Introduction
Osteoarthritis (OA) is characterized by simultaneous
bone destruction and osteophyte formation; the main clinical
symptoms of OA include joint pain and dysfunction (1). Cartilage degradation induced by
chondrocyte inflammation has been hypothesized to be a crucial
event in the development of OA. Chondrocytes, which are the only
resident cell type present in articular cartilage, serve an
important role in the proliferation and degradation of cartilage
(2,3). It has been demonstrated that the
apoptosis of chondrocytes is significantly correlated with the
severity of OA, as it can cause matrix degradation and destruction,
finally resulting in OA (4,5).
It has been hypothesized that inflammation is
involved in the development and progression of OA, even in the
early stages of the disease. Tumor necrosis factor (TNF)-α is one
of the pivotal inflammatory factors in the pathogenesis of OA.
During the onset and progression of OA, the concentration of TNF-α
in the synovial fluid has been reported to increase, resulting in
the induction of inflammation, promotion of the release of bone
matrix-degrading enzymes, and the enhancement of fibroblastic
hyperplasia and histological damage (6). The Ras-Raf-mitogen-activated protein
kinase kinase (MEK)1/2-extracellular signal-regulated kinase
(ERK)1/2 signaling pathway regulates the TNF-α-induced inflammatory
response, and finally leads to cartilage degradation (7).
The Ras-Raf-MEK1/2-ERK1/2 signaling pathway is
involved in cell inflammation and is closely associated with the
progression of various diseases. TNF-α interacts with its receptor
on the cell surface and then initiates the Ras-Raf-MEK1/2-ERK1/2
signaling pathway inducing gene transcription. In addition, the
pathway regulates the activity of apoptotic molecules that are
normally sequestered in the mitochondria. Previous studies have
demonstrated that the pathway is inappropriately activated in
numerous types of cancer, leading to the activation of ERK via its
phosphorylation. Phosphorylated (p)-ERK interacts with various
substrates in the nucleus and cytosol, thus controlling the
transcription and expression of numerous genes involved in
inflammatory responses and cellular apoptosis (8,9).
Acupuncture has been widely used as an alternative
therapy for the treatment of several diseases (10–12).
Electroacupuncture (EA) is a modified acupuncture technique that
utilizes electrical stimulation. EA is an effective and simple
approach, which has been widely used for the treatment of OA, not
only to alleviate pain and disability, but also to decrease
inflammation and the progression of pathogenesis (13,14).
Our previous study demonstrated that EA treatment promotes
chondrocyte proliferation via promotion of G1/S
checkpoint transition in the cell cycle (15). However, the precise mechanism
underlying the effects of EA serum (EAS) on inflammation-induced
cartilage degradation remains to be elucidated. Therefore, the
present study investigated the effects of EAS on the TNF-α-mediated
inflammation of chondrocytes, in order to explore the underlying
molecular mechanisms.
Materials and methods
Animals
All animal procedures conducted in the present study
were approved by the Institutional Animal Care and Use Committee of
the Fujian University of Traditional Chinese Medicine (Fuzhou,
China). A total of 30 Sprague Dawley rats (age, 2 months; weight,
210±10 g) were purchased from the Shanghai Slack Laboratory Animal
Co., Ltd. (Shanghai, China; certification no. SCXK: 2012-0002). All
rats were housed in cages at a relative humidity of 55±5% and at
room temperature (24±2°C) under a 12-h light/dark cycle. The rats
were given ad libitum access to water and food.
EA treatment
Following 1 week of acclimation, the rats were
randomly divided into three groups: The blank group and two
experimental groups (n=10 rats/group). Rats in the two experimental
groups were treated with EA using the bilateral Neixiyan (EX-LE4)
and Waixiyan (ST 35) acupuncture points, which are located on the
inside and outside of the depression of the patellar ligament,
respectively. Disposable needles (diameter, 0.25 mm; length, 15 mm;
Huatuo; Suzhou Medical Appliance Factory, Suzhou, China) were
inserted to a depth of 4 mm at the acupuncture points. The
acupuncture points of rats in experimental groups I and II were
stimulated using the SDZ-II nerve and muscle stimulator (Huatuo;
Suzhou Medical Appliance Factory) with an intensity of 2 mA and a
frequency of 2 Hz for 15 and 30 min, respectively. The positively
and negatively charged clips were connected to the right and left
needles of the bilateral knee joints for 15 or 30 min each day for
3 days.
Preparation of serum
Arterial blood was collected from the abdominal
aorta. Processing and storage of blood was conducted in accordance
with a previously described method (16). The collected blood was placed at
room temperature for 4 h and centrifuged at 1,500 × g for 15 min at
room temperature. The serum fraction was isolated and was then
filtered through a 0.22 µm filter. The resulting EAS was then
aliquoted and stored at −20°C.
Chondrocyte culture and
identification
Chondrocytes were isolated (stepwise 0.2%
collagenase type II digestion) from the knee cartilage of
4-week-old male Sprague Dawley rats (weight, 80±10 g), which were
purchased from the Shanghai Slack Laboratory Animal Co., Ltd.
(certification no. SCXK: 2012-0001). The chondrocytes were cultured
and identified by collagen type II immunohistochemistry (17–19).
Briefly, chondrocytes from the knee articular cartilage of
4-week-old rats were isolated using 0.2% type II collagenase
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) in magnesium- and
calcium-free PBS (pH 7.4; HyClone; GE Healthcare, Logan, UT, USA)
for 1 h at 37°C. The cells were then resuspended in Dulbecco's
modified Eagle's medium (DMEM; HyClone; GE Healthcare) supplemented
with 10% FBS (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA), 100 µg/ml streptomycin and 100 U/ml penicillin (HyClone; GE
Healthcare), and were seeded in a monolayer at a density of
5×105 cells/cm2. Third generation
chondrocytes were seeded on cover slips and cultured for 72 h, the
cover slips were then rapidly washed in ethanol, dried and mounted
for microscopic examination. Subsequently, the chondrocytes were
incubated with rat monoclonal antibodies against type II collagen
(BS1071; 1:100; Bioworld Technology, Inc., St. Louis Park, MN, USA)
at 4°C overnight. The expression of type II collagen was observed
using a phase-contrast microscope (BH2; Olympus Corporation, Tokyo,
Japan). The primary chondrocytes were termed passage (P)1, and the
P3 chondrocytes were used in subsequent experiments.
EAS treatment of chondrocytes
P3 chondrocytes were seeded into 6-well plates at a
density of 1.3×105/ml, and were incubated in DMEM
(HyClone; GE Healthcare) supplemented with 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.) for 72 h at 37°C in a 5% CO2
incubator. The cells were divided into four groups: Control group,
which was treated with 10% serum (100 µl/ml) obtained from the
blank group; model group, which was cotreated with 10 µg/l TNF-α
and 10% serum (100 µl/ml) obtained from the blank group; EAS I
group, which was cotreated with 10 µg/l TNF-α and 10% serum (100
µl/ml) obtained from experimental group I; and EAS II group, which
was cotreated with 10 µg/l TNF-α and 10% serum (100 µl/ml) obtained
from experimental group II. All groups were treated for 24, 48, and
72 h at 37°C in a 5% CO2 incubator.
Assessment of cell viability
Following treatment with EAS, the chondrocytes were
seeded into 6-well plates at a density of 1.3×105/ml for
24 h. Following treatment with or without TNF-α and EAS for 24, 48
and 72 h, the medium was then discarded and 100 µl 0.5% MTT
solution was added to replace the medium. Following a 4 h
incubation at 37°C, the wells were emptied, and 150 µl dimethyl
sulfoxide was added to the plate and agitated for 10 min. The
absorbance was measured at 490 nm using an ELISA reader (BioTek
Instruments, Inc., Winooski, VT, USA), and the means were
calculated.
Observation of morphologic
alterations
The cells were cultured in 6-well plates at a
density of 1.3×105/ml for 24 h. Following treatment with
or without TNF-α and EAS for 48 h, cell morphology was observed
using a phase-contrast microscope (BH2; Olympus Corporation).
ELISA
The concentration of interleukin (IL)-1β in
conditioned medium was measured using an ELISA kit (RLB00; R&D
Systems, Inc., Minneapolis, MN, USA) according to the
manufacturer's protocol. Briefly, following treatment with or
without EAS for 48 h at 37°C in a 5% CO2 incubator, the
cell culture medium was collected and centrifuged at 1,500 × g for
15 min at room temperature to remove cell fragments, and IL-1β
concentration was determined. Absorbance of the samples was
measured using a microplate spectrophotometer (Omega Bio-Tek, Inc.,
Norcross, GA, USA). A standard curve was generated, from which
IL-1β concentration was determined.
Assessment of chondrocyte apoptosis by
DAPI staining
Treated chondrocytes were collected and fixed in 4%
paraformaldehyde for 15 min. Subsequently, the cells were stained
with 5 µg/ml DAPI for 5 min and washed three times with PBS. The
cells were then observed under a Fluo-View confocal fluorescent
microscope (Fluo-View FV10i; Olympus Corporation). A total of 10
visual fields in each group were randomly selected and the number
of apoptotic chondrocytes in each of these fields was counted using
ZEN 2009 Light Edition (Carl Zeiss AG, Oberkochen, Germany).
Western blot analysis
Total proteins were isolated from cells using
radioimmunoprecipitation assay lysis buffer (P0013B; Beyotime
Institute of Biotechnology, Shanghai, China), after which they were
stored on ice for 30 min and the protein concentration was
determined using the bicinchoninic acid assay (P0009; Beyotime
Institute of Biotechnology). Protein (20 µg) was separated by 10 or
12% SDS-PAGE. Following electrophoresis, proteins were transferred
to polyvinylidene fluoride membranes using a semidry blotting
system, and the membranes were blocked with 5% w/v nonfat dry milk
for 1 h at room temperature (20).
The membranes were then incubated with rat monoclonal antibodies
against Ras (ab52939; 1:5,000), Raf (ab33899; 1:2,000), MEK1/2
(ab178876; 1:5,000) and β-actin (ab6276; 1:1,000) (Abcam,
Cambridge, UK), and ERK1/2 (4695s; 1:1,000) and p-ERK1/2 (4094s;
1:1,000) (Cell Signaling Technology, Inc., Beverly, MA, USA) at 4°C
overnight, followed by incubation with horseradish
peroxidase-conjugated secondary antibodies (ZB-2301; 1:5,000;
OriGene Technologies, Inc., Beijing, China; or bs-0296G; 1:5,000;
BIOSS, Beijing, China) for 1 h at room temperature. Blots were
visualized using Pierce™ Enhanced Chemiluminescence Western
Blotting Substrate (32106; Thermo Fisher Scientific, Inc.). The
intensity of each band was semi-quantified using the Image Lab gel
analyzing system (Image Lab 3.0™ Software; Bio-Rad Laboratories,
Inc., Hercules, CA, USA) and was normalized to the band intensity
of β-actin.
Laser confocal scanning
microscopy
Following treatment, the cells were fixed in
ice-cold methanol and permeabilized with 1% Triton X-100 for 10
min, after which they were blocked with 5% bovine serum albumin
(A8010; Beijing Solarbio Science & Technology Co., Ltd.,
Beijing, China) for 1 h at room temperature. The fixed cells were
washed and incubated with rat monoclonal antibodies against matrix
metalloproteinase (MMP)-3 (sc-30070; 1:100; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) and MMP-13 (PA5-16566; 1:100;
Thermo Fisher Scientific, Inc.) at 4°C overnight, followed by
incubation with Alexa Fluor 488 goat anti-rabbit immunoglobulin G
(H+L) secondary antibodies (A-11008; 1:300; Thermo Fisher
Scientific, Inc.) and DAPI (D1306; 1:100; Thermo Fisher Scientific,
Inc.) for 5 min at room temperature in the dark. The signal was
visualized and images were acquired using a laser confocal scanning
microscope (Olympus Corporation).
Statistical analysis
All data were collected from at least three
independent experiments. Statistical analysis was performed using
SPSS 20.0 software (IBM Corp., Armonk, NY, USA). All data are
presented as the mean ± standard deviation. The differences among
the four groups were compared using one-way analysis of variance,
and multiple comparisons were performed using the
Student-Newman-Keuls-q test. P<0.05 was considered to indicate a
statistically significant difference.
Results
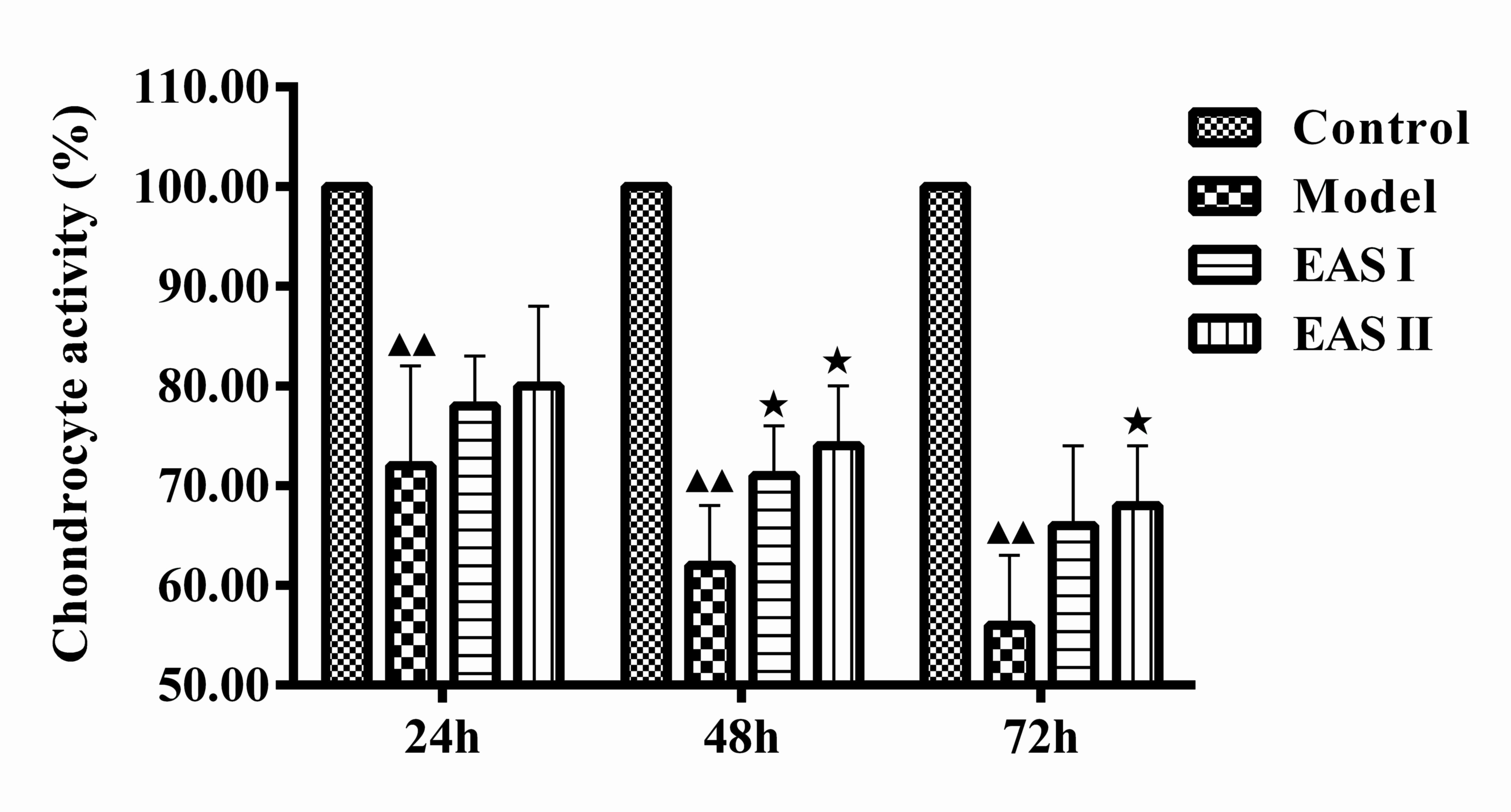
EAS enhances chondrocyte
viability
The present study examined whether EAS could promote
the viability of TNF-α-treated chondrocytes using an MTT assay. The
groups were treated with or without EAS for various durations. The
results demonstrated that the viability of the model group was
significantly lower than that of the control group (P<0.01), and
the viability of the EAS I and II groups was significantly higher
than that of the model group after 48 h (P<0.05; Fig. 1). Therefore, EAS treatment for 48 h
was used in the subsequent experiments.
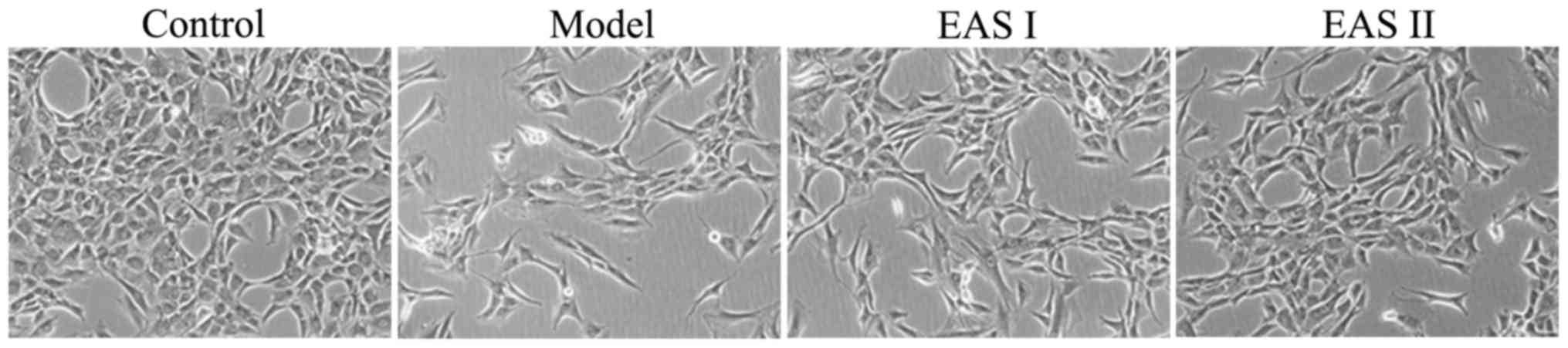
Effects of EAS on the morphology of
TNF-α-treated chondrocytes
To determine the effects of EAS treatment on the
morphology of cells, the morphological alterations of the cells in
the various groups were observed by phase-contrast microscopy
(Fig. 2). The morphology of the
untreated control cells exhibited a healthy status, whereas
TNF-α-treated chondrocytes presented more apoptotic cells that
detached from each other and became bright, elongated and shrunken,
or floated in the medium, as compared with cells in the EAS I and
II groups. However, the TNF-α-induced alterations in cell
morphology were not observed, or were less evident, in the
TNF-α-stimulated chondrocytes treated with EAS.
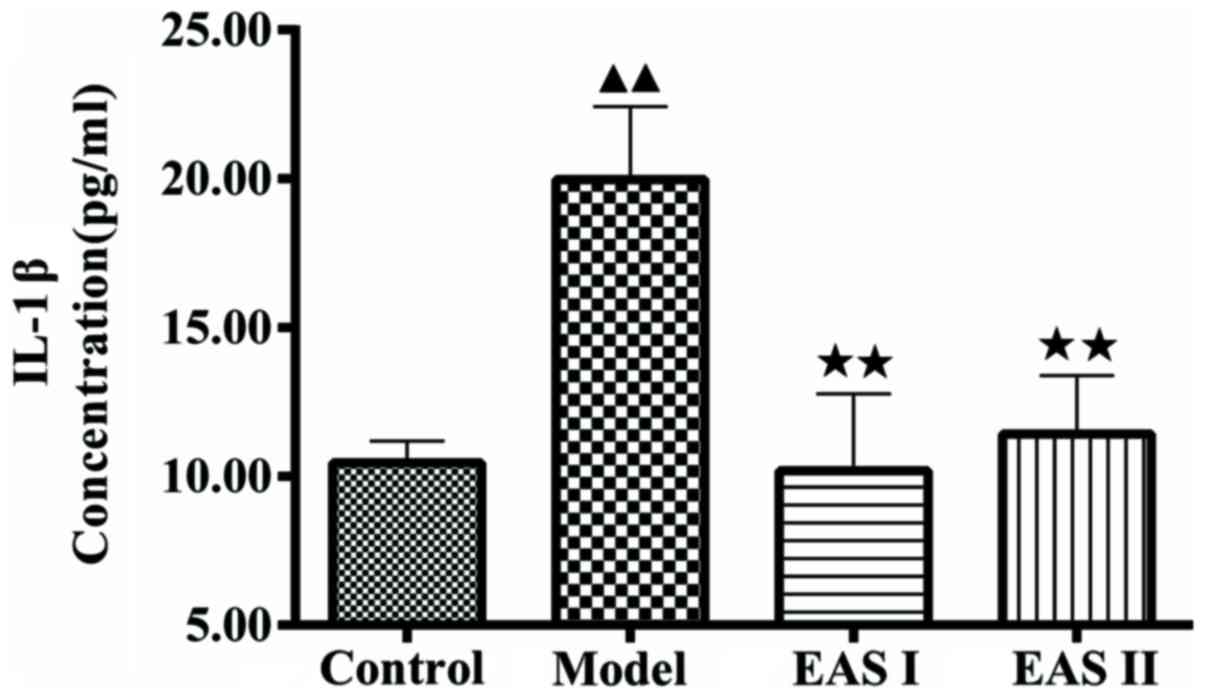
EAS inhibits IL-1β and apoptosis of
TNF-α-treated chondrocytes
To examine the effects of EAS on inflammation in
TNF-α-treated chondrocytes, an ELISA was used to measure IL-1β
concentration. As presented in Fig.
3, IL-1β concentration in the model group was significantly
higher compared with in the control group (P<0.01); however,
treatment with EAS significantly reduced IL-1β concentration
compared with the model group (P<0.01). To further determine
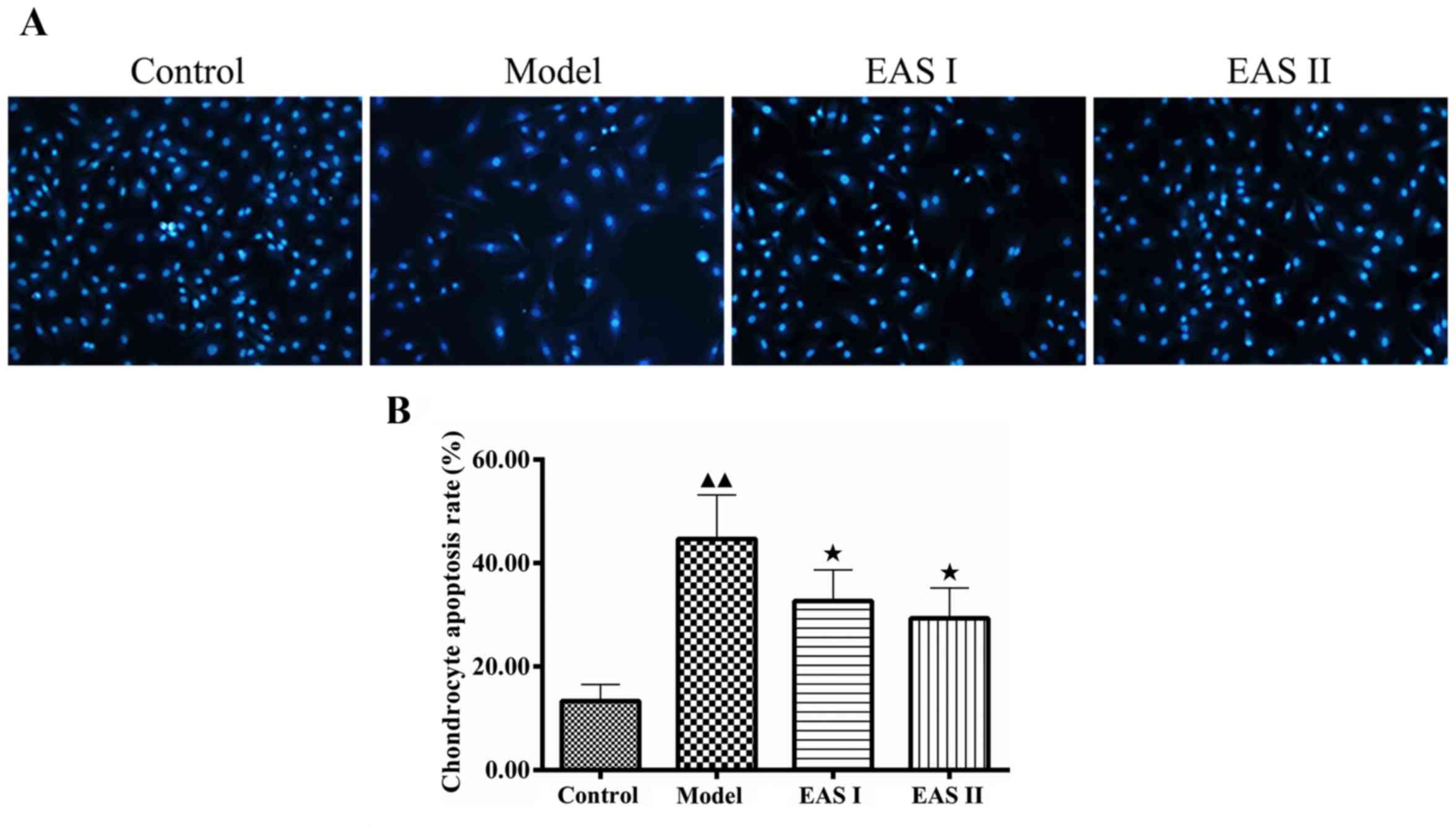
whether EAS inhibited inflammation via apoptotic processes, DAPI
staining was used to assess chondrocyte apoptosis. Apoptotic cells
exhibited typical alterations, including reduced cellular volume,
bright blue staining, and condensed or fragmented nuclei. This
phenomenon was more obvious in the model group compared with in the
EAS I and II groups (Fig. 4). The
percentage of apoptotic cells in the EAS groups was significantly
lower than in the model group (P<0.05), thus suggesting that EAS
may inhibit the apoptosis of TNF-α-treated chondrocytes via the
regulation of IL-1β.
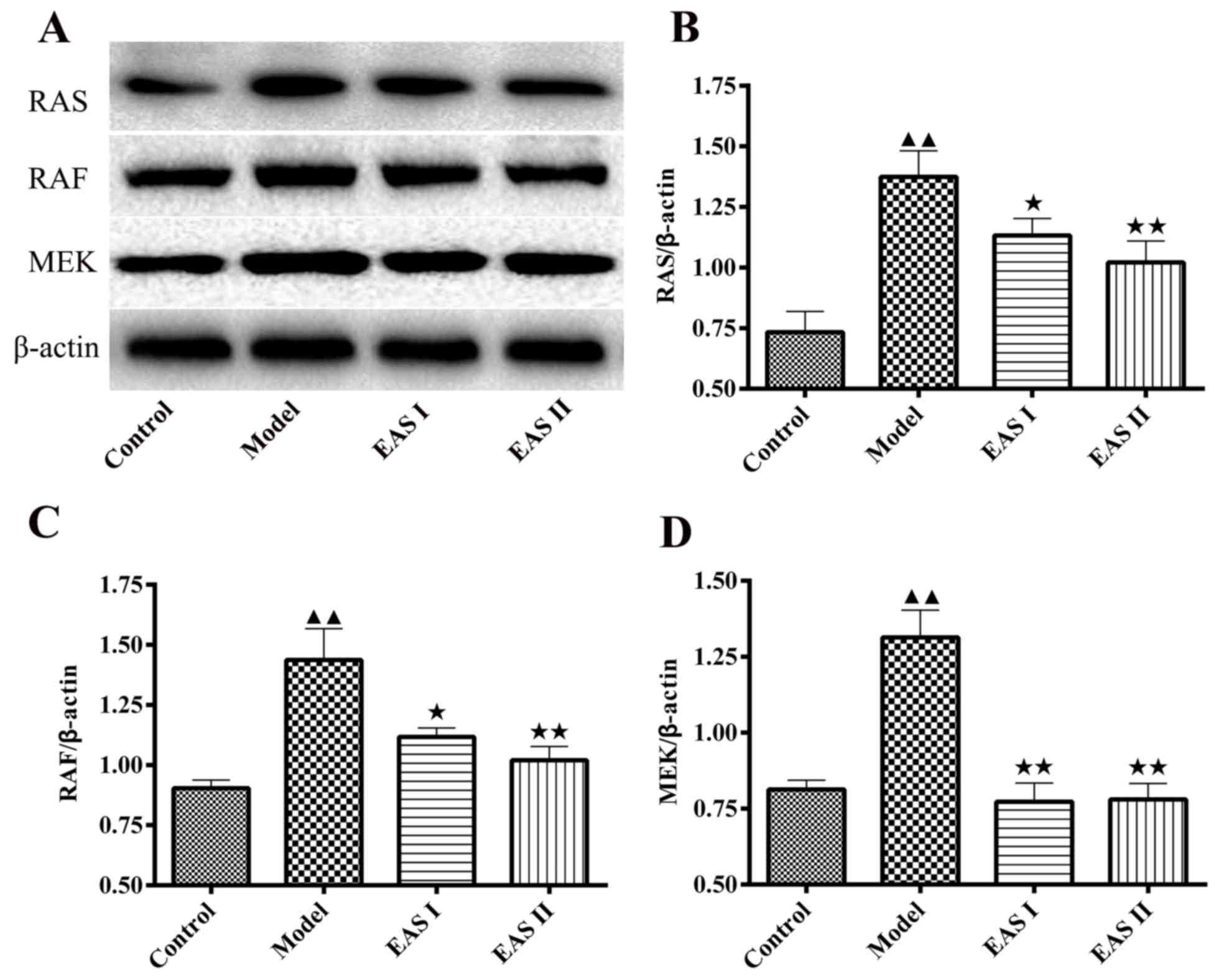
EAS inhibits the protein expression
levels of Ras, Raf, MEK1/2 and p-ERK1/2 in TNF-α-treated
chondrocytes
To gain insight into the mechanisms underlying the
effects of EAS on the inflammation of TNF-α-treated chondrocytes,
the protein expression levels of Ras, Raf and MEK1/2 were detected.
The results demonstrated that the protein expression levels of Ras,
Raf and MEK1/2 were lower in the EAS I and II groups compared with
in the model group (P<0.05; Fig.
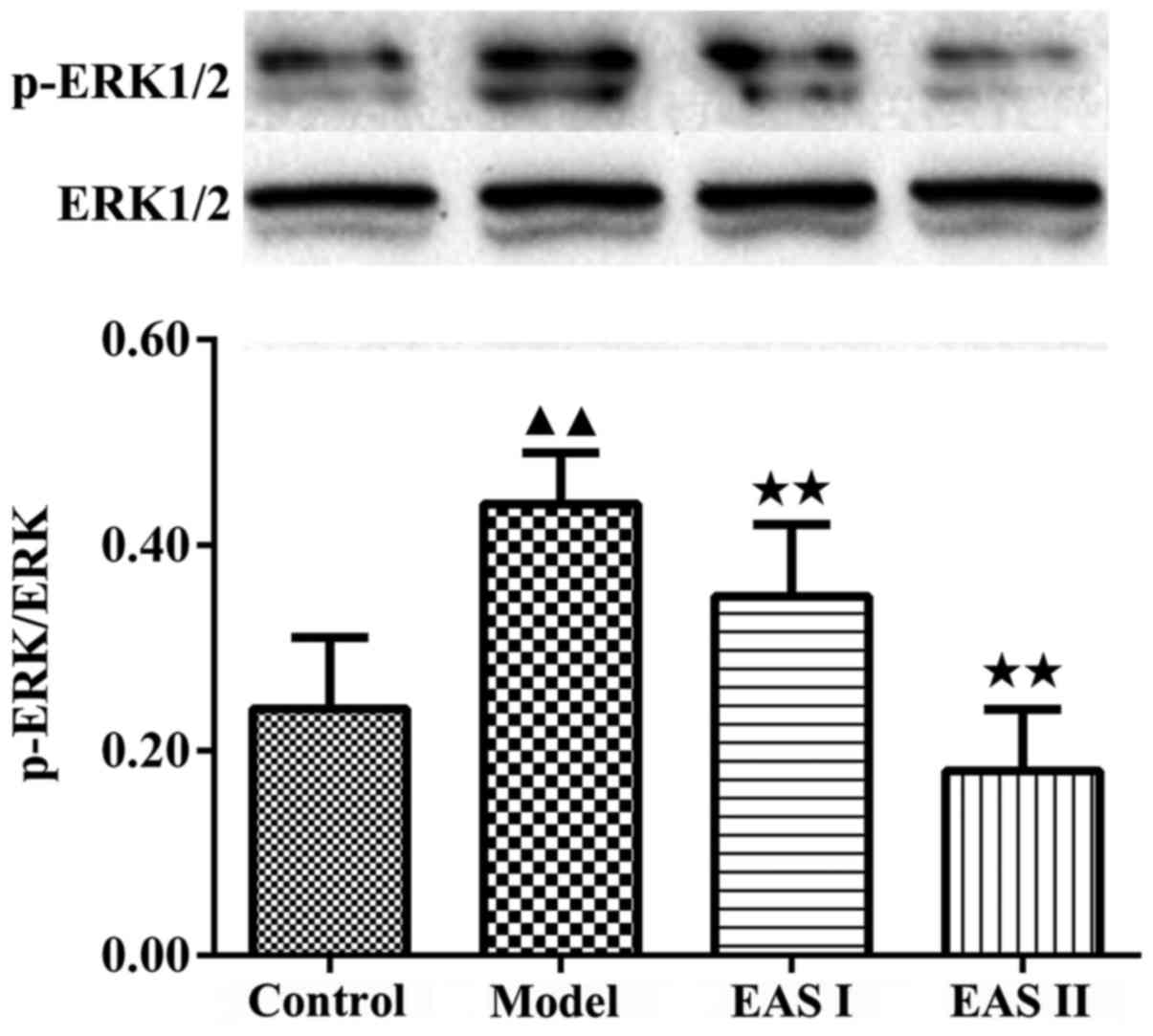
5). Furthermore, activation of the ERK1/2 signaling pathway has
been reported to participate in chondrocyte inflammation induced by
various stimuli. p-ERK1/2 is an intracellular signaling molecule
that transduces extracellular responses, serves a well-known role
in regulating chondrocyte inflammation and contributes to loss of
the chondral matrix (21,22). Therefore, to understand the
molecular mechanism by which EAS inhibits TNF-α-induced
inflammation, the expression of p-ERK1/2 was detected. The results
demonstrated that the protein expression levels of p-ERK1/2 were
significantly reduced in the EAS groups compared with in the model
group (P<0.01; Fig. 6). Taken
together, these results indicated that EAS may inhibit
TNF-α-mediated chondrocyte inflammation via the
Ras-Raf-MEK1/2-ERK1/2 signaling pathway.
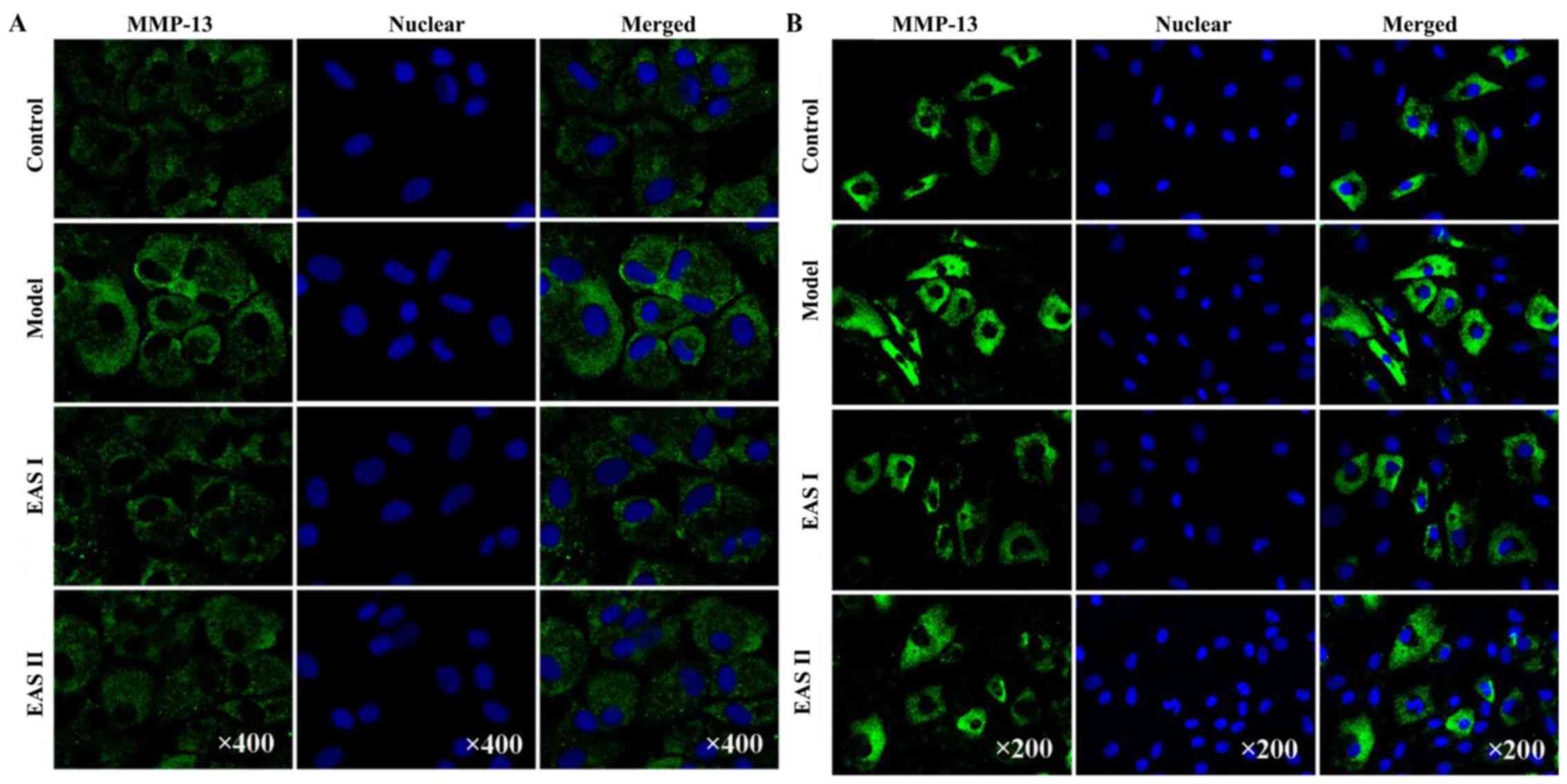
EAS decreases the protein expression
levels of MMP-3 and MMP-13 in TNF-α-treated chondrocytes
It is well known that MMPs serve a critical role in
OA. Furthermore, a previous study reported that EA inhibited the
expression of MMP-3 in chondrocytes (23). Therefore, the present study
investigated the effects of EAS on the protein expression of MMP-3
and MMP-13 using immunofluorescent staining. Chondrocytes
stimulated with TNF-α exhibited an increased release of MMP-3 and
MMP-13 compared with the untreated controls. However, treatment of
chondrocytes with EAS markedly inhibited the TNF-α-mediated release
of MMP-3 and MMP-13 (Fig. 7),
which indicated that EAS may inhibit the Ras-Raf-MEK1/2-ERK1/2
pathway, resulting in reduced MMP-3 and MMP-13 expression.
Discussion
EA is a modern modification of the traditional
acupuncture method, which stimulates acupuncture points with
electrical current instead of manual manipulations and has been
used to treat OA. Previous studies have demonstrated that EA
treatment is able to induce a series of changes in serum (24–26).
The present study used EAS to explore the anti-inflammatory effects
and underlying mechanisms of EA on OA. EA treatment leads to the
production of a series of factors involved in the regulation of
cartilage inflammation. These factors are translocated to tissues
and cells through the blood circulation; therefore, cells treated
with EAS in vitro are in a similar condition to in
vivo cells (27,28). The treatment of cells in
vitro with EAS is easily regulated and effects are easily
detected; therefore, EAS treatment is convenient in cell and
molecular biology, as it can reveal the mechanisms underlying
acupuncture treatment (29,30).
In the present study, chondrocytes were treated with EAS with the
aim to further investigate the potential mechanisms underlying the
effects of EAS treatment on the regulation of chondrocyte
function.
Inflammation leads to cartilage degradation;
therefore, inhibiting chondrocyte inflammation may be an efficient
method for the treatment of OA. Previous studies have reported that
TNF-α is a potent proinflammatory cytokine that induces apoptosis
of chondrocytes, which had been enzymatically dissociated from OA
knee cartilage (31,32). In order to study the effects of EAS
treatment on TNF-α-mediated chondrocyte inflammation, a chondrocyte
culture was established in vitro via stepwise 0.2%
collagenase type II digestion; subsequently, the chondrocytes were
treated with 10 µg/l TNF-α for various durations. The results of
the present study are similar to those of a previous study, which
indicated that TNF-α suppresses viability and induces apoptotic
signaling in chondrocytes ex vivo (33). Furthermore, IL-1β serves a crucial
role in the progression of OA, where it induces cartilage damage
through modulating the expression of MMPs, and promotes
proteoglycan degradation and apoptosis (34). In the present study, IL-1β
concentration was increased in TNF-α-treated chondrocytes; however,
treatment with EAS significantly reduced the concentration of IL-1β
in TNF-α-treated chondrocytes. Furthermore, EAS significantly
reduced the percentage of TNF-α-mediated apoptotic chondrocytes,
thus suggesting that EA may prevent the degradation of articular
cartilage by inhibiting chondrocyte inflammation. The concentration
of TNF-α (10 µg/l) used in the present study was based on the
concentration range of TNF-α (5–40 µg/l) used in a previous study,
which focused on TNF-α-mediated inflammation in chondrocytes in
vitro (33).
The Ras-Raf-MEK1/2-ERK1/2 signaling pathway is the
best characterized of three mitogen-activated protein kinase
pathways that transduce extracellular signals through an
intracellular signal transduction cascade. In vitro, IL-1β
leads to the activation of ERK; activation of the Erk1/2 signaling
pathway can induce numerous protein kinase cascade reactions, and
transmits extracellular signals into the cells. Under the
stimulation of extracellular signals, Ras can be activated by
binding with guanosine triphosphate (35). Through a complex series of events,
activated Ras then directs plasma membrane recruitment and
activation of Raf. Through the formation of homo- and heterodimers,
Raf propagates downstream signaling by activating MEK1/2 at one of
two serine residues. Subsequently, MEK1/2 propagates the signal by
phosphorylating ERK at two threonine residues (36,37).
In accordance with previous reports, the present study demonstrated
that TNF-α increased p-ERK1/2 expression in chondrocytes (38). It has previously been reported that
EA may reduce the protein expression levels of p-ERK1/2 in cells
(39). The present study
demonstrated that EAS was able to decrease the protein expression
levels of p-ERK1/2 in TNF-α-treated chondrocytes. p-ERK1/2 enters
the nuclei and triggers the activity of transcription factors and
MMPs, including MMP-3 and MMP-13, causing a series of cellular
responses that can regulate cell apoptosis (40–43).
Therefore, it may be hypothesized that cartilage degradation can be
reduced via inhibiting the activity of MMPs. The present study
demonstrated that EAS markedly decreased the protein expression of
MMP-3 and MMP-13 in TNF-α-treated chondrocytes.
In conclusion, EAS treatment efficiently suppressed
inflammation by inhibiting activation of the Ras-Raf-MEK1/2-ERK1/2
signaling pathway and the expression of MMP-3 and MMP-13 in
TNF-α-treated chondrocytes. Further studies are required to
investigate the association between EAS and other signals involved
in the inflammation of chondrocytes. An enhanced understanding of
the underlying molecular mechanisms of EAS will aid the improvement
of diagnoses and the development of novel therapeutic targets for
the treatment of OA-associated inflammation.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (grant no.
81373719).
References
|
1
|
van den Berg WB: Osteoarthritis year 2010
in review: Pathomechanisms. Osteoarthritis Cartilage. 19:338–341.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Huang Z, Li J, Du S, Chen G, Qi Y, Huang
L, Xiao L and Tong P: Effects of UCP4 on the proliferation and
apoptosis of chondrocytes: Its possible involvement and regulation
in osteoarthritis. PLoS One. 11:e1506842016.
|
|
3
|
Li X, Liu C, Liang W, Ye H, Chen W, Lin R,
Li Z, Liu X and Wu M: Millimeter wave promotes the synthesis of
extracellular matrix and the proliferation of chondrocyte by
regulating the voltage-gated K+ channel. J Bone Miner
Metab. 32:367–377. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Thomas CM, Fuller CJ, Whittles CE and
Sharif M: Chondrocyte death by apoptosis is associated with
cartilage matrix degradation. Osteoarthritis Cartilage. 15:27–34.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Loeser RF: Aging and osteoarthritis: The
role of chondrocyte senescence and aging changes in the cartilage
matrix. Osteoarthritis Cartilage. 17:971–979. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hussein MR, Fathi NA, El-Din AM, Hassan
HI, Abdullah F, Al-Hakeem E and Backer EA: Alterations of the
CD4(+), CD8(+) T cell subsets, interleukins-1beta, IL-10, IL-17,
tumor necrosis factor-alpha and soluble intercellular adhesion
molecule-1 in rheumatoid arthritis and osteoarthritis: Preliminary
observations. Pathol Oncol Res. 14:321–328. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gui L, Zeng Q, Xu Z, Zhang H, Qin S, Liu
C, Xu C, Qian Z, Zhang S, Huang S and Chen L: IL-2, IL-4, IFN-γ or
TNF-α enhances BAFF-stimulated cell viability and survival by
activating Erk1/2 and S6K1 pathways in neoplastic B-lymphoid cells.
Cytokine. 84:37–46. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Clausen BH, Degn M, Sivasaravanaparan M,
Fogtmann T, Andersen MG, Trojanowsky MD, Gao H, Hvidsten S, Baun C,
Deierborg T, et al: Conditional ablation of myeloid TNF increases
lesion volume after experimental stroke in mice, possibly via
altered ERK1/2 signaling. Sci Rep. 6:292912016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hu C, Yang H, Zhao Y, Chen X, Dong Y, Li
L, Dong Y, Cui J, Zhu T, Zheng P, et al: The role of inflammatory
cytokines and ERK1/2 signaling in chronic prostatitis/chronic
pelvic pain syndrome with related mental health disorders. Sci Rep.
6:286082016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hinman RS, McCrory P, Pirotta M, Relf I,
Forbes A, Crossley KM, Williamson E, Kyriakides M, Novy K, Metcalf
BR, et al: Acupuncture for chronic knee pain: A randomized clinical
trial. JAMA. 312:1313–1322. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
White A and Cummings M: Acupuncture for
knee osteoarthritis: Study by Hinman et al represents missed
opportunities. Acupunct Med. 33:84–86. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen X, Spaeth RB, Retzepi K, Ott D and
Kong J: Acupuncture modulates cortical thickness and functional
connectivity in knee osteoarthritis patients. Sci Rep. 4:64822014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu MX, Li XH, Lin MN, Jia XR, Mu R, Wan
WR, Chen RH, Chen LH, Lin WQ, Huang CY, et al: Clinical study on
the treatment of knee osteoarthritis of Shen-Sui insufficiency
syndrome type by electroacupuncture. Chin J Integr Med. 16:291–297.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Plaster R, Vieira WB, Alencar FA, Nakano
EY and Liebano RE: Immediate effects of electroacupuncture and
manual acupuncture on pain, mobility and muscle strength in
patients with knee osteoarthritis: A randomised controlled trial.
Acupunct Med. 32:236–241. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang Y, Wu G, Fan H, Ye J and Liu X:
Electroacupuncture promotes chondrocyte proliferation via
accelerated G1/S transition in the cell cycle. Int J Mol Med.
31:1443–1448. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu G, Fan H, Huang Y, Zheng C, Ye J and
Liu X: Duhuo Jisheng Decoction-containing serum promotes
proliferation of interleukin-1β-induced chondrocytes through the
p16-cyclin D1/CDK4-Rb pathway. Mol Med Rep. 10:2525–2534. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li X, Du M, Liu X, Chen W, Wu M, Lin J and
Wu G: Millimeter wave treatment promotes chondrocyte proliferation
by upregulating the expression of cyclin-dependent kinase 2 and
cyclin A. Int J Mol Med. 26:77–84. 2010.PubMed/NCBI
|
|
18
|
Li H, Li X, Liu G, Chen J, Weng X, Liu F,
Xu H, Liu X and Ye H: Bauhinia championi (Benth.) Benth.
Polysaccharides upregulate Wnt/β-catenin signaling in chondrocytes.
Int J Mol Med. 32:1329–1336. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li XH, Wu MX, Ye HZ, Chen WL, Lin JM,
Zheng LP and Liu XX: Experimental study on the suppression of
sodium nitroprussiate-induced chondrocyte apoptosis by Tougu
Xiaotong capsule containing serum. Chin J Integr Med. 17:436–443.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu F, Weng X, Lin P, Zheng C, Xu H, Liu
X, Ye H and Li X: Duhuo Jisheng decoction inhibits endoplasmic
reticulum stress in chondrocytes induced by tunicamycin through the
downregulation of miR-34a. Int J Mol Med. 36:1311–1318. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim KS, Oh DH, Choi HM, Bang JS, Ryu CJ,
Kim JH, Yoo MC and Yang HI: Pyrrolidine dithiocarbamate, a
NF-kappaB inhibitor, upregulates MMP-1 and MMP-13 in
IL-1beta-stimulated rheumatoid arthritis fibroblast-like
synoviocytes. Eur J Pharmacol. 613:167–175. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li X, Li J, Cheng K, Lin Q, Wang D, Zhang
H, An H, Gao M and Chen A: Effect of low-intensity pulsed
ultrasound on MMP-13 and MAPKs signaling pathway in rabbit knee
osteoarthritis. Cell Biochem Biophys. 61:427–434. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liang CX, Guo Y, Tao L, Xiao H, Liu QG, Ma
HF and Guo CQ: Effects of acupotomy intervention on regional
pathological changes and expression of carti-lage-mechanics related
proteins in rabbits with knee osteoarthritis. Zhen Ci Yan Jiu.
40119–124. (140)2015.(In Chinese). PubMed/NCBI
|
|
24
|
Yang EJ, Jiang JH, Lee SM, Hwang HS, Lee
MS and Choi SM: Electroacupuncture reduces neuroinflammatory
responses in symptomatic amyotrophic lateral sclerosis model. J
Neuroimmunol. 223:84–91. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liao HY, Sun MF, Lin JG, Chang SL and Lee
YC: Electroacupuncture plus metformin lowers glucose levels and
facilitates insulin sensitivity by activating MAPK in
steroid-induced insulin-resistant rats. Acupunct Med. 33:388–394.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liao Y, Li X, Li N and Zhou J:
Electroacupuncture protects against articular cartilage erosion by
inhibiting mitogen-activated protein kinases in a rat model of
osteoarthritis. Acupunct Med. 34:290–295. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pavão TS, Vianna P, Pillat MM, Machado AB
and Bauer ME: Acupuncture is effective to attenuate stress and
stimulate lymphocyte proliferation in the elderly. Neurosci Lett.
484:47–50. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim SK and Bae H: Acupuncture and immune
modulation. Auton Neurosci. 157:38–41. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu M, Li X, Li L, Liu B and Lin J: Effect
of electro acupuncture serum on MAPK signal pathway of apoptosis
chondrocyte induced by TNF-α. Fujian J TCM. 42:43–45. 2011.
|
|
30
|
Wu M, Li X, Wu G, Li L, Chen W and Liu X:
Effects of serum on tumor necrosis factor alpha induced chondrocyte
apoptosis following electro-acupuncture. J Clin Rehabilitative
Tissue Engineering Res. 15:8551–8555. 2011.
|
|
31
|
Markway BD, Cho H, Anderson DE, Holden P,
Ravi V, Little CB and Johnstone B: Reoxygenation enhances tumour
necrosis factor alpha-induced degradation of the extracellular
matrix produced by chondrogenic cells. Eur Cell Mater. 31:425–439.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Malemud CJ, Sun Y, Pearlman E, Ginley NM,
Awadallah A, Wisler BA and Dennis JE: Monosodium Urate and tumor
necrosis factor-α increase apoptosis in human chondrocyte cultures.
Rheumatology (Sunnyvale). 2:1132012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li X, Wu G, Wu M, Chen W and Liu X: In
vitro study of inhibitory millimeter wave treatment effects on the
TNF-α-induced NF-κB signal transduction pathway. Int J Mol Med.
27:71–78. 2011.PubMed/NCBI
|
|
34
|
Lu S, Xiao X and Cheng M: Matrine inhibits
IL-1β-induced expression of matrix metalloproteinases by
suppressing the activation of MAPK and NF-κB in human chondrocytes
in vitro. Int J Clin Exp Pathol. 8:4764–4772. 2015.PubMed/NCBI
|
|
35
|
Wang X, Li F, Fan C, Wang C and Ruan H:
Effects and relationship of ERK1 and ERK2 in interleukin-1β-induced
alterations in MMP3, MMP13, type II collagen and aggrecan
expression in human chondrocytes. Int J Mol Med. 27:583–589.
2011.PubMed/NCBI
|
|
36
|
Uehling DE and Harris PA: Recent progress
on MAP kinase pathway inhibitors. Bioorg Med Chem Lett.
25:4047–4056. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xu WH, Zhang JB, Dang Z, Li X, Zhou T, Liu
J, Wang DS, Song WJ and Dou KF: Long non-coding RNA URHC regulates
cell proliferation and apoptosis via ZAK through the ERK/MAPK
signaling pathway in hepatocellular carcinoma. Int J Biol Sci.
10:664–676. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Appleton CT, Usmani SE, Mort JS and Beier
F: Rho/ROCK and MEK/ERK activation by transforming growth
factor-alpha induces articular cartilage degradation. Lab Invest.
90:20–30. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen H, Peng B, Li FY, YU X, Lu HY and Wei
XY: Study on the effect of electro-acupuncture ouch point in
rabbits with muscle regeneration after contusion based on the ERK
signal pathway inhibited by U0126. China J Traditional Chin Med
Pharmacy. 29:2304–2308. 2014.
|
|
40
|
Chambard JC, Lefloch R, Pouysségur J and
Lenormand P: ERK implication in cell cycle regulation. Biochim
Biophys Acta. 1773:1299–1310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yu SM and Kim SJ: The thymoquinone-induced
production of reactive oxygen species promotes dedifferentiation
through the ERK pathway and inflammation through the p38 and PI3K
pathways in rabbit articular chondrocytes. Int J Mol Med.
35:325–332. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen J, Wang Z, Xu D, Liu Y and Gao Y:
Aquaporin 3 promotes prostate cancer cell motility and invasion via
extracellular signal-regulated kinase 1/2-mediated matrix
metalloproteinase-3 secretion. Mol Med Rep. 11:2882–2888. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhong HM, Ding QH, Chen WP and Luo RB:
Vorinostat, a HDAC inhibitor, showed anti-osteoarthritic activities
through inhibition of iNOS and MMP expression, p38 and ERK
phosphorylation and blocking NF-κB nuclear translocation. Int
Immunopharmacol. 17:329–335. 2013. View Article : Google Scholar : PubMed/NCBI
|