Introduction
The annual incidence of stroke in China is
120–180/100,000, with >2 million new cases every year (1). At least two-thirds of stroke patients
experience varying degrees of physical disability (2). Because of the rapid progression
following acute cerebral infarction, there has long been a lack of
particularly effective treatments. Intravascular thrombolytic
therapy is currently considered to be one of the most promising
treatment methods; however, it is only useful in a small number of
acute phase patients (3).
Therefore, it is important that research continues to develop drugs
that can intervene in the progression of ischemia.
Hypothermia therapy is considered to be an effective
means for treating cerebral ischemia and hypoxia, high intracranial
pressures and other complications that are caused by early stage
ischemic cerebrovascular disease (4). Hypothermia is thought to reduce the
rate of cerebral metabolism and oxygen consumption, and reduce the
synthesis and release of excitotoxic neurotransmitters and
inflammatory mediators (5).
Despite this, a previous study demonstrated that hypothermia did
not reduce the mortality in patients with severe brain damage or
massive cerebral infarction (6).
This clinical and laboratory discordance sparked renewed discussion
on the protection mechanisms of hypothermia, requiring further
molecular exploration of hypothermia protection mechanisms.
Small ubiquitin-related modifier (SUMO) functions as
a post-translational modification system that maintains
protein-protein interactions, cytoplasmic and nuclear
translocations, stabilizes the genome and antagonizes
ubiquitination (7). SUMO
modification is a reversible dynamic equilibrium event, which
suggests that SUMO-associated disease may be reversed. For example,
the increased conjugation of SUMO with protein is observed during
neuroprotection after cerebral ischemia (8). Lee et al (9) found that a large number of proteins
had been modified by SUMOs in squirrels during hibernation so as to
resist glucose and oxygen depletion. Based on previous results, it
is hypothesized that the SUMOylation of neuronal proteins may be
increased to assist neuroprotection during hypothermia; however,
the mechanisms remain unclear.
In the current study, a human cerebral ischemia
event was simulated by an in vitro ischemia model that used
oxygen-glucose deprivation (OGD), and an in vivo rat model
of middle cerebral artery occlusion (MCAO). Results suggest that in
early cerebral ischemia, SUMO-2/3, but not SUMO-1, is rapidly
activated, and covalently bound to abundant proteins to prevent
them from being hydrolyzed by ubiquitin, which greatly increases
the tolerance of injured neurons to the hypoxic environment. These
findings indicate a novel neuroprotective mechanism of hypothermia.
That is, abundant proteins can conjugate to, and be protected by,
SUMO-2/3 during early ischemia. These finding may provide an avenue
for the development of treatments for patients with cerebral
ischemia in the future.
Materials and methods
Experimental animals
Male Sprague-Dawley (SD) rats (n=36; 12-weeks-old)
and 2 female pregnant rats (15-weeks-old) were purchased from the
Animal Center of the Cancer Institute of Chinese Academy of Medical
Science (Beijing, China), and housed in the Animal Experimental
Center, Capital Medical University (Beijing, China), with humidity
of 50±5% at 20–25°C. a 12-h light/dark cycle (8:00-20:00) and free
access to food and water. All the experiments were performed
according to the Principles of Laboratory Animal Care (10) and approved by the Ethics Committee
of Capital Medical University (Beijing, China).
Primary neuronal cell cultures
Primary neuronal cell cultures were prepared from
the cortex of embryonic rat brains at gestation day 18. The entire
cortex was dissected and digested by 0.125% trypsin over 10 min.
The cells were plated at a density of 100,000 cells/cm2
in neurobasal medium (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with B27, glutamax I, 5% fetal
bovine serum (Invitrogen; Thermo Fisher Scientific, Inc.), and 1
µg/ml gentamicin, at 37°C with 5% CO2. After 3 days in
culture, cytosine-β-D-arabino-furanoside was added to a final
concentration of 5 µM. Cells were then fed twice a week with
serum-free neurobasal/B27 medium for an additional 7–9 days.
Immunocytochemistry
The primary neuronal cells were fixed with 4%
paraformaldehyde at room temperature for 10 min, and blocked with
5% goat serum (Beijing Zhongshan Bio Corp., Beijing, China), 2%
bovine serum albumin (BSA; Beijing Zhongshan Bio Corp.) and 0.1%
Triton-X-100 at 37°C for 1 h. Primary antibodies against neuron
specific enolase (NSE; 1:1,000 dilution; cat. no. sc-51880; Santa
Cruz Biotechnology Inc., Dallas, TX, USA) and glial fibrillary
acidic protein (GFAP; 1:4,000 dilution; cat. no. TA500335; Beijing
Zhongshan Bio Corp.) were incubated with the cells in moisture box
at 4°C overnight. The sections were subsequently incubated with
goat anti-mouse IgG-CruzFluor™ 488 (1:400 dilution; cat. no.
sc-362257; Santa Cruz Biotechnology, Inc.) or goat anti-mouse
IgG-CruzFluor™ 594 (1:400 dilution; cat. no. sc-362277; Santa Cruz
Biotechnology, Inc.) secondary antibodies at 37°C for 1 h, followed
by mounting and observation under a fluorescence microscope
(Olympus DP 70; Olympus Corporation, Tokyo, Japan).
OGD model and hypothermia
treatment
To simulate cerebral ischemia, neuronal cells were
cultured in an anoxic chamber (Forma Scientific Anaerobic System).
Neurobasal medium free of glucose, L-aspartic acid, L-glutamic
acid, and sodium pyruvate was equilibrated overnight in the anoxic
chamber with 85% N2, 10% H2, and 5% CO2. At ~90%
confluence, cultures were transferred to the anoxic chamber and
washed three times with the anoxic medium. After 10, 30 and 60 min,
and 2, 4, 8, 12 and 48 h of OGD exposure, the anoxic medium was
replaced with neurobasal/B27 medium and cells were transferred back
to the incubator at 37°C with a gas mixture of 95% air and 5%
CO2 for an additional 24 h. For induction of
hypothermia, which was performed simultaneously with OGD, the
neurobasal/B27 medium was replaced with neurobasal medium
pre-cooled to 33°C and cultures were placed in a 33°C
environment.
Western blot analysis
The cells were solubilized in 1% Nonidet P-40 lysis
buffer. Lysates (40 µg) were separated by 8% sodium dodecyl
sulfate-acrylamide electrophoresis. Separated proteins were
transferred to polyvinylidene fluoride membranes (EMD Millipore,
Billerica, MA, USA) and blocking was performed with 5% skimmed milk
at room temperature for 1 h, which was followed by incubation with
primary antibodies against SUMO-1 (1:100 dilution; cat. no.
sc-130275; Santa Cruz Biotechnology, Inc.) and SUMO-2/3 (1:400
dilution; cat. no. sc-32873; Santa Cruz Biotechnology, Inc.),
followed by incubation with horseradish peroxidase (HRP)-conjugated
goat anti-mouse IgG secondary antibody (1:2,000 dilution; cat. no.
31430; Thermo Fisher Scientific, Inc.) or goat anti-rabbit IgG
secondary antibodies (1:5,000 dilution; cat. no. 31466; Thermo
Fisher Scientific, Inc.) at room temperature for 1 h. Conjugated
and free SUMO proteins were detected using the same primary
antibodies, and distinguished on the same blot based on the
different molecular weights of the free and conjugated forms of the
proteins. Specific proteins were detected using a SuperSignal
protein detection kit (Pierce; Thermo Fisher Scientific, Inc.). The
membrane was stripped by Restore™ Plus Western Blot Stripping
Buffer (Invitrogen; Thermo Fisher Scientific, Inc.) and reprobed
with a primary monoclonal antibody against β-actin (1:5,000
dilution; cat. no. MA5-15739; Thermo Fisher Scientific, Inc.) for 2
h at room temperature. The membrane was washed in TBS-0.1% Tween-20
and probed with HRP-conjugated goat anti-mouse IgG secondary
antibody (1:2,000 dilution; cat. no. 31430; Thermo Fisher
Scientific, Inc.) at room temperature for 1 h. Optical density was
measured and analyzed using Quantity One 4.62 software (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
Lactate dehydrogenase (LDH) activity
detection
Samples were divided into control group, OGD group,
and OGD + hypothermia group. At 24 h after experimental conditions
were established, LDH content in the conditioned medium was
measured by enzyme-linked immunosorbent assay Lactate Dehydrogenase
Assay kit (Colorimetric; cat. no. ab102526; Abcam, Cambridge, UK)
in accordance with the manufacturer's instructions.
Apoptosis detection
Neurons were harvested 24 h after experimental
conditions were established, washed with phosphate-buffered saline
(PBS), immersed in permeabilization solution (0.1% Triton-X-100 +
0.1% sodium citrate) for 5 min, and incubated with 25 µl TUNEL
reaction mixture (Click-iT® TUNEL Alexa
Fluor® 488 Imaging assay; Thermo Fisher Scientific,
Inc.) in a wet box at 37°C for 60 min. After washing with PBS,
nuclei were counter-stained with Hoechst 33258. Samples were washed
with PBS and deionized water, mounted and observed with a
fluorescence microscope. The total number of cells (Hoechst 33258
staining) and the number of apoptotic cells (green fluorescence)
were quantified. Apoptotic rate (%) was equal to the number of
apoptotic cells/number of total cells ×100.
Establishment of a rat model of MCAO
and hypothermia treatment
Adult SD rats (n=48) were randomly divided into
three groups, with 16 rats in each group. The experiment was
performed using a small animal ventilator. Body temperature was
monitored with a rectal thermometer. A 1-cm-long medial
longitudinal incision was made from the manubrium to mandible. The
left common carotid artery, external carotid artery, and internal
carotid artery were isolated under a microscope. The distal segment
of the common carotid artery and the proximal segment of the
external carotid artery were ligated. A nylon suture with a
0.23-mm-diameter tip and 0.18-mm trunk was inserted into the middle
cerebral artery through the common carotid artery (~12 mm) and
fixed. In the sham surgery group (control group), the arteries were
exposed, but without the insertion of a nylon suture. In the
hypothermia group, the experiment was carried out on controlled
temperature blankets. After anesthesia, SD rat body temperature was
controlled between 32 and 34°C with a rectal thermometer. Following
occlusion, body temperature was allowed to gradually recover to
normal over 10 h. Laboratory animals were housed in a quiet clean
room. At 1, 7, 14 and 21 day after the models were established, the
neurological deficit of all rats was evaluated as previously
described (11).
Immunofluorescence analysis of SUMO
proteins
At 4 days after model establishment, 4 rats in each
group were sacrificed, and the brain tissue was isolated and fixed
with 10% formaldehyde at 4°C for 24 h, and subsequently embedded in
paraffin and sliced into 5-µm-thick coronal sections. Hippocampal
sections were de-waxed with xylene and alcohol, and hydrated
through an alcohol gradient. Following antigen retrieval with 0.01
M citric acid buffer (pH=6.0), sections were blocked with 5% goat
serum at room temperature for 10 min and incubated with mouse
anti-SUMO-1 (1:100 dilution; cat. no. sc-5308; Santa Cruz
Biotechnology, Inc.) and rabbit anti-SUMO-2/3 (1:500 dilution; cat.
no. sc-32873; Santa Cruz Biotechnology, Inc.) primary antibodies at
4°C overnight. The samples were washed repeatedly, incubated with
goat anti-mouse IgG-CruzFluor™ 488 (1:400 dilution; cat. no.
sc-362257; Santa Cruz Biotechnology, Inc.) or goat anti-rabbit IgG-
CruzFluor™ 594 (1:400 dilution; cat. no. sc-362282; Santa Cruz
Biotechnology, Inc.) secondary antibodies at 37°C for 1 h,
counter-stained with DAPI, mounted with fluorescence mounting
medium and observed using a fluorescence microscope.
Statistical analysis
Data are expressed as the mean ± standard deviation.
Data were analyzed using one-way analysis of variance by SPSS 13.0
software (SPSS Inc., Chicago, IL, USA). The Tukey test was
performed for multiple comparisons. P<0.05 was considered to
indicate a statistically significant difference.
Results
Neuronal culture
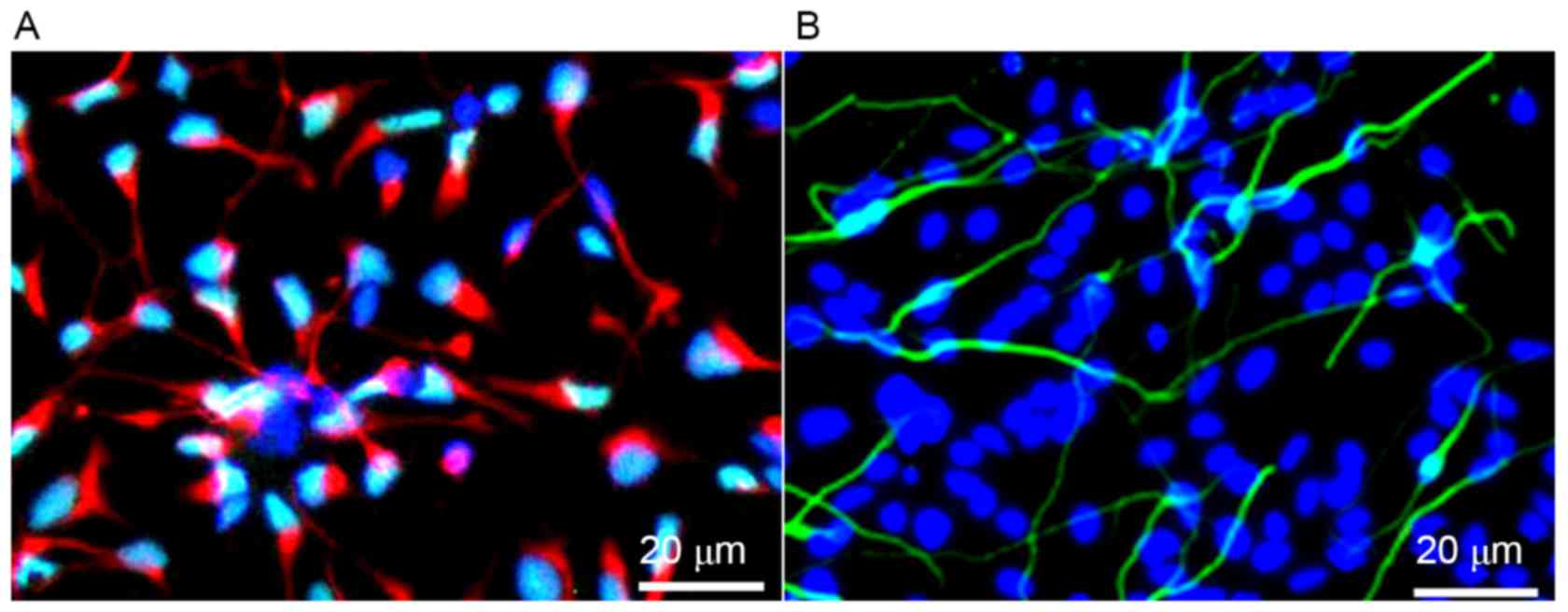
Immunofluorescence demonstrated that after 10 days
of primary culture, >90% of the primary cells expressed
neuron-specific protein NSE, and <10% of cells expressed the
glial cell specific marker proteins, GFAP (Fig. 1). These results indicated that
neurons were successfully cultured at a high purity.
Western blot analysis
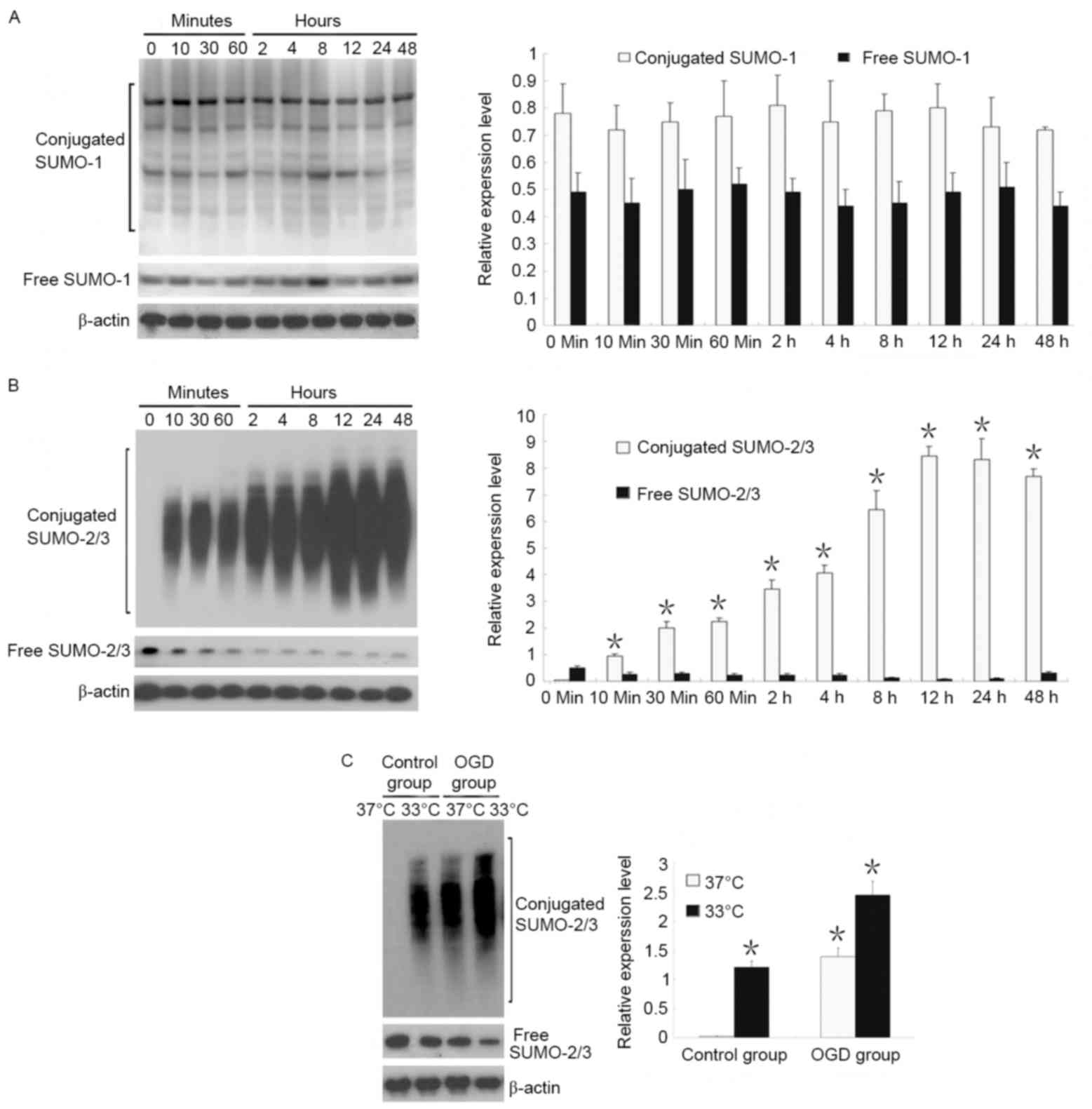
SUMO-1 and SUMO-2/3 expression was identified in the
early stage (48 h) of OGD. Results revealed that free and
conjugated SUMO-1 expression did not change within 48 h of OGD,
whereas conjugated SUMO-2/3 expression increased starting from 10
min of OGD and reached a peak at 12 h of OGD, and a gradual
decrease in free SUMO-2/3 (Fig. 2A and
B).
The effects of temperature on SUMO-1 and SUMO-2/3
expression were also observed. Results demonstrated that when
neurons were cultured at a normal temperature (37°C), there was
almost no conjugated SUMO-2/3, and that SUMO-2/3 was present only
in an unconjugated state. When neurons were cultured in hypothermia
conditions (33°C), conjugated SUMO-2/3 expression markedly
increased, suggesting that abundant proteins were bound to
SUMO-2/3. Under OGD conditions, hypothermia induced a 1.8-fold
increase in conjugated SUMO-2/3 expression compared with OGD at
37°C (Fig. 2C).
Neuroprotective effect of hypothermia
on OGD-induced injury
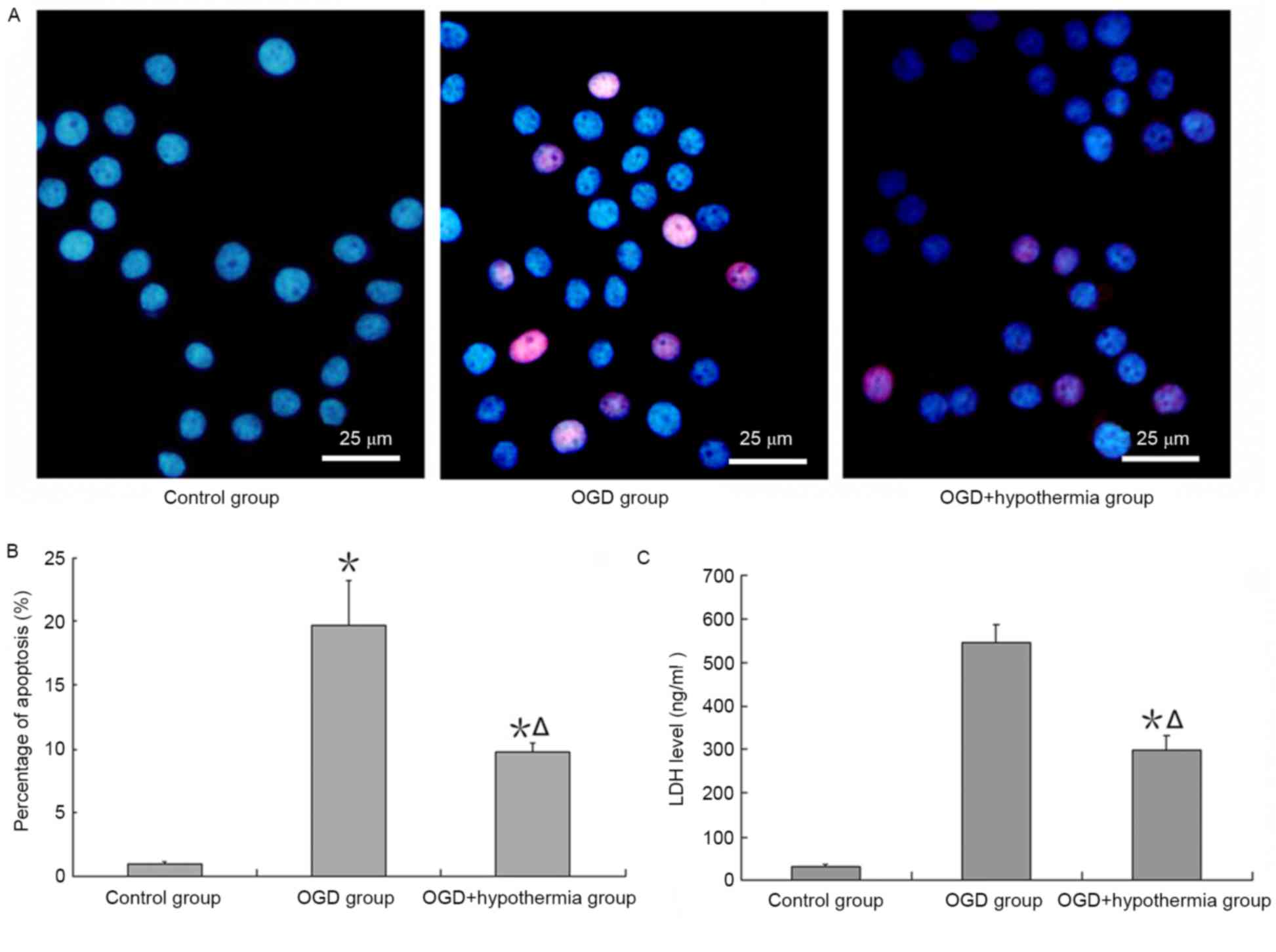
An apoptosis assay demonstrated that apoptosis was
increased by ODG, and this effect of apoptosis was reduced by
hypothermia (Fig. 3A and B).
Additionally, the results revealed that LDH expression was very low
in neurons cultured in normal conditions, but high when neurons
were exposed to OGD conditions. LDH expression was significantly
lower in OGD-injured neurons in the hypothermia group than in the
ODG normal temperature group (F=38.4, P<0.05; Fig. 3C).
Neurological deficit scores
Compared with the control group, neurological
deficit scores were higher in the MCAO group at the same time
point. Neurological deficit scores in the MCAO + hypothermia group
were lower than in the MCAO group in the early stage of cerebral
ischemia, but not in the late stage of ischemia (>21 days;
Table I).
 | Table I.Neurological impairment scores in
rats. |
Table I.
Neurological impairment scores in
rats.
|
| Time after MCAO
(days) |
|---|
|
|
|
|---|
| Group | 1 | 7 | 14 | 21 |
|---|
| Control | 0.14±0.03 | 0.11±0.03 | 0.08±0.02 | 0.09±0.02 |
| MCAO |
33.12±3.26a |
23.15±4.56a |
16.24±3.18a |
12.31±2.51a |
| MCAO +
hypothermia |
32.34±3.09a |
19.71±3.68a,b |
12.73±3.17a,b |
11.72±2.46a |
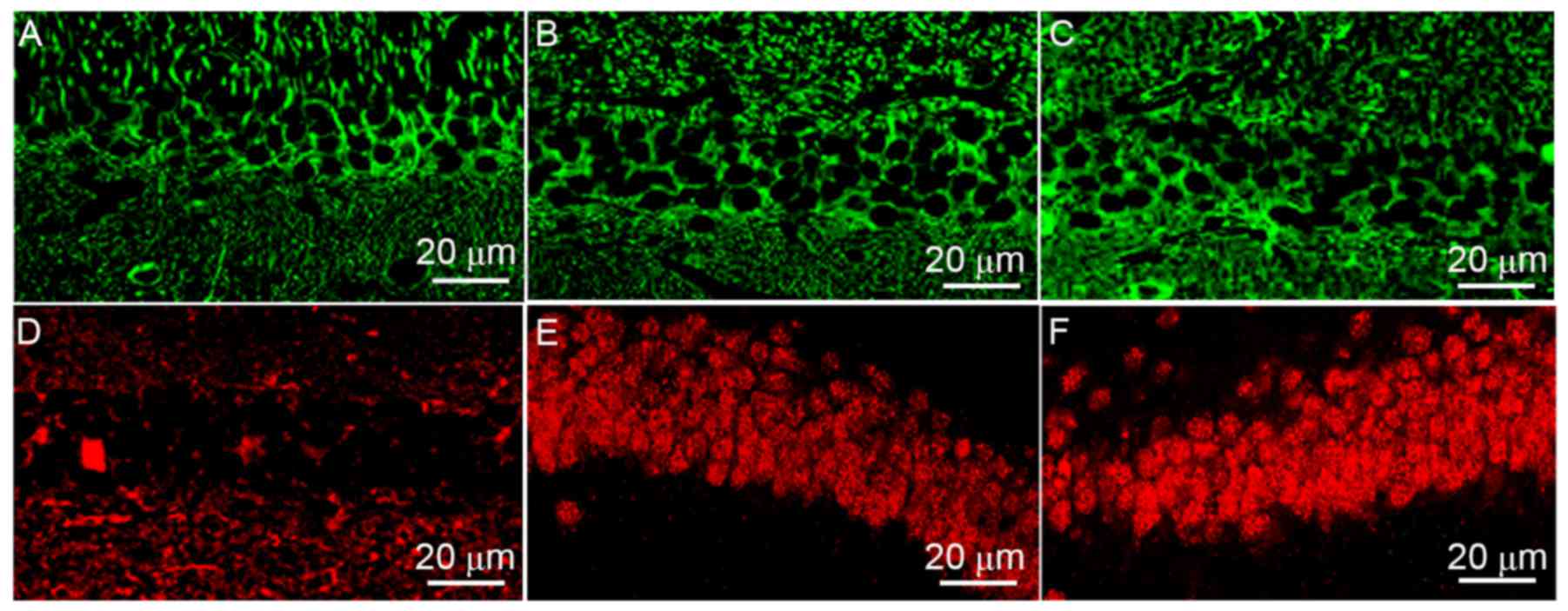
SUMO-2/3 transport from cytoplasm to
nucleus
The localization of SUMO-1 (Fig. 4A-C) and SUMO-2/3 (Fig. 4D-F) in rat hippocampal neurons
differed according to the treatment group. Results revealed that
SUMO-1 was mainly located in the cytoplasm in the control group.
MCAO and hypothermia did not impact on SUMO-1 expression or
localization. However, MCAO induced SUMO-2/3 translocation from the
cytoplasm to the nucleus, and hypothermia enhanced this
transport.
Discussion
Massive cerebral infarction has a high mortality and
disability rate due to abrupt onset and rapid progression.
Hypothermia has been considered as one of the early and effective
treatment methods for cerebral infarction-induced severe ischemia
and hypoxia. A recent large-scale clinical investigation suggested
that hypothermia does not significantly reduce the mortality in
patients with severe brain damage or massive cerebral infarction,
but may assist rescuing ischemic penumbra neurons (12). The current understanding of
hypothermia as a treatment encompasses the following (13–17):
i) Hypothermia decreases the cellular metabolism rate and increases
energy reserve for recovery; ii) hypothermia reduces the
accumulation of inflammatory substances in the local region, and
improves the local microenvironment; and iii) hypothermia enhances
the tolerance of injured nerve cells to the microenvironment by
reducing the activity of proteolytic enzymes. Based on the above,
it seems feasible to use targeted hypothermia neuroprotection
therapies in the early, critical window.
The biological functions of SUMO include maintaining
genomic stability, regulating protein interactions, adjusting
cytoplasmic protein transport and localization, controlling the
activity of transcription factors and antagonizing ubiquitination
(18–22). SUMO modification is a reversible
dynamic process, which indicates that SUMOylation-associated
disease can be reversed. Yang et al (23,24)
reported that cerebral ischemia activates a SUMO-2/3-mediated
molecular mechanism pathway and exert a neuroprotective effect. A
previous study demonstrated that following ischemic cerebrovascular
disease, proteins that are strongly associated with oxidative
stress, inflammation, DNA synthesis, energy transfer and metabolism
via SUMO can remove foreign substances, resist inflammation and
balance cell proliferation and apoptosis (25). In 2011, Lee et al (9) reported that the SUMO modification of
a large number of proteins increased in squirrels during
hibernation, presumably to resist glucose and oxygen depletion.
Thus, we hypothesized that SUMOs may have an important role in the
molecular mechanisms of hypothermia-mediated neuroprotection.
In the present study, we used an in vitro OGD
neuronal model that incorporated a hypothermia therapeutic element.
SUMO-2/3 was activated in neurons under ischemic and hypoxic
conditions, corresponding to increasing conjugated SUMO-2/3 protein
expression. This result suggests that a certain amount of SUMO-2/3
is reserved in neurons for use during stress responses, yet the
quantity of SUMO-2/3 in cells is ultimately limited. The amount of
SUMO-2/3 appeared to reach a peak at 12 h after the onset of
stress. After this time, more SUMO-2/3 would presumably need to be
synthesized to supplement the lost SUMO-2/3.
Subsequent experimental results found that
hypothermia could not only increase the expression level of
conjugated SUMO-2/3 in normal neurons, but also enhance the
conjugation of SUMO-2/3 to target proteins in OGD-injured neurons.
These findings indicate that hypothermia could initially increase
the conjugation of proteins to SUMO-2/3 in a physical capacity, and
may also enhance the conjugation of SUMO-2/3 to target proteins via
intervention from other stress factors.
To verify that SUMO-2/3 activation under hypothermia
is a neuroprotective response, the degree of apoptosis of
OGD-injured neurons cultured in vitro was investigated.
Hypothermia decreased the degree of apoptosis in early OGD. In the
MCAO model, hypothermia reduced rat neurological deficits in the
early stage, but this effect diminished with time. Ultimately,
there was no statistical difference between the neurological
deficit score of the MACO group and the MCAO + hypothermia group at
days 21. Previous findings indicate that hypothermia may delay the
apoptosis of injured cells in the early stage, but could not
ultimately stop this process, and may even result in the
acceleration of apoptosis upon rewarming (26). Our evidence supports this theory.
The precise mechanism of how this occurs requires further
investigation. Part of this may relate to the localization of SUMO,
where our in vivo results suggest that hypothermia increases
SUMO-2/3 transport from the cytoplasm to the nucleus, involving
increased conjugation with target protein. Future studies may be
able to determine whether the enhanced SUMO-2/3 conjugation is only
a temporary feature induced by hypothermia, or a genuine phenomenon
involving DNA repairing leading to neuron regeneration.
In conclusion, the current study confirmed that
hypothermia may enhance the conjugation of SUMO-2/3 to target
proteins and halt cellular apoptosis in early stage of ischemia.
This result serves to enrich the understanding of the molecular
mechanism of brain protection in hypothermia. The development of
small molecule drugs based on SUMO-2/3 may provide a putative
therapeutic target for patients with cerebral ischemia. Ultimately,
improving the understanding of the hypothermia process will lead to
more effective protocols for use in the critical, early stage of
ischemia therapy.
Acknowledgements
The present study was supported by the China
National Natural Scientific Fund (grant nos. 81471175 and
81371292).
Glossary
Abbreviations
Abbreviations:
|
SUMO
|
small ubiquitin-related modifier
|
|
OGD
|
oxygen-glucose deprivation
|
|
MCAO
|
middle cerebral artery occlusion
|
|
LDH
|
lactate dehydrogenase
|
References
|
1
|
Tu Y, Chen C, Sun HT, Cheng SX, Liu XZ, Qu
Y, Li XH and Zhang S: Combination of temperature-sensitive stem
cells and mild hypothermia: A new potential therapy for severe
traumatic brain injury. J Neurotrauma. 29:2393–2403. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kirton A and Deveber G: Life after
perinatal stroke. Stroke. 44:3265–3271. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wu S, Sena E, Egan K, Macleod M and Mead
G: Edaravone improves functional and structural outcomes in animal
models of focal cerebralischemia: A systematic review. Int J
Stroke. 9:101–106. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu TC and Grotta JC: Hypothermia for acute
ischaemic stroke. Lancet Neurol. 12:275–284. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang M, Wang H, Zhao J, Chen C, Leak RK,
Xu Y, Vosler P, Chen J, Gao Y and Zhang F: Drug-induced hypothermia
in stroke models: Does it always protect? CNS Neurol Disord Drug
Targets. 12:371–380. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kallmünzer B, Kollmar R and Schwab S:
Therapeutic hypothermia in acute brain injury. Nervenarzt.
83:975–981. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Droescher M, Chaugule VK and Pichler A:
SUMO rules: Regulatory concepts and their implication in neurologic
functions. Neuromolecular Med. 15:639–660. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cubeñas-Potts C and Matunis MJ: SUMO: A
multifaceted modifier of chromatin structure and function. Dev
Cell. 24:1–12. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee YJ, Mou Y, Maric D, Klimanis D, Auh S
and Hallenbeck JM: Elevated global SUMOylation in Ubc9 transgenic
mice protects their brains against focal cerebral ischemic damage.
PLoS One. 6:e258522011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guide for the care and use of laboratory
animals. Public Health Service, National Institute of Health: NIH
Publication No. 86–23. Revised. 1985.
|
|
11
|
Reglodi D, Tamás A and Lengvári I:
Examination of sensorimotor performance following middle cerebral
artery occlusion in rats. Brain Res Bull. 59:459–466. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Meixensberger J and Renner C: Therapeutic
hypothermia in the intensive care unit. Anaesthesist. 56:945–948.
2007.(In German). View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jordan JD and Carhuapoma JR: Hypothermia:
Comparing technology. J Neurol Sci. 261:35–38. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ramani R: Hypothermia for brain protection
and resuscitation. Curr Opin Anaesthesiol. 19:487–491. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Alzaga AG, Cerdan M and Varon J:
Therapeutic hypothermia. Resuscitation. 70:369–380. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nogoy N: Neuroproteomics: The hunt for
biomarkers of neurotrauma. Andrew ottens talks to nicole nogoy.
Expert Rev Proteomics. 4:343–345. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Han HS, Qiao Y, Karabiyikoglu M, Giffard
RG and Yenari MA: Influence of mild hypothermia on inducible nitric
oxide synthase expression and reactive nitrogen production in
experimental stroke and inflammation. J Neurosci. 22:3921–3928.
2002.PubMed/NCBI
|
|
18
|
Guo C and Henley JM: Wrestling with
stress: Roles of protein SUMOylation and deSUMOylation in cell
stress response. IUBMB Life. 66:71–77. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Feligioni M and Nisticò R: SUMO: A
(oxidative) stressed protein. Neuromolecular Med. 15:707–719. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sriramachandran AM and Dohmen RJ:
SUMO-targeted ubiquitin ligases. Biochim Biophys Acta. 1843:75–85.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang CJ, Wu D, Khan FA and Huo LJ:
DeSUMOylation: An important therapeutic target and protein
regulatory event. DNA Cell Biol. 34:652–660. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Abeywardana T and Pratt MR: Extent of
inhibition of α-synuclein aggregation in vitro by SUMOylation is
conjugation site- and SUMOisoform-selective. Biochemistry.
54:959–961. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang W, Thompson JW, Wang Z, Wang L, Sheng
H, Foster MW, Moseley MA and Paschen W: Analysis of
oxygen/glucose-deprivation-induced changes in SUMO3 conjugation
using SILAC-based quantitative proteomics. J Proteome Res.
11:1108–1117. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang W, Wang L and Paschen W: Development
of a high-throughput screening assay for inhibitors of small
ubiquitin-like modifier proteases. J Biomol Screen. 18:621–628.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang W, Sheng H, Homi HM, Warner DS and
Paschen W: Cerebral ischemia/stroke and small ubiquitin-like
modifier (SUMO) conjugation-a new target for therapeutic
intervention? J Neurochem. 106:989–999. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Olsen TS, Weber UJ and Kammersgaard LP:
Therapeutic hypothermia for acute stroke. Lancet Neurol. 2:410–416.
2003. View Article : Google Scholar : PubMed/NCBI
|