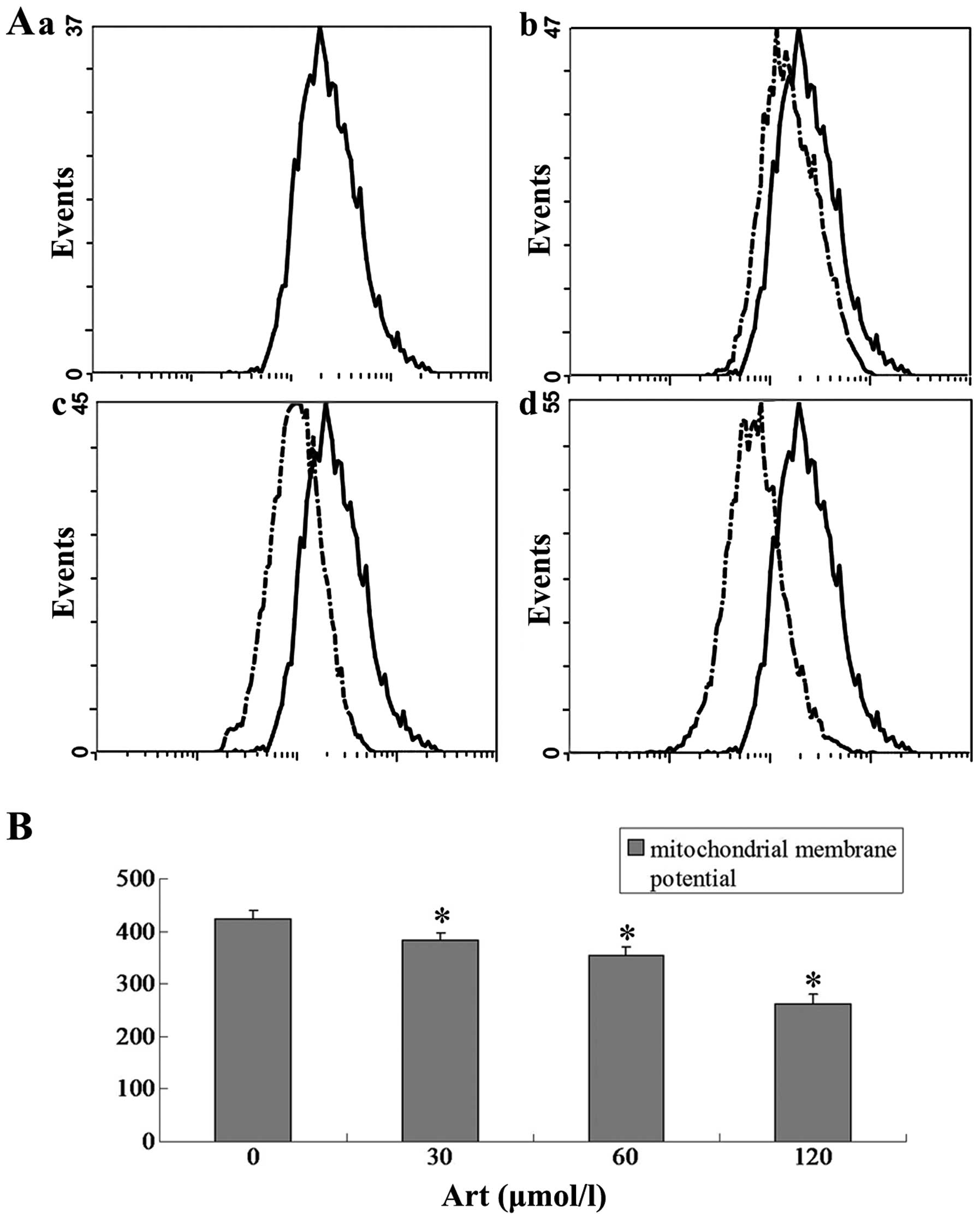
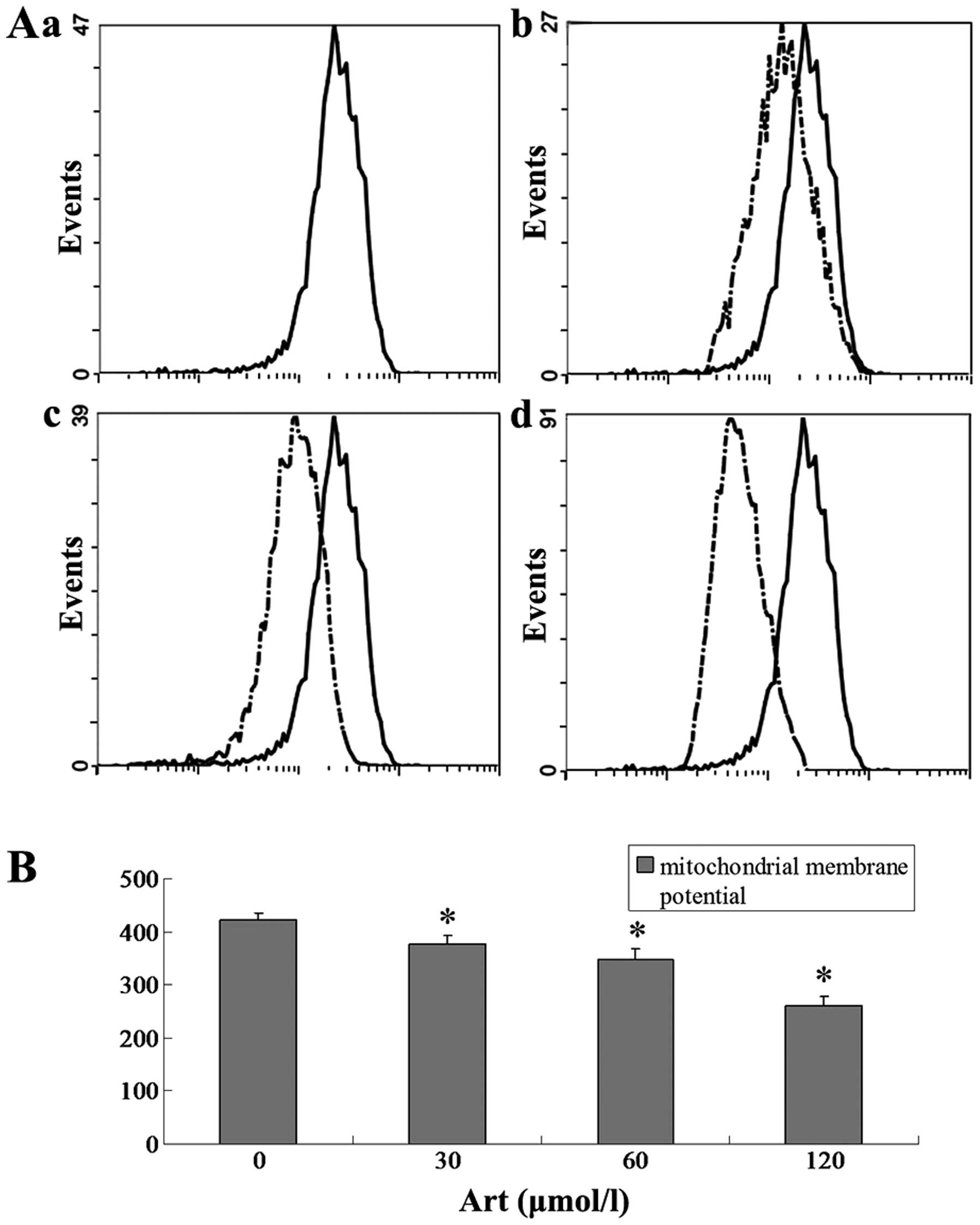
Effect of Art on the mitochondrial
membrane potential in Eca109 and Ec9706 cells
Eca109 and Ec9706 cells were treated with 0, 30, 60
and 120 μmol/l Art for 24 h, washed with cold PBS and
analyzed for the mitochondrial membrane potential using flow
cytometry. The mitochondrial membrane potential in the Art group
was significantly lower than that in the control group (P<0.01).
The mitochondrial membrane potential in the 120 μmol/ml Art
group was significantly lower than that in the 30 μmol/ml
and 60 μmol/ml Art group (P<0.01) (Figs. 13 and 14).
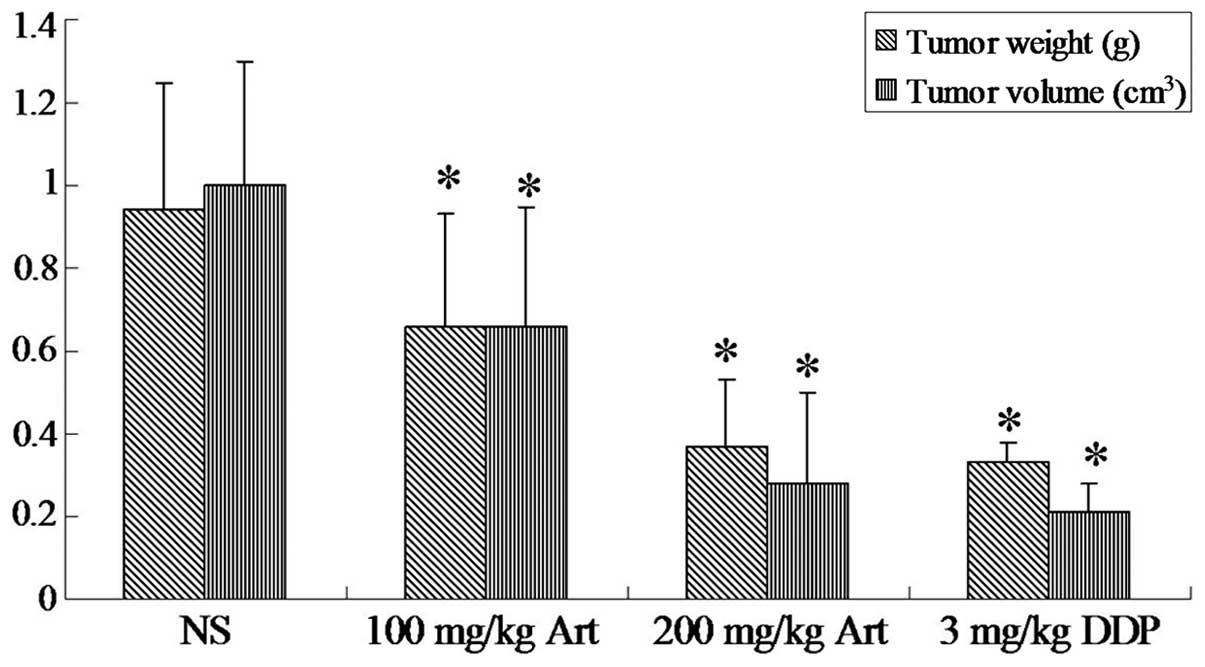
Inhibitory effect of Art on the growth of
xenograft tumors
On the basis of the in vitro results, the
present study examined the effects of Art on the growth of Eca109
cells in mouse xenograft models. An Eca109-transplanted tumor nude
mouse model was established successfully. As shown in Fig. 15, Art inhibited the growth of
Eca109-transplanted tumors in nude mice. The volume and weight of
transplanted tumors in the Art and cisplatin groups were lower than
those in the normal saline-injected control group (P<0.05).
Discussion
Cancer therapy is often based on surgery,
chemotherapy and radiation therapy. Chemotherapy is one of the main
therapies for the majority of cancers, particularly for those with
a proneness to invade adjacent tissues and to metastasize to other
organs. The effectiveness of chemotherapy is often limited by
toxicity to other normal tissues in the body, thereby resulting in
chemotherapy failure. Esophageal carcinoma is one of the most
common malignant gastrointestinal tumors (20). The incidence of esophageal
carcinoma is high in China and it is a great threat to human
health. As chemotherapy with normal anti-cancer drugs is far from
providing satisfactory clinical outcomes for patients with
esophageal cancer, more efficient drugs and drug combinations are
urgently required. Artemisins are a class of compounds that are
first-line treatment options for malaria (21,22),
and its water-soluble derivative, Art is a remarkable anti-malarial
agent, particularly in severe and drug-resistant cases (23,24).
Recently, Art has also been demonstrated to exert profound
anti-cancer activity (25).
Efferth et al (5) reported
a profound cytotoxic action of Art against cancer cell lines of
different tumor types. However, a detailed understanding of the
molecular mechanisms during Art-induced cell death in cancer cells
is limited. The present study reported that Art exerted potent
cytotoxic effects on the human esophageal cancer Eca109 and Ec9706
cell lines in vitro and in vivo. The cytotoxicity of
Art was mediated by apoptosis and cell cycle arrest, which was
supported by assessment of apoptosis, cell cycle analysis and
determination of expression levels of apoptosis- and cell
cycle-associated proteins. Apoptosis has important roles in
maintaining cell homeostasis and dysfunction of apoptotic
signalling may cause serious conditions such as cancer.
At present, apoptosis has been the most studied
mechanism in anti-cancer therapy (26,27).
Cell apoptosis is a programmed and gene-controlled cell death and
involves complex regulatory mechanisms. Loss of the mitochondrial
transmembrane potential is an indicator and inducer of apoptosis
(26). In the present study, the
apoptosis-inducing effect of Art on Eca109 and Ec9706 cells through
the mitochondrial pathway was studied in vitro. Treatment
with Art induced apoptosis in Eca109 and Ec9706 cells in a
dose-dependent manner, which was detected by annexin V/PI staining.
In addition, the in-vivo study showed that Art exerted a
significant growth inhibitory effect on xenograft tumors in nude
mice. Hamacher-Brady et al (6) showed that Art was able to induce
mitochondrial apoptosis in the breast cancer cell line MCF-7, which
was consistent with the results of the present study. On the other
hand, it has been reported that the anti-tumor mechanism of Art was
not caused by apoptosis. Zhou et al (28) showed that Art suppressed the
proliferation of gastric cancer via oncosis rather than apoptosis.
The results of the present study demonstrated that in esophageal
cancer cells, Art exerted multifarious effects on proteins involved
in apoptosis signaling. Downstream of BCL-2 protein expression and
upstream of Bax protein expression, mitochondrial apoptotic cell
death was activated. The BCL-2 and Bax proteins are members of the
BCL-2 family, which consists of pro-apoptotic and anti-apoptotic
proteins that exert opposing effects on mitochondria (29). Increased expression of the
anti-apoptotic protein BCL-2 is involved in the development and
progression of numerous tumor types. BCL-2 localizes to cellular
membranes, particularly in mitochondria, where it stabilizes the
transmembrane potential and reduces membrane permeability. BCL-2
appears to contribute to tumor cell survival by enhancing the rate
of cell proliferation and by allowing tumor cells to escape
destruction by effector cells of the immune system, whereas Bax can
promote the release of cytochrome C from mitochondria into the
cytosol to enhance apoptosis. In the present study, it was found
that the BCL-2 protein expression of esophageal cancer cells was
downregulated following treatment with various concentrations of
Art in a dose-dependent manner, whereas Bax protein expression was
upregulated. Furthermore, the mitochondrial membrane potential of
esophageal cancer cells was downregulated by Art in a
dose-dependent manner. Furthermore, caspase-3 protein expression
levels were upregulated following treatment with Art in a
dose-dependent manner. In the present study, Art induced esophageal
cancer cell apoptosis by modulating the BCL-2 and Bax expression
levels, downregulating the mitochondrial membrane potential and
upregulating caspase-3 expression levels. At the same time, it was
found that Art caused G0/G1 phase arrest of esophageal cancer
cells. The cell-cycle protein CDC25A activates CDK2, promoting
entry into S-phase of the cell cycle (30,31).
Overexpression of CDC25A, which has been previously shown to have
oncogenic potential (32),
eliminates this checkpoint. According to the results of the present
study, Art reduced CDC25A expression in esophageal cancer cells in
a dose-dependent manner in vivo and in vitro. The
results of the present study revealed that CDC25A was a molecular
target of Art, which suggested that the Art may impair G1/S
transition, thereby contributing to the prevention of uncontrolled
cell growth.
In conclusion, the present study revealed that Art
inhibited the growth of esophageal cancer cells by inducing
apoptosis and cell cycle arrest. Art is a clinically approved drug
for the treatment of malaria and showed low side effects in nude
mice. Future studies will further explore the potential clinical
application of Art as a chemotherapeutic drug for esophageal
cancer.
Acknowledgments
This study was supported by a grant from The Hebei
Province Medical Scientific Research Key Project (no.
ZL20140126).
References
|
1
|
He SP, Tan GY, Li G, et al: Development of
a sensitive monoclonal antibody-based enzyme-linked immunosorbent
assay for the antimalaria active ingredient artemisinin in the
Chinese herb Artemisia annua L. Anal Bioanal Chem. 393:1297–1303.
2009. View Article : Google Scholar
|
|
2
|
Tanaka H, Putalun W, De-Eknamkul W,
Matangkasombut O and Shoyama Y: Preparation of a novel monoclonal
antibody against the antimalarial drugs, artemisinin and Art.
Planta Med. 73:1127–1132. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Harris P, Price S, Senthuran S,
Cochupanachimootil J and Norton R: Automated erythrocytapheresis
for severe falciparum malaria. Intern Med J. 41:60–63. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
McLean WG and Ward SA: In vitro
neurotoxicity of artemisinin derivatives. Med Trop (Mars). 58(Suppl
3): 28–31. 1998.
|
|
5
|
Efferth T, Dunstan H, Sauerbrey A, Miyachi
H and Chitambar CR: The anti-malarial Art is also active against
cancer. Int J Oncol. 18:767–773. 2001.PubMed/NCBI
|
|
6
|
Hamacher-Brady A, Stein HA, Turschner S,
et al: Art activates mitochondrial apoptosis in breast cancer cells
via iron-catalyzed lysosomal reactive oxygen species production. J
Biol Chem. 286:6587–6601. 2011. View Article : Google Scholar :
|
|
7
|
Blomberg I and Hoffmann I: Ectopic
expression of Cdc25A accelerates the G (1)/S transition and leads
to premature activation of cyclin E- and cyclin A-dependent
kinases. Mol Cell Biol. 19:6183–6194. 1999.PubMed/NCBI
|
|
8
|
Chen MS, Ryan CE and Piwnica-Worms H: Chk1
kinase negatively regulates mitotic function of Cdc25A phosphatase
through 14-3-3 binding. Mol Cell Biol. 23:7488–7497. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Galaktionov K, Lee AK, Eckstein J, et al:
CDC25 phosphatases as potential human oncogenes. Science.
269:1575–1577. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mailand N, Podtelejnikov AV, Groth A, Mann
M, Bartek J and Lukas J: Regulation of G (2)/M events by Cdc25A
through phosphorylation-dependent modulation of its stability. EMBO
J. 21:5911–5920. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Falck J, Mailand N, Syljuasen RG, Bartek J
and Lukas J: The ATM-Chk2-Cdc25A checkpoint pathway guards against
radioresistant DNA synthesis. Nature. 410:842–847. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mailand N, Falck J, Lukas C, et al: Rapid
destruction of human Cdc25A in response to DNA damage. Science.
288:1425–1429. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Molinari M, Mercurio C, Dominguez J,
Goubin F and Draetta GF: Human Cdc25 A inactivation in response to
S phase inhibition and its role in preventing premature mitosis.
EMBO Rep. 1:71–79. 2000. View Article : Google Scholar
|
|
14
|
Gasparotto D, Maestro R, Piccinin S, et
al: Overexpression of CDC25A and CDC25B in head and neck cancers.
Cancer Res. 57:2366–2368. 1997.PubMed/NCBI
|
|
15
|
Dixon D, Moyana T and King MJ: Elevated
expression of the cdc25A protein phosphatase in colon cancer. Exp
Cell Res. 240:236–243. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nishioka K, Doki Y, Shiozaki H, et al:
Clinical significance of CDC25A and CDC25B expression in squamous
cell carcinomas of the oesophagus. Br J Cancer. 85:412–421. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu W, Fan YH, Kemp BL, Walsh G and Mao L:
Overexpression of cdc25A and cdc25B is frequent in primary
non-small cell lung cancer but is not associated with
overexpression of c-myc. Cancer Res. 58:4082–4085. 1998.PubMed/NCBI
|
|
18
|
Siddiqui WA, Ahad A and Ahsan H: The
mystery of BCL2 family: Bcl-2 proteins and apoptosis: An update.
Arch Toxicol. 89:289–317. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shi LS, Wang H, Wang F, et al: Effects of
gastrokine-2 expression on gastric cancer cell apoptosis by
activation of extrinsic apoptotic pathways. Mol Med Rep.
10:2898–2904. 2014.PubMed/NCBI
|
|
20
|
Gamliel Z: Incidence, epidemiology and
etiology of esophageal cancer. Chest Surg Clin N Am. 10:441–450.
2000.PubMed/NCBI
|
|
21
|
Danis M and Jauréguiberry S: The
artemisinin derivatives must be in France the first-line treatment
of all P. falciparum malaria cases simple or severe. La Prat.
63:896–898. 2013.In French.
|
|
22
|
Kumnuan R, Pattaradilokrat S,
Chumpolbanchorn K, et al: In vivo transmission blocking activities
of Art on the avian malaria parasite Plasmodium gallinaceum. Vet
Parasitol. 197:447–454. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shanks GD: For severe malaria, Art is the
answer. Lancet. 376:1621–1622. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Okebe J and Eisenhut M: Pre-referral
rectal Art for severe malaria. Cochrane Database Syst Rev.
5:CD0099642014.
|
|
25
|
Steinbruck L, Pereira G and Efferth T:
Effects of Art on cytokinesis and G (2)/M cell cycle progression of
tumour cells and budding yeast. Cancer Genomics Proteomics.
7:337–346. 2010.
|
|
26
|
Yang H, Tian ST, Wu RY, et al:
Glycoborinine induces apoptosis through mitochondrial pathway in
HepG2 cells. J Asian Nat Prod Res. 16:991–999. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wimardhani YS, Suniarti DF, Freisleben HJ,
Wanandi SI, Siregar NC and Ikeda MA: Chitosan exerts anticancer
activity through induction of apoptosis and cell cycle arrest in
oral cancer cells. J Oral Sci. 56:119–126. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhou X, Sun WJ, Wang WM, et al: Art
inhibits the growth of gastric cancer cells through the mechanism
of promoting oncosis both in vitro and in vivo. Anticancer Drugs.
24:920–927. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Oh KJ, Barbuto S, Pitter K, Morash J,
Walensky LD and Korsmeyer SJ: A membrane-targeted BID BCL-2
homology 3 peptide is sufficient for high potency activation of BAX
in vitro. J Biol Chem. 281:36999–37008. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Viry E, Anwar A, Kirsch G, Jacob C,
Diederich M and Bagrel D: Antiproliferative effect of natural
tetrasulfides in human breast cancer cells is mediated through the
inhibition of the cell division cycle 25 phosphatases. Int J Oncol.
38:1103–1111. 2011.PubMed/NCBI
|
|
31
|
Young LM and Pagano M: Cdc25 phosphatases:
differential regulation by ubiquitin-mediated proteolysis. Cell
Cycle. 9:4613–4614. 2010. View Article : Google Scholar
|
|
32
|
Sengupta S, Jana S and Bhattacharyya A:
TGF-β-Smad2 dependent activation of CDC 25A plays an important role
in cell proliferation through NFAT activation in metastatic breast
cancer cells. Cell Signal. 26:240–252. 2014. View Article : Google Scholar
|