Introduction
Hochu-ekki-to (HET) (bu-zhong-yi-qi-tang in Chinese)
is a Kampō herbal medicine. Kampō medicines are traditional
remedies composed of several herbs, and have been used for several
hundred years in Japan (1). To
date, the Japanese Ministry of Health, Labour and Welfare has
approved >120 Kampō prescriptions for clinical use (2). These Kampō medicines have been
considered to be clinically effective since antiquity in Japan.
However, studies on the biochemical and pharmacological mechanisms
of action of Kampō medicines are not extensive.
HET is mainly used for the treatment of appetite
loss (3), general fatigue
(4) and loss of vigor (1). It is particularly used for treating
elderly people and, more recently, patients with infectious and
malignant diseases (5,6), or after chemotherapy or radiation
therapy of malignant tumors (7,8). It
has been reported that HET activates macrophages (9) and natural killer cells (10) and restores impaired immune
function, which is beneficial for the prevention of cancer
(11,12). In studies on mice, HET suppressed
the development and metastasis of several malignant tumor types,
including biliary carcinoma and uterine cancer (3,13).
In an in vitro study, inhibition of viability measured by
MTT assay was observed when Hep3B hepatocellular carcinoma cells
were incubated with HET (14). In
addition, flow cytometric analysis showed that HET induced cell
cycle arrest and apoptosis in Hep3B cells (14). These results suggested that the
underlying mechanisms of the anti-tumor activity of HET involve the
suppression of cell viability and induction of apoptosis.
However, high concentrations of HET in culture
medium may have non-specific cytotoxic effects owing to components
including saponins (e.g. saikosaponin in HET) and other
detergent-like compounds. When cells are incubated with >5,000
µg/ml, necrosis has been identified to be markedly increased
in ovarian cancer cells by flow cytometric analysis (15). To rule out the non-specific
cytotoxic effects, the present study used a low dose of HET (50
µg/ml) that did not suppress cell viability, and assessed
the synergistic effect of HET pre-treatment on the potency of
cisplatin against HeLa cells. Cell survival was measured by colony
survival and crystal violet assays, and the apoptotic rate was
analyzed by flow cytometry. Proteins associated with cell viability
and apoptosis, including phosphorylated Akt (p-Akt), p53, B-cell
lymphoma 2 (Bcl-2), Bcl-2-associated X protein (Bax) and active
caspase-3 were analyzed by immunoblotting.
Materials and methods
Reagents and drugs
HET was obtained from Tsumura Co. (Tokyo, Japan).
HET is a mixture of spray-dried powder from hot water extracts
obtained from the following ten medicinal plants: Ginseng radix
(16.7%), Atractylodis rhizoma (16.7%), Astragali radix (16.7%),
Angelicae radix (12.5%), Ziziphi fructus (8.3%), Bupleuri radix
(8.3%), Glycyrrhizae radix (6.3%), Zingiberis rhizoma (2.0%),
Cimicifuga rhizoma (4.2%) and Aurantii nobilis pericarpium (8.3%).
HET powder was dissolved at 10 mg/ml in culture medium at 37°C for
10 min and thoroughly mixed. The solution was passed through a
0.22-µm filter to sterilize and remove any insoluble
components. Cisplatin was purchased from Sigma-Aldrich (St. Louis,
MO, USA).
Cells and culture conditions
The human cervical cancer cell line HeLa, obtained
as previously described (16) was
used in the present study. The cells were cultured in Eagle's
Minimum Essential Medium (EMEM; Nissui, Tokyo, Japan) supplemented
with 10% calf serum (Gibco-BRL, Invitrogen Life Technologies,
Carlsbad, CA, USA). Cells were grown at 37°C in a humidified
atmosphere containing 5% CO2.
Cell survival assay
The colony survival assay was used to determine the
dose of HET that did not affect cell viability (17). Briefly, HeLa cells were plated in
100-mm dishes (1×103 cells/dish) in medium containing
HET at various concentrations (0, 5, 25, 50, 250 and 500
µg/ml). After culturing for 14 days, the colonies were
stained with 0.2% methylene blue (Wako Pure Chemical Industries,
Ltd., Osaka, Japan) in 30% methanol (Wako Pure Chemical Industries,
Ltd.), and visible colonies with a diameter greater than 3 mm
(containing greater than 50 cells) were counted by eye.
The sensitivity of cells to cisplatin was measured
by colony survival and crystal violet assays (18). For the colony survival assay, cells
were plated in 100-mm dishes (1×103 cells/dish) in
medium with or without 50 µg/ml HET. After culturing for 24
h, the medium was changed to serum-free medium containing the
indicated concentrations of cisplatin (0, 0.31, 0.63, 1.3, 2.5 or
5.0 µM). After incubation in the cisplatin-containing medium
for 2 h, the HET-pre-treated cells (cells treated with cisplatin
following HET pre-treatment) or HET-untreated cells (cells treated
with cisplatin without HET pre-treatment) were cultured in fresh
medium for 14 days followed by colony counting.
For the crystal violet assay, cells were plated in
60-mm dishes (5×105 cells/dish) in medium with or
without 50 µg/ml HET and incubated for 24 h. Cells were
harvested, plated onto 96-well plates (4×103 cells/well)
containing cisplatin at the indicated concentrations (0, 0.39,
0.78, 1.6 or 3.1 µM), and cultured for 2 days. After
culture, viable cells were stained with crystal violet (Wako Pure
Chemical Industries, Ltd.) followed by measurement of absorbance at
595 nm. Cell viability was calculated from the absorbance using an
Emax Precision Microplate reader (Molecular Devices, LLC,
Sunnyvale, CA, USA) and expressed as the percentage of surviving
cells with regard to the group treated without cisplatin.
Flow cytometric analysis of the apoptotic
cell population in sub-G1 phase
Cells that were pre-treated with and without HET (50
µg/ml) for 24 h were then treated with cisplatin (0, 15 and
30 µM) at the indicated concentrations for 24 h. The
HET-pre-treated or -untreated cells were harvested and the cell
population in sub-G1 phase (apoptotic fraction) was
analyzed with Guava Cell Cycle Reagent (EMD Millipore, Billerica,
CA, USA) and an Accuri C6 cytometer (Tomy Digital Biology,
Encyclopedia Circle, Fremont, CA, USA). The data were analyzed with
FlowJo software, version 7.6 (FlowJo, LLC, Ashland, OT, Canada) as
described previously (19).
Immunoblotting
Apoptosis-associated molecules were analyzed by
immunoblotting (19). Cells were
cultured in medium in the presence or absence of 50 µg/ml
HET for 24 h and then treated with cisplatin at indicated
concentrations for 24 h. Next, whole-cell lysates in an SDS
sampling buffer (62.5 mM Tris-Cl, pH 6.8, 2.3% SDS, 10% glycerol,
5% 2-mercaptoethanol, 0.01% bromophenol blue) were prepared from
the HET-pre-treated or -untreated cells. Briefly, cells were
harvested with phosphate-buffered saline, cell number was counted,
then whole cells were dissolved directly with the SDS sampling
buffer. The whole-cell lysates from approximately 2×104
cells were applied to each lane of SDS-PAGE. Electrophoresis and
blotting were performed with constant 15 mA and 250 mA current
using a Tris-glycine buffer system, for 2 h and 3 h, respectively.
All reagents for the western blotting were obtained from Wako Pure
Chemical Industries, Ltd. The active form of caspase-3 protein was
detected using rabbit anti-cleaved caspase-3 antibody (5A1E; Cell
Signaling Technology, Inc., Beverly, MA, USA; 1:500 dilution).
Bcl-2, Bax, p-Akt and p53 proteins were detected using mouse
anti-Bcl-2 monoclonal antibody (sc-7832; Santa Cruz Biotechnology,
Santa Cruz, CA, USA; 1:1,000 dilution), mouse anti-Bax monoclonal
antibody (sc-7480; Santa Cruz Biotechnology; 1:1,000 dilution),
rabbit anti-p-Akt antibody (193H12; Cell Signaling Technology,
Inc.; 1:1,000 dilution), and mouse anti-p53 monoclonal antibody
(sc-126; Santa Cruz Biotechnology; 1:1,000 dilution), respectively.
The secondary antibodies used were as follows: Horseradish
peroxidase (HRP)-linked anti-rabbit antibody (NA934V; GE Healthcare
Life Sciences, Chalfont, UK; 1:4,000 dilution) and HRP-linked
anti-mouse antibody (NA931V; GE Healthcare Life Sciences; 1:4,000
dilution). The protein signals were visualized by the Enhanced
Chemiluminescence system reaction (GE Healthcare Life Sciences).
Protein levels of actin were also analyzed using mouse anti-actin
antibody (C4; ICN Biomedicals, Costa Mesa, CA, USA; 1:10,000
dilution) as a loading control. The intensities of the protein
signals were quantified using Multi Gauge 2.2 image analysis
software (Fuji Foto Film, Tokyo, Japan) and expressed as a relative
value to that of actin. Immunoblotting analysis was performed three
times using cell samples prepared twice independently.
Statistical analysis
P-values were calculated to compare the
HET-pre-treated cells and the HET-untreated cells by Student's
t-test with Microsoft Excel 2010 software (Microsoft Corporation,
Redmond, WA, USA). Values are expressed as the mean ± standard
deviation. P<0.05 was considered to indicate a statistically
significant difference between values.
Results
HET sensitizes HeLa cells to
cisplatin-induced cytotoxicity
The present study first examined the effect of HET
alone on HeLa cell viability. No growth suppression by HET was
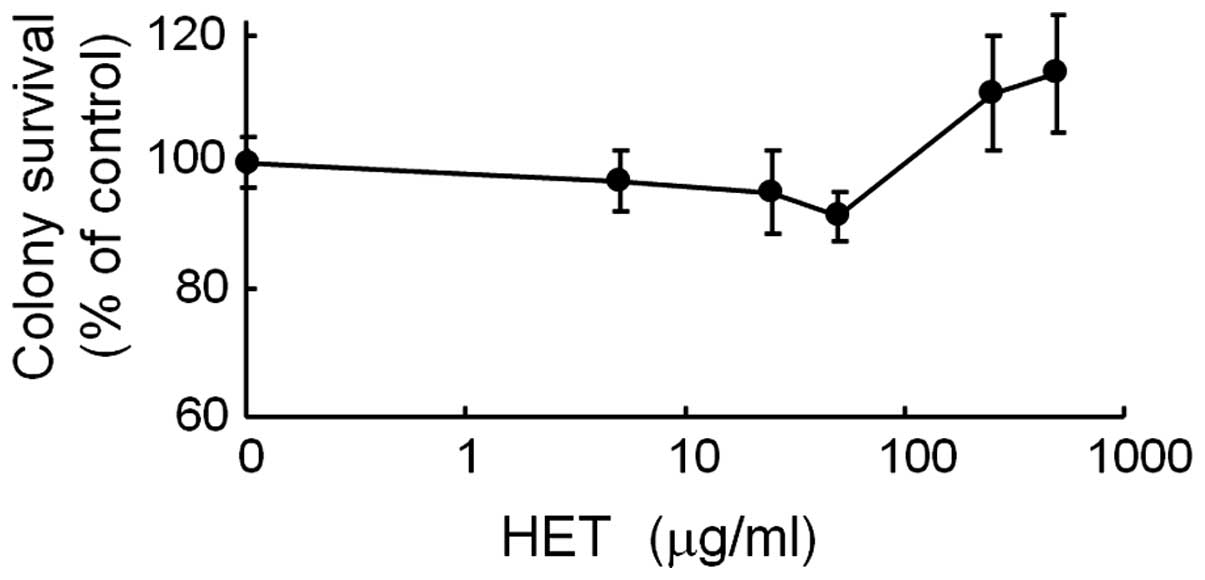
observed at concentrations of up to 500 µg/ml (Fig. 1).
The present study therefore adopted 50 µg/ml
HET to exclude any cytotoxic effects. Next, the sensitivity of HeLa
cells to cisplatin after pre-treatment in medium containing 50
µg/ml of HET for 24 h was assessed. The colony survival
assay indicated that the HET-pre-treated cells had a significantly
higher sensitivity to cisplatin-induced cell death than the
HET-untreated cells (Fig. 2A).
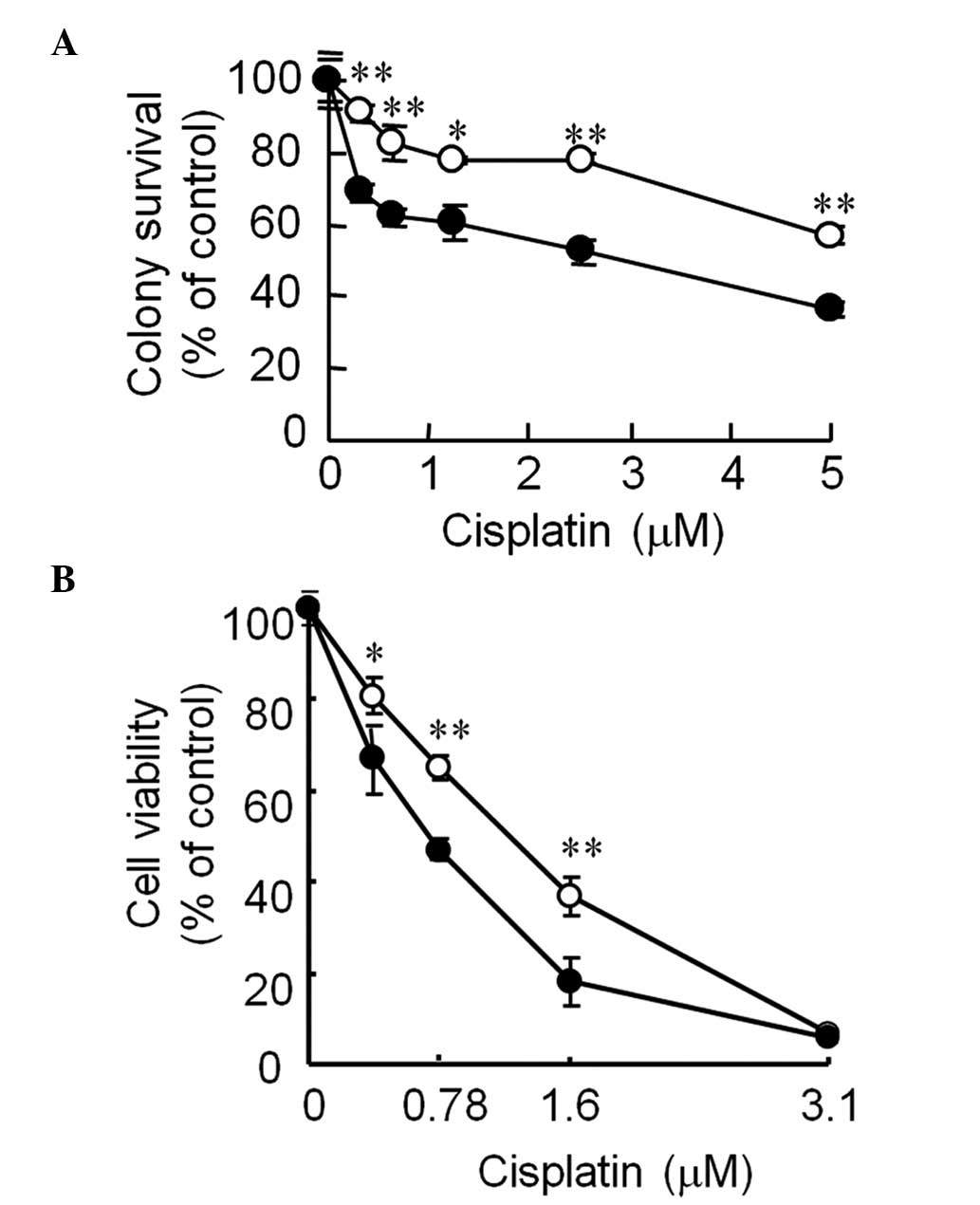 | Figure 2Effect of pre-treatment with HET on
the sensitivity of HeLa cells to cisplatin-induced cell death. (A)
Colony survival assay. Cells were cultured in the medium with or
without 50 µg/ml HET for 24 h and then incubated with 0,
0.31, 0.63, 1.3, 2.5 or 5.0 µM cisplatin in serum-free
medium for 2 h. After incubation, the cells were cultured with
fresh medium for 14 days followed by colony counting. (B) Crystal
violet assay. Cells were cultured in medium with or without 50
µg/ml of HET for 24 h, harvested, plated onto 96-well
plates, and incubated with cisplatin at 0, 0.39, 0.78, 1.6 or 3.1
µM for 2 days. Viable cells were stained with crystal violet
and cell viability was assessed. Values are expressed as the mean ±
standard deviation. *P<0.05; **P<0.005,
for HET-pre-treated cells vs. HET-untreated cells. HET,
Hochu-ekki-to. |
In addition, the crystal violet assay showed that
HET-pre-treated HeLa cells were more sensitive to the inhibitory
effect of cisplatin on cell viability than HET-untreated HeLa cells
(Fig. 2B). These findings
suggested that the anti-cancer effect of cisplatin was enhanced by
pre-treatment with HET.
HET- and cisplatin-induced cell death
proceeds via a caspase-dependent apoptotic pathway
A previous study by our group reported that
cisplatin induced apoptosis in HeLa cells and other cancer cell
lines (20). Therefore, the
present study examined whether the cell death induced by cisplatin
and HET is induced via apoptotic pathways. Flow cytometric analysis
indicated a distinct increase in the population of cells in the
sub-G1 phase (apoptotic fraction) after cisplatin
treatment in the cells pre-treated with HET compared with that in
the HET-untreated cells (Fig. 3).
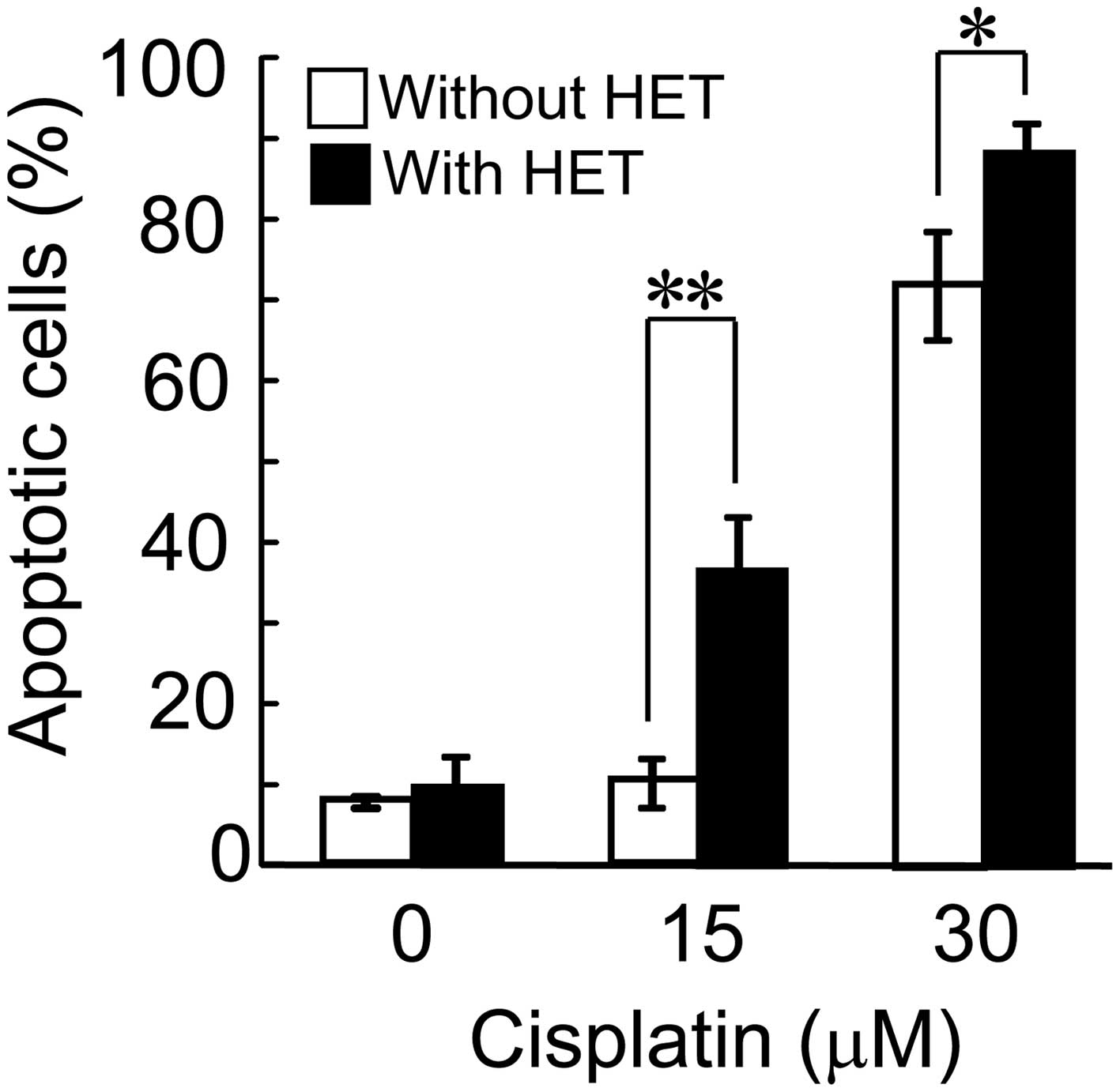
While the sub-G1 population in the HET-untreated HeLa
cells was 7.8±0.71, 10.3±3.0 and 72.2±6.7% after treatment with 0,
15 and 30 µM cisplatin, respectively, the sub-G1
fractions were increased to 9.8±3.6, 36.8±6.4 (P<0.005) and
88.6±4.3% (P<0.05), respectively, in the HET-pre-cultured HeLa
cells.
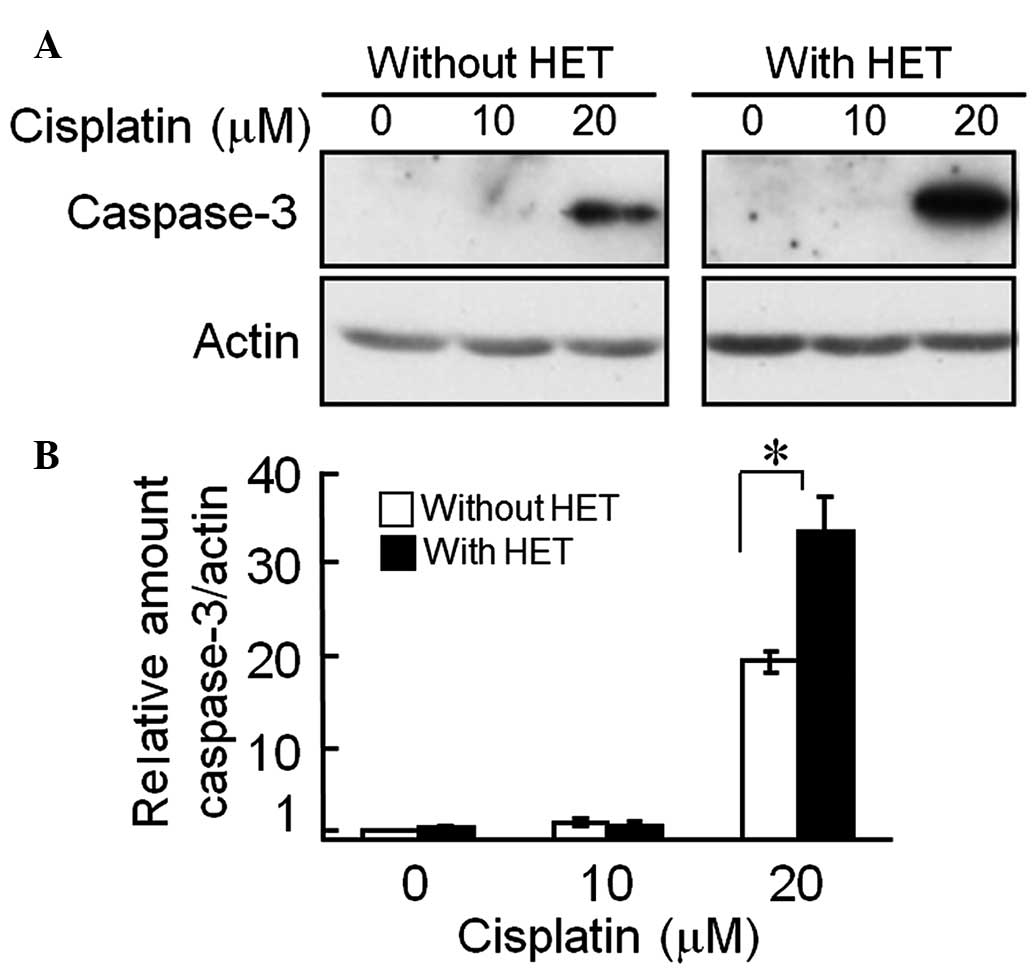
Caspase-3 is one of the effector caspases, which is
cleaved and activated by initiator caspases, and executes apoptosis
when activated (21). The levels
of active caspase-3 increased by ~1.7-fold after incubation of
HET-pre-treated HeLa cells with 20 µM cisplatin, compared
with those in the HET-untreated cells (Fig. 4). These results supported that
HET-pre-treatment enhanced cisplatin-induced apoptosis, which
proceeded via a caspase-dependent pathway.
HET-pre-treatment enhances apoptosis
signaling in HeLa cells treated with cisplatin
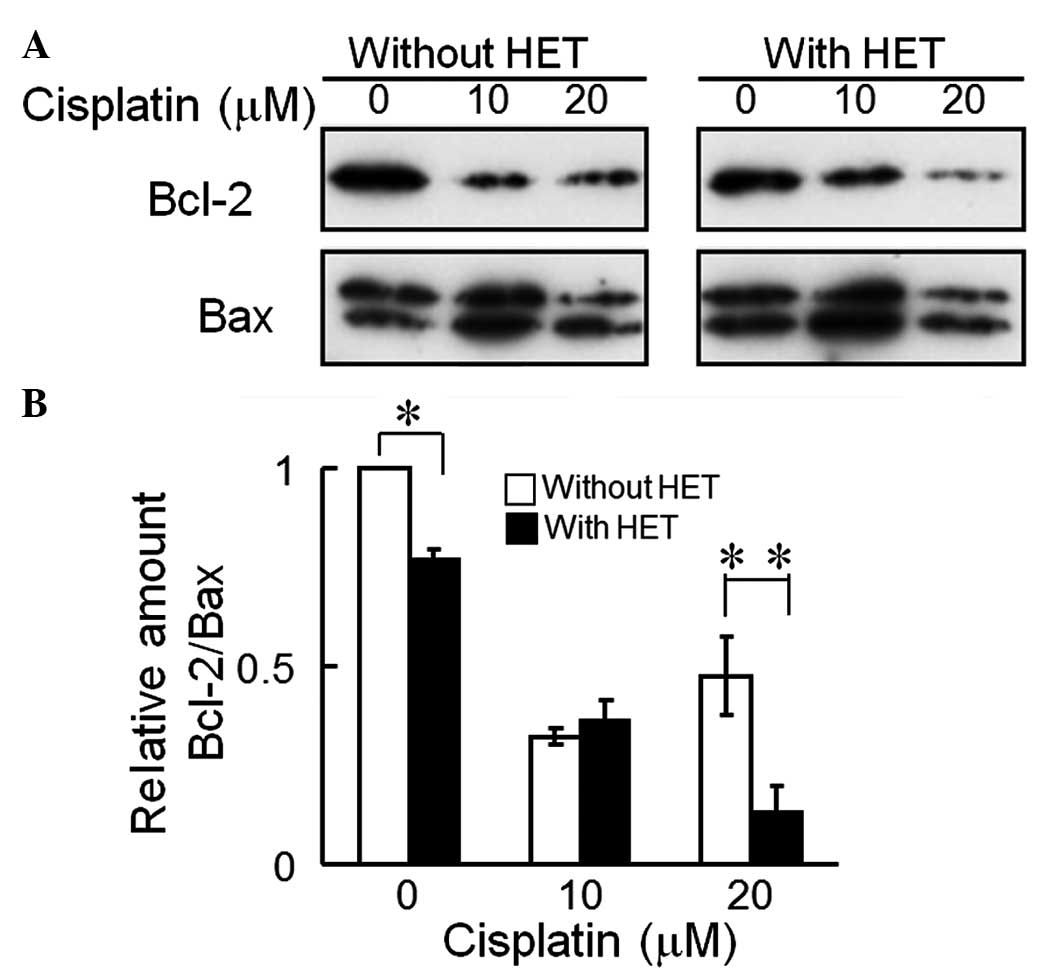
Bcl-2 is an anti-apoptotic protein, and Bax is a
pro-apoptotic protein (22). The
relative expression ratios of anti-apoptotic proteins to
pro-apoptotic proteins have been reported to correlate with
cellular sensitivity to the lethal effects of anti-cancer drugs
(23). The Bcl-2-to-Bax ratio at 0
and 20 µM cisplatin in the HET-pre-cultured HeLa cells was
decreased by 23 and 68%, respectively, compared with that in the
HET-untreated cells (Fig. 5).
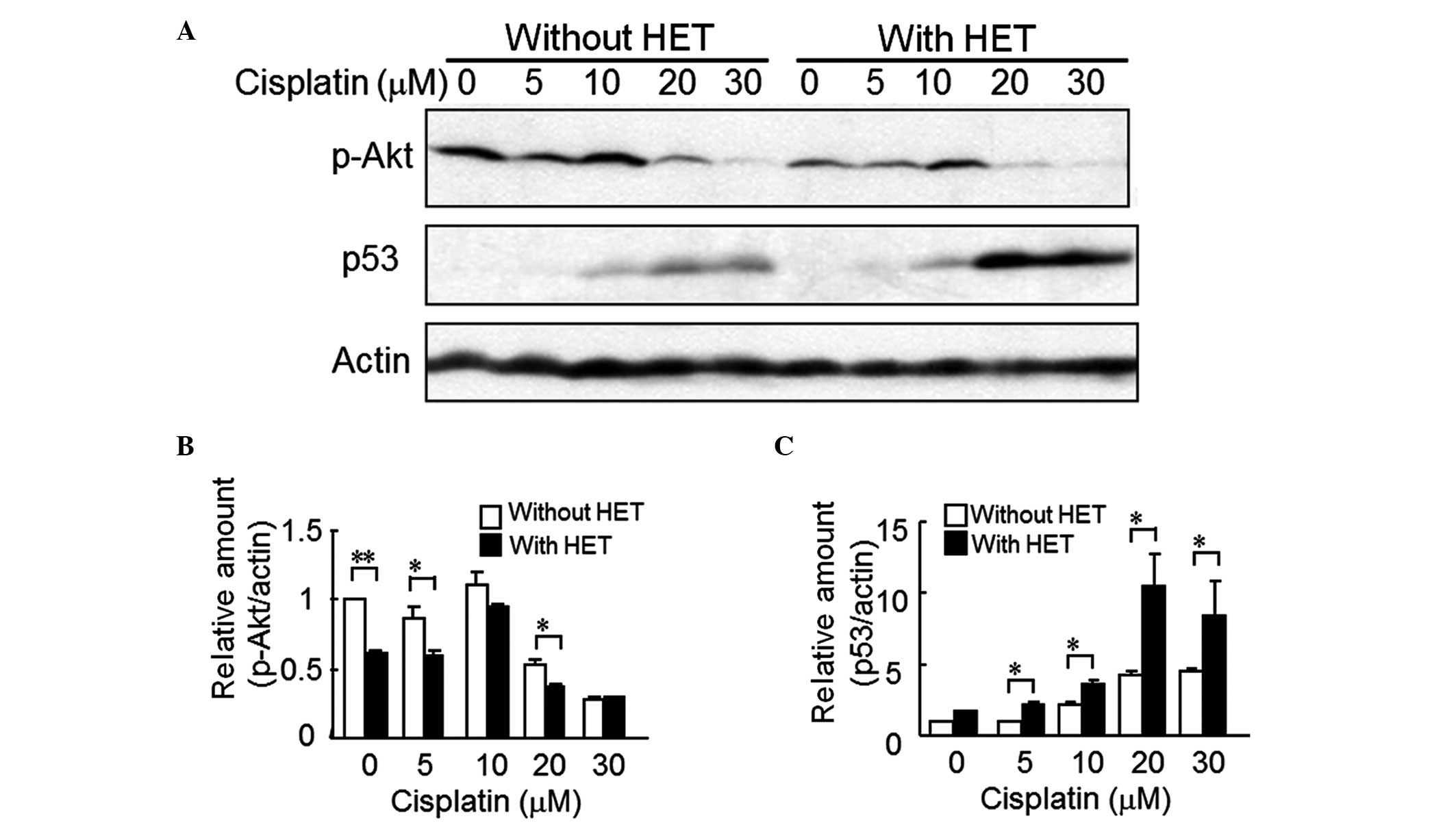
Next, the present study determined the cellular
levels of p-Akt and p53 as candidates for upstream molecules to
regulate the enhancement of the cisplatin-induced apoptosis by HET.
Akt is negatively regulated by p53 (24) when apoptosis occurs. Fig. 6 shows the effect of HET on the
protein levels of p-Akt and p53 in HeLa cells incubated with
various concentrations of cisplatin. In the HET-untreated cells as
well as in HET-treated cells, p-Akt expression was decreased at
cisplatin concentra tions of >20 µM, compared with that
in control cells without cisplatin treatment. In addition, at
almost all of the tested concentrations of cisplatin (0, 5, 10 and
20 µM), the p-Akt levels in the HET-pre-treated cells were
lower than those in the HET-untreated cells. The difference between
the two groups was significant at cisplatin concentrations other
than 10 and 30 µM (Fig. 6A and
B). Conversely, the protein levels of p53 were increased at
cisplatin concentrations of >10 µM compared with those in
the cisplatin-untreated groups. Of note, p53 levels were markedly
elevated in the HET-pre-treated cells as compared with those in the
HET-untreated cells (Fig. 6A and
C).
Discussion
Zhu et al (15) reported a necrotic effect of HET at
concentrations greater than 5000 μg/ml in ovarian cancer
cells. In the present study, no growth suppression was observed at
concentrations up to 100 µg/ml, while the colony survival
appeared to increase marginally (no significant difference) at
>100 µg/ml (Fig. 1).
Thus in the current study, 50 µg/ml HET was selected for
use. The findings of the present study suggested that HET enhanced
the anti-cancer effects of cisplatin by stimulation of
cisplatin-induced apoptosis signaling, including upregulation of
caspase-3 activation and downregulation of the Bcl-2 to Bax ratio.
Furthermore, cellular levels of p-Akt were decreased, while p53 was
increased in cells pre-treated with HET, leading to the enhancement
of the cisplatin-induced apoptosis.
In several cancer cell lines, the proliferation is
enhanced through constitutive activation of the phosphoinositide 3
kinase (PI3K)/Akt pathway through growth factor receptors including
epidermal growth factor receptor (25) and platelet-derived growth factor
receptor (26) as well as
oncogenes including Ras and Her2/Neu (27,28).
The results of the present study showed that p-Akt levels were
decreased, which was in parallel with decreases in the Bcl-2 to Bax
ratio (Fig. 5) in the
HET-pre-treated HeLa cells compared with those in the HET-untreated
cells, even without cisplatin treatment. Therefore, a mild
suppression of PI3K/Akt signaling by HET at the low concentration
at which it was used in the present study (50 µg/ml) may be
involved in its enhancement of anti-cancer drug-induced apoptosis,
even though HET alone did not suppress cell viability.
When p-Akt (activated Akt) is outnumbered by
counteracting p53, apoptosis is activated (29). Interplay between Akt and p53
determines the cell fate with regard to proliferation or apoptosis
(24). Under stress conditions,
such as the presence of cytotoxic cisplatin, Akt is negatively
regulated by p53 (24).
Furthermore, inactivation of Akt leads to suppression of Bcl-2
(30), and upregulation of p53
also suppresses the expression of Bcl-2 (31). Therefore, the downregulation of Akt
and the upregulation of p53 (Fig.
6) may have contributed to the apoptosis-enhancing effect of
HET.
This apoptosis-enhancing effect of HET was not
observed in pancreatic cancer cell lines, including AsPC-1 and
MiaPaCa-2, which carry a p53 mutation (32,33).
Another study reported that the hepatoma cell line HepG2 (wild-type
p53) was more sensitive to growth suppression induced by HET than
HA22T (p53 mutation) (14). HeLa
cells carry wild-type p53 (34).
These findings suggested that the apoptosis-inducing effect of HET
may be dependent on the p53 status, as wild-type p53 is required
for its efficiency. In this respect, further studies are required
to elucidate the underlying upstream mechanisms.
In conclusion, the present study was the first, to
the best of our knowledge, to demonstrate that pre-treatment with
HET at a low dose, which did not suppress cell viability, enhanced
cisplatin-induced apoptosis in HeLa cells, as indicated by a
distinct increase in the population of cells in the
sub-G1 phase by flow cytometry, downregulation of p-Akt
and the Bcl-2 to Bax ratio, as well as stimulation of
cisplatin-induced up-regulation of p53 and active caspase-3. The
HET-involved augmenting effect on cisplatin-induced apoptosis may
therefore be beneficial for enhancing the efficiency of anti-cancer
drug treatment.
Acknowledgments
This study was supported in part by grants-in-aid
from the following organizations: The Smoking Research Foundation
and the Japan Society for the Promotion of Science (no.
24510065).
Abbreviations:
References
|
1
|
Tohda M, Hayashi H, Sukma M and Tanaka K:
BNIP-3: A novel candidate for an intrinsic depression-related
factor found in NG108-15 cells treated with Hochu-ekki-to, a
traditional oriental medicine, or typical antidepressants. Neurosci
Res. 62:1–8. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
de Caires S and Steenkamp V: Use of
Yokukansan (TJ-54) in the treatment of neurological disorders: A
review. Phytother Res. 24:1265–1270. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Onogi K, Niwa K, Tang L, Yun W, Mori H and
Tamaya T: Inhibitory effects of Hochu-ekki-to on endometrial
carcinogenesis induced by N-methyl-N-nitrosourea and
17beta-estradiol in mice. Oncol Rep. 16:1343–1348. 2006.PubMed/NCBI
|
|
4
|
Jeong JS, Ryu BH, Kim JS, Park JW, Choi WC
and Yoon SW: Bojungikki-tang for cancer-related fatigue: A pilot
randomized clinical trial. Integr Cancer Ther. 9:331–338. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Satoh H, shikawa H, Ohtsuka M and Sekizawa
K: Japanese herbal medicine in patients with advanced lung cancer:
Prolongation of survival. J Altern Complement Med. 8:107–108. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tatsumi K, Shinozuka N, Nakayama K, Sekiya
N, Kuriyama T and Fukuchi Y: Hochuekkito improves systemic
inflammation and nutritional status in elderly patients with
chronic obstructive pulmonary disease. J Am Geriatr Soc.
57:169–170. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kaneko M, Kawakita T, Kumazawa Y, Takimoto
H, Nomoto K and Yoshikawa T: Accelerated recovery from
cyclophosphamide-induced leukopenia in mice administered a Japanese
ethical herbal drug, Hochu-ekki-to. Immunopharmacology. 44:223–231.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kim SH, Lee SE, Oh H, Kim SR, Yee ST, Yu
YB, Byun MW and Jo SK: The radioprotective effects of
bu-zhong-yi-qi-tang: A prescription of traditional Chinese
medicine. Am J Chin Med. 30:127–137. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nakayama M, Sugiyama Y, Yamasawa H, Soda
M, Mato N, Hosono T and Bando M: Effect of hochuekkito on alveolar
macrophage inflammatory responses in hyperglycemic mice.
Inflammation. 35:1294–1301. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Harada M, Seta K, Ito O, Tamada K, Li T,
Terao H, Takenoyama M, Kimura G and Nomoto K: Concomitant immunity
against tumor development is enhanced by the oral administration of
a kampo medicine, Hochu-ekki-to (TJ-41: Bu-Zhong-Yi-Qi-Tang).
Immunopharmacol Immunotoxicol. 17:687–703. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kuroiwa A, Liou S, Yan H, Eshita A, Naitoh
S and Nagayama A: Effect of a traditional Japanese herbal medicine,
hochu-ekki-to (Bu-Zhong-Yi-Qi Tang), on immunity in elderly
persons. Int Immunopharmacol. 4:317–324. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Utsuyama M, Seidlar H, Kitagawa M and
Hirokawa K: Immunological restoration and anti-tumor effect by
Japanese herbal medicine in aged mice. Mech Ageing Dev.
122:341–352. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tsuneoka N, Tajima Y, Kitasato A, Fukuda
K, Kitajima T, Adachi T, Mishima T, Kuroki T, Onizuka S and
Kanematsu T: Chemopreventative effect of hochu-ekki-to (TJ-41) on
chemically induced biliary carcinogenesis in hamsters. J Surg Res.
151:22–27. 2009. View Article : Google Scholar
|
|
14
|
Kao ST, Yeh CC, Hsieh CC, Yang MD, Lee MR,
Liu HS and Lin JG: The Chinese medicine Bu-Zhong-Yi-Qi-Tang
inhibited proliferation of hepatoma cell lines by inducing
apoptosis via G0/G1 arrest. Life Sci. 69:1485–1496. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhu K, Fukasawa I, Furuno M, Inaba F,
Yamazaki T, Kamemori T, Kousaka N, Ota Y, Hayashi M, Maehama T and
Inaba N: Inhibitory effects of herbal drugs on the growth of human
ovarian cancer cell lines through the induction of apoptosis.
Gynecol Oncol. 97:405–409. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Suzuki T, Lu J, Hu G, Kita K and Suzuki N:
Retrovirus-mediated transduction of a short hairpin RNA gene for
GRP78 fails to downregulate GRP78 expression but leads to cisplatin
sensiti-zation in HeLa cells. Oncol Rep. 25:879–885. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wano C, Kita K, Takahashi S, Sugaya S,
Hino M, Hosoya H and Suzuki N: Protective role of HSP27 against
UVC-induced cell death in human cells. Exp Cell Res. 298:584–592.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Saotome K, Morita H and Umeda M:
Cytotoxicity test with simplified crystal violet staining method
using microtitre plates and its application to injection drugs.
Toxicol In Vitro. 3:317–321. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kita K, Sugita K, Chen SP, Suzuki T,
Sugaya S, Tanaka T, Jin YH, Satoh T, Tong XB and Suzuki N:
Extracellular recombinant annexin II confers UVC-radiation
resistance and increases the Bcl-xL to Bax protein ratios in human
UVC-radiation-sensitive cells. Radiat Res. 176:732–742. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sato T, Kita K, Sugaya S, Suzuki T and
Suzuki N: Extracellular release of annexin II from pancreatic
cancer cells and resistance to anticancer drug-induced apoptosis by
supplementation of recombinant annexin II. Pancreas. 41:1247–1254.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang ZB, Liu YQ and Cui YF: Pathways to
caspase activation. Cell Biol Int. 29:489–496. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Green DR and Kroemer G: The
pathophysiology of mitochondrial cell death. Science. 305:626–629.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dong M, Chen SP, Kita K, Ichimura Y, Guo
WZ, Lu S, Sugaya S, Hiwasa T, Takiguchi M, Mori N, et al:
Anti-proliferative and apoptosis-inducible activity of Sarcodonin G
from Sarcodon scabrosus in HeLa cells. Int J Oncol. 34:201–207.
2009.
|
|
24
|
Haupt S, Berger M, Goldberg Z and Haupt Y:
Apoptosis-the p53 network. J Cell Sci. 116:4077–4085. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Normanno N, Maiello MR and De Luca A:
Epidermal growth factor receptor tyrosine kinase inhibitors
(EGFR-TKIs): Simple drugs with a complex mechanism of action? J
Cell Physiol. 194:13–19. 2003. View Article : Google Scholar
|
|
26
|
Ostman A and Heldin CH: PDGF receptors as
targets in tumor treatment. Adv Cancer Res. 97:247–274. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Field JK and Spandidos DA: The role of ras
and myc oncogenes in human solid tumours and their relevance in
diagnosis and prognosis (review). Anticancer Res. 10:1–22.
1990.PubMed/NCBI
|
|
28
|
Slamon DJ, Clark GM, Wong SG, Levin WJ,
Ullrich A and McGuire WL: Human breast cancer: Correlation of
relapse and survival with amplification of the HER-2/neu oncogene.
Science. 235:177–182. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ogawara Y, Kishishita S, Obata T, Isazawa
Y, Suzuki T, Tanaka K, Masuyama N and Gotoh Y: Akt enhances
Mdm2-mediated ubiquitination and degradation of p53. J Biol Chem.
277:21843–21850. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pugazhenthi S, Nesterova A, Sable C,
Heidenreich KA, Boxer LM, Heasley LE and Reusch JE: Akt/protein
kinase B up-regulates Bcl-2 expression through cAMP-response
element-binding protein. J Biol Chem. 275:10761–10766. 2000.
View Article : Google Scholar
|
|
31
|
Prokop A, Wieder T, Sturm I, Essmann F,
Seeger K, Wuchter C, Ludwig WD, Henze G, Dörken B and Daniel PT:
Relapse in childhood acute lymphoblastic leukemia is associated
with a decrease of the Bax/Bcl-2 ratio and loss of spontaneous
caspase-3 processing in vivo. Leukemia. 14:1606–1613. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
King JC, Lu QY, Li G, Moro A, Takahashi H,
Chen M, Go VL, Reber HA, Eibl G and Hines OJ: Evidence for
activation of mutated p53 by apigenin in human pancreatic cancer.
Biochim Biophys Acta. 1823:593–604. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Redston MS, Caldas C, Seymour AB, Hruban
RH, da Costa L, Yeo CJ and Kern SE: p53 mutations in pancreatic
carcinoma and evidence of common involvement of homocopolymer
tracts in DNA microdeletions. Cancer Res. 54:3025–3033.
1994.PubMed/NCBI
|
|
34
|
Jia LQ, Osada M, Ishioka C, Gamo M, Ikawa
S, Suzuki T, Shimodaira H, Niitani T, Kudo T, Akiyama M, et al:
Screening the p53 status of human cell lines using a yeast
functional assay. Mol Carcinog. 19:243–253. 1997. View Article : Google Scholar : PubMed/NCBI
|