Introduction
Laryngeal squamous cell carcinoma (LSCC) is a common
malignant tumor of the head and neck region, and it is the eighth
leading cause of cancer-associated mortality worldwide (1). LSCC is likely to metastasize to
regional lymph nodes, which impacts cure rates and survival.
Numerous advanced methods have been developed for diagnosis and
treatment, the mortality rate of LSCC has not improved (2). Thus, an improved understanding of the
underlying mechanisms of LSCC development is key for the
development of novel diagnostic and prognostic markers, and novel
therapeutic targets.
Epigenetic modifications have been recognized as an
important mechanism underlying carcinoma progression. DNA
methylation is one of the best-understood mechanisms of epigenetic
regulation of gene expression. Hypermethylation of CpG islands,
which are located in the promoter regions of tumor-associated
genes, is the predominant mechanism of gene inactivation in cancer
(3). However, different types of
tumor have a different pattern of hypermethylated genes (4). Identification of hypermethylated
genes in LSCC may be important for finding molecular markers to aid
diagnosis, treatment monitoring, and prognosis of LSCC.
Thrombospondin-1 (THBS-1) is a glycoprotein
containing multiple domains, which is important in cell
proliferation, adhesion, angiogenesis, migration, and tumor
metastasis via interaction with numerous proteins and cell
receptors (5). However, the
involvement of THBS-1 in cancer progression remains controversial.
Inhibition of tumor growth by THBS-1 is considered to be associated
with its antiangiogenic activity, which has been well described
(6). By contrast, a number of
previous studies have demonstrated that THBS-1 promotes tumor cell
invasion and metastasis in breast cancer (7), gastric carcinoma (8) and pancreatic carcinoma (9). Thus, the effects of THBS-1 appear to
be specific for the type of tumor examined and the experimental
model used. Promoter hypermethylation of the THBS-1 gene has been
observed in certain primary human carcinomas, including colorectal
cancer (10), melanoma (11) and gastric cardia adenocarcinoma
(12). It has been suggested that
hypermethylated THBS-1 may promote tumorigenesis via its effects on
angiogenesis (10–12). To the best of our knowledge, the
role and methylation status of THBS-1 in LSCC remains to be
elucidated. The present study investigated the expression and role
of THBS-1, and evaluated the association between expression and
methylation status of THBS-1 and clinicopathological parameters of
LSCC. In addition, the current study detected the ability of
5-aza-2′-deoxy-cytidine (5-aza-dC) to induce THBS-1 gene
re-expression and its effect on proliferation and invasion of Hep-2
cells.
Materials and methods
Patients and tissue samples
The present study was conducted on 66 LSCC patients
(24 patients with glottic lesions, 34 with supraglottic lesions and
8 with subglottic lesions), who were histologically and clinically
diagnosed at Chongqing Cancer Institute and the First Affiliated
Hospital of Chongqing Medical University (Chongqing, China) between
2012 and 2013. All patients received no radiotherapy, chemotherapy
or biotherapy prior to operation. Among the 66 LSCC patients, 54
were male and 12 were female, age ranged from 34–76 years, with a
mean age of 62.3 years. A total of 24 patients had positive lymph
node metastasis and 42 patients had negative lymph node metastasis.
Histological grade determined 37 patients were of high grade, 20 of
middle grade, and 9 of low grade. According to the
tumor-node-metastasis (TNM) classification by the Union for
International Cancer Control (1997), 36 were in stage I-II and 30
were in stage III–IV. Fresh tumor tissues and adjacent non-tumorous
tissues were obtained immediately following tumor resection. Each
tissue was snap-frozen in liquid nitrogen and stored at −80°C.
The present study was approved by the ethics
committee of Chongqing Cancer Institute and the First Affiliated
Hospital of Chongqing Medical University. Informed consent was
obtained from all patients. All specimens were handled and made
anonymous according to the ethical and legal standards.
Cell culture and treatment with
5-aza-dC
The Hep-2 human laryngeal carcinoma cell line was
obtained from the Cell Biology Institute of Shanghai, Chinese
Academy of Science (Shanghai, China). The cells were grown in
RPMI-1640 (GE Healthcare Life Sciences, Logan, UT, USA) with 10%
fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), penicillin (100 U/ml) and streptomycin (100
µg/ml) at 37°C in a 5% CO2 humidified atmosphere.
Following culture in a 6-well plate for 24 h, the Hep-2 cells were
treated for times ranging from 6–96 h with the demethylating
reagent, 5-aza-dC (Sigma-Aldrich, St. Louis, MO, USA) at a final
concentration of 0.1, 1, or 5 µM. Dimethyl sulfoxide
(Sigma-Aldrich) served as a control for non-specific solvent
effects on cells.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from human tissue samples
and Hep-2 cells using TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Total RNA was reverse transcribed into cDNA
using a PrimeScript™ RT reagent kit (Perfect Real Time) obtained
from Takara Bio, Inc. (Otsu, Japan) according to the manufacturer's
protocol. qPCR was conducted using the SYBR® Premix Ex
Taq™ II (Tli RNaseH Plus) kit (Takara Bio, Inc.) and the Rotor-Gene
Q Cycler (Qiagen GmbH, Hilden, Germany). The primers for were as
follows: Forward, 5′-TGT TTG TGC AGG AAG ACA GG-3′ and reverse,
5′-TTG TCA AGG GTG AGG AGG AC-3′ for THBS-1; and forward, 5′-CTC
TCT GCT CCT CCT GTT CGAC-3′ and reverse, 5′-TGA GCG ATG TGG CTC
GGCT-3′ for GAPDH. The thermocycling conditions were as follows: 2
min at 95°C; followed by 40 cycles of 95°C for 15 sec, 58°C for 30
sec and 58°C for 30 sec. All samples were analyzed using GAPDH gene
expression as an internal control. The relative mRNA level of
THBS-1 gene expression was determined by the 2−ΔΔCq
method (13).
Western blotting
Briefly, human tissue samples and Hep-2 cells were
homogenized in radioimmunoprecipitation lysis buffer
(Sigma-Aldrich; Merck Millipore, Darmstadt, Germany) to extract the
protein. Total protein concentration was quantified using a Protein
Assay kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Total
protein (50 µg) was separated by electrophoresis on 10%
SDS-PAGE gel and transferred to polyvinylidene difluoride
membranes. The membranes were then blocked with 5% non-fat dry milk
in 1 × Tris buffered-saline-Tween-20 (TBST; Bio-Rad Laboratories,
Inc.) for 1 h at 25°C, membranes were probed with primary
monoclonal anti-THBS-1 antibody (cat. no. sc-59887; 1:1,000) and
β-actin antibody (cat. no. sc-47778; 1:1,000; both from Santa Cruz
Biotechnology, Inc, Dallas, TX, USA) overnight at 4°C. The
membranes were then washed once with TBST and were incubated with
horseradish peroxidase (HRP)-labeled anti-mouse secondary antibody
(cat. no. BA1051, 1:5,000, Boster Biological Technology, Ltd.,
Wuhan, China) at 37°C for 1 h. β-actin served as an internal
control. Membranes were incubated with Immobilon™ Western
chemiluminescent HRP substrate (EMD Millipore, Billerica, MA, USA)
after washing. Signals were detected by ChemiDoc XRS imaging system
(Bio-Rad Laboratories, Inc.). Quantification of bands on western
blots was performed using Quantity One software version 4.6.2
(Bio-Rad Laboratories, Inc.).
DNA extraction and methylation-specific
PCR
Genomic DNA was extracted from human tissue samples
or Hep-2 cells using MasterPure DNA Purification kit (Epicentre
Biotechnologies; Illumina, Inc., San Diego, CA, USA) according to
the manufacturer's protocols. Genomic DNA was modified using the EZ
DNA Methylation™ Bisulfite kit following manufacturer's protocol
(Zymo Research, Irvine, CA, USA). This modification resulted in a
conversion of unmethylated cytosine to thymine, whereas methylated
cytosine remained unchanged. The specific PCR was then used to
distinguish between methylated and unmethylated DNA sequences. The
bisulfite-treated DNA was amplified using EpiTaq™ HS kit (TaKaRa
Bio Inc.). The primer sequences were as follows: Forward, 5′-TTG
AGT ACG TTA AGG TTG CGT GGGC-3′ and reverse, 5′-AAC GCT AAA ACT ACC
GAT ACG CCG AA-3′ (212 bp) for the methylated form; and forward,
5′-GGT TGA GTA TGT TAA GGT TGT GTG GGT-3′ and reverse, 5′-TAA AAA
CAC TAA AAC TAC CAA TAC ACC AAA-3′ (230 bp) for the unmethylated
form. The thermocycling conditions were as follows: 5 min at 94°C;
followed by 35 cycles of 94°C for 30 sec, 60°C for 30 sec, and 72°C
for 30 sec; with a final extension at 72°C for 10 min. PCR products
were analyzed on 2% agarose gels with ethidium bromide and
visualized under UV illumination. Genomic DNA, treated in
vitro with Sss I methyltransferase served as a positive control
for methylated DNA, and DNA from peripheral blood lymphocytes of
healthy individuals served as a control for unmethylated DNA. A
water blank served as a negative control.
Cell viability assay
Cell viability was determined using a Cell Counting
Kit-8 (CCK-8; Beyotime Institute of Biotechnology, Haimen, China).
Hep-2 cells (5×103) were suspended in 100 µl
RPMI-1640 medium containing 10% FBS were seeded in 96-well plates.
After 24 h, the cells were treated with different concentrations
(0.1, 1, or 5 µM) of 5-aza-dC for 24, 48, 72, and 96 h,
respectively. CCK-8 solution (10 µl) was added to each well
and the cultures were incubated at 37°C for 90 min. Absorbance at a
wavelength of 450 nm was measured using an microplate reader. The
results were plotted as the mean ± standard deviation from three
separate experiments with four determinations per experiment for
each experimental condition.
Cell invasion assay
Cell invasive ability was examined using a 24-well
Transwell assay with 8 µm pore polycarbonate membrane
inserts (Corning Incorporated, Corning, NY, USA). Hep-2 cells
incubated with 5 µM 5-Aza-dC for 24, 48 and 72 h,
respectively were detached from the tissue culture plates, washed
with PBS, and planted at the density of 5×104/upper well
in 200 µl of serum-free RPMI-1640 medium. RPMI-1640 (500
µl) supplemented with 10% FBS was added to the lower
chambers as a chemoattractant. Cells were incubated at 37°C in a
humidified 5% CO2 atmosphere for 24 h. Cells that had
successfully invaded through the inserts were fixed in 4%
paraformaldehyde for 30 min and stained with crystal violet. The
invaded cells were counted from five preselected microscopic fields
(magnification, ×200). All experiments were performed in
triplicate.
Statistical analysis
For statistical evaluation, the SPSS 17.0 software
(SPSS Inc., Chicago, IL, USA) was used. Experiments were
independently repeated three times. Differences between groups were
assessed by Student's t-test, one-way analysis of variance followed
by Bonferroni's multiple comparison test or χ2 test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
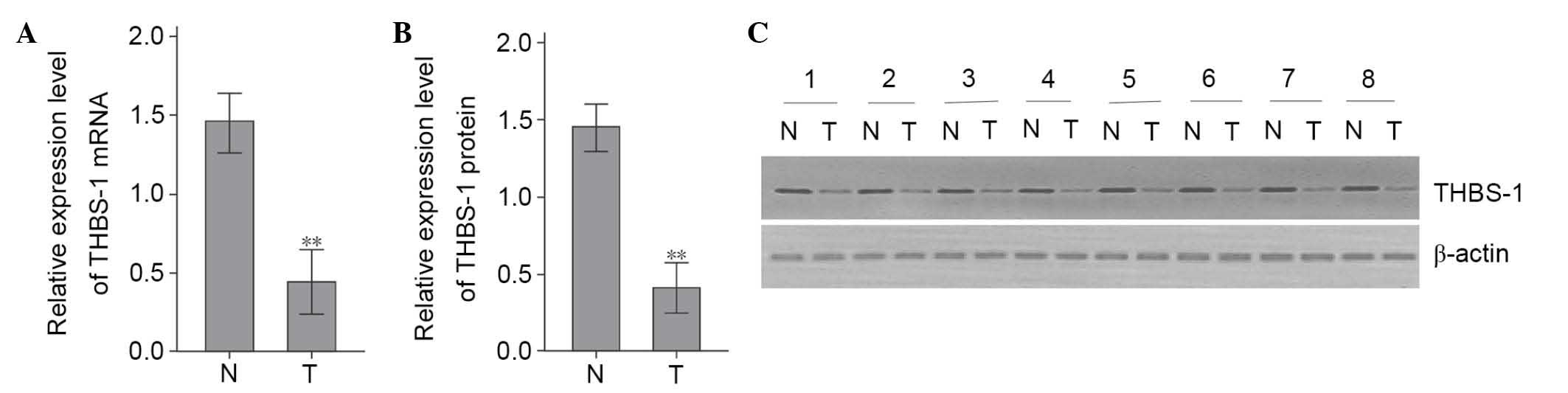
THBS-1 expression was downregulated in
LSCC
Protein and mRNA expression levels of THBS-1 were
examined by western blotting and RT-qPCR, respectively, in 66 LSCC
samples and corresponding adjacent non-tumorous tissue. The THBS-1
mRNA expression levels in LSCC tumor tissues were significantly
lower than in corresponding adjacent non-tumorous tissues (t=6.349,
P<0.01; Fig. 1A). To further
verify this alteration, the protein expression levels of THBS-1
were evaluated. Compared with adjacent non-tumorous tissues,
protein levels of THBS-1 were significantly decreased in LSCC tumor
tissues (t=8.420, P<0.01; Fig. 1B
and C).
Association between clinicopathological
features and expression levels of THBS-1 in LSCC
The association between THBS-1 expression in LSCC
and clinicopathological features is presented in Table I. Protein and mRNA expression
levels of THBS-1 were significantly lower in patients with lymph
node metastasis compared with those without (for mRNA levels:
t=5.118, P<0.01; for protein levels: t=4.117, P<0.05). THBS-1
mRNA and protein expression in patients with stage III–IV LSCC were
significantly decreased compared with those in patients with stage
I–II LSCC (for mRNA levels: t=5.417, P<0.01; for protein levels:
t=5.136, P<0.01). The expression of THBS-1 was not associated
with age, gender, primary site of tumor or histological
differentiation of LSCC patients (P>0.05).
 | Table IAssociations between THBS-1 expression
and clinicopathological features of LSCC patients. |
Table I
Associations between THBS-1 expression
and clinicopathological features of LSCC patients.
| Clinicopathological
characteristic | n | THBS-1 mRNA | P-value | THBS-1 protein | P-value |
|---|
| Age (years) |
| <60 | 23 | 0.411±0.157 | 0.181a | 0.398±0.121 | 0.341a |
| ≥60 | 43 | 0.397±0.191 | | 0.401±0.141 | |
| Gender |
| Male | 54 | 0.399±0.182 | 0.143a | 0.395±0.119 | 0.105a |
| Female | 12 | 0.409±0.166 | | 0.404±0.143 | |
| Classification |
| Supraglottic
LSCC | 34 | 0.399±0.171 | 0.179b | 0.389±0.124 | 0.227b |
| Glottic LSCC | 24 | 0.410±0.177 | | 0.408±0.137 | |
| Subglottic LSCC | 8 | 0.403±0.174 | | 0.402±0.132 | |
| Differentiation |
| High | 37 | 0.409±0.184 | 0.156b | 0.386±0.131 | 0.209b |
| Moderate | 20 | 0.404±0.164 | | 0.406±0.138 | |
| Poor | 9 | 0.399±0.174 | | 0.407±0.124 | |
| Lymph node
metastasis |
| Negative | 42 | 0.695±0.178 | 0.008a | 0.573±0.135 | 0.012a |
| Positive | 24 | 0.113±0.174 | | 0.226±0.129 | |
| TNM stage |
| I–II | 36 | 0.701±0.164 | 0.003a | 0.584±0.139 | 0.008a |
| III–IV | 30 | 0.107±0.184 | | 0.215±0.125 | |
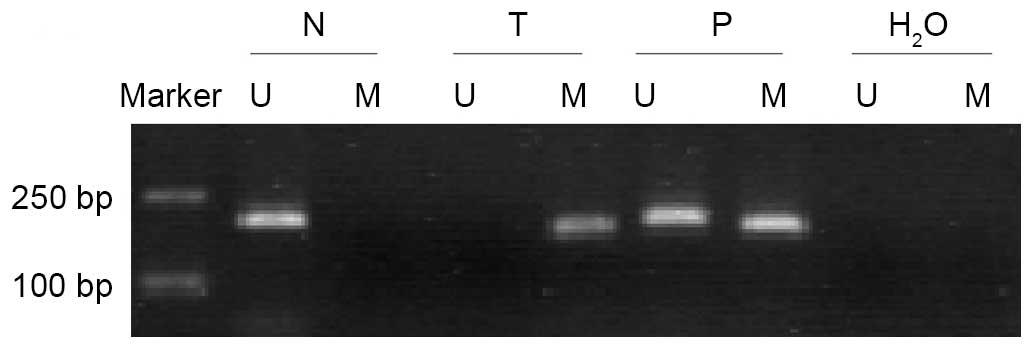
Aberrantly methylated THBS-1 was present
in LSCC
The methylation status of THBS-1 was analyzed in 66
LSCC samples and corresponding adjacent non-tumorous tissues
(Fig. 2). Of the LSCC samples, 32
of 66 (48.50%) exhibited THBS-1 methylation, while only 4 of 66
(6.06%) of paired adjacent non-tumorous tissues were demonstrated
to exhibit THBS-1 methylation. Methylation frequency of THBS-1 in
tumor tissues was significantly higher than in paired adjacent
non-tumorous tissues (χ2=29.90, P<0.001).
Downregulation of THBS-1 is associated
with aberrant methylation in LSCC
The present study investigated the association
between methylation status and expression levels of THBS-1 in LSCC.
Of the 32 methylated LSCC samples, it was observed that mRNA and
protein expression levels of THBS-1 were 0.164±0.067 and
0.098±0.440, respectively. In the remaining 34 unmethylated LSCC,
mRNA and protein levels of THBS-1 were 0.618±0.081 and 0.584±0.079,
respectively. The THBS-1 mRNA and protein levels in methylated
samples were significantly downregulated compared to those in the
unmethylated samples (mRNA: t=10.480, P<0.001; protein: t=9.990,
P<0.001, Table II).
| Table IIAssociation between THBS-1 expression
levels and DNA methylation status in laryngeal squamous cell cancer
tissues. |
Table II
Association between THBS-1 expression
levels and DNA methylation status in laryngeal squamous cell cancer
tissues.
| DNA methylation
status | n | THBS-1 mRNA | P-value | THBS-1 protein | P-value |
|---|
| Methylated | 32 | 0.164±0.067 | P<0.001 | 0.098±0.440 | P<0.001 |
| Unmethylated | 34 | 0.618±0.081 | | 0.584±0.079 | |
Association between clinicopathological
features and methylation status of THBS-1 in LSCC
The association between methylation frequency of
THBS-1 and clinicopathological features of LSCC was examined
(Table III). Methylation
frequency of THBS-1 in patients with lymph node metastasis was
significantly higher than that in patients without lymph node
metastasis (χ2=7.542, P<0.01). When stratified for
TNM stages, frequencies of THBS-1 methylation of patients with
stage III or IV cancer were significantly higher than patients with
stage I or II cancer (χ2=4.855, P<0.05). No other
significant associations were observed between the methylation
status of THBS-1 and the clinicopathological findings, including
age, gender, primary site and histological differentiation
(P>0.05).
| Table IIIAssociations between
clinicopathological features and methylation status of THBS-1 in
LSCC patients. |
Table III
Associations between
clinicopathological features and methylation status of THBS-1 in
LSCC patients.
| Clinicopathological
characteristic | Patients, n | THBS-1 methylation
status, n (%)
| P-value |
|---|
| M | U |
|---|
| Age (years) |
| <60 | 23 | 12 (52.17) | 11 (47.83) | 0.661 |
| ≥60 | 43 | 20 (46.51) | 23 (53.49) | |
| Gender |
| Male | 54 | 26 (48.15) | 28 (51.85) | 0.908 |
| Female | 12 | 6 (50.00) | 6 (50.00) | |
| Classification |
| Supraglottic
LSCC | 34 | 15 (44.11) | 19 (55.88) | 0.634 |
| Glottic LSCC | 24 | 12 (50.00) | 12 (50.00) | |
| Subglottic
LSCC | 8 | 5 (62.50) | 3 (37.5) | |
|
Differentiation |
| High | 37 | 19 (51.35) | 18 (48.65) | 0.870 |
| Moderate | 20 | 9 (45.00) | 11 (55.00) | |
| Poor | 9 | 4 (44.44) | 5 (55.56) | |
| Lymph node
metastasis |
| Negative | 42 | 15 (35.71) | 27 (64.29) | 0.006 |
| Positive | 24 | 17 (70.83) | 7 (29.17) | |
| TNM stage |
| I–II | 36 | 13 (36.11) | 23 (63.89) | 0.028 |
| III–I | 30 | 19 (63.33) | 11 (36.67) | |
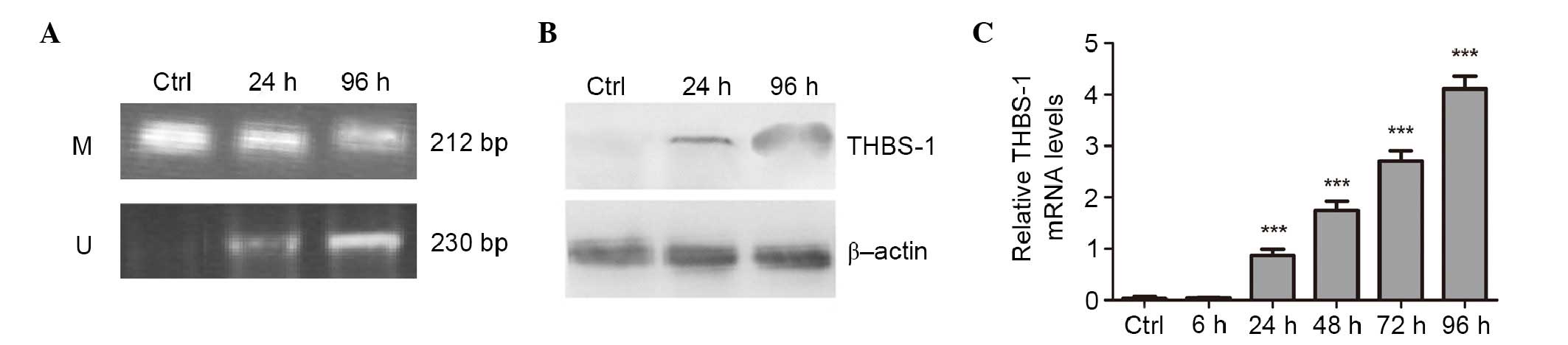
Reactivation of THBS-1 expression
following treatment with 5-aza-dC
To confirm that aberrant methylation was responsible
for silencing THBS-1 expression, Hep-2 cells were treated with 1
µM of demethylating agent 5-aza-dC for 6, 24, 48, 72, and 96
h, respectively. The methylation status of THBS-1 and the effect of
aberrant methylation of THBS-1 on THBS-1 mRNA and protein
expression were evaluated (Fig.
3). In Hep-2 cells, complete methylation of THBS-1 was observed
in control cells, however, following 5-Aza-dC treatment for 96 h,
the methylation status of THBS-1 changed from methylated to a
partially methylated state (Fig.
3A).
The current study also investigated whether the
change in methylation status of THBS-1 was associated with change
in expression of THBS-1 following 5-aza-dC treatment. It was
observed that the expression level of THBS-1 mRNA was increased by
5-aza-dC in a time-dependent manner (0.867±0.129, 1.747±0.170,
2.703±0.207 and 4.110±0.250, at 24, 48, 72 and 96 h, respectively)
compared with those in the control group and 6 h (P<0.001;
Fig. 3C). Increased THBS-1 protein
levels were identified following 5-aza-dC treatment using western
blotting (P<0.05; Fig. 3B).
These results suggest the aberrant methylation of THBS-1 suppresses
THBS-1 mRNA and protein expression in Hep-2 human laryngeal
carcinoma cell line.
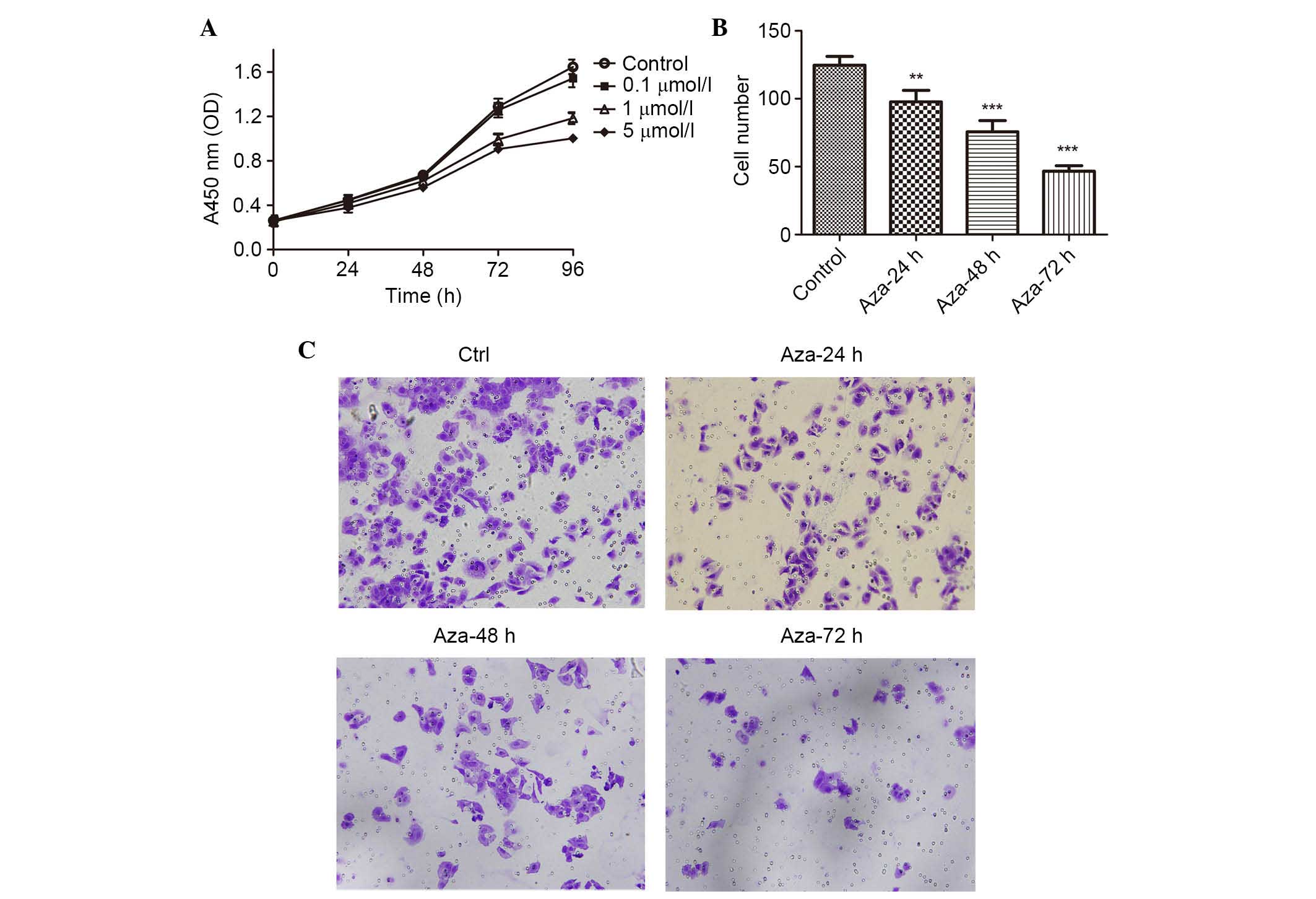
Reactivation of THBS-1 expression with
5-aza-dC inhibits the viability and invasion of Hep-2 cell
To further examine whether the reactivation of
THBS-1 expression can regulate LSCC proliferation and invasion, the
present study analyzed the viability and invasive capability of
Hep-2 cells using the CCK-8 and invasion assay. As presented in
Fig. 4A and Table IV, a concentration- and time-
dependent growth inhibition of cell proliferation was observed in
the Hep-2 cells (all P<0.05). The invasion assay demonstrated
that the number of invading Hep-2 cells was 124.67±6.51,
97.67±8.50, 75.67±8.33, and 46.67±4.04, in the control, 5-aza-dC
24, 48 and 72 h groups, respectively. The number of invading Hep-2
cells was significantly reduced following 5-aza-dC treatment when
compared with the control group (P<0.01 for 24 h 5-aza-dC
treatment; P<0.001 for 48 and 7 h 5-aza-dC treatment; Fig. 4B and C).
| Table IVInhibitory rate (%) for different
concentrations of 5-aza-dC at different time points in Hep-2
cells. |
Table IV
Inhibitory rate (%) for different
concentrations of 5-aza-dC at different time points in Hep-2
cells.
| 5-Aza-dC
concentration | 24 h | 48 h | 72 h | 96 h |
|---|
| 0 µmol/l
(control) | – | – | – | – |
| 0.1
µmol/l | 0.69±0.51a,b | 2.22±0.73a,b | 2.37±0.35a,b | 6.28±1.43a,b |
| 1
µmol/l | 7.20±0.94a,b | 7.90±1.62a,b | 22.91±2.59a,b | 28.07±0.23a,b |
| 5
µmol/l | 16.0±2.07a,b | 16.5±1.82a,b | 28.75±2.61a,b | 39.1±1.83a,b |
Discussion
Thrombospondins are a family of homologous proteins
involved in the regulation of cellular phenotype and extracellular
structure during tissue genesis and remodeling (14). The first to be identified was
THBS-1, it has been demonstrated to modulate progression and
metastasis of tumors (15).
However, the role of THBS-1 in tumor progression and metastasis
remains controversial and presents stimulatory and inhibitory
effects (6–9). The present study demonstrated for the
first time, to the best of our knowledge, that the levels of THBS-1
mRNA and protein expression were significantly decreased in LSCC
tissues compared with adjacent non-tumorous tissues, and were
negatively correlated with lymph node metastasis and advanced
clinical stage. These observations are consistent with the results
from previous studies on non-small cell lung cancer (16), cutaneous squamous cell carcinoma
(17), and melanoma (11), and suggested that THBS-1 acts as a
tumor suppressor in LSCC, which can inhibit the development and
metastasis of LSCC.
DNA methylation on the gene promoter region, which
often results in the suppression of transcription, is considered to
be an underlying mechanism of tumor suppressor gene inactivation
(18–19). Aberrant THBS-1 methylation has been
reported in other types of cancer (10–12).
To elucidate whether decreased expression of THBS-1 in LSCC is a
result of DNA methylation, the methylation status of THBS-1 in LSCC
was investigated and the association between methylation status and
expression levels of THBS-1 in LSCC was determined. The results
demonstrated that the THBS-1 gene was identified to exhibit a more
frequent methylation rate in LSCC compared with adjacent
non-tumorous tissues, which may indicate that aberrant THBS-1
methylation is important in the development of LSCC. Furthermore,
the THBS-1 expression levels in methylated LSCC tissues were
significantly lower than those in the unmethylated LSCC tissues.
These data suggested a potential association between THBS-1
methylation and loss of THBS-1 expression. To further confirm that
aberrant DNA methylation results in inhibition of THBS-1
expression, the effect of the demethylation by 5-aza-dC on THBS-1
gene methylation and THBS-1 re-expression was examined in the Hep-2
cell line. The results indicated that the unmethylated status of
THBS-1 increased with increasing THBS-1 mRNA and protein expression
levels. Thus, methylation of THBS-1 gene directly induced THBS-1
inactivation.
A number of previous studies have considered THBS-1
methylation may be associated with clinicopathological features of
tumors (12,20–22).
However, to the best of our knowledge, there has been no study
evaluating the association between the methylation status of THBS-1
and clinicopathological characteristics of LSCC. The present study
demonstrated that THBS-1 hypermethylation was associated with lymph
node metastasis and TNM stage of LSCC, however, not associated with
age, gender, primary site and histological differentiation. These
significant associations suggested a functional role for THBS-1
gene methylation in invasion and metastasis of LSCC, consistent
with results from gastric cardia adenocarcinoma (12), gastric carcinoma (20), penile squamous cell carcinoma
(21) and meningiomas (22). To provide further evidence on the
association between THBS-1 methylation and progression of LSCC, the
effect of the demethylation reagent 5-aza-dC on the proliferation
and invasion ability of Hep-2 cells was investigated. The results
from the current study suggested that the suppression of
proliferation and invasive ability due to 5-aza-dC may result from
DNA demethylation and reactivation of THBS-1. These findings were
consistent with previous results that indicated increased
expression of THBS-1 decreased angiogenesis, tumor growth, and
metastasis in melanoma (11) and
human neuroblastoma (23).
Although the influence of other possible methylation-silenced tumor
suppressor genes cannot be eliminated, the findings of the present
study suggest that hypermethylation status of THBS-1 may result in
decreased THBS-1 and accelerated LSCC progression and invasion.
5-Aza-dC is a strong inducer of DNA demethylation
that acts by binding methyltransferase enzymes, resulting in the
reactivation of the corresponding genes silenced by DNA methylation
(24). Previously, 5-aza-dC has
been demonstrated to synergize with progesterone therapy to inhibit
endometrial cancer cell growth and invasion (25). In Hep-2 cells, 5-aza-dC
significantly inhibited tumor cell proliferation and invasion.
Although the focus of the present study was THBS-1, these results
provide an additional rationale for investigating 5-aza-dC in the
treatment of LSCC.
There are also a number of limitations in the
present study. Due to contaminations in the available stocks of
Hep-2 cells, it is likely that the cells used in the current study
are HeLa contaminants. However, this may suggest that the
demethylation of THBS-1 in human cervical cancer cell results in
increased THBS-1 expression to inhibit tumor growth and invasion.
Thus, it can be hypothesized that the methylation of THBS-1 is
involved in tumor invasion and metastasis in cervical cancer
patients, which could be further demonstrated in future
studies.
In conclusion, the present study is the first to
determine that THBS-1 is a tumor suppressor gene in LSCC and that
DNA methylation of THBS-1 is an epigenetic event that silences this
gene. Aberrant hypermethylation and reduced expression of THBS-1
promote the invasion of LSCC, which may be a useful biomarker of
tumor progression. The current study also suggested an improved
understanding of DNA methylation may provide a potential
therapeutic target for LSCC. Further research is required to
elucidate the tumor-suppressive mechanism of THBS-1 in LSCC.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant no. 81402606).
References
|
1
|
Choong N and Vokes E: Expanding role of
the medical oncologist in the management of head and neck cancer.
CA Cancer J Clin. 58:32–53. 2008. View Article : Google Scholar
|
|
2
|
Belcher R, Hayes K, Fedewa S and Chen AY:
Current treatment of head and neck squamous cell cancer. J Surg
Oncol. 110:551–574. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shivapurkar N and Gazdar AF: DNA
methylation based biomarkers in non-invasive cancer screening. Curr
Mol Med. 10:123–132. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Heyn H and Esteller M: DNA methylation
profiling in the clinic: Applications and challenges. Nat Rev
Genet. 13:679–692. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Silverstein RL and Febbraio M:
CD36-TSP-HRGP interactions in the regulation of angiogenesis. Curr
Pharm Des. 13:3559–3567. 2007. View Article : Google Scholar
|
|
6
|
Kazerounian S, Yee KO and Lawler J:
Thrombospondins in cancer. Cell Mol Life Sci. 65:700–712. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yee KO, Connolly CM, Duquette M,
Kazerounian S, Washington R and Lawler J: The effect of
thrombospondin-1 on breast cancer metastasis. Breast Cancer Res
Treat. 114:85–96. 2009. View Article : Google Scholar :
|
|
8
|
Lin XD, Chen SQ, Qi YL, Zhu JW, Tang Y and
Lin JY: Overexpression of thrombospondin-1 in stromal
myofibroblasts is associated with tumor growth and nodal metastasis
in gastric carcinoma. J Surg Oncol. 106:94–100. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kasper HU, Ebert M, Malfertheiner P,
Roessner A, Kirkpatrick CJ and Wolf HK: Expression of
thrombospondin-1 in pancreatic carcinoma: Correlation with
microvessel density. Virchows Arch. 438:116–120. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rojas A, Meherem S, Kim YH, Washington MK,
Willis JE, Markowitz SD and Grady WM: The aberrant methylation of
TSP1 suppresses TGF-beta1 activation in colorectal cancer. Int J
Cancer. 123:14–21. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lindner DJ, Wu Y, Haney R, Jacobs BS,
Fruehauf JP, Tuthill R and Borden EC: Thrombospondin-1 expression
in melanoma is blocked by methylation and targeted reversal by
5-Aza-deoxycytidine suppresses angiogenesis. Matrix Biol.
32:123–132. 2013. View Article : Google Scholar :
|
|
12
|
Guo W, Dong Z, He M, Guo Y, Guo J, Chen Z,
Yang Z and Kuang G: Aberrant methylation of thrombospondin-1 and
its association with reduced expression in gastric cardia
adenocarcinoma. J Biomed and Biotechnol. 2010:7214852010.
View Article : Google Scholar
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
14
|
Chen H, Herndon ME and Lawler J: The cell
biology of thrombos-pondin-1. Matrix Biol. 19:597–614. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Baenziger NL, Brodie GN and Majerus PW: A
thrombin-sensitive protein of human platelet membranes. Proc Natl
Acad Sci USA. 68:240–243. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fleitas T, Martínez-Sales V, Vila V,
Reganon E, Mesado D, Martín M, Gómez-Codina J, Montalar J and
Reynés G: VEGF and TSP1 levels correlate with prognosis in advanced
non-small cell lung cancer. Clin Transl Oncol. 15:897–902. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Streit M, Velasco P, Brown LF, Skobe M,
Richard L, Riccardi L, Lawler J and Detmar M: Overexpression of
thrombospondin-1 decrease angiogenesis and inhibits the growth of
human cutaneous squamous cell carcinomas. Am J Pathol. 155:441–452.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang C, Li H, Wang Y, Liu W, Zhang Q,
Zhang T, Zhang X, Han B and Zhou G: Epigenetic inactivation of the
tumor suppressor gene RIZI in hepatocellular carcinoma involves
both DNA methylation and histone modifications. J Hepatol.
53:889–895. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cherrier T1, Suzanne S, Redel L, Calao M,
Marban C, Samah B, Mukerjee R, Schwartz C, Gras G, Sawaya BE, et
al: p21 (WAF1) gene promoter is epigenetically slienced by CTIP2
and SUV39H1. Oncogene. 28:3380–3389. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Oue N, Matsumura S, Nakayama H, Kitadai Y,
Taniyama K, Matsusaki K and Yasui W: Reduced expression of the TSP1
gene and its association with promoter hypermethylation in gastric
carcinoma. Oncology. 64:423–429. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Guerrero D, Guarch R, Ojer A, Casas JM,
Ropero S, Mancha A, Pesce C, Lloveras B, Garcia-Bragado F and Puras
A: Hypermethylation of the thrombospondin-1 gene is associated with
poor prognosis in penile squamous cell carcinoma. BJU Int.
102:747–755. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gong J, Zhu SG, Wu CY, Li XG, Liu YG, Ren
XH and Zhang Y: Aberrant methylation of NF2, TIMP-3 and THBS1 genes
and their diagnostic values in meningiomas. Zhonghua Yi Xue Za Zhi.
92:2889–2892. 2012.In Chinese.
|
|
23
|
Yang QW, Liu S, Tian Y, Salwen HR,
Chlenski A, Weinstein J and Cohn SL: Methylation-associated
silencing of the thrombospondin-1 gene in human neuroblastoma.
Cancer Res. 63:6299–6310. 2003.PubMed/NCBI
|
|
24
|
Meng CF, Zhu XJ, Peng G and Dai DQ: Role
of histone modifications and DNA methylation in the regulation of
O6-methylguanine-DNA methyltransferase gene expression in human
stomach cancer cells. Cancer Invest. 28:331–339. 2010. View Article : Google Scholar
|
|
25
|
Hu Q, Yu L, Chen R, Wang YL, Ji L, Zhang
Y, Xie Y and Liao QP: 5-aza-2′-deoxycytidine improves the
sensitivity of endometrial cancer cells to progesterone therapy.
Int J Gynecol Cancer. 22:951–959. 2012. View Article : Google Scholar : PubMed/NCBI
|