Introduction
Mesenchymal stem cells (MSCs), which were first
derived from bone marrow, have self-renewal properties and are able
to differentiate into a variety of mesenchymal tissue types
(1–3). In stem cell therapy, human bone
marrow-derived MSCs (BM-MSCs) are expanded in vitro and
subsequently autoimplanted, which eliminates the risk of immune
rejection. BM-MSCs are able to differentiate into osteoblasts,
chondrocytes and adipocytes (4),
and are a major source of bone regeneration and remodeling during
homeostasis (5–8). In addition, immunophenotype
evaluation demonstrated that mouse BM-MSCs express Sca-1 and CD44,
but not CD11b or CD45 (9).
The transforming growth factor (TGF)-β superfamily
includes the TGF-β/activin/Nodal family and the bone morphogenetic
protein (BMP)/growth and differentiation factor (GDF)/Mullerian
inhibiting substance (MIS) family (10). On the cell surface, binding of
ligands to receptors triggers the formation of a tetrameric complex
of type I and II receptors. Type II receptor kinase activates type
I receptor kinase, which transduces the signal through
phosphorylation of receptor-activated Smads (R-Smads) (11–14).
Smad proteins are the central mediators of TGF-β superfamily
signaling. R-Smads, including Smad 1, Smad 5 and Smad 8, are
primarily activated by BMP-specific type I receptors, whereas Smad
2 and Smad 3 are activated by the TGF-β-specific type I receptors.
Activated R-Smads form complexes with the common mediator Smads
(Co-Smads; e.g., Smad 4), which translocate into the nucleus, where
they and their partner proteins regulate the transcription of
specific target genes. Abnormal intensity of Smad-mediated
TGF-β/BMP signals is associated with various human diseases,
including bone and immune disorders, fibrosis, and cancer
progression or metastasis (15).
Of note, TGF-β superfamily-induced intracellular signals affect
osteogenesis and adipogenesis of MSCs; for instance, BMP has been
observed to potentiate osteogenic and adipogenic differentiation of
undifferentiated mesenchymal cells (16). By contrast, TGF-β potentiates
osteogenic differentiation of BM-MSCs (17,18),
although none of these results have been confirmed in
vivo.
Recent studies have focused on controlling TGF-β/BMP
signals for the discovery of pharmacotherapeutics; however, the
detection of therapeutic molecular targets in these pathways has
not been successful, probably because most trials are performed
in vitro, not in vivo (19,20).
Therefore, it is important to establish appropriate in vivo
experimental models to evaluate the role of TGF-β/BMP signaling in
disease development or healing. The present study aimed to
establish MSC cell lines derived from bone marrow of green
fluorescent protein (GFP)-transgenic mice; the cells and their
diverse, intracellular BMP and TGF-β signals can be tracked after
transplantation into in vivo experimental models. These cell
lines are available for in vivo molecular studies that aim
to determine how the TGF-β superfamily affects MSC proliferation
and differentiation in diseases including fibrosis and cancer
progression or metastasis (21,22),
and in tissue repair processes, including tissue reconstruction and
anti-inflammatory responses (23).
Materials and methods
Bone marrow-derived cells from
GFP-transgenic mice
All experimental procedures were performed in
accordance with the guidelines established by the Animal Studies
Committee at Iwate Medical University (Iwate, Japan). A total of
four GFP-transgenic mice (24)
were obtained from the Center for in vivo Science, Iwate
Medical University (Iwate, Japan). The mice were sacrificed by
excessive inhalation of CO2. Cells were flushed from the
tibia of three-week-old GFP-transgenic mice with phosphate-buffered
saline (PBS) containing 0.5% fetal bovine serum (FBS; PAA
Laboratories, GE Healthcare, Piscataway, NJ, USA) and 2 mM EDTA,
and then seeded into plastic cell culture dishes (Nunc; Thermo
Fisher Scientific, Waltham, MA, USA) with Dulbecco's modified
Eagle's medium (DMEM; Sigma-Aldrich, St. Louis, MO, USA) containing
10% FBS. The cells were cultured for 1 week under hypoxic
conditions (5% O2, 5% CO2 and 90%
N2). Cells were re-plated upon reaching 80%
confluence.
Co-transfection of hTERT and SV40 large T
antigen (SV40LT) genes
The expanded cells were transfected with
pBABE-neo-hTERT and pBABE-pur-SV40LT plasmids encoding neomycin and
puromycin resistance (provided by Addgene, Cambridge, MA, USA) with
Lipofectamine LTX (Invitrogen Life Technologies, Carlsbad, CA, USA)
according to manufacturer instructions. Cells were then incubated
in DMEM containing 10% FBS, 150 µg/ml G418 (Gibco-BRL, Thermo
Fisher Scientific) or 1 µg/ml puromycin (Gibco-BRL) under hypoxic
conditions for 12–15 days. The surviving cells were trypsinized and
allowed to grow in 90-mm culture dishes.
Single-cell cloning
Single-cell clones were obtained using the limited
dilution method. After hTERT and SV40LT transfection and selection
with G418 and puromycin, the surviving cells were seeded on a
96-well plate (Nunc) at 0.5% cells per well, and then cultured
under hypoxic conditions. After 10 days, the cells were
sub-cultured in 24-well plates (Nunc). This was repeated until
confluence was reached at 20 days after single cell cloning.
Population doubling (PD) was defined as the number of doublings
required for a single cell to reach confluence in a 60-mm culture
dish (Nunc) under hypoxic conditions. PD was estimated for clones
SG-2, -3, -5 and -6.
Telomeric repeat amplification
protocol
Telomerase activity in bone marrow-derived cell
lines was assayed by the stretch PCR method using the Quantitative
Telomerase Detection kit (Allied Biotech, Vallejo, CA, USA)
according to manufacturer's instructions. The PCR mixture contained
QTD premixed buffer and SYBR green one dye (cat. no. MT3010; Allied
Biotech, Inc., St. Benicia, CA, USA). Quantification of telomerase
activity was performed under the following amplification conditions
using a Thermal Cycler Dice Real Time system (Takara Bio, Otsu,
Japan) according to manufacturer's instructions: 25°C for 20 min,
initial activation at 95°C for 10 min, denaturation at 90°C for 30
s, annealing at 60°C for 30 s, and a final extension of 40 cycles
at 72°C for 30 s. The PCR products were separated by 10–20%
polyacrylamide gel electrophoresis and stained with ethidium
bromide.
Detection of SV40LT by
immunocytochemistry
Bone marrow-derived cell lines were seeded onto
eight-well culture slides (BD Biosciences, Franklin Lakes, NJ,
USA). After 24 h, the cells were fixed with 4% paraformaldehyde and
washed five times with PBS. For detection of SV40LT, cells were
incubated with mouse monoclonal anti-SV40LT (1:100; cat. no.
ab16879; Abcam, Cambridge, UK) antibody for 1 h at room
temperature. The cells were then incubated with Alexa Fluor 594
goat polyclonal anti-mouse secondary antibodies (1:500; cat. no.
A11005; Thermo Fisher Scientific) and DAPI (1:500; cat. no. D9542;
Sigma-Aldrich) for 30 min at room temperature. Fluorescence was
examined by using a fluorescence microscope (Olympus ix70; Olympus
Corporation, Tokyo, Japan).
Detection of MSC markers by flow
cytometry
A total of 1.0×105 bone marrow-derived
cell lines (SG-2, -3, -5 and -6) were suspended in PBS containing
0.5 FBS and 2 mM EDTA and incubated with phycoerythrin-conjugated
monoclonal anti-mouse Sca-1 (1:10; cat. no. 130-093-224),
monoclonal anti-mouse CD44 (1:10; cat. no. 130-096-838), monoclonal
anti-mouse CD11b (1:10; cat. no. 130-091-240) or monoclonal
anti-mouse CD45 (1:10; cat. no. 130-091-610) antibody for 1 h at
4°C in the dark. All antibodies were purchased from Miltenyi Biotec
(Bergisch Gladbach, Germany). Acquisition was performed using an
EPICS XL ADC system (Beckman Coulter, Brea, CA, USA).
Adipogenic and osteogenic
differentiation
In vitro differentiation was performed
according to a previous study by our group (25). To induce osteogenic
differentiation, confluent cells were incubated in osteogenic
differentiation medium (ODM) under hypoxic conditions for two
weeks. Bone matrix mineralization was evaluated by Alizarin red S
(Sigma-Aldrich) staining. To induce adipogenic differentiation,
cells were cultured to near confluence and cultured in adipogenic
differentiation medium (ADM) under hypoxic conditions for two
weeks. At the end of the differentiation period, lipid droplets
were stained with Oil Red O (Sigma-Aldrich).
Expression profiling of cytokines and
cytokine receptors
Gene expression profiling was performed using a
PrimerArray of mouse cytokine-cytokine receptor interaction (Takara
Bio) in combination with a Thermal Cycler Dice Real Time System
(Takara Bio) according to manufacturer's instructions. This
PrimerArray is a set of real-time reverse transcription-polymerase
chain reaction (RT-PCR) primers used for the analysis of RNA
expression. The array contains a mixture of 96 primer pairs for 88
target genes and eight housekeeping genes. Quantification of gene
expression was performed using a PrimerArray Analysis Tool version
2.0 (Takara Bio).
Western blot analysis
SG-2, -3 and -5 cells were serum-starved overnight
and stimulated with 50 ng/ml BMP-2 (R&D Systems, Minneapolis,
MN, USA) or 50 ng/ml TGF-β1 (R&D Systems). The cells were
washed twice with ice-cold PBS and then lysed in
radioimmunoprecipitation assay buffer (50 mM Tris-HCl, pH 7.2, 150
mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, and 0.1% SDS)
containing protease and phosphatase inhibitor cocktails
(Sigma-Aldrich). The protein content was quantified using the
bicinchoninic acid method (Pierce; Thermo Fisher Scientific).
Samples containing equal amounts of protein were separated by 12.5%
SDS-PAGE and transferred onto a polyvinylidene difluoride membrane
(Merck Millipore, Darmstadt, Germany). After blocking with 5%
nonfat dry milk in 50 mM Tris-HCl, pH 7.2, containing 150 mM NaCl
and 0.1% Tween-20 for 2 h at room temperature, the membrane was
incubated with primary rabbit monoclonal anti-phospho-Smad 5
(1:1,000; cat. no. ab92698; Abcam), anti-Smad 1/5/8 (1:1,000, cat.
no. 12656; Cell Signaling Technology, Danvers, MA, USA), rabbit
monoclonal anti-phospho-Smad 2 (1:1,000; cat. no. 04-953; Merck
Millipore), or anti-Smad 2/3 (1:1,000; cat. no. 610843; BD
Biosciences) antibody overnight at 4°C, with mouse monoclonal
anti-β-actin (1:1,000; cat. no. sc-47778; Santa Cruz Biotechnology,
Dallas, TX, USA) antibody as a loading control. The blots were
incubated with alkaline phosphatase-conjugated secondary antibody
and developed using the
5-bromo-4-chloro-3′-indolyphosphate/nitro-blue tetrazolium membrane
phosphatase substrate system (cat. no. 50-81-00; Kirkegaard &
Perry Laboratories, Gaithersburg, MD, USA).
RNA isolation and RT-quantitative
(q)PCR
SG-2, -3 and -5 cells were stimulated with TGF-β1
(1–50 ng/ml; R&D Systems) for 24 h. Total RNA was isolated
using ISOGEN reagent (Nippongene, Tokyo, Japan) according to the
manufacturer's instructions. First-strand cDNA was synthesized from
total RNA with the PrimeScript RT Reagent kit (Takara Bio). RT-qPCR
was performed on a Thermal Cycler Dice Real Time System (Takara
Bio) with SYBR Premix Ex Taq II (Takara Bio). Expression of
CCAAT/enhancer binding protein-β (C/EBPβ) was normalized to β-actin
and relative expression levels were calculated as a fold-increase
or -decrease relative to the control. Transcripts were detected
with primers (designed using a Perfect Real Time Support system;
Takara Bio) for C/EBPβ (sense, 5′-GACAAGCTGAGCGACGAGTA-3′; and
anti-sense, 5′-AGCTGCTCCACCTTCTTCTG-3′) and β-actin (sense,
5′-CATCCGTAAAGACCTCTATGCCAAC-3′ and anti-sense,
5′-ATGGAGCCACCGATCCACA-3′). For each PCR run, cDNA derived from 50
ng total RNA was used. Following initial denaturation at 95°C for
30 s, a two-step cycle procedure was used (denaturation at 95°C for
5 s and annealing and extension at 60°C for 30 s) for 40 cycles.
The relative mRNA expression levels in each sample were calculated
using the 2−ΔΔCT method (26).
Statistical analysis
All experiments were repeated at least three times.
Representative images or data are shown. Values are expressed as
the mean ± standard deviation. Differences between control and test
samples were analyzed using paired two-tailed Student's t-tests.
P<0.05 was considered to indicate a statistically significant
difference between values.
Results
Establishment of cell lines from the bone
marrow of GFP-transgenic mice
Bone marrow cells were flushed from the tibia of GFP
mice and cultured under hypoxic conditions. Adherent cells were
transformed with hTERT and SV40LT vectors, yielding four single
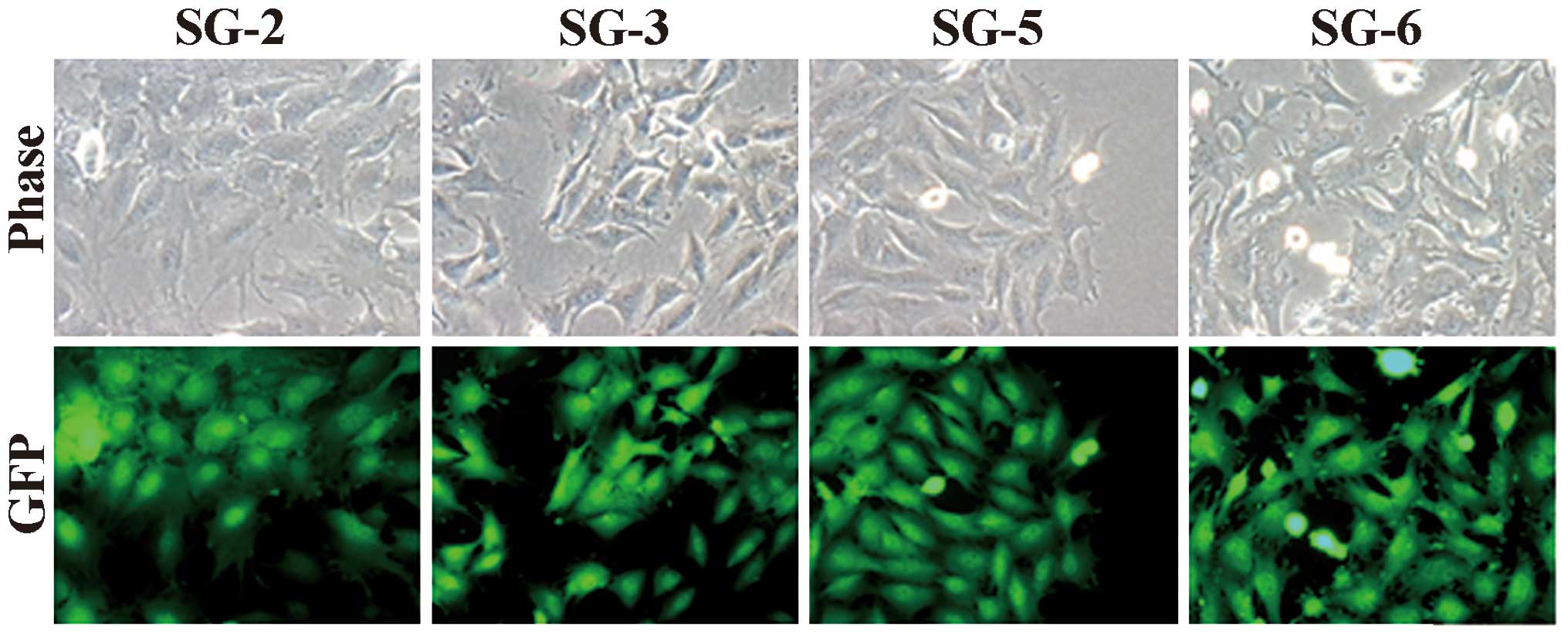
cell-derived cell lines SG-2, -3, -5 and -6. As shown in Fig. 1, the cell lines exhibited
fibroblastic morphology (Fig. 1,
upper panel) and GFP fluorescence (Fig. 1, lower panel). All cell lines
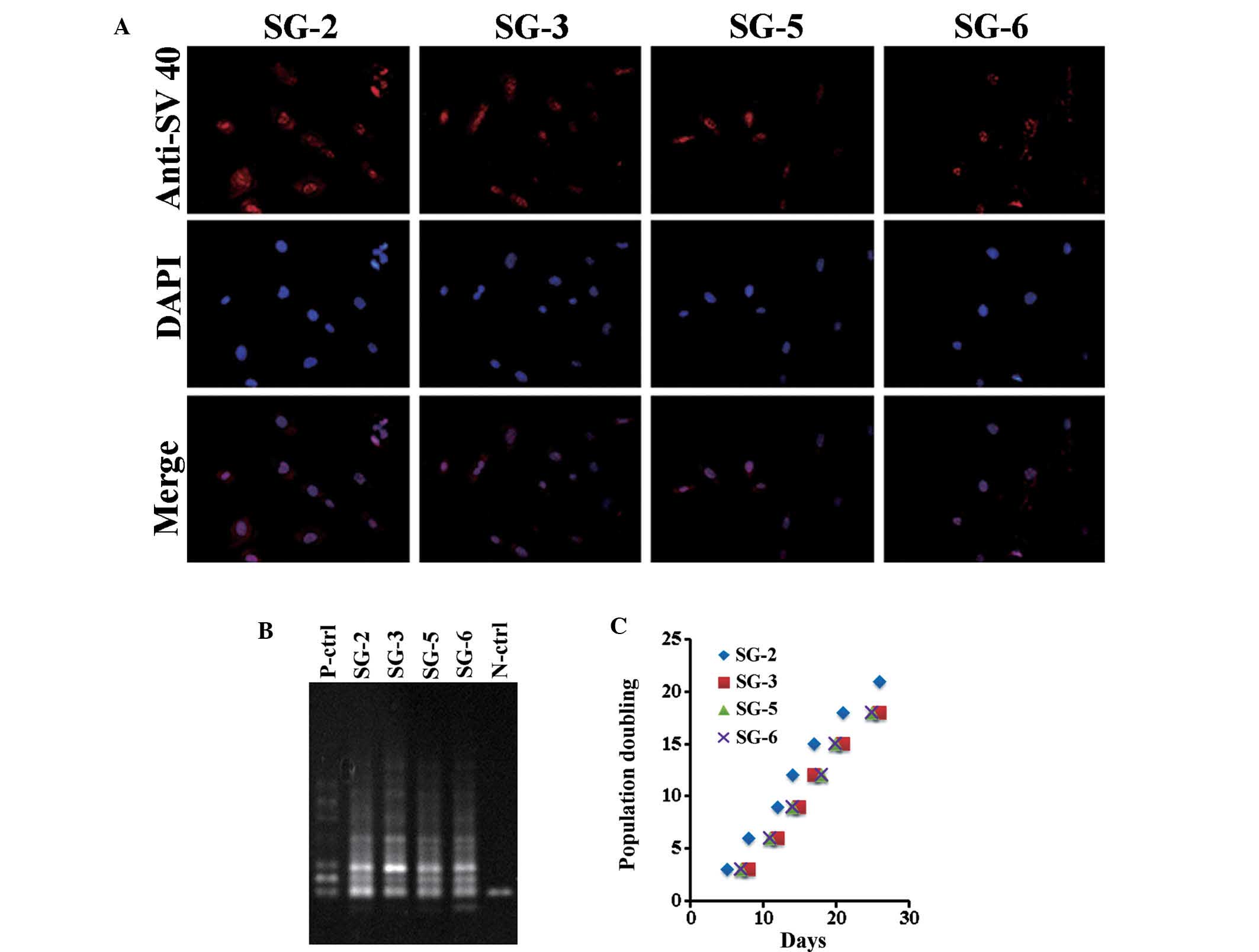
exhibited nuclear SV40LT expression (Fig. 2A). At PD 20, a stretch PCR assay
indicated telomerase activity in all cell lines, but not in the
negative control (Fig. 2B). Thus,
the bone marrow-derived cell lines exhibited telomerase activity
and SV40LT expression. All cell lines grew at a similar rate of ~1
PD every two days (Fig. 2C). The
cells divided at least 60 times and were passaged >30 times,
thus demonstrating successful immortalization.
Bone marrow-derived SG cell lines have
MSC-like features
To determine their MSC character, the SG-2, -3, 5,
and -6 cell lines were analyzed for the expression of mouse MSC
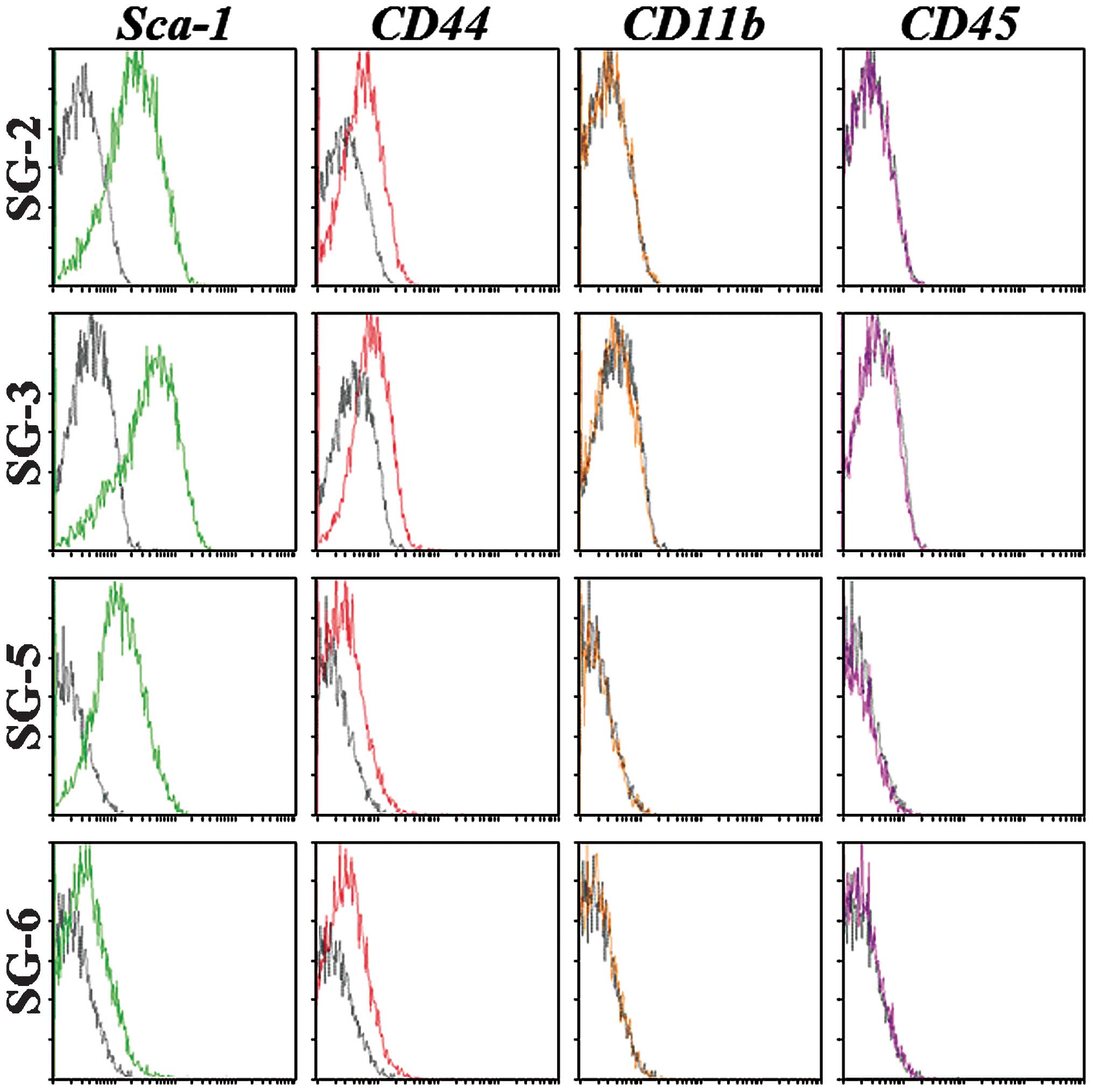
markers and differentiation potential. Sca-1 is the most reliable
MSC marker in mice and was strongly expressed in SG-2, -3 and -5
cells, but only weakly expressed in SG-6 cells (Fig. 3). CD44 was detected at similar
levels in all cell lines, although the hematopoietic stem cell
markers CD11b and CD45 were not detected. The expression patterns
of MSC markers suggested that the SG-2, -3 and -5 cell lines were
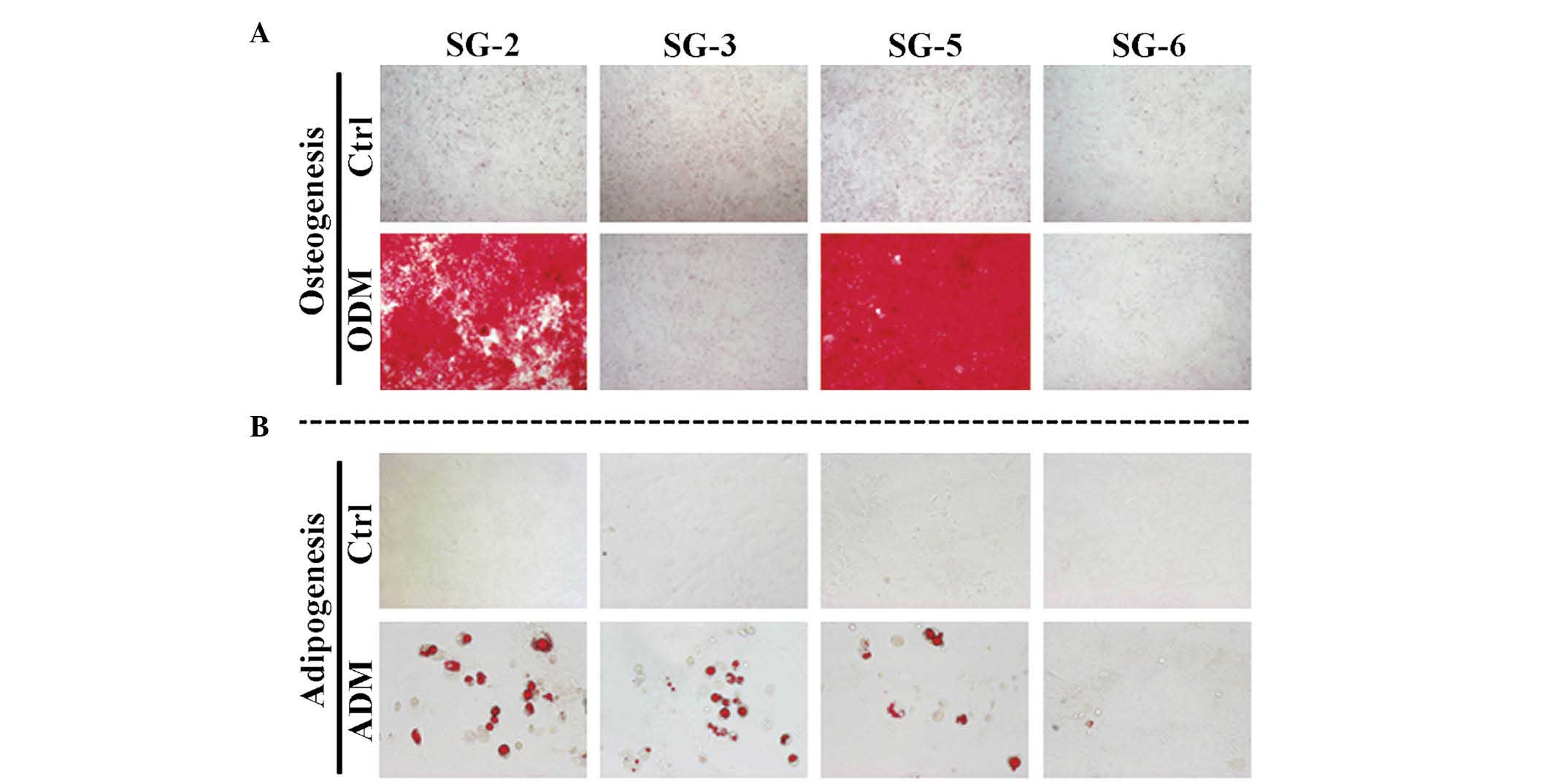
MSCs. Next, the present study evaluated the osteogenic and
adipogenic differentiation potentials of these cell lines. Bone
matrix mineralization indicated by Alizarin red staining was
highest in SG-5 cells (Fig. 4A).
Although miner-alization was observed in SG-2 cells, it was
markedly lower than that in SG-5 and was not detected in SG-3 and
SG-6 cells. Thus, SG-2 and -5 cells retained their osteogenic
differentiation potential, although at different levels of
efficiency. Lipid droplet formation indicated by Oil Red O staining
was more intense in SG-2 cells than in SG-3 and -5 cells (Fig. 4B). Thus, SG-2, -3, and -5 retained
their adipogenic differentiation potential to various degrees.
Intercellular signaling by TGF-β and BMP
in SG cells
In order to identify the expression profiles of
cytokines and cytokine receptors in SG cells, the present study
performed PrimerArray analyses and compared the results derived
from SG-2, -3 and -5 cells. Differentially expressed genes are
shown in Table I. BMP receptor 1B
(Bmpr1b) was most highly expressed in SG-3 cells, while
TGF-β receptor II (Tgfbr2) was most highly expressed in SG-3
and -5 cells, indicating differential sensitivities to BMP-2 and
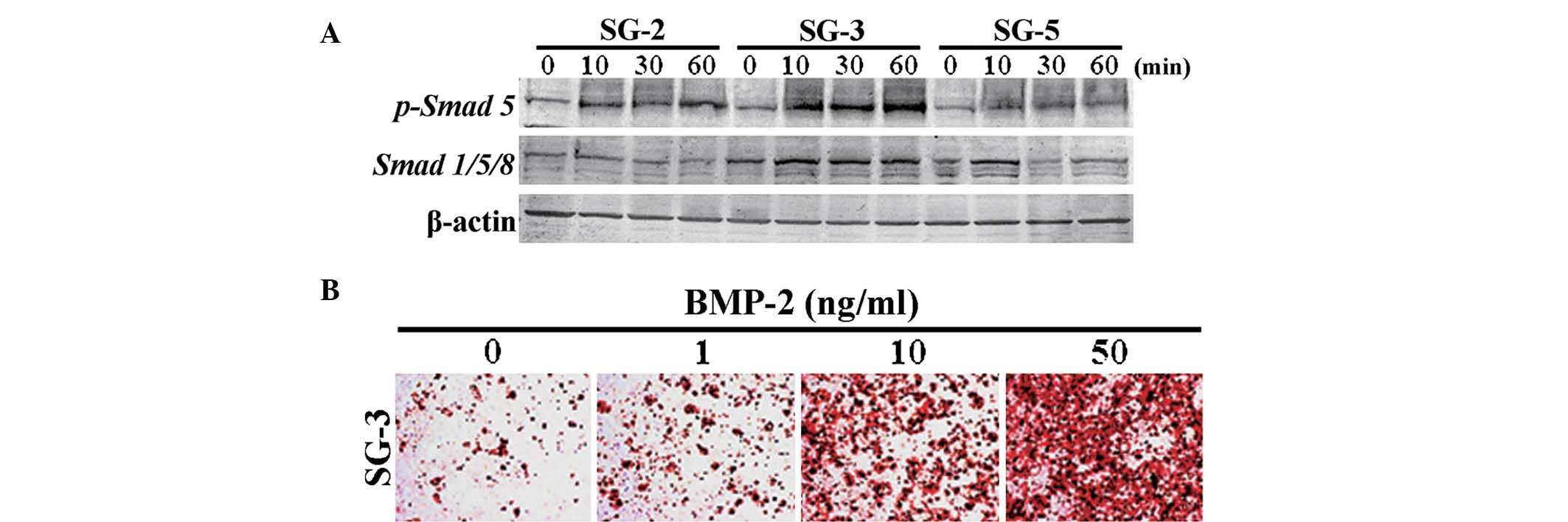
TGF-β. Smad 5, which is a major signaling factor activated by BMP,
was most significantly phosphorylated in BMP 2-stimulated SG-3
cells (Fig. 5A). In addition,
osteogenic differentiation in SG-3 cells was induced by stimulation
with BMP-2 in a dose-dependent manner (Fig. 5B), suggesting these cells retained
the capacity to differentiate into adipogenic (Fig. 4B) and osteogenic lineages. However,
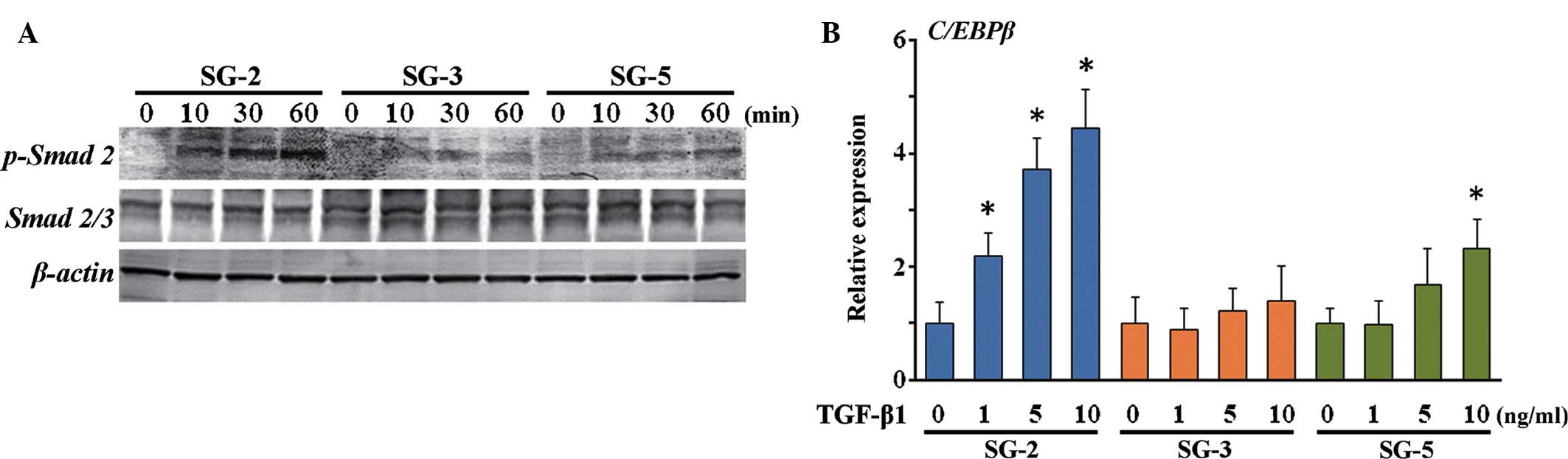
phosphorylation of Smad 2, which is activated by TGF-β, was highest
in the MSC-like SG-2 cells in response to TGF-β1 (Fig. 6A). Of note, mRNA expression of
C/EBPβ, which is an immune- and inflammatory response-associated as
well as Smad-interacting transcription factor, was induced in SG-2
cells by TGF-β1 in a dose-dependent manner (Fig. 6B). Thus, the SG-3 cells were
BMP-responsive and the SG-2 cells were TGF-β-responsive, while SG-5
cells were BMP/TGF-β-unresponsive MSCs.
 | Table IGenes for which expression changed by
>2-fold in SG-2 vs. SG-3 and SG-5. |
Table I
Genes for which expression changed by
>2-fold in SG-2 vs. SG-3 and SG-5.
| Gene symbol | Gene name | Fold change with
SG-2
|
|---|
| vs. SG-3 | vs. SG-5 |
|---|
| Bmpr1b | Bone morphogenetic
protein receptor, type 1B | 45.89 | −2.362 |
| Egfr | Epidermal growth
factor receptor | 4.228 | 6.453 |
| Ifngr2 | Interferon γ
receptor 2 | 1.181 | −3.534 |
| Il17ra | Interleukin 17
receptor A | 2.266 | 1.790 |
| Il18r1 | Interleukin 18
receptor 1 | 3.811 | 1.007 |
| Pdgfrb | Platelet derived
growth factor receptor, beta polypeptide | 2.514 | 1.625 |
| Cx3cl1 | Chemokine (C-X3-C
motif) ligand 1 | 5.315 | 2.346 |
| Tgfbr1 | Transforming growth
factor, beta receptor I | 2.056 | 1.778 |
| Tgfbr2 | Transforming growth
factor, beta receptor II | 2320 | 2702 |
| Hgf | Hepatocyte growth
factor | 14.83 | 5.098 |
| Kdr | Kinase insert
domain protein receptor | 8.574 | 2.888 |
| Lepr | Leptin
receptor | 2.144 | 1.495 |
| Pdgfb | Platelet-derived
growth factor, B polypeptide | 4.823 | 4.993 |
| Pdgfra | Platelet-derived
growth factor receptor, alpha polypeptide | 2.329 | 1.444 |
| Prlr | Prolactin
receptor | −2.071 | −1.014 |
| Ccl9 | Chemokine (C-C
motif) ligand 9 | −2.174 | 1.050 |
|
Tnfrsf1b | Tumor necrosis
factor receptor superfamily, member 1b | 2.129 | 1.879 |
| Cd40 | CD40 antigen | −1.319 | −27.48 |
| Il2rg | Interleukin 2
receptor, γ chain | 7.160 | 2.789 |
| Lifr | Leukemia inhibitory
factor receptor | 1.602 | 4.112 |
| Kitl | Kit ligand | 2.000 | 1.283 |
| Cntfr | Ciliary
neurotrophic factor receptor | 5.938 | 5.242 |
| Cxcl14 | Chemokine (C-X-C
motif) ligand 14 | 7.835 | 5.205 |
| Il17rb | Interleukin 17
receptor B | −2.250 | 3.074 |
| Ccl8 | Chemokine (C-C
motif) ligand 8 | 1.919 | 2.346 |
Discussion
In the present study, the bone marrow of GFP mice
was used to establish three MSC lines immortalized by transfection
with SV40LT and hTERT. SV40LT-transformed cells are not tumorigenic
(27–30); therefore, SV40LT is commonly used
to immortalize primary mammalian cells. Another commonly used gene
for immortalization is TERT, which maintains telomere length to
enable cells to indefinitely proliferate. TERT expression is high
in stem cells, while it is reduced upon differentiation.
Restoration of TERT activity in normal somatic cells can lead to
their immortalization and may be associated with the acquisition of
characteristics associated with cellular transformation (31). It has been indicated that ectopic
expression of the mouse TERT catalytic subunit does not affect
embryonic stem cell proliferation or differentiation in
vitro, but protects them from cell death during differentiation
(32). Therefore, the cell lines
generated in the present study may be used to study stem cell
proliferation and differentiation.
The expression of MSC markers Sca-1+ and
CD44+ and the absence of hematopoietic stem cell markers
CD11b− and CD45− were confirmed in SG-2, -3
and -5 cells, which exhibited osteogenic and adipogenic
differentiation potential. These results strongly suggested that
the MSC-like potential of the cells was preserved. The present
study focused on cytokines and cytokine receptors that are
expressed specifically in each cell line to clarify the
differentiation mechanism of MSCs: Bmpr1b was most highly
expressed in SG-3 but not in SG-2 or -5 cells, whereas
Tgfbr2 was most highly expressed in SG-3 and -5 but not in
SG-2 cells. BMP-2 induced phosphorylation of Smad 5 in SG-3 but not
in SG-2 and -5 cells. Furthermore, BMP-2 induced osteogenic
differentiation of SG-3 cells but did not affect osteogenic
differentiation in SG-2 and -5 cells (data not shown). By contrast,
TGF-β unexpectedly but clearly induced Smad 2 phosphorylation in
SG-2 cells, in which expression of TGF-β receptors I and II was
lower than in SG-3 and -5 cells, suggesting that Smad 2 itself or
signal transduction molecules upstream of Smad 2 may have been
inactivated in SG-3 and -5 cells. Thus, the present study
established TGF-β-responsive SG-2 cells, BMP-responsive SG-3 cells
and TGF-β/BMP-unresponsive SG-5 cells that can be traced by GFP
fluorescence after transplantation into in vivo experimental
models.
Of note, TGF-β stimulation of SG-2 induced
expression of C/EBPβ, a Smad-interacting transcription factor
(33). C/EBPβ is a member of the
C/EBP family of transcription factors (C/EBPα, C/EBPβ, C/EBPγ,
C/EBPδ, C/EBPε and C/EBPξ) (34–36).
C/EBPβ was first described in 1990 as a basic leucine
zipper-structured factor that binds to the interleukin
(IL)-1-responsive element in the IL-6 promoter (37). C/EBPβ is highly expressed in
myelomonocytic cells and macrophages (38–41).
Extracellular signals, including differentiation- or
proliferation-inducing agents, hormones, cytokines and inflammatory
substances, as well as bacterial and other microbial products can
activate or inhibit C/EBPβ via distinct signal transduction
pathways. The expression and/or activation of C/EBPβ is regulated
by transcriptional mechanisms, mammalian target of rapamycin
(mTOR)-mediated alternative translation, post-translational
modifications and protein-protein interactions (35,42,43).
Upon its activation, C/EBPβ induces or represses a variety of
genes, including cytokines, chemokines and their receptors, other
pro-inflammatory genes and pro-proliferative or
differentiation-associated markers, as well as metabolic enzymes
(34). C/EBPβ thereby affects
associated cellular functions, including proliferation (40,42),
differentiation (39,41,44),
metabolic regulation (45,46) and orchestration of the immune
response (47–49). Furthermore, C/EBPβ is implicated in
the pathogenesis of various common diseases, including cancer,
hyper-/hypo-inflammation and bacterial/viral infections (34,50).
Expression and activation of C/EBPβ induces the production of
monocyte chemotactic protein-1 (MCP-1) (51–54),
a member of the C-C motif chemokine ligand-2, which induces
leukocyte migration to inflamed tissues and organs (55,56).
In addition, MCP-1 is secreted by primary breast tumors and
stimulates migration of MSCs to tumor lesions (57). MCP-1, stromal cell-derived
factor-1, macrophage inflammatory protein-1α and monocyte
chemotactic protein-3 are the most widely reported MSC homing
factors (58–61). Stem cell therapy relies on the
appropriate homing and engraftment capacity of stem cells (62). Therefore, SG-2 can be used for
in vivo studies of TGF-β-dependent anti-inflammation and
stem cell homing.
Abnormal intensity of Smad-mediated TGF-β/BMP
signals in MSCs is associated with various human diseases,
including bone and immune disorders, fibrosis, and cancer
progression or metastasis. However, the detection of therapeutic
molecular targets for these diseases in the TGF-β/BMP signaling
pathways has not been successful as most studies have been
performed in vitro. The present study established MSC lines,
including TGF-β-responsive SG-2, BMP-responsive SG-3, and
TGF-β/BMP-unresponsive SG-5 cells, which can be traced by GFP
fluorescence after transplantation into in vivo experimental
models. These cell lines can be used to explore how
TGF-β/BMP-induced Smad-mediated signals affect proliferation and
differentiation of MSCs in vivo, providing insight into
various human diseases, including bone and immune disorders,
fibrosis and cancer progression or metastasis.
Acknowledgments
The present study was supported in part by KAKENHI
grants-in-aid from the Japan Society for the Promotion of Science
(grant nos. 25893221 to S.S., 25463053 to N.C., 26893248 to J.Y.,
26462932 to H.K. and 26670852 to A.I.), a grant-in-aid for
Strategic Medical Science Research Centre from the Ministry of
Education, Culture, Sports, Science and Technology of Japan
(2010–2014), a grant from the Keiryokai Research Foundation (no.
120 to S.S., 2013) and the Fund of Academic Society for Research in
Otolaryngology (Kansai Medical University, Hirakata, Japan).
References
|
1
|
Prockop DJ: Marrow stromal cells as stem
cells for nonhematopoietic tissues. Science. 276:71–74. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pittenger MF, Mackay AM, Beck SC, Jaiswal
RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S and
Marshak DR: Multilineage potential of adult human mesenchymal stem
cells. Science. 284:143–147. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Docheva D, Popov C, Mutschler W and
Schieker M: Human mesenchymal stem cells in contact with their
environment: Surface characteristics and the integrin system. J
Cell Mol Med. 11:21–38. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Baksh D, Song L and Tuan RS: Adult
mesenchymal stem cells: Characterization, differentiation and
application in cell and gene therapy. J Cell Mol Med. 8:301–316.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kassem M, Abdallah BM and Saeed H:
Osteoblastic cells: Differentiation and transdifferentiation. Arch
Biochem Biophys. 473:183–187. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jones E and Yang X: Mesenchymal stem cells
and bone regeneration: Current status. Injury. 42:562–568. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Proff P and Römer P: The molecular
mechanism behind bone remodelling: A review. Clin Oral Investig.
13:355–362. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lazar-Karsten P, Dorn I, Meyer G, Lindner
U, Driller B and Schlenke P: The influence of extracellular matrix
proteins and mesenchymal stem cells on erythropoietic cell
maturation. Vox Sang. 101:65–76. 2011. View Article : Google Scholar
|
|
9
|
Yew TL, Chang MC, Hsu YT, Weng WH, Tsai
CC, Chiu FY, Hung SC and He FY: Efficient expansion of mesenchymal
stem cells from mouse bone marrow under hypoxic conditions. J
Tissue Eng Regen Med. 7:984–993. 2013. View Article : Google Scholar
|
|
10
|
Weiss A and Attisano L: The TGF beta
superfamily signaling pathway. Wiley Interdiscip Rev Dev Biol.
2:47–63. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Miyazawa K, Shinozaki M, Hara T, Furuya T
and Miyazono K: Two major Smad pathways in TGF-beta superfamily
signalling. Genes Cells. 7:1191–1204. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Heldin CH, Landström M and Moustakas A:
Mechanism of TGF-beta signaling to growth arrest, apoptosis and
epithelial-mesenchymal transition. Curr Opin Cell Biol. 21:166–176.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Meulmeester E and Ten Dijke P: The dynamic
roles of TGF-β in cancer. J Pathol. 223:205–218. 2011. View Article : Google Scholar
|
|
14
|
Imamura T, Oshima Y and Hikita A:
Regulation of TGF-β family signaling by ubiquitination and de
ubiquitination. J Biochem. 154:481–489. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Herhaus L and Sapkota GP: The emerging
roles of deubiquitylating enzymes (DUBs) in the TGFβ and BMP
pathways. Cell Signal. 26:2186–2192. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee SY, Lee JH, Kim JY, Bae YC, Suh KT and
Jung JS: BMP2 increases adipogenic differentiation in the presence
of dexamethasone, which is inhibited by the treatment of TNF-α in
human adipose tissue-derived stromal cells. Cell Physiol Biochem.
34:1339–1350. 2014. View Article : Google Scholar
|
|
17
|
Pountos I, Georgouli T, Henshaw K, Bird H,
Jones E and Giannoudis PV: The effect of bone morphogenetic
protein-2, bone morphogenetic protein-7, parathyroid hormone and
platelet-derived growth factor on the proliferation and osteogenic
differentiation of mesenchymal stem cells derived from osteoporotic
bone. J Orthop Trauma. 24:552–556. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yokota J, Chosa N, Sawada S, Okubo N,
Takahashi N, Hasegawa T, Kondo H and Ishisaki A: PDGF-induced
PI3K-mediated signal enhances TGF-β-induced osteogenic
differentiation of human mesenchymal stem cells in the
TGF-β-activated MEK-dependent manner. Int J Mol Med. 33:534–542.
2014.PubMed/NCBI
|
|
19
|
Ray P and Chapman SC: Cytoskeletal
reorganaization drives mesenchymal condensation and regulates
downstream molecular signaling. PLoS One. 10:e01347022015.
View Article : Google Scholar
|
|
20
|
Sengle G, Carlberg V, Tufa SF, Charbonneau
NL, Smaldone S, Carlson EJ, Ramirez F, Keene DR and Sakai LY:
Abnormal activation of BMP signaling causes myopathy in Fbn2 null
mice. PLoS Genet. 11:e10053402015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kramman R and Humphreys BD: Kidney
pericytes: Roles in regeneration and fibrosis. Semin Nephrol.
34:374–383. 2014. View Article : Google Scholar
|
|
22
|
Touboul C, Vidal F, Pasquier J, Lis R and
Rafii A: Role of mesenchymal cells in the natural history of
ovarian cancer: A review. J Transl Med. 12:2712014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wise AF and Ricardo SD: Mesenchymal stem
cells in kidney inflammation and repair. Nephrology (Carlton).
17:1–10. 2012. View Article : Google Scholar
|
|
24
|
Okabe M, Ikawa M, Kominami K, Nakanishi T
and Nishimune Y: 'Green mice' as a source of ubiquitous green
cells. FEBS Lett. 407:313–319. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Aomatsu E, Takahashi N, Sawada S, Okubo N,
Hasegawa T, Taira M, Miura H, Ishisaki A and Chosa N: Novel
SCRG1/BST1 axis regulates self-renewal, migration and osteogenic
differentiation potential in mesenchymal stem cells. Sci Rep.
4:36522014. View Article : Google Scholar
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
27
|
Gee CJ and Harris H: Tumorigenicity of
cells transformed by Simian virus 40 and of hybrids between such
cells and normal diploid cells. J Cell Sci. 36:223–240.
1979.PubMed/NCBI
|
|
28
|
Howell N: Suppression of transformation
and tumorigenicity in interspecies hybrids of human
SV40-transformed and mouse 3T3 cell lines. Cytogenet Cell Genet.
34:215–229. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kahn P, Topp WC and Shin S: Tumorigenicity
of SV40-transformed human and monkey cells in immunodeficient mice.
Virology. 126:348–360. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nitta M, Katabuchi H, Ohtake H, Tashiro H,
Yamaizumi M and Okamura H: Characterization and tumorigenicity of
human ovarian surface epithelial cells immortalized by SV40 large T
antigen. Gynecol Oncol. 81:10–17. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kang MK and Park NH: Extension of cell
life span using exogenous telomerase. Methods Mol Biol.
371:151–165. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee MK, Hande MP and Sabapathy K: Ectopic
mTERT expression in mouse embryonic stem cells does not affect
differentiation but confers resistance to differentiation- and
stress-induced p53-dependent apoptosis. J Cell Sci. 118:819–829.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Abraham S, Sweet T, Khalili K, Sawaya BE
and Amini S: Evidence for activation of the TGF-beta1 promoter by
C/EBP beta and its modulation by Smads. J Interferon Cytokine Res.
29:1–7. 2009. View Article : Google Scholar
|
|
34
|
Ramji DP and Foka P:
CCAAT/enhancer-binding proteins: Structure, function and
regulation. Biochem J. 365:561–575. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zahnow CA: CCAAT/enhancer-binding protein
beta: Its role in breast cancer and associations with receptor
tyrosine kinases. Expert Rev Mol Med. 11:e122009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lekstrom-Himes J and Xanthopoulos KG:
Biological role of the CCAAT/enhancer-binding protein family of
transcription factors. J Biol Chem. 273:28545–28548. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Akira S, Isshiki H, Sugita T, Tanabe O,
Kinoshita S, Nishio Y, Nakajima T, Hirano T and Kishimoto T: A
nuclear factor for IL-6 expression (NF-IL6) is a member of a C/EBP
family. EMBO J. 9:1897–1906. 1990.PubMed/NCBI
|
|
38
|
Williams SC, Cantwell CA and Johnson PF: A
family of C/EBP-related proteins capable of forming covalently
linked leucine zipper dimers in vitro. Genes Dev. 5:1553–1567.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Katz S, Kowenz-Leutz E, Müller C, Meese K,
Ness SA and Leutz A: The NF-M transcription factor is related to
C/EBP beta and plays a role in signal transduction, differentiation
and leukemogenesis of avian myelomonocytic cells. EMBO J.
12:1321–1332. 1993.PubMed/NCBI
|
|
40
|
Haas SC, Huber R, Gutsch R, Kandemir JD,
Cappello C, Krauter J, Duyster J, Ganser A and Brand K: ITD- and
FL-induced FLT3 signal transduction leads to increased C/EBP
beta-LIP expression and LIP/LAP ratio by different signalling
modules. Br J Haematol. 148:777–790. 2010. View Article : Google Scholar
|
|
41
|
Gutsch R, Kandemir JD, Pietsch D, Cappello
C, Meyer J, Simanowski K, Huber R and Brand K:
CCAAT/enhancer-binding protein beta inhibits proliferation in
monocytic cells by affecting the retinoblastoma protein/E2F/cyclin
E pathway but is not directly required for macrophage morphology. J
Biol Chem. 286:22716–22729. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Nerlov C: The C/EBP family of
transcription factors: A paradigm for interaction between gene
expression and proliferation control. Trends Cell Biol. 17:318–324.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tsukada J, Yoshida Y, Kominato Y and Auron
PE: The CCAAT/enhancer (C/EBP) family of basic-leucine zipper
(bZIP) transcription factors is a multifaceted highly-regulated
system for gene regulation. Cytokine. 54:6–19. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Pham TH, Langmann S, Schwarzfischer L, El
Chartouni C, Lichtinger M, Klug M, Krause SW and Rehli M: CCAAT
enhancer-binding protein beta regulates constitutive gene
expression during late stages of monocyte to macrophage
differentiation. J Biol Chem. 282:21924–21933. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liu S, Croniger C, Arizmendi C,
Harada-Shiba M, Ren J, Poli V, Hanson RW and Friedman JE:
Hypoglycemia and impaired hepatic glucose production in mice with a
deletion of the C/EBP beta gene. J Clin Invest. 103:207–213. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Croniger CM, Millward C, Yang J, Kawai Y,
Arinze IJ, Liu S, Harada-Shiba M, Chakravarty K, Friedman JE, Poli
V and Hanson RW: Mice with a deletion in the gene for
CCAAT/enhancer-binding protein beta have an attenuated response to
cAMP and impaired carbohydrate metabolism. J Biol Chem.
276:629–638. 2001. View Article : Google Scholar
|
|
47
|
Poli V: The role of C/EBP isoforms in the
control of inflammatory and native immunity functions. J Biol Chem.
273:29279–29282. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zwergal A, Quirling M, Saugel B, Huth KC,
Sydlik C, Poli V, Neumeier D, Ziegler-Heitbrock HW and Brand K:
C/EBP beta blocks p65 phosphorylation and thereby NF-kappa
B-mediated transcription in TNF-tolerant cells. J Immunol.
177:665–672. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cappello C, Zwergal A, Kanclerski S, Haas
SC, Kandemir JD, Huber R, Page S and Brand K: C/EBP beta enhances
NF-kappaB-associated signalling by reducing the level of
IkappaB-alpha. Cell Signal. 21:1918–1924. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Liu Y, Nonne macher MR and Wigdahl B:
CCAAT/enhancer-binding proteins and the pathogenesis of retrovirus
infection. Future Microbiol. 4:299–321. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Spooner CJ, Guo X, Johnson PF and Schwartz
RC: Differential roles of C/EBP beta regulatory domains in
specifying MCP-1 and IL-6 transcription. Mol Immunol. 44:1384–1392.
2007. View Article : Google Scholar
|
|
52
|
Abraham M, Shapiro S, Karni A, Weiner HL
and Miller A: Gelatinases (MMP-2 and MMP-9) are preferentially
expressed by Th1 vs. Th2 cells. J Neuroimmunol. 163:157–164. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Guo J, Zhang H, Xiao J, Wu J, Ye Y, Li Z,
Zou Y and Li X: Monocyte chemotactic protein-1 promotes the
myocardial homing of mesenchymal stem cells in dilated
cardiomyopathy. Int J Mol Sci. 14:8164–8178. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Belema-Bedada F, Uchida S, Martire A,
Kostin S and Braun T: Efficient homing of multipotent adult
mesenchymal stem cells depends on FROUNT-mediated clustering of
CCR2. Cell Stem Cell. 2:566–575. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Gerard C and Rollins BJ: Chemokines and
disease. Nat Immunol. 2:108–115. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
56
|
Rot A and von Andrian UH: Chemokines in
innate and adaptive host defense: Basic chemokinese grammar for
immune cells. Annu Rev Immunol. 22:891–928. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Dwyer RM, Potter-Beirne SM, Harrington KA,
Lowery AJ, Hennessy E, Murphy JM, Barry FP, O'Brien T and Kerin MJ:
Monocyte chemotactic protein-1 secreted by primary breast tumors
stimulates migration of mesenchymal stem cells. Clin Cancer Res.
13:5020–5027. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Schenk S, Mal N, Finan A, Zhang M,
Kiedrowski M, Popovic Z, McCarthy PM and Penn MS: Monocyte
chemotactic protein-3 is a myocardial mesenchymal stem cell homing
factor. Stem Cells. 25:245–251. 2007. View Article : Google Scholar
|
|
59
|
Zhou YL, Zhang HF, Li XL, Di RM, Yao WM,
Li DF, Feng JL, Huang J, Cao KJ and Fu M: Increased
stromal-cell-derived factor 1 enhances the homing of bone marrow
derived mesenchymal stem cells in dilated cardiomyopathy in rats.
Chin Med J (Engl). 123:3282–3287. 2010.
|
|
60
|
Zhuang Y, Chen X, Xu M, Zhang LY and Xiang
F: Chemokine stromal cell-derived factor 1/CXCL12 increases homing
of mesenchymal stem cells to injured myocardium and
neovascularization following myocardial infarction. Chin Med J
(Engl). 122:183–187. 2009.
|
|
61
|
Abbott JD, Huang Y, Liu D, Hickey R,
Krause DS and Giordano FJ: Stromal cell-derived factor-1 alpha
plays a critical role in stem cell recruitment to the heart after
myocardial infarction but is not sufficient to induce homing in the
absence of injury. Circulation. 110:3300–3305. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Mozid AM, Arnous S, Sammut EC and Mathur
A: Stem cell therapy for heart diseases. Br Med Bull. 98:143–159.
2011. View Article : Google Scholar : PubMed/NCBI
|