Introduction
Hepatocellular carcinoma (HCC) has been deemed the
fifth most common type of malignant tumor. HCC has the third
highest cancer-related mortality rate and a previous study have
estimated that 500,000–1,000,000 novel cases of HCC are diagnosed
annually (1). Invasion, metastasis
and postoperative recurrence are predominant causes of mortality in
patients with HCC and are the primary factors that affect clinical
treatment and prognosis. Therefore, due to aggressive metastasis
and invasion, HCC is frequently diagnosed at an advanced stage in
patients and has a high lethality (2). A previous study demonstrated that
aberrant activation of epithelial mesenchymal transition (EMT) is
important for the process of tumor cell invasion and metastasis.
EMT triggers multiple biochemical changes in normal hepatic
epithelial cells, which ultimately enable them to assume a
mesenchymal cell phenotype. This leads to enhanced migratory
capacity, invasiveness, elevated resistance to apoptosis and
increased production of ECM components (3). EMT is involved in multiple molecular
mechanisms and signal transduction pathways. Their main function
being the downregulation of the cell adhesion molecule E-cadherin
and the upregulation of the mesenchymal molecule vimentin. In
addition, a number of transcriptional factors have been identified
as crucial for the induction of EMT, including Snail homologues,
basic helix-loop-helix and zinc-finger E-box-binding homeobox (ZEB)
(4). The ZEB family (consisting of
ZEB1 and ZEB2) has a consistent inverse correlation with E-cadherin
and vimentin in numerous types of cancer (5).
Long non-coding RNAs (lncRNAs) are a type of RNA
molecule that are over 200 nucleotides in length and do not have
protein-coding potential (6). They
have been identified to affect multiple biological regulatory
processes, including development, differentiation and
carcinogenesis (7,8). A previous study suggested that the
dysregulation of lncRNAs may be associated with the development and
metastasis of various types of cancer (9). Antisense long non-coding RNAs
(aslncRNAs) are a type of lncRNAs that are oriented in antisense
direction with respect to a protein coding locus. They are
transcribed frequently in human cells and 61–72% of all transcribed
regions possess aslncRNAs (10).
Additionally, aslncRNAs have been associated with tumor recurrence,
metastasis and prognosis in various types of cancer.
A recent study by Li et al (11) identified the ZEB 1 antisense RNA 1
(ZEB1-AS1) gene as a non-coding oncogene involved in the
tumorigenesis of HCC (11). It was
determined that higher expression levels of lncRNA ZEB1-AS1 were
observed in HCC tissues, compared with the adjacent normal tissues.
Li et al (11) also
described the effects of the ZEB1-AS1 lncRNA on cell proliferation,
migration and invasion, and cell cycle regulation. In addition, it
was determined that ZEB1-AS1 may control ZEB1 expression levels and
promote tumor growth and metastasis in patients with HCC. Sequence
analysis was performed and revealed that there was also a
non-coding antisense transcript in the promoter sequence of ZEB2
(12). The function of ZEB2
antisense RNA 1 (ZEB2-AS1) remains to be elucidated. Therefore, the
present study investigated whether lncRNA ZEB2-AS1 contributes to
HCC development and how it may alter cell growth and metastasis by
regulating ZEB2, vimentin, fibronectin, E-cadherin and N-cadherin
expression levels. Additionally, the clinical characteristics of
patients that may influence the expression levels of ZEB2-AS1 and
provide data for the prognosis and targeted therapy of HCC were
determined.
Materials and methods
Patient and tumor samples
HCC and adjacent normal tissues were collected
following curative surgical resection from 40 patients with HCC in
the Zhongnan Hospital of Wuhan University (Wuhan, China) between
May 2010 and May 2012. The present study was approved by the ethics
committee of the Zhongnan Hospital of Wuhan University and written
informed consent was obtained from all patients. Curative resection
was defined as the removal of all recognizable tumor tissue with a
clear microscopic margin. Patients were not treated using any
preoperative therapy, such as transcatheter arterial
chemoembolization, radiofrequency ablation or percutaneous ethanol
injection. The collected tissue samples were immediately frozen in
liquid nitrogen and stored at −80°C until they were used for RNA
extraction. Tumor tissues were divided into low-expression (n=20)
and high expression (n=20) groups based on the mean expression
level of all tumor tissues (n=40). Using the patient medical
documents, a 48-month follow-up survival survey was conducted.
Cell lines and culture
The Hep3B, HepG2, Huh-7 and SMMC-7721 HCC cell lines
were obtained from the American Type Culture Collection (Manassas,
VA, USA). The HCCLM9 cell line was obtained from the Liver Cancer
Institute, Fudan University (Shanghai, China) (13). All cells were cultured in RPMI-1640
medium, which was supplemented with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and 1%
penicillin and streptomycin in a humidified atmosphere of 5% CO2 at
37°C.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from HCC and normal adjacent
tissues, and human HCC cell lines using TRIzol reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. RNA was converted into cDNA (124 µg/20 µl) using a Takara
RNA PCR kit (Takara Biotechnology Co., Ltd., Dalian, China). qPCR
was performed using a iQ SYBR-Green Supermix (Bio-Rad Laboratories,
Inc., Hercules, CA, USA) with 124 ng/2 µl cDNA. All reactions were
performed in triplicate on iCycler IQ multi-color Detection System
(Bio-Rad Laboratories, Inc.). The thermocycling conditions were as
follows: 95°C for 5 min; 40 cycles of denaturation at 95°C for 30
sec; annealing at 60°C for 30 sec and extension at 72°C for 20 sec.
The following primers were used: ZEB2-AS1 forward (F)
5′-ATGAAGAAGCCGCGAAGTGT-3′ and reverse (R)
5′-CACACCCTAATACACATGCCCT-3′; and β-actin F
5′-AGTTGCGTTACACCCTTTCTTGAC-3′ and R 5′-GCTCGCTCCAACCGACTGC-3′. The
comparative cycle quantification (Cq) method (14) was used to quantify the relative
expression levels of mRNA. The relative amount was calculated using
the equation 2−ΔΔCq where ΔCq = (Cq of interest mRNA -
Cq of β-actin). All the qPCR reactions were performed in
duplicate.
ZEB2-AS1 small interfering RNAs
(siRNAs) and transfection
The siRNA-ZEB2-AS1 sequence used was
5′-CAAAGGACACCTTTGGTTACCTGAA-3′. Synthetic sequence-scrambled siRNA
(Santa Cruz Biotechnology Inc., Dallas, TX, USA) was used as a
negative control (siRNA-NC). The siRNAs were transfected into Huh-7
cells during the logarithmic growth phase using Lipofectamine 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. Cells were incubated with the siRNA
transfection complex for 24 h. Next, cells were harvested in order
to assess mRNA and protein expression level changes using RT-qPCR
and western blot analysis at 24 h. For the conventional PCR, the
relative mRNA expression level was represented as the grayscale
ratio of the mRNA of interest to β-actin.
Western blot analysis
Proteins from HCC human cell lines were extracted
using a radioimmunoprecipitation lysis buffer (Beyotime Institute
of Biotechnology, Shanghai, China). The proteins were quantified by
the bicinchoninic acid method and samples (7530 µg/ml) were then
separated on 10% sodium dodecyl sulfate-polyacrylamide gel and
transferred onto polyvinylidene fluoride membranes. The membranes
were blocked with 5% skimmed milk powder in Tris-buffered saline
and Tween 20 (TBST; 10 mM Tris-HCl, pH 7.5, 150 mM NaCl and 0.1%
Tween-20) at room temperature for 1 h. The following primary
antibodies were added and membranes were incubated overnight at
4°C: Mouse monoclonal anti-ZEB2 antibody (1:200; Santa Cruz
Biotechnology, Inc.; cat. no. sc-271984), mouse monoclonal
anti-vimentin (1:200; Santa Cruz Biotechnology, Inc.; cat. no.
sc-6260), mouse monoclonal E-cadherin (1:500; Abcam, Cambridge, UK;
cat. no. ab1416), rabbit polyclonal anti-N-cadherin (1:1,000;
Abcam; cat. no. ab18203) and mouse monoclonal anti-fibronectin
(1:1,000; Abcam; cat. no. ab6328). After washing with TBST, goat
anti-mouse (cat. no. SA00001-1) and goat anti-rabbit (SA00001-2)
horseradish peroxidase-conjugated secondary antibody (1:2,000;
Proteintech Group, Inc., Chicago, IL, USA) were added and the
membrane was incubated at room temperature for 1 h. The proteins
were visualized using autoradiography with an enhanced
chemiluminescence reagent (Wuhan Boster Biological Technology,
Ltd., Wuhan, China). The relative protein expression levels were
determined by using the grayscale ratio of the protein of interest
to glyceraldehyde 3-phosphate dehydrogenase and the results were
analyzed using GraphPad Prism version 5.0 software (GraphPad
Software, Inc., La Jolla, CA, USA).
In vitro cell proliferation assay
Huh-7 cells were seeded at a density of 8.0×103
cells/well in a 96-well plate and incubated for 18 h. Water-soluble
tetrazolium salts-8 (10 µl) from the Cell Counting Kit-8 (Wuhan
Boster Biological Technology, Ltd., Wuhan, China) was then added to
each well. The plates were incubated at 37°C in 5% CO2 for 12, 24,
48, 72 and 96 h, and the absorbance of each sample was determined
at a wavelength of 450 nm using a microplate reader.
Bromodeoxyuridine (BrdU) labeling
assay
The Huh-7 cells (1×106/cm2)
were seeded on a slide and incubated in RPMI-1640 with 10% FBS with
or without siRNA-ZEB1 transfection for 24 h. BrdU (10 µM;
Sigma-Aldrich; Merck Millipore, Darmstadt, Germany) was added to
the culture for 1 h. Cells were then fixed with 4% paraformaldehyde
and exposed to ice-cold methanol for 2 min. Cells were incubated in
2 M HCl for 1 h at room temperature and then incubated in 0.1 M
sodium borate for 2 min. Finally, immunocytochemistry for BrdU
labeling of proliferative Huh-7 cells was performed. Nuclei were
stained with 1 µg DAPI solution for 5 min. Images were captured
using an epifluorescence microscope (Zeiss AG, Oberkochen, Germany)
with a charge-coupled device camera and AxioVision software SE64
4.9.1 (Zeiss AG).
In vitro invasion assay
In order to perform the invasion assay, Transwell
chambers precoated with Matrigel (BD Bioscience, San Jose, CA, USA)
were used. Huh-7 cells were cultured in serum-free medium (GE
Healthcare Life Sciences, Logan, UT, USA), where the upper chambers
of a Transwell plate (Corning, Corning, NY, USA) were separated
from the lower chambers with a permeable 8 µm polycarbonate
membrane. Medium with 10% FBS was added into the lower chambers as
a chemoattractant. Cells were incubated for 24 h, cells remaining
on the upper membrane were carefully removed, those that had
invaded through the membrane were fixed in 75% ethanol and stained
with crystal violet. The number of stained cells was counted under
a light microscope from five different visual fields of each
filter. Three independent experiments were performed.
In vitro migration assay
The Huh-7 cells were seeded at a density of 1.0×106
cells/well in 6-well plates. Confluent cell monolayers were
disrupted by standardized wound scratching using a sterile 10 µl
pipette tip and incubated in serum-free medium for 60 h. Migration
of cells into the disrupted area and the recovery of the monolayer
was assessed and photographed every 12 h for 60 h using a phase
contrast microscope.
Statistical analysis
Data are presented as the mean ± standard deviation
and analyzed using GraphPad Prism 5.0 software (GraphPad Software,
Inc.). Continuous variables were compared using a Kruskal-Wallis
test and categorical data were compared using χ2 or
Fisher exact tests. The Kaplan-Meier method was used for survival
analysis and differences in survival were determined using the
log-rank test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Expression level of ZEB2-AS1 in HCC
tissues
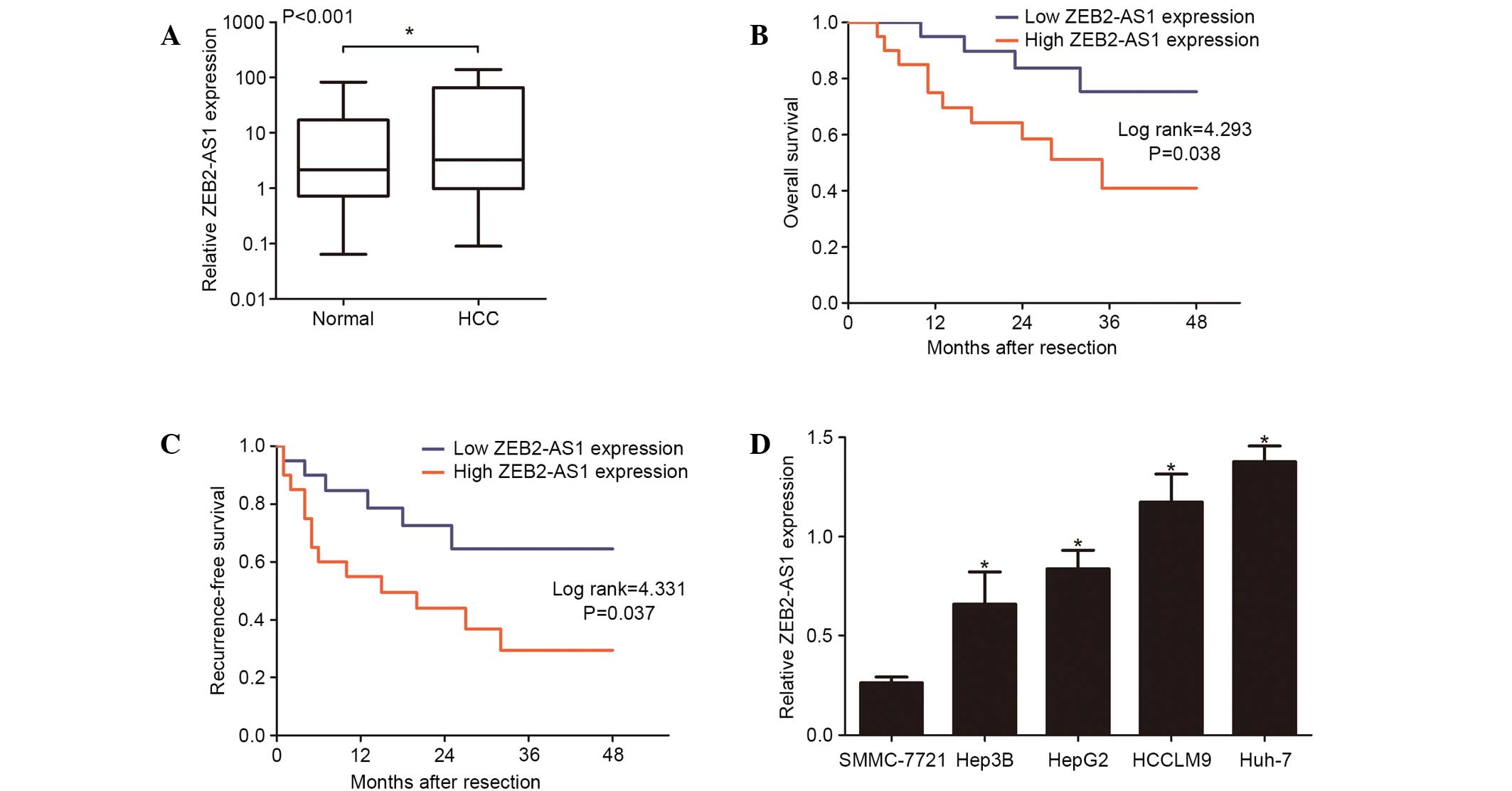
In order to determine whether there was a difference
in the expression level of lncRNA ZEB2-AS1 in HCC tissues and the
adjacent normal tissues the ZEB2-AS1 expression levels in 40 paired
tissue samples were detected using RT-qPCR. The expression of
lncRNA ZEB2-AS1 was significantly upregulated in HCC tissues when
compared with adjacent normal tissues (P<0.001; Fig. 1A). Additionally, the correlation
between lncRNA ZEB2-AS1 expression level and clinicopathological
variables of patients with HCC patients was assessed in order to
evaluate its clinical significance. As presented in Table I, ZEB2-AS1 expression levels in HCC
were significantly associated with a primary tumor size of ≥5 cm
(P=0.027), intrahepatic metastasis (P=0.025) and TNM stage
(P=0.011). However, ZEB2-AS1 expression was not correlated with
other clinicopathological variables, including age (P=0.342), liver
cirrhosis (P=0.113) or differentiation (P=0.784). Additionally,
patients with HCC and high ZEB2-AS1 expression were revealed to
have significantly shorter overall survival (P=0.038; Fig. 1B) and significantly higher
recurrence rates (P=0.037; Fig.
1C) compared with those with low expression of ZEB2-AS1. These
findings demonstrate that the upregulation of lncRNA ZEB-AS1 may be
associated with tumor metastasis and poor prognosis in patients
with HCC.
 | Table I.Correlation between ZEB2-AS1
expression and clinicopathological variables of patients with
hepatocellular carcinoma. |
Table I.
Correlation between ZEB2-AS1
expression and clinicopathological variables of patients with
hepatocellular carcinoma.
|
|
| Relative ZEB2-AS1
expression |
|
|---|
|
|
|
|
|
|---|
| Variable | Total (n=40) | High (n=20) | Low (n=20) | P-value |
|---|
| Gender |
|
|
|
|
| Male | 29 | 16 | 13 | 0.479 |
|
Female | 11 | 4 | 7 |
|
| Age (years) |
|
|
|
|
|
<60 | 19 | 11 | 8 | 0.342 |
| ≥60 | 21 | 9 | 12 |
|
| Size of primary tumor
(cm) |
|
|
|
|
|
<5 | 19 | 6 | 13 | 0.027 |
| ≥5 | 21 | 14 | 7 |
|
| PVTT |
|
|
|
|
|
Present | 6 | 4 | 2 | 0.658 |
|
Absent | 34 | 16 | 18 |
|
| Intrahepatic
metastasis |
|
|
|
|
|
Present | 17 | 12 | 5 | 0.025 |
|
Absent | 23 | 8 | 15 |
|
| Serum AFP
(ng/ml) |
|
|
|
|
|
<200 | 15 | 5 | 10 | 0.103 |
| ≥200 | 25 | 15 | 10 |
|
| Liver cirrhosis |
|
|
|
|
|
Present | 19 | 12 | 7 | 0.113 |
|
Absent | 21 | 8 | 13 |
|
| Differentiation |
|
|
|
|
| Well | 7 | 3 | 4 | 0.784 |
|
Moderate | 28 | 15 | 13 |
|
| Poor | 5 | 2 | 3 |
|
| TNM stage |
|
|
|
|
| I–II | 22 | 7 | 15 | 0.011 |
|
III–IV | 18 | 13 | 5 |
|
Expression levels of ZEB2-AS1 in
various human HCC cell lines
RT-qPCR was performed to determine ZEB2-AS1
expression in 5 human HCC cell lines, SMMC-7721, Hep3B, HepG2,
HCCLM9 and Huh-7. The highest expression level of ZEB2-AS1 was
detected in Huh-7 cells when compared with the remaining cell lines
(P<0.05; Fig. 1D). Therefore,
the Huh-7 HCC cell line was selected for the remaining experiments
in the present study.
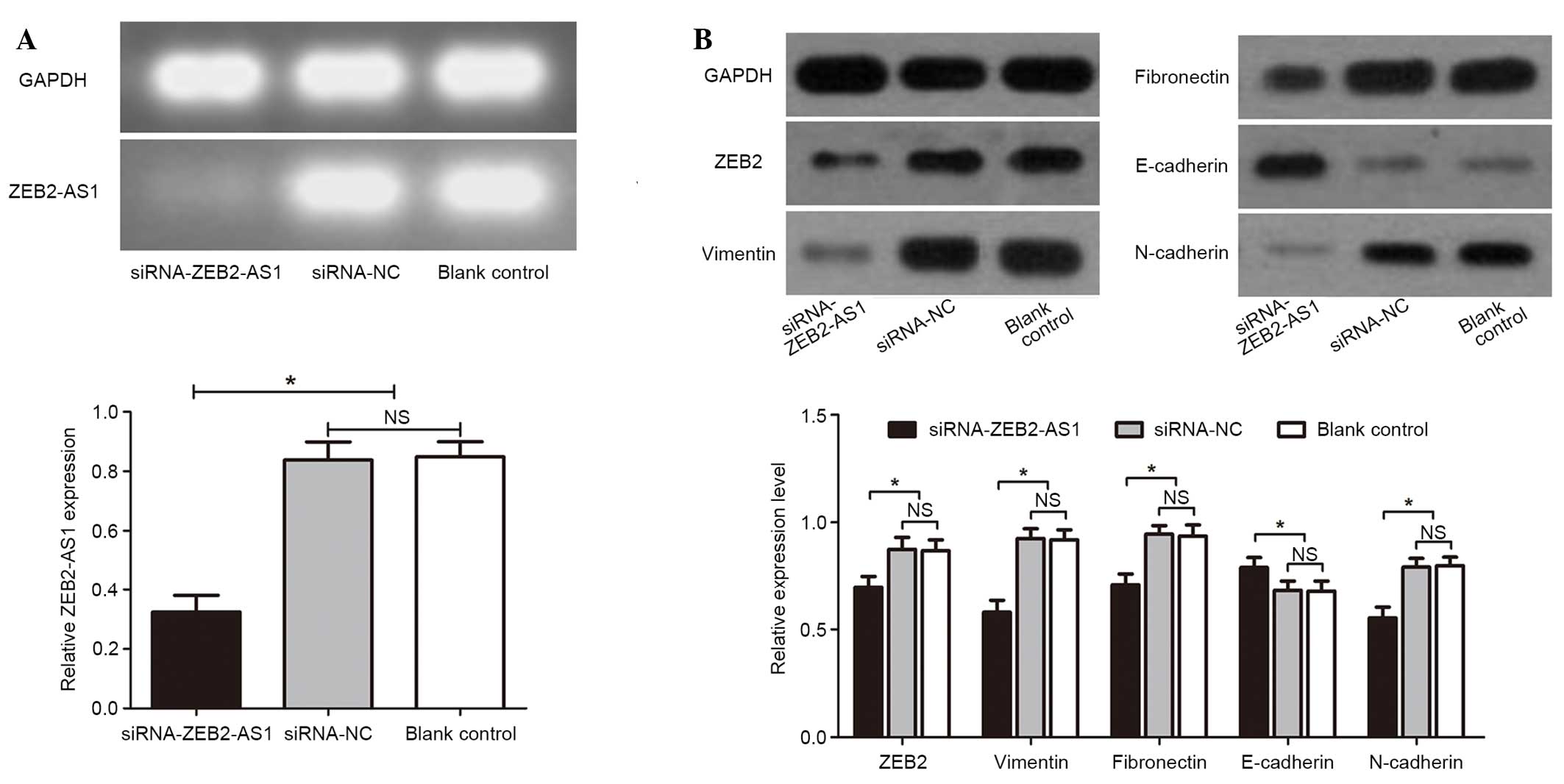
Effects of ZEB2-AS1 siRNAs on the
expression levels of EMT-induced markers
The present study aimed to determine the association
between ZEB2-AS1 expression levels and EMT-induced markers, and
determine the function of ZEB2-AS1 in human HCC cells. Therefore,
siRNAs were used to knock down ZEB2-AS1 in the Huh-7 HCC cell line.
Huh-7 cells were divided into three treatment groups: i)
siRNA-ZEB2-AS1-transfected group; ii) siRNA-NC group; and iii)
blank control group. The knockdown effect of siRNA-ZEB2-AS1 was
confirmed by conventional PCR. This revealed that the expression
levels of ZEB1-AS1 were significantly reduced in the
siRNA-ZEB1-transfected group compared with the siRNA-NC and the
blank control groups (P<0.05; Fig.
2A), indicating the interference ability of siRNA on ZEB2-AS1
was effective and specific. Subsequently, the result of the western
blot analysis demonstrated that the successful knock down of
ZEB2-AS1 may lead to a significant downregulation of ZEB2
(P<0.05; Fig. 2B), vimentin
(P<0.05; Fig. 2B), fibronectin
(P<0.05; Fig. 2B) and
N-cadherin (P<0.05; Fig. 2B),
and an upregulation of E-cadherin (P<0.05; Fig 2B), which suggested that the
expression of EMT-induced markers may be regulated by ZEB2-AS1.
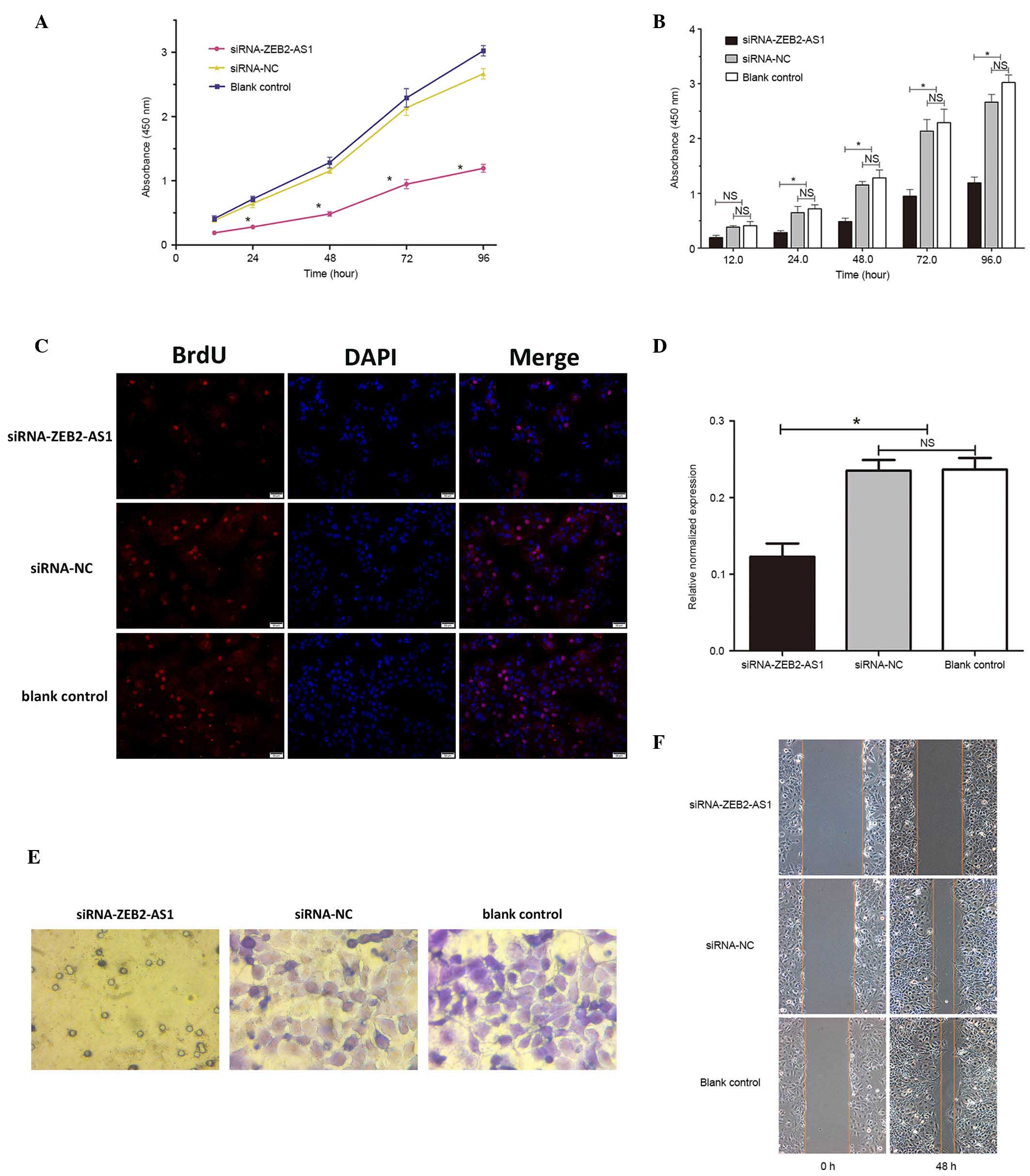
Effects of ZEB2-AS1 siRNAs on cell
proliferation and cell cycle in vitro
CCK-8 assay was used to assess the effect of
ZEB2-AS1 suppression on cell vitality over a period of 96 h. It was
determined that cell viability was significantly reduced in the
siRNA-ZEB2-AS1-transfected group compared with the negative and
blank control groups (P<0.05; Fig.
3A and B). Therefore, it is possible that cell viability was
suppressed by the knockdown of ZEB2-AS1. The BrdU labeling assay
revealed that DNA synthesis was also significantly inhibited in the
siRNA-ZEB2-AS1-transfected group compared with the remaining
control groups (P<0.05; Fig. 3C and
D). These findings reveal that Huh-7 cell proliferation was
inhibited by the silencing of ZEB2-AS1.
Effects of ZEB2-AS1 siRNAs on invasion
and migration of Huh-7 cells in vitro
Cell invasion and migration assays were performed to
determine how ZEB2-AS1 affected these characteristics in Huh-7
cells. The in vitro invasiveness of Huh-7 cells was detected
using a Transwell invasion chamber assay. Crystal violet staining
revealed that the numbers of Huh-7 cells per high power field
(three fields analyzed) in siRNA-ZEB2-AS1, siRNA-NC and blank
control groups were 7.33±2.49, 70.33±3.86 and 66.33±3.68,
respectively, this suggests that the number of Huh-7 cells that had
passed through the polycarbonate membrane of the Transwell invasion
chamber was significantly reduced in the siRNA-ZEB2-AS1-transfected
group compared with the negative and blank control groups
(P<0.05; Fig. 3E). No
significant difference was identified between the invasiveness of
the Huh-7 cells siRNA-NC group and the blank control group
(P>0.05; Fig. 3E). The
wound-healing assays were performed to investigate the migration
ability of Huh-7 cells in the three treatment groups. It was
determined, from three high power fields that the movement of cells
in siRNA-ZEB2-AS1, siRNA-NC and blank control groups were
1.93±0.26, 3.53±0.33 and 3.23±0.21 cm, respectively which
demonstrates that the migration ability of cells the
siRNA-ZEB2-AS1-transfected group was significantly reduced compared
with the two remaining control groups, 48 h after wounding
(Fig. 3F). These findings
demonstrated that the downregulation of ZEB2-AS1 reduced the
migration and invasion abilities of Huh-7 cells.
Discussion
Previous studies have demonstrated that lncRNAs have
important functions in the regulation of gene expression and their
dysregulation may participate in the proliferation and metastasis
of cancer cells (15–17). Notably, aslncRNAs, which are
oriented in antisense direction with respect to a protein coding
loci, usually act as regulators of the gene located on the opposite
strand (18). Previous studies
have determined that aslncRNAs are involved in the complex network
of signaling pathways in cancer cells and may function as enhancers
of tumor-associated gene expression (19,20).
ZEB has been identified as a transcriptional factor
and has been demonstrated to be important for the process of EMT,
which is closely associated with carcinogenesis. Previous studies
have revealed that abnormal expression of ZEB in various types of
cancer has been associated with aggressive disease, low
differentiation, the development of metastases and poor prognosis
in patients (21,22). Li et al (11) reported that the expression levels
of ZEB1-AS1 were upregulated in HCC tissues when compared with
paired adjacent normal tissues. ZEB1-AS1 expression was observed to
increase with HCC progression. Patients with HCC that had high
ZEB1-AS1 expression were identified to have shorter overall
survival and higher recurrence rates compared with those that had
low expression levels of ZEB-AS1. ZEB1-AS1 promoted the
proliferation and metastasis of HCC in vitro and in
vivo; therefore, acting as an oncogene in HCC. Using the Pubmed
gene database, it was determined that there was a non-coding
antisense transcript located from the promoters of ZEB2 (ZEB2-AS1)
(23). The present study
determined that the expression of ZEB2-AS1 in HCC tissues was
significantly higher compared with adjacent normal tissues and this
was significantly associated with the size of the primary tumor
(P=0.027), intrahepatic metastasis (P=0.025) and TNM stage
(P=0.011). Additionally, according to the 48-month follow-up
survival survey the present study demonstrated that patients with
HCC and high ZEB2-AS1 expression levels had shorter overall
survival and higher recurrence rates compared with those with low
expression levels of ZEB2-AS1 (P<0.05). Therefore, ZEB2-AS1
expression levels may serve as a prognostic factor for HCC
pathogenesis. Furthermore, the successful downregulation of
ZEB2-AS1 reduced HCC cell viability significantly and DNA synthesis
was also inhibited. Simultaneously, the invasion and migration
ability of HCC cells was suppressed by the knockdown of the
ZEB2-AS1 gene. Therefore, the current study indicates that ZEB2-AS1
may serve as an oncogene and may be important for HCC development
and progression.
In conclusion, the present study demonstrated that
the downregulation of aslncRNA ZEB2-AS1 was associated with the
reduction of tumor growth and metastasis in HCC tissues. The
importance of ZEB2-AS1 in tumor progression and survival prediction
in HCC patients was elucidated, indicating that ZEB2-AS1 may be a
valuable prognostic biomarker and a novel therapeutic target for
HCC.
Acknowledgements
The present study was supported by the Science and
Technology Project of Wuhan (grant no. 2013060501010153).
References
|
1
|
El-Serag HB and Rudolph KL: Hepatocellular
carcinoma: Epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lau WY and Lai EC: Hepatocellular
carcinoma: Current management and recent advances. Hepatobiliary
Pancreat Dis Int. 7:237–257. 2008.PubMed/NCBI
|
|
3
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Iwatsuki M, Mimori K, Yokobori T, Ishi H,
Beppu T, Nakamori S, Baba H and Mori M: Epithelial-mesenchymal
transition in cancer development and its clinical significance.
Cancer Sci. 101:293–299. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Guo F, Kerrigan BC Parker, Yang D, Hu L,
Shmulevich I, Sood AK, Xue F and Zhang W: Post-transcriptional
regulatory network of epithelial-to-mesenchymal and
mesenchymal-to-epithelial transitions. J Hematol Oncol. 7:192014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shi X, Sun M, Liu H, Yao Y and Song Y:
Long non-coding RNAs: A new frontier in the study of human
diseases. Cancer Lett. 339:159–166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang JL, Zheng L, Hu YW and Wang Q:
Characteristics of long non-coding RNA and its relation to
hepatocellular carcinoma. Carcinogenesis. 35:507–514. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fatica A and Bozzoni I: Long non-coding
RNAs: New players in cell differentiation and development. Nat Rev
Genet. 15:7–21. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He Y, Vogelstein B, Velculescu VE,
Papadopoulos N and Kinzler KW: The antisense transcriptomes of
human cells. Science. 322:1855–1857. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li T, Xie J, Shen C, Cheng D, Shi Y, Wu Z,
Deng X, Chen H, Shen B, Peng C, et al: Upregulation of long
noncoding RNA ZEB1-AS1 promotes tumor metastasis and predicts poor
prognosis in hepatocellular carcinoma. Oncogene. 35:1575–1584.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhuang J, Lu Q, Shen B, Huang X, Shen L,
Zheng X, Huang R, Yan J and Guo H: TGFβ1 secreted by
cancer-associated fibroblasts induces epithelial-mesenchymal
transition of bladder cancer cells through lncRNA-ZEB2NAT. Sci Rep.
5:119242015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li Y, Tang ZY, Ye SL, Liu YK, Chen J, Xue
Q, Chen J, Gao DM and Bao WH: Establishment of cell clones with
different metastatic potential from the metastatic hepatocellular
carcinoma cell line MHCC97. World J Gastroenterol. 7:630–636.
2001.PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tian D, Sun S and Lee JT: The long
noncoding RNA, Jpx, is a molecular switch for X chromosome
inactivation. Cell. 143:390–403. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ørom UA, Derrien T, Beringer M, Gumireddy
K, Gardini A, Bussotti G, Lai F, Zytnicki M, Notredame C, Huang Q,
et al: Long noncoding RNAs with enhancer-like function in human
cells. Cell. 143:46–58. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao T, Xu J, Liu L, Bai J, Xu C, Xiao Y,
Li X and Zhang L: Identification of cancer-related lncRNAs through
integrating genome, regulome and transcriptome features. Mol
Biosyst. 11:126–136. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nolasco S, Bellido J, Gonçalves J, Tavares
A, Zabala JC and Soares H: The expression of tubulin cofactor A
(TBCA) is regulated by a noncoding antisense Tbca RNA during testis
maturation. PloS One. 7:e425362012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lepoivre C, Belhocine M, Bergon A, Griffon
A, Yammine M, Vanhille L, Zacarias-Cabeza J, Garibal MA, Koch F,
Maqbool MA, et al: Divergent transcription is associated with
promoters of transcriptional regulators. BMC Genomics. 14:9142013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kurihara M, Shiraishi A, Satake H and
Kimura AP: A conserved noncoding sequence can function as a
spermatocyte-specific enhancer and a bidirectional promoter for a
ubiquitously expressed gene and a testis-specific long noncoding
RNA. J Mol Biol. 426:3069–3093. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sánchez-Tilló E, de Barrios O, Siles L,
Amendola PG, Darling DS, Cuatrecasas M, Castells A and Postigo A:
ZEB1 Promotes invasiveness of colorectal carcinoma cells through
the opposing regulation of uPA and PAI-1. Clin Cancer Res.
19:1071–1082. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Graham TR, Zhau HE, Odero-Marah VA,
Osunkoya AO, Kimbro KS, Tighiouart M, Liu T, Simons JW and O'Regan
RM: Insulin-like growth factor-I-dependent up-regulation of ZEB1
drives epithelial-to-mesenchymal transition in human prostate
cancer cells. Cancer Res. 68:2479–2488. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Beltran M, Puig I, Peña C, García JM,
Alvarez AB, Peña R, Bonilla F and de Herreros AG: A natural
antisense transcript regulates Zeb2/Sip1 gene expression during
Snail1-induced epithelial-mesenchymal transition. Genes Dev.
22:756–769. 2008. View Article : Google Scholar : PubMed/NCBI
|