Introduction
Atherosclerosis (AS) is characterised by autoimmune
and immunological mechanisms leading to plaque formation (1–5).
Endothelial dysfunction and apoptosis are often seen as the
initiating factors (3–6). A high-fat diet promotes the
development of AS by increasing endothelial permeability and
apoptosis (7–10). The discovery of microRNAs (miRNAs)
has provided a novel perspective for AS research. miRNAs are small,
single-stranded, non-coding RNA molecules (~22 nucleotides) that
are encoded within the genome and derived from endogenous small
hairpin precursors (3,4,11).
miRNAs are involved in the post-transcriptional regulation of gene
expression via binding to the 3′-untranslated regions (3′-UTRs) of
specific mRNAs, which subsequently inhibits the transcription or
translation of the target mRNAs (12,13).
It has been reported that miRNAs have key roles in various
physiological cellular activities, including development, growth,
proliferation, metabolism, differentiation and apoptosis. The
occurrence and development of numerous cardiovascular diseases,
including AS, hypertension and cardiac fibrosis, is associated with
aberrant expression of miRNAs (14–17).
miRNA (miR)-126 is currently the only known miRNA
expressed specifically in endothelial cells and hematopoietic stem
cells, and is closely associated with AS, coronary heart disease
and other cardiovascular diseases (18–22).
Our recent study demonstrated that miR-126 exerts anti-apoptotic
effects in human umbilical vein endothelial cells (HUVECs) by
inhibiting the expression of tumour necrosis factor
receptor-associated factor 7 and the production of reactive oxygen
species (23).
Transforming growth factor β (TGFβ) is a cytokine
that has key roles in numerous biological processes, including
inflammatory responses and apoptosis (24–26).
Studies have indicated that TGFβ is closely associated with
endothelial cells and the pathogenesis of AS (27–31).
The effect of miR-126 on high-fat diet-induced endothelial
permeability and apoptosis remains unclear. The results of the
present study suggested that miR-126 inhibited endothelial
permeability and apoptosis by inhibiting TGFβ expression via
binding to the 3′-UTR of TGFβ mRNA.
Materials and methods
Cell culture and reagents
Primary HUVECs were purchased from Lonza, Inc.
(Allendale, NJ, USA). Sulfo-NHS-LC-biotin was purchased from
Pierce; Thermo Fisher Scientific, Inc. (Waltham, MA, USA). miRNA
oligonucleotides (miR-126 mimic, 5′-CAGUACUUUUGUGUACAA-3′; miR-126
antagomir, 5′-CGCAUUAUUACUCACGGUACGA-3′; and negative control
miRNA, 5′-UUCUCCGAACGUGUCACGUTT-3′) and TransMessenger Transfection
reagent were obtained from Qiagen China Co., Ltd. (Shanghai,
China). Rhodamine600 (XRITC)-avidin was obtained from Pierce;
Thermo Fisher Scientific, Inc. HUVECs were cultured in endothelial
cell growth medium supplemented with 10% fetal bovine serum and 100
U/ml penicillin-streptomycin (Invitrogen; Thermo Fisher Scientific,
Inc.), and maintained at 37°C in a 5% CO2 atmosphere.
The cells at passage 3–4 were used for experiments. All experiments
were repeated at least three times.
Animal experimental procedures
All of the experiments performed in the present
study were approved by the Ethics Committee of The First Affiliated
Hospital of Anhui Medical University (Hefei, China). A total of 60
male apolipoprotein E (apoE)−/− mice (age, 4 weeks;
weight, 22±5 g) were obtained from the Institute of Basic Medical
Sciences of Peking Union Medical College (Beijing, China). Mice
were housed individually in screen-bottomed plastic cages in a
temperature-controlled room (25°C) under a 12-h light/dark cycle
with free access to food and water. Mice were divided into two
groups: Standard diet (n=16) and standard diet plus 5% lard oil and
2% cholesterol (n=44); diets were maintained for 16 weeks. At 12
weeks, 8 control and 8 high-fat diet mice were euthanized for
detection of miR-126 expression levels, and the remaining control
and high-fat diet mice were randomly divided into the following
three groups: Control miRNA oligonucleotides (n=8; 4 control mice
and 4 high-fat diet mice); miR-126 antagomir oligonucleotides
(n=18; 9 control mice and 9 high-fat diet mice); and miR-126 mimic
oligonucleotides (n=18; 9 control mice and 9 high-fat diet mice).
Control miRNA, or miR-126 antagomir or mimic was injected
subcutaneously at a dose of 10 mg/kg twice in the first week,
followed by once a week for 4 weeks. During the study period, 1
mouse died in the control miRNA oligonucleotide group, 2 died in
the miR-126 antagomir oligonucleotide group and 2 died in the
miR-126 mimic oligonucleotide group. At the end of the experiment,
the aorta was removed from mice as previously described (8). Part of the aorta was embedded in
optimal cutting temperature compound and snap frozen. The remaining
aorta was stored at −80°C.
miR-126 expression assay
Total RNA was extracted from the aorta using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). miR-126 expression was determined using a miRNA Plate Assay
kit (cat no. MA-0101; Signosis, Inc., Santa Clara, CA, USA) and an
oligo mix specific for miR-126 (cat no. MO-0040; Signosis, Inc.),
according to the manufacturer's protocol. The RNA content was
normalized to U6 small nuclear (sn)RNA. miR-126 expression was also
determined using reverse transcription-quantitative polymerase
chain reaction (RT-qPCR) with the following primers: miR-126,
forward 5′-UCGUACCGUGAGUAAUAAUGCG-3′, reverse
5′-CAUUAUUACUCACGGUACGAUU-3′; and U6 snRNA, forward
5′-CTCGCTTCGGCAGCACA-3′ and reverse 5′-AACGCTTCACGAATTTGCGT-3′.
Amplification conditions were as follows: Initial denaturation at
95°C for 10 min, followed by 40 cycles at 95°C for 15 sec, and at
60°C for 60 sec. The reaction mixture contained the following: 2X
Power SYBR Green Master Mix (Applied Biosystems; Thermo Fisher
Scientific, Inc.), cDNA template, 10 pmol/µl of each primer and
sterile H2O. Relative gene expression was quantified
according to the comparative Cq method (32) using GraphPad Prism software version
5.01 (GraphPad Software, Inc., La Jolla, CA, USA).
Permeability assay
The permeability assay was performed as previously
described (33). Briefly, frozen
aorta sections (10-µm thick) were stained at 37°C with
Sulfo-NHS-LC-biotin for 30 min, and blocked with 5% non-fat milk at
4°C overnight and subsequently submerged into blocking buffer
containing XRITC-avidin (1:500) at room temperature for 1 h. The
slides were washed three times with PBS and dried. Images were
captured using an Olympus Provis AX70 fluorescence microscope
(Olympus Corporation, Tokyo, Japan).
Caspase-3 activity measurement
Caspase-3 activity was detected in aortic samples
using a Caspase-3 Fluorometric assay kit (Enzo Life Sciences, Inc.,
Farmingdale, NY, USA), as previously described (34). Briefly, aortic samples were
isolated from anesthetized mice and stored in liquid nitrogen.
Samples were homogenized with lysis buffer containing 50 mM
Tris-HCl (pH 6.8), 10% glycerol and 2% SDS for 3 min on ice,
maintained on ice for 10 min, centrifuged at 21,130 × g for 10 min
at 4°C, and supernatants were collected and cryopreserved at −70°C
until further use. Protein concentration was determined using a
bicinchoninic acid (BCA) assay with the Micro BCA™ Protein Assay
kit (Pierce; Thermo Fisher Scientific, Inc.). Equal amounts of
extracted protein samples (50 µg) were incubated at 37°C overnight
with N-acetyl-Asp-Glu-Val-Asp p-nitroanilide (Ac-DEVD-pNA). The
quantity of pNA that was released was estimated at 405 nm using a
microplate ELISA reader. Caspase-3 relative activity was calculated
as follows: Caspase-3 activity=(mean experimental absorbance/mean
control absorbance)x100%.
Western blot analysis
Aortic samples were homogenized, lysed for 3 min on
ice in 1X SDS lysis buffer containing 50 mM Tris-HCl (pH 6.8), 10%
glycerol and 2% SDS and boiled for 10 min, followed by
centrifugation at 16,000 × g for 10 min at room temperature.
Protein concentration was determined using a Micro BCA™ Protein
Assay kit (Pierce; Thermo Fisher Scientific, Inc.). Equal amounts
of extracted protein samples (20 µg) were separated by 10% SDS-PAGE
and transferred onto polyvinylidene difluoride membranes (GE
Healthcare Life Sciences, Chalfont, UK). Membranes were blocked
using 5% (w/v) bovine serum albumin (BSA; Amresco, LLC, Solon, OH,
USA) for 2 h at room temperature, followed by incubation with the
following primary antibodies: Anti-TGFβ (cat no. 3711; 1:1,000),
anti-B-cell lymphoma (Bcl)-2 (cat no. 2872; 1:1,000), purchased
from Cell Signaling Technology, Inc. (Danvers, MA, USA); and
anti-GAPDH (cat no. TA505454; 1:5,000; Beijing Zhongshan Jinqiao
Biotechnology Co., Ltd., Beijing, China) diluted in TBS containing
0.05% Tween-20 (TBST) at 4°C overnight. Following 3 washes with
TBST, horseradish peroxidase (HRP)-conjugated secondary antibodies
(cat nos. ZB-2305 and ZB-2301; 1:5,000; Beijing Zhongshan Jinqiao
Biotechnology Co., Ltd.) were added to membranes and incubated at
room temperature for 2 h. Protein bands were visualized by enhanced
chemiluminescence using ECL reagents (Pierce; Thermo Fisher
Scientific, Inc.). GAPDH was used as the loading control. Blots
were semi-quantified by densitometric analysis using the Image-Pro
Plus software version 6.0 (Media Cybernetics, Inc., Rockville, MD,
USA). Experiments were repeated 3 times.
Immunohistochemistry
Immunohistochemistry was performed as previously
described (8). Sections (10 µm) of
the frozen aorta tissue samples were blocked using PBS containing
0.05% Tween-20 and 1% BSA at room temperature for 30 min, and
incubated at 4°C overnight with an anti-TGFβ (cat no. 3711;
1:1,000) primary antibody, purchased from Cell Signaling
Technology, Inc. Subsequently, sections were incubated with a
HRP-conjugated goat secondary antibody (cat no. ZB-2301; 1:5,000;
Beijing Zhongshan Jinqiao Biotechnology Co., Ltd.) at room
temperature for 60 min. A Metal Enhanced 3,3′-diaminobenzidine
Substrate kit (Pierce; Thermo Fisher Scientific, Inc.) was used for
3 min to develop the colour, and sections were counterstained with
hematoxylin (10%) at room temperature for 30 sec. The integral
absorbance was examined under a light microscope and results were
quantified using the Image-Pro Plus software version 6.0 (Media
Cybernetics, Inc.).
Luciferase assay
The region of the TGFβ 3′-UTR containing the
potential binding site of miR-126 (source: NCBI GenBank; https://www.ncbi.nlm.nih.gov/nuccore/NM_011577.2)
was predicted using TargetScan version 7.1 (http://www.targetscan.org/vert_71/). The sequence was
inserted into the 3′ region of the luciferase gene in a luciferase
vector (wt-Luc-TGFβ; Shanghai GeneChem Co., Ltd., Shanghai, China),
and a mutated version of this sequence was inserted into the vector
(mu-Luc-TGFβ). Plasmid DNA (300 ng) and miR-126 mimic, antagomir or
control oligonucleotide (80 nmol/l) were co-transfected using
TransMessenger Transfection reagent into HUVECs seeded into 6-well
plates at a density of 1–2×105 cells/well (confluence,
60–70%) for 24 h at 37°C. The pRL-TK vector (Promega Corporation,
Madison, WI, USA) expressing Renilla luciferase served as a
control. The luciferase assay was performed using a
Dual-Luciferase® Reporter assay system (Promega
Corporation) 24 h following transfection, according to the
manufacturer's protocol.
Statistical analysis
Data are presented as the mean ± standard deviation
of 3 independent experiments. The statistical significance of the
differences between groups was assessed using one-way analysis of
variance followed by a Neuman-Keuls post hoc test for multiple
comparisons. Comparisons between two groups were based on least
significant differences. P<0.05 was considered to indicate a
statistically significant difference. Statistical analysis was
performed using SPSS software version 21.0 (IBM Corp., Armonk, NY,
USA).
Results
Reduction of miR-126 expression and
increase in apoptosis of arterial endothelial cells in high-fat
diet fed mice
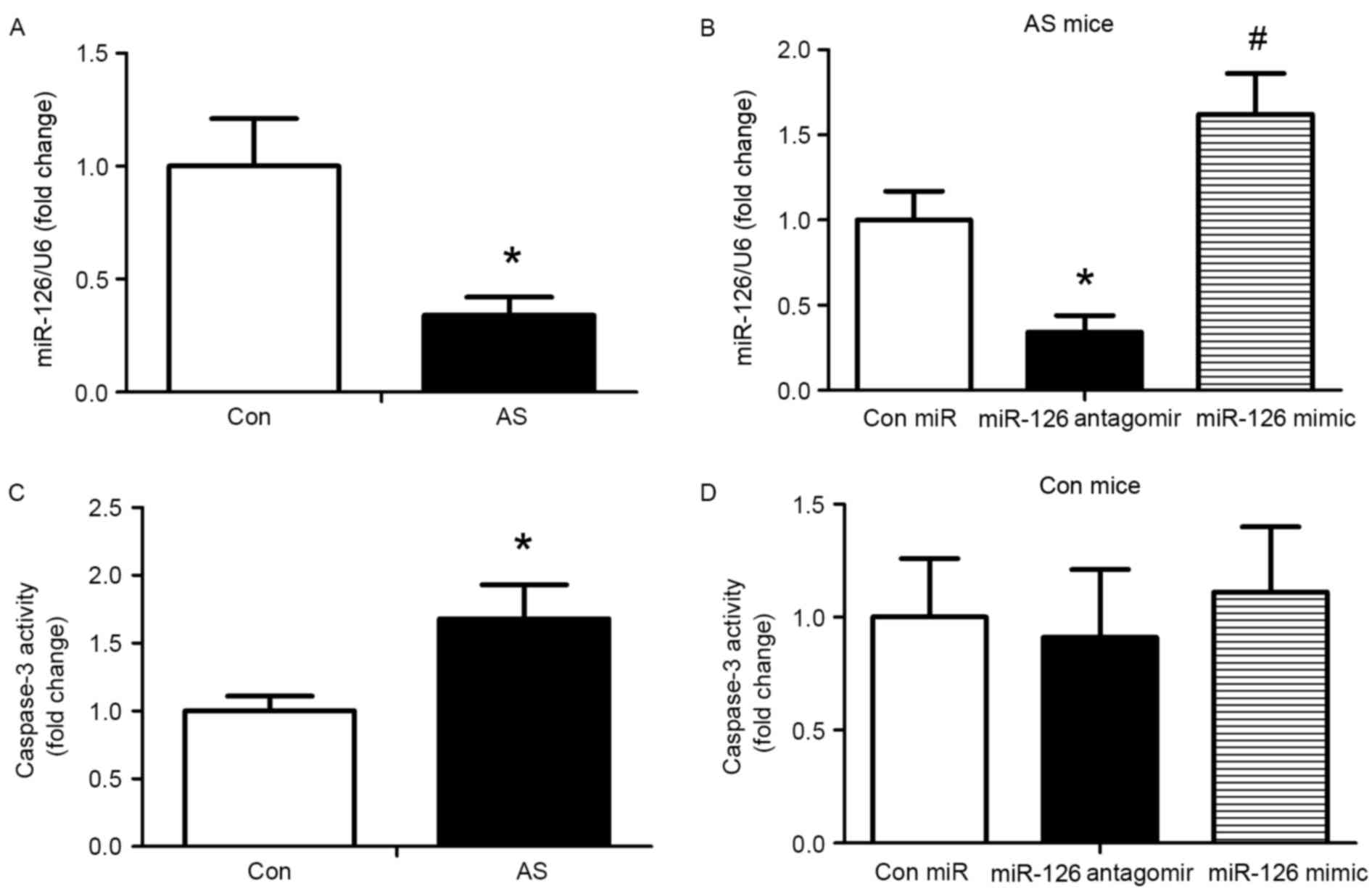
To evaluate the impact of miR-126 on an established
AS model, apoE−/− mice were fed a high-fat diet for 12
weeks, followed by subcutaneous injection of 10 mg/kg control
miRNA, miR-126 antagomir or miR-126 mimic twice for one week, and
once a week for 4 weeks. Subsequently, the expression of miR-126 in
the aorta was determined using a miRNA Plate assay kit. miR-126
levels were reduced in AS mice compared with control mice
(P<0.05; Fig. 1A). Following 4
weeks of treatment, the miR-126 mimic significantly increased
miR-126 expression and the miR-126 antagomir significantly reduced
the expression of miR-126 compared with AS mice injected with
control miRNA (P<0.05; Fig.
1B). In addition, caspase-3 activity was measured following 4
weeks of treatment with the miRNAs. Compared with the control
group, caspase-3 activity was greater in AS mice (P<0.05;
Fig. 1C). Treatment with miR-126
antagomir and mimic exhibited no effect on caspase-3 activity in
mice fed a standard diet (Fig.
1D). Thus, the standard diet group was not analysed in
subsequent experiments.
miR-126 expression alleviates
endothelial permeability in AS model mice
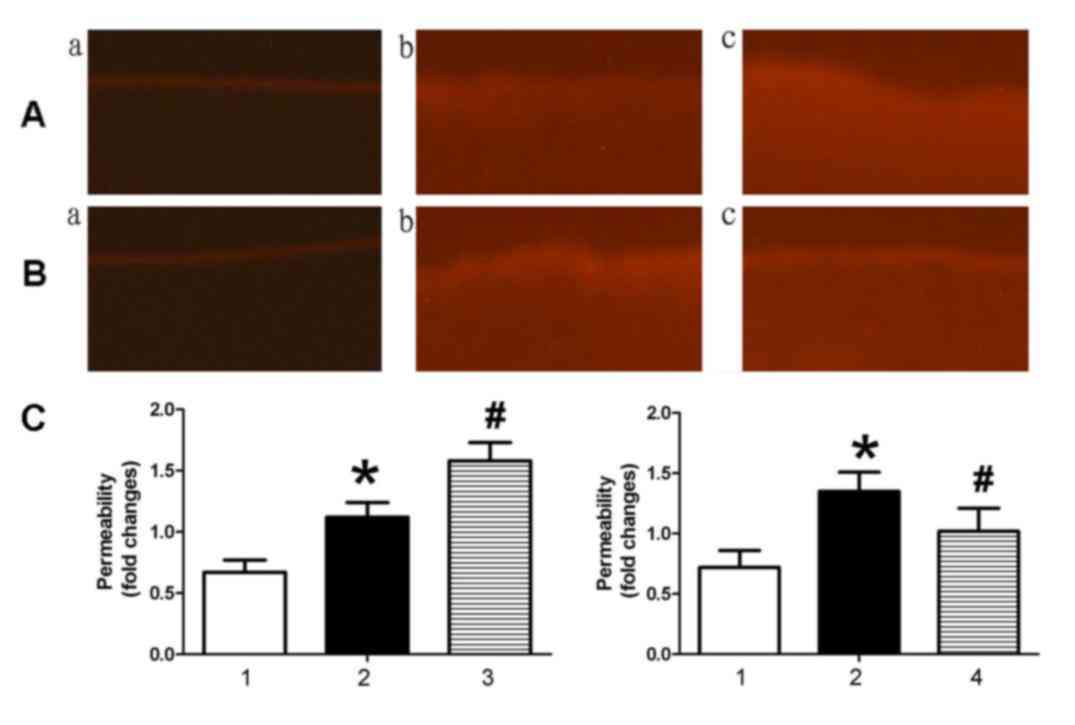
To investigate the effect of miR-126 on endothelial
permeability, NHS-LC-biotin and XRITC-avidin were used to detect
the endothelial permeability of aortic intima. Results presented in
Fig. 2 demonstrated that the
aortic intima from AS mice exhibited greater staining with
NHS-LC-biotin compared with control mice, in which only the
endothelial surface was stained. When AS mice were treated with an
miR-126 antagomir (Fig. 2Ac), an
increased quantity of NHS-LC-biotin leaked into the aortic intima
compared with AS mice treated with control miRNA (Fig. 2Ab). By contrast, miR-126 mimic
(Fig. 2Bc) treatment reduced
endothelial permeability compared with control miRNA-treated AS
mice (Fig. 2Bb).
miR-126 inhibits apoptosis in AS model
mice
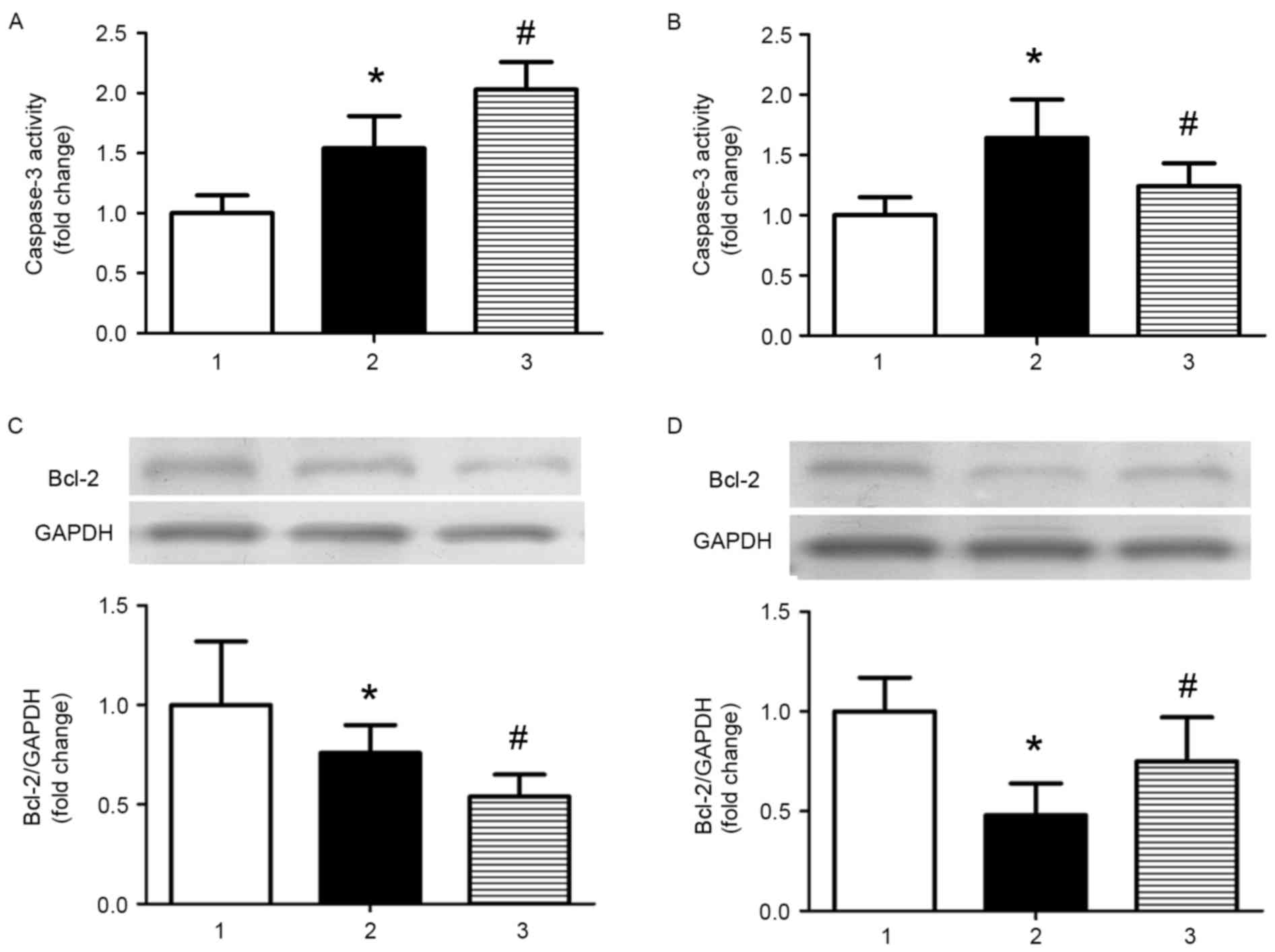
As apoptosis is involved in the development of AS,
the effect of miR-126 on apoptosis was investigated in the current
study. Caspase-3 activity was significantly increased in AS model
mice treated with control miRNA compared with control mice treated
with control miRNA, and miR-126 antagomir treatment further
increased caspase-3 activity (P<0.05; Fig. 3A). Treatment with an miR-126 mimic
partially reversed the increased activity of caspase-3 induced by a
high-fat diet (P<0.05; Fig.
3B). The opposite results were observed when the expression
levels of the anti-apoptotic protein B-cell lymphoma 2 (Bcl-2) were
analysed (Fig. 3C and D).
miR-126 modulates TGFβ expression in
AS model mice
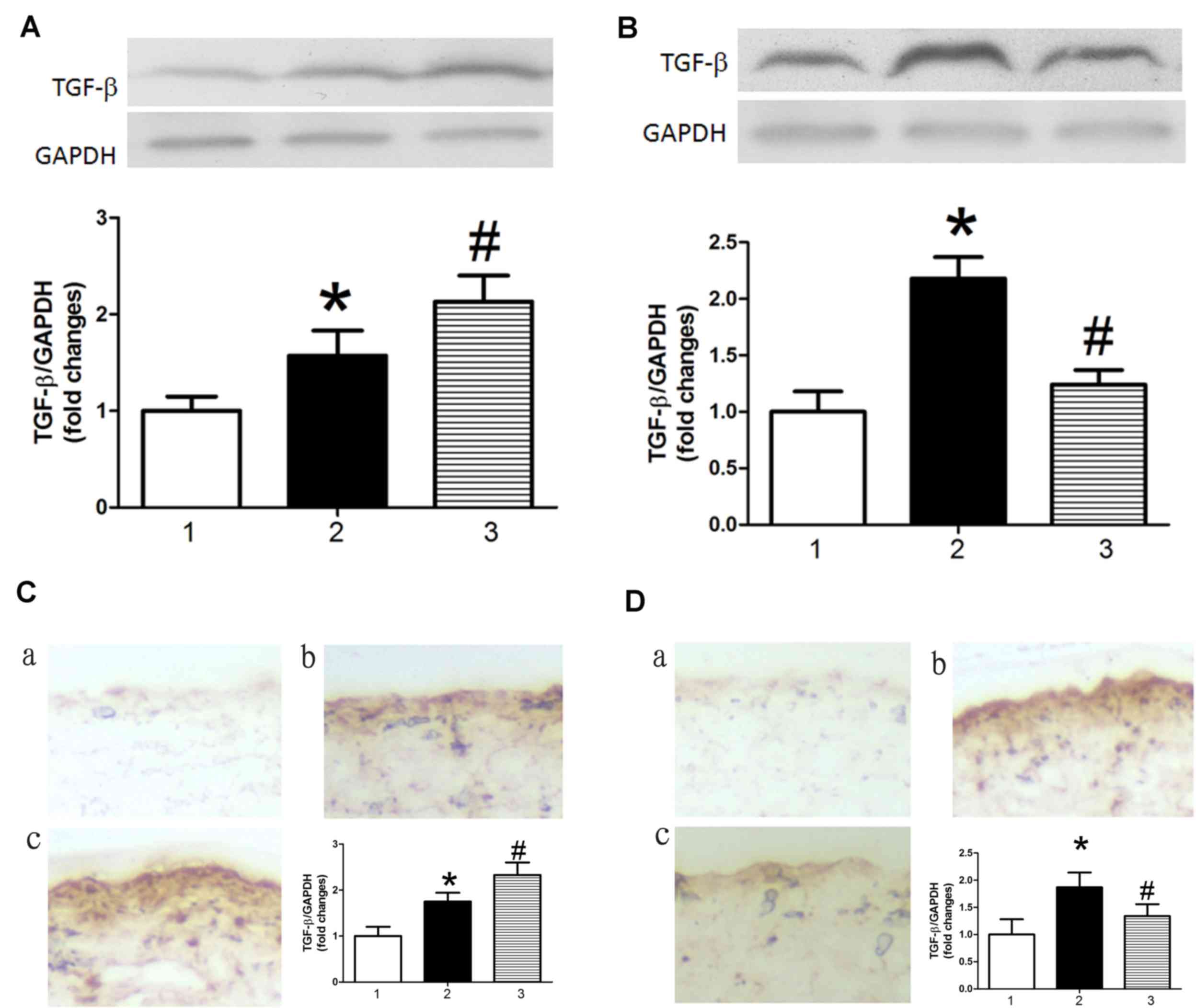
Previous studies have demonstrated that TGFβ
promotes apoptosis in endothelial cells (22–24).
Therefore, the present study investigated the effect of miR-126 on
the expression of TGFβ. TGFβ protein expression levels were
increased in AS model mice treated with control miRNA compared with
control mice treated with control miRNA, as determined by western
blot analysis (Fig. 4A and B) and
immunohistochemistry (Fig. 4C and
D). This result indicated that TGFβ may be a target gene of
miR-126. Results in Fig. 4
demonstrated that miR-126 antagomir treatment significantly
increased levels of TGFβ in AS model mice (Fig. 4A and C), whereas an miR-126 mimic
had the opposite effect (Fig. 4B and
D).
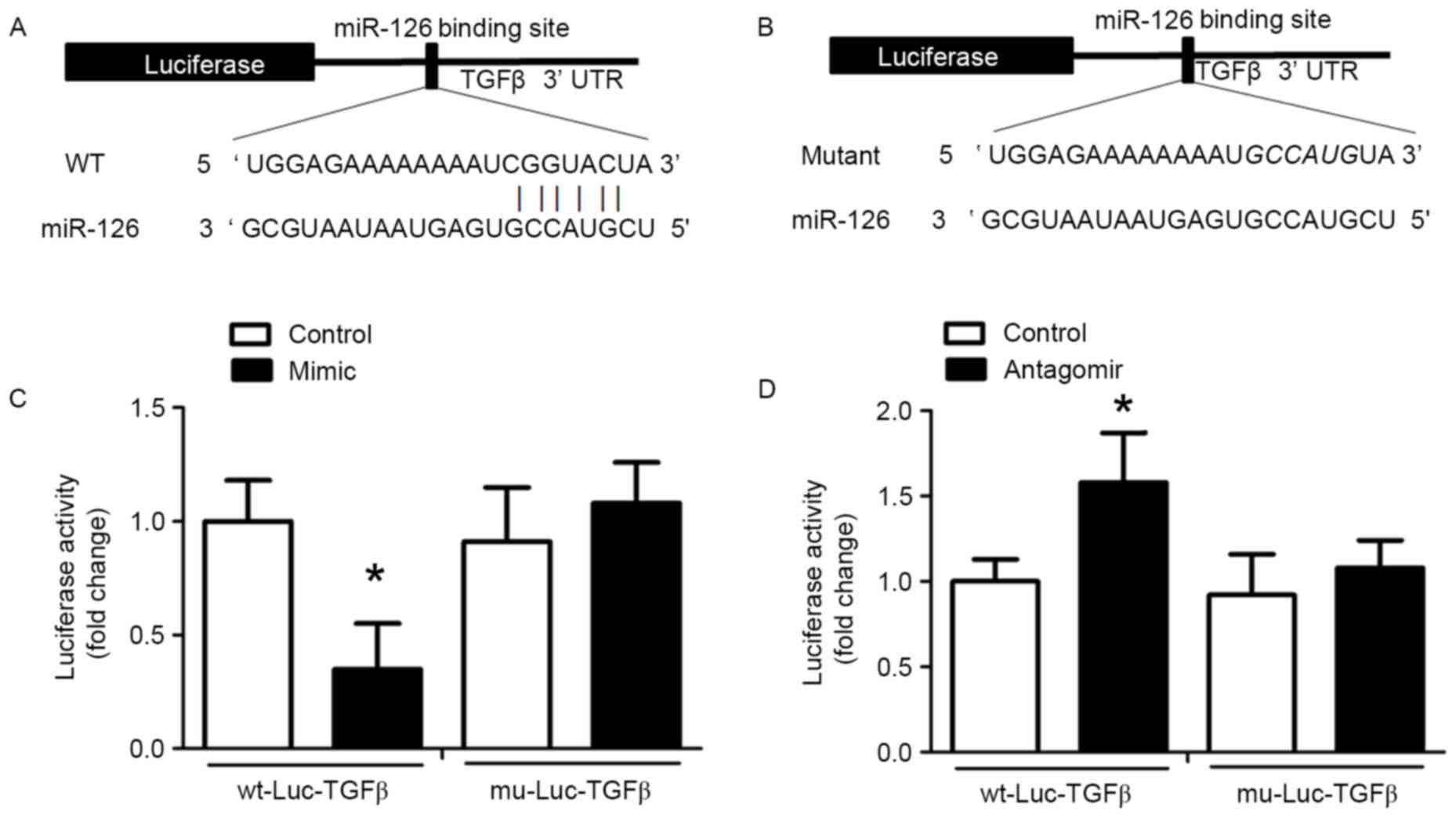
TGFβ is a target of miR-126
miRNAs specifically bind to the 3′-UTR of target
mRNAs and induce transcript degradation or translational
repression. An miR-126 binding site in the TGFβ 3′-UTR was
identified and the alignment between miR-126 and the wild-type and
mutant TGFβ 3′-UTR are presented in Fig. 5A and B, respectively. This
indicated that miR-126 may cause translation inhibition of TGFβ. To
validate this result of bioinformatics analysis, wt-Luc-TGFβ and
mu-Luc-TGFβ were inserted into the luciferase vector, which was
co-transfected into HUVECs with control miRNA, miR-126 antagomir or
miR-126 mimic. Treatment with miR-126 mimic reduced luciferase
activity in HUVECs co-transfected with wt-Luc-TGFβ compared with
HUVECs co-transfected with wt-Luc-TGFβ and control miRNA (Fig. 5C). In HUVECs co-transfected with
mu-Luc-TGFβ and an miR-126 mimic, no inhibition was observed
(Fig. 5C). miR-126 antagomir
significantly promoted luciferase activity in the
wt-Luc-TGFβ-transfected HUVECs, but had no effect on
mu-Luc-TGFβ-transfected HUVECs (Fig.
5D).
Discussion
The present study investigated the effect of
alterations in miR-126 expression on endothelial cell permeability
and apoptosis in a mouse model of AS. The present results
demonstrated that TGFβ was a direct target gene for miR-126, thus
suggesting that miR-126 may bind to TGFβ and suppresses its
expression. These findings indicated a novel mechanism by which
miR-126 may have a vital role in endothelial permeability and
apoptosis in AS model mice.
It has been reported that a high-fat diet is one of
the factors that promotes AS development (7,31,35).
Jakob et al (36)
demonstrated that the expression of miR-126 in patients with
chronic heart failure was significantly reduced compared with
healthy controls. Furthermore, cardiac function was markedly
improved by transfection with miR-126. The present study
demonstrated that arterial wall permeability and apoptosis was
increased in AS, and inhibition of miR-126 promoted this
pathological process. Overexpression of miR-126, using an miR-126
mimic, attenuated these effects. Thus, normal levels of miR-126 may
be necessary for the integrity of the arterial wall. However,
miR-126 inhibited apoptosis as demonstrated by reduced caspase-3
activity and increased protein expression levels of Bcl-2 in mice
fed high-fat diets. miR-126 had no effect on basal apoptosis in
mice fed standard chow. This indicated that miR-126 may only
inhibit apoptosis in mice on a high-fat diet, which inhibits
miR-126 expression. However, this requires further
investigation.
Typically, miRNAs influence gene expression by
inducing post-transcriptional inhibition, mRNA degradation or
translation suppression (12,13).
It has been reported that TGFβ has key roles in inflammatory
responses and apoptosis (24–26),
and is closely associated with the pathogenesis of AS (27,28).
The present study demonstrated that miR-126 directly binds to TGFβ
mRNA and inhibits its expression. The results of the current study
demonstrated that a reduction in miR-126 expression caused by a
high-fat diet increased TGFβ protein expression levels and caused
an increase in endothelial permeability and apoptosis.
In conclusion, the present study identified TGFβ as
a direct target gene of miR-126, and demonstrated that reduced
miR-126 expression increased TGFβ protein expression. These
findings suggested that TGFβ downregulation may be implicated in
the increased endothelial leakage and apoptosis that is observed in
AS. Based on the results of the current study, miR-126 may have
potential as a novel target for the treatment of cardiovascular
diseases.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81570419, 81470568
and 81270372), the National Natural Science Fund for Distinguished
Young Scholars of China (grant no. 81302150) and Grants for
Cultivating Program of National Natural Science Foundation for
Young Scholars of the First Affiliated Hospital of Anhui Medical
University (grant no. 2012KJ10).
References
|
1
|
Staff AC, Johnsen GM, Dechend R and Redman
CW: Preeclampsia and uteroplacental acute atherosis: Immune and
inflammatory factors. J Reprod Immunol. 101:120–126. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ooi CY, Sutcliffe MP, Davenport AP and
Maguire JJ: Changes in biomechanical properties of the coronary
artery wall contribute to maintained contractile responses to
endothelin-1 in atherosclerosis. Life Sci. 118:424–429. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schober A, Nazari-Jahantigh M and Weber C:
MicroRNA-mediated mechanisms of the cellular stress response in
atherosclerosis. Nat Rev Cardiol. 12:361–174. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Svoboda P: A toolbox for miRNA analysis.
FEBS Lett. 589:1694–1701. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xu S, Liu Z and Liu P: Targeting hydrogen
sulfide as a promising therapeutic strategy for atherosclerosis.
Int J Cardiol. 172:313–317. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lloyd MM, Grima MA, Rayner BS, Hadfield
KA, Davies MJ and Hawkins CL: Comparative reactivity of the
myeloperoxidase-derived oxidants hypochlorous acid and
hypothiocyanous acid with humancoronary artery endothelial cells.
Free Radic Biol Med. 65:1352–1362. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nahrendorf M and Swirski FK: Lifestyle
effects on hematopoiesis and atherosclerosis. Circ Res.
116:884–894. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhu HQ, Li Q, Dong LY, Zhou Q, Wang H and
Wang Y: MicroRNA-29b promotes high-fat diet-stimulated endothelial
permeability and apoptosis in apoE knock-out mice by
down-regulating MT-1 expression. Int J Cardiol. 176:764–770. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kraakman MJ, Kammoun HL, Allen TL,
Deswaerte V, Henstridge DC, Estevez E, Matthews VB, Neill B, White
DA, Murphy AJ, et al: Blocking IL-6 trans-signaling prevents
high-fat diet-induced adipose tissue macrophage recruitment but
does not improve insulin resistance. Cell Metab. 21:403–416. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
van Bussel BC, Henry RM, Ferreira I, van
Greevenbroek MM, van der Kallen CJ, Twisk JW, Feskens EJ,
Schalkwijk CG and Stehouwer CD: A healthy diet is associated with
less endothelial dysfunction and less low-grade inflammation over a
7-year period in adults at risk of cardiovascular disease. J Nutr.
145:532–540. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Romaine SP, Tomaszewski M, Condorelli G
and Samani NJ: MicroRNAs in cardiovascular disease: An introduction
for clinicians. Heart. 101:921–928. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Andreou I, Sun X, Stone PH, Edelman ER and
Feinberg MW: miRNAs in atherosclerotic plaque initiation,
progression, and rupture. Trends Mol Med. 21:307–318. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang W, Zhang E and Lin C: MicroRNAs in
tumor angiogenesis. Life Sci. 136:28–35. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Thum T and Condorelli G: Long noncoding
RNAs and microRNAs in cardiovascular pathophysiology. Circ Res.
116:751–762. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Loyer X, Mallat Z, Boulanger CM and Tedgui
A: MicroRNAs as therapeutic targets in atherosclerosis. Expert Opin
Ther Targets. 19:489–496. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nazari-Jahantigh M, Egea V, Schober A and
Weber C: MicroRNA-specific regulatory mechanisms in
atherosclerosis. J Mol Cell Cardiol. 89:35–41. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Orenes-Piñero E, Montoro-García S, Patel
JV, Valdés M, Marín F and Lip GY: Role of microRNAs in cardiac
remodelling: New insights and future perspectives. Int J Cardiol.
167:1651–1659. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fish JE, Santoro MM, Morton SU, Yu S, Yeh
RF, Wythe JD, Ivey KN, Bruneau BG, Stainier DY and Srivastava D:
microRNA-126 regulates angiogenic signaling and vascular integrity.
Dev Cell. 15:272–284. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Synetos A, Toutouzas K, Stathogiannis K,
Latsios G, Tsiamis E, Tousoulis D and Stefanadis C: microRNAs in
arterial hypertension. Curr Top Med Chem. 13:1527–1532. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hulsmans M and Holvoet P:
MicroRNA-containing microvesicles regulating inflammation in
association with atherosclerotic disease. Cardiovasc Res. 100:7–18.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun X, Zhang M, Sanagawa A, Mori C, Ito S,
Iwaki S, Satoh H and Fujii S: Circulating microRNA-126 in patients
with coronary artery disease: Correlation with LDL cholesterol.
Thromb J. 10:162012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Long G, Wang F, Duan Q, Chen F, Yang S,
Gong W, Wang Y, Chen C and Wang DW: Human circulating microRNA-1
and microRNA-126 as potential novel indicators for acute myocardial
infarction. Int J Biol Sci. 8:811–818. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Wang F, Wu Y, Zuo L, Zhang S, Zhou
Q, Wei W, Wang Y and Zhu H: MicroRNA-126 attenuates
palmitate-induced apoptosis by targeting TRAF7 in HUVECs. Mol Cell
Biochem. 399:123–130. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Franken R, den Hartog AW, de Waard V,
Engele L, Radonic T, Lutter R, Timmermans J, Scholte AJ, van den
Berg MP, Zwinderman AH, et al: Circulating transforming growth
factor-β as a prognostic biomarker in Marfan syndrome. Int J
Cardiol. 168:2441–2446. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yan F, Wang Y, Wu X, Peshavariya HM,
Dusting GJ, Zhang M and Jiang F: Nox4 and redox signaling mediate
TGF-β-induced endothelial cell apoptosis and phenotypic switch.
Cell Death Dis. 5:e10102014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Frei K, Gramatzki D, Tritschler I,
Schroeder JJ, Espinoza L, Rushing EJ and Weller M: Transforming
growth factor-β pathway activity in glioblastoma. Oncotarget.
6:5963–5977. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rath D, Chatterjee M, Müller I, Müller K,
Böckmann C, Droppa M, Stimpfle F, Karathanos A, Borst O, Seizer P,
et al: Platelet expression of transforming growth factor beta 1 is
enhanced and associated with cardiovascular prognosis in patients
with acute coronary syndrome. Atherosclerosis. 237:754–759. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hwang JS, Eun SY, Ham SA, Yoo T, Lee WJ,
Paek KS, Do JT, Lim DS and Seo HG: PPARδ modulates oxLDL-induced
apoptosis of vascular smooth muscle cells through a TGF-β/FAK
signaling axis. Int J Biochem Cell Biol. 62:54–61. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tian H, Liu J, Chen J, Gatza ML and Blobe
GC: Fibulin-3 is a novel TGF-β pathway inhibitor in the breast
cancer microenvironment. Oncogene. 34:5635–5647. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Marcantoni E, Dovizio M, Gaora OP, Di
Francesco L, Bendaya I, Schiavone S, Trenti A, Guillem-Llobat P,
Zambon A, Nardelli GB, et al: Dysregulation of gene expression in
human fetal endothelial cells from gestational diabetes in response
to TGF-β1. Prostaglandins Other Lipid Mediat. 120:103–114. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Climent M, Quintavalle M, Miragoli M, Chen
J, Condorelli G and Elia L: TG-Fβ triggers miR-143/145 transfer
from smooth muscle cells to endothelial cells, thereby modulating
vessel stabilization. Circ Res. 116:1753–1764. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2 (−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhu HQ, Zhou Q, Jiang ZK, Gui SY and Wang
Y: Association of aorta intima permeability with myosin light chain
kinase expression in hypercholesterolemic rabbits. Mol Cell
Biochem. 347:209–215. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhu H, Yang Y, Wang Y, Li J, Schiller PW
and Peng T: MicroRNA-195 promotes palmitate-induced apoptosis in
cardiomyocytes by down-regulating Sirt1. Cardiovasc Res. 92:75–84.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hu ZP, Fang XL, Fang N, Wang XB, Qian HY,
Cao Z, Cheng Y, Wang BN and Wang Y: Melatonin ameliorates vascular
endothelial dysfunction, inflammation, and atherosclerosis by
suppressing the TLR4/NF-κB system in high-fat-fed rabbits. J Pineal
Res. 55:388–398. 2013.PubMed/NCBI
|
|
36
|
Jakob P, Doerries C, Briand S, Mocharla P,
Kränkel N, Besler C, Mueller M, Manes C, Templin C, Baltes C, et
al: Loss of angiomiR-126 and 130a in angiogenic early outgrowth
cells from patients with chronic heart failure: Role for impaired
in vivo neovascularization and cardiac repair capacity.
Circulation. 126:2962–2975. 2012. View Article : Google Scholar : PubMed/NCBI
|