Introduction
Bovine mastitis can directly result in the reduction
of milk quality and production leading to a large economic loss
(1). Gram-negative (GN) bacterial
infection is the main reason for bovine mastitis, which leads to
acute mastitis with clinical symptoms (2–4).
Lipopolysaccharide (LPS) is a key component of the outer membrane
of all GN bacteria, which can be secreted by them under different
physiological conditions (5,6). LPS
can stimulate secretion of cytokines-mediated inflammation pathways
(7). In addition, mammals, through
a cluster of differentiation (CD)14-dependent pathway can detect
LPS concentration, which activates host cells to start an
inflammatory response for bacteria resistance (6).
CD14 serves as a receptor for the complex of
LPS-liposaccharide binding protein (LPS-LBP) complex in animals
(8,9). When LPS is released from bacterial
outer membrane and intrudes into the host organism, CD14 binds to
LPS-LBP to form the LPS-LBP-CD14 complex (10). Subsequently, macrophages can secret
inflammatory cytokines such as tumor necrosis factor-α (TNF-α) and
interleukins (IL)-1β, −6 and −8 (11) activating the toll-like receptor-4
(TLR-4) signaling pathway. As a result, this binding induces an
innate defense response against GN bacteria (12). Investigation of CD14 involved in
the activation of the TLR signaling pathway may contribute to
blocking the inflammatory cascade prior to inflammatory cytokines
release (13).
In addition, it is well known that there are two
forms of CD14, a membrane-bound CD14 (mCD14) and a soluble one
(sCD14), which serves inessential role in the recognition of GN
bacteria. mCD14 is expressed on the cell surface of monocytes,
macrophages and polymorphonuclear neutrophils (PMNs) (9–11).
Under low LPS concentrations, mCD14 activate phagocytes in the
presence of LBP. Notably, sCD14 is only present in body fluid such
as the serum or breast milk (7,8).
Either mCD14 or sCD14 can directly strengthen LPS response and
associated signaling pathways in epithelial and endothelial cells,
even in the absence of mCD14 (14–17).
sCD14 binding to LPS can stimulate innate response to GN bacterial
infection under low concentrations (6,17),
alleviate stress induced by mastitis and weakens LPS toxicity
(8). Previous studies revealed the
difference in susceptibility to GN bacterial infection between
dairy cows (18) and the
polymorphisms in the coding and promoter region of CD14
gene-associated with the risk of several diseases in humans and
animals (11,19–22).
Therefore, it seems that genetic variations in cow CD14 may affect
the risk for GN bacterial infections (23). However, little is known about the
correlation between CD14 polymorphisms and mastitis. For this
reason, the single-strand conformation polymorphism (SSCP) method
was used to analyze CD14 polymorphisms and their correlation with
clinical mastitis in Chinese Holstein, which will be important for
breeding anti-mastitis dairy cows.
Materials and methods
Animal selection, ethical approval and
genomic DNA extraction
In the present study, 134 dairy cows (Chinese
Holstein, Wuhu Dairy Co., Ltd., Wuhu, China) had complete records
of Dairy Herd Improvement. They shared the same season (Spring,
February) of the last deliveries and physical conditions (2 years
old, 2 fetus, same lactation period). They were raised under the
same housing and management conditions, in which 66 cows were
regarded as clinical mastitis cases defined by the red and swollen
udder and somatic cell count (SCC) of milk (SCC >1,000,000/ml).
These data were used to evaluate animal status (24). The other 68 cows without clinical
symptoms were regarded as non-mastitis cows. Animals and their care
protocols followed a previous study (5) and were approved by the Animal Care
and Use Committee of Nanjing Agricultural University and Wuhu Dairy
Ltd (approval no. 20150930). Blood samples were harvested from
caudal veins of cattle and stored at −81°C. DNA extraction from the
blood samples was followed by the conventional phenol-chloroform
method. DNA was dissolved in TE buffer (made in our laboratory) and
stored at −20°C until further use.
Primer design and polymerase chain
reaction (PCR)
Primers were designed to amplify coding sequence of
CD14 gene (NCBI Reference Sequence: AC_000164.1). PCR
reaction contained: 2.5 µl of 10XPCR buffer (Takara Biotechnology
Co., Ltd., Dalian, China) 1.5 µl Mg2+ (25 mM), 0.5 µl
dNTP (10 mM), 0.2 µl Taq DNA polymerase (0.5 U/0.5 µl; Takara
Biotechnology Co., Ltd.), 1 µl each primer (10 µM/µl), and 2 µl
template DNA (50 ng/µl) diluted with ddH2O to a final
volume of 25 µl. PCR amplification reaction program:
Pre-denaturation for 4 min at 94°C; then 35 cycles (denaturation
for 40 sec at 94°C, renaturation for 30 sec at annealing
temperature, elongation for 1 min at 72°C); finally elongation for
7 min at 72°C. The primer sequences, sequence site, predicted size
of segment and annealing temperatures are listed in Table I.
 | Table I.Primer pairs of CD14 gene designed
for its amplification. |
Table I.
Primer pairs of CD14 gene designed
for its amplification.
| Primer | Forward
(5′-3′) | Reverse
(5′-3′) | Annealing
temperature (°C) | Expected segment
size (bp) | Amplified site size
(bp) |
|---|
| 1 |
GTGAGCCACTGTAAAGGAAAGA |
TGGAAAGACAGCGGAGGT | 57.8 | 172 | −80–91 |
| 2 |
GGCTCTGAGAATCTACCGACTA |
ACTGCTTCGGGTTGGTGT | 54.4 | 352 |
−21–330 |
| 3 |
GCCGTTCAGTGTATGGTTGCCGTCG |
GGCACCTCCTGTTGTCCACGATACG | 54.4 | 330 | 234–563 |
| 4 |
AGGAACTGACGCTTGAGGA |
CGCCGAGACTGGGATTGT | 57.2 | 270 | 442–711 |
| 5 |
CAGGGTGCTGAACATTGCC |
GCGAGTTGTGGCTGAGGT | 57.2 | 266 | 602–867 |
| 6 |
GACCTCAGCCACAACTCGC |
GGCTCCAGGGTCCAGAAA | 55.4 | 243 |
849–1,091 |
| 7 |
GCTGCCCGAGGTAAATGA | TAGAAGGCTGGTTGGTT
GAG | 58 | 272 | 1,034–1,305 |
PCR-SSCP assay
PCR products (5 µl) were denatured with the
denaturation buffer solution (Takara Biotechnology Co., Ltd.) at
97°C for 7 min and put on ice for 20 min, separated with 30%
polyacrylamide gel at 200 V/12-15 h and stained with silver nitrate
(Tiangen Co., Ltd., Beijing) for 15–20 min at room temperature.
DNA sequencing and analysis of
different genotypes
PCR products were purified using a kit from Tiangen
Biotech Co., Ltd. (Beijing, China) and sent for sequencing and
analysis. The differences and polymorphism of different genotypes
were analyzed with MegAlign software version 2.0 (DNASTAR, Inc.,
Madison, WI, USA).
Prediction of protein and functional
site of CD14 protein
Protean software version 2.0 (DNASTAR, Inc.) was
applied to analyze the hydrophilicity and secondary structure. The
3-dimensional (3D) structure was also built through the Hopp-Woods
and Homology-based modeling method (25–28).
The modeling construction can be divided into two
steps: In the first, protein crystal structure data was selected
from the ExPDB database; in the second, protein sequences were
placed to the SWISS-MODEL website (www.expasy.org/swissmod/SWISS-MODEL.html).
Subsequently, the results of the analysis of homology modeling,
modification and optimization of the protein were obtained. The
quality (QMEAN-score) of 3D structure was evaluated in QMEAN
website (swissmodel.expasy.org/qmean) (29,30).
Coding sequences and functional sites were also predicted on the
ExPASy Molecular Biology Server (www.expasy.org/tools/). The website (http://prosite.expasy.org/) of PROSITE database was
used to assay protein kinase C (PKC) phosphorylation site. A
Z-score of the model was also analyzed with QMEAN including four
regular statistical aspects (C beta interaction, all-atom pair
wise, salvation and torsion angle energy). Finally, 3D structures
were presented with Cn3d 4.1 software (NIH, Bethesda, MD, USA).
Reverse transcription-quantitative PCR
(RT-qPCR)
Blood samples of mastitis cows were collected and
anti-coagulated with sodium heparin. Ficoll reagent (Shanghai
Shenggong Co., Ltd., Shanghai, China) was used to dissociate
lymphocytes from peripheral blood. Total RNA was extracted from
lymphocytes with TRIzol following manufacturer's protocol
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA), and
then SuperScript First-Strand Synthesis System (Invitrogen; Thermo
Fisher Scientific, Inc.) was applied to perform the reverse
transcription following manufacturer's protocol. The 2XiTaq
Universal SYBR-Green Supermix (Bio-Rad Laboratories Inc., Hercules,
CA, USA) was used to amplify the target genes. PCR reactions (40
cycles for 95°C for 15 sec and 60°C for 1 min) were run on ABI
StepOne machine (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The mRNA level of target genes was calculated with StepOne
software version 2.1, using 18s rRNA (18s rRNA kit, Ambion; Thermo
Fisher Scientific, Inc.) as internal control. The following primers
of TNF-α and IL-6 were used: Bovine TNF-α forward,
5′-CAGGGCTCCAGAAGTTGCTTG-3′ and reverse,
5′-GAACCAGAGGGCTGTTGATGG-3′; Bovine IL-6 forward,
5′-GTGATGACTTCTGCTTTCCCTACC-3′ and reverse,
5′-TCTGCCAGTGTCTCCTTGCTG-3′.
Statistical analysis
Pop Gene software version 3.1 (https://sites.ualberta.ca/~fyeh/popgene.html)
(5) was used to assess if
frequencies of the alleles, diploid data of the single-population
and the significance of differences of single locus [including
allele number, frequency, loci and polymorphism information content
(PIC)] had a P-value <0.05. Data are expressed as the mean ±
standard deviation. Association between single nucleotide
polymorphisms (SNPs) in the CD14 gene and the morbidity of
clinical mastitis was analyzed with crosstabs methods (including
Pearson χ2, Continuity correction and Fisher' exact
test) in SPSS version 13.0 (SPSS, Inc., Chicago, IL, USA).
Results
Gene amplification
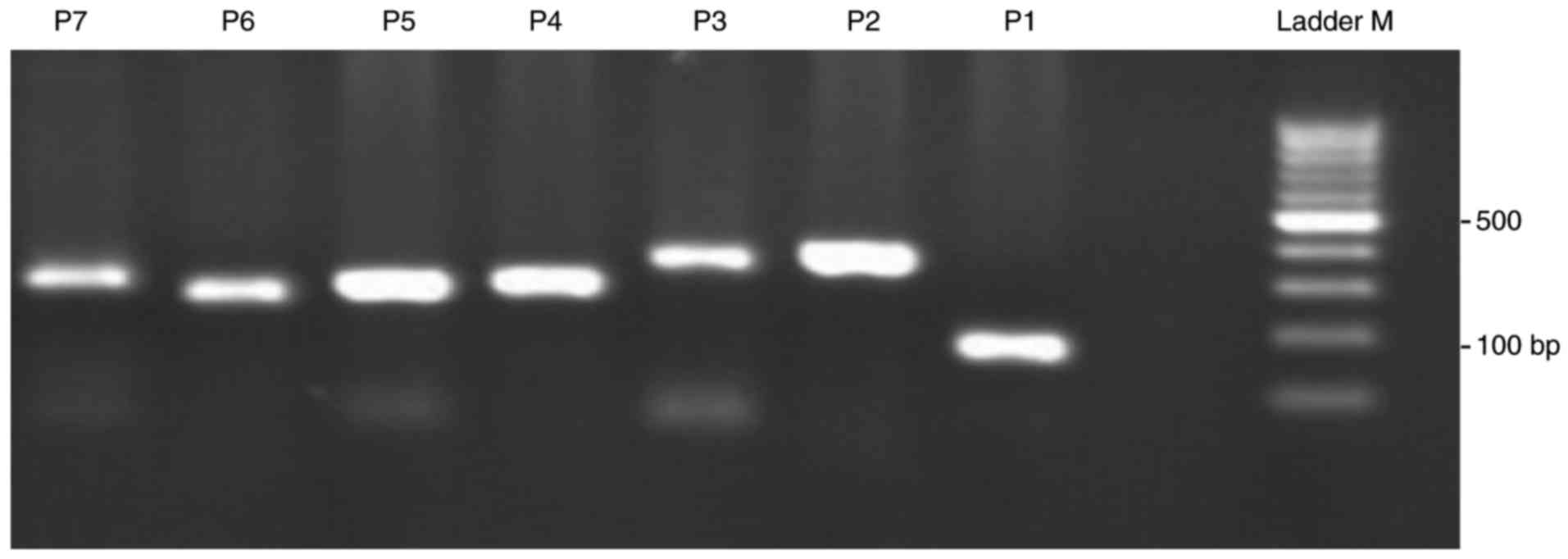
Seven pairs of primers were tested to amplify
CD14 and product sizes were as expected (Table I; Fig.
1). Therefore, CD14 genotypes were analyzed further by
SSCP.
Analysis of SSCP and sequences
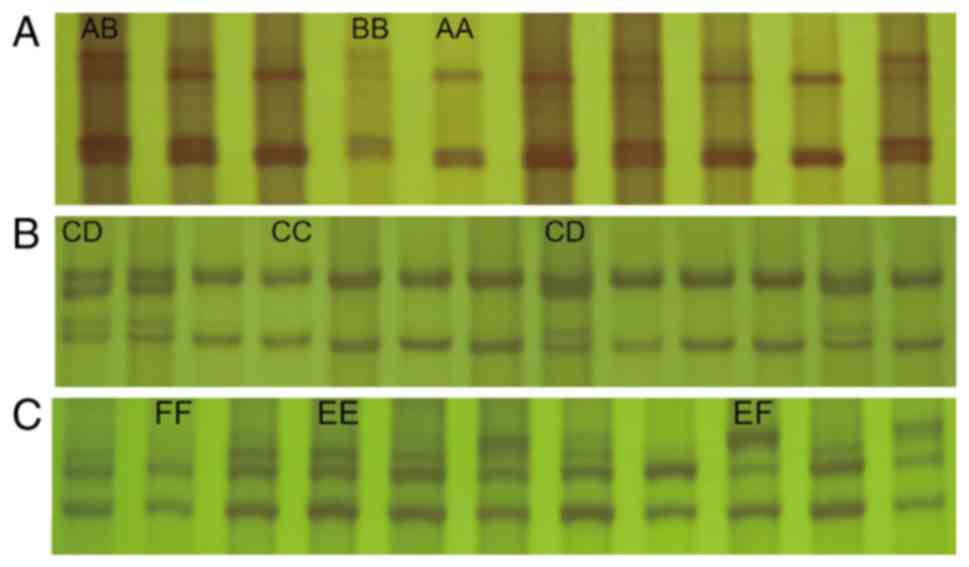
The SSCP analysis revealed the polymorphisms in the
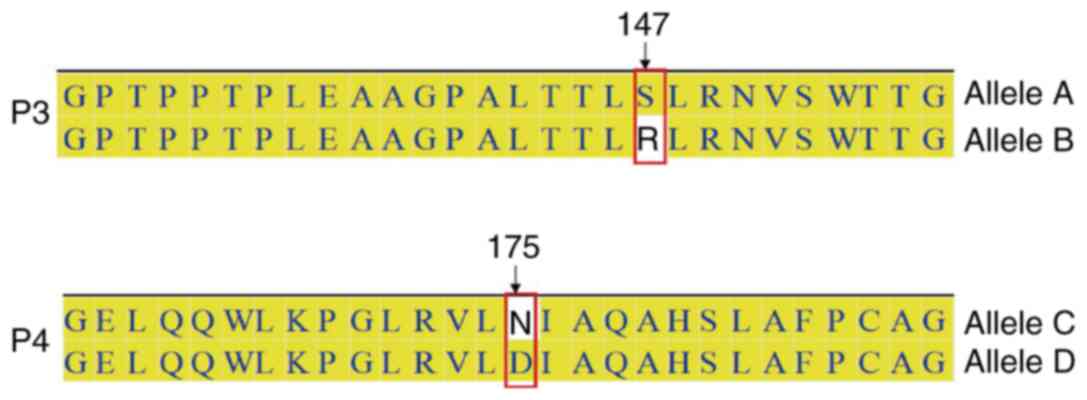
CD14 gene all of which were found in exon 2 (Fig. 2). Three genotypes (AA, AB and BB)
were found in PCR products amplified with primer pair 3 (P3)
including one SNP: g.528 A→C in allele B (Fig. 2A). Two genotypes (CC, CD) were
found in PCR products amplified with primer pair 4 (P4) including
one SNP: g.612 A→G in allele D (Fig.
2B); three genotypes (EE, EF and FF) were found in PCR products
amplified with primer pair 6 (P6) including one SNP: g.1022 A→G in
allele F (Fig. 2C). All genotypes
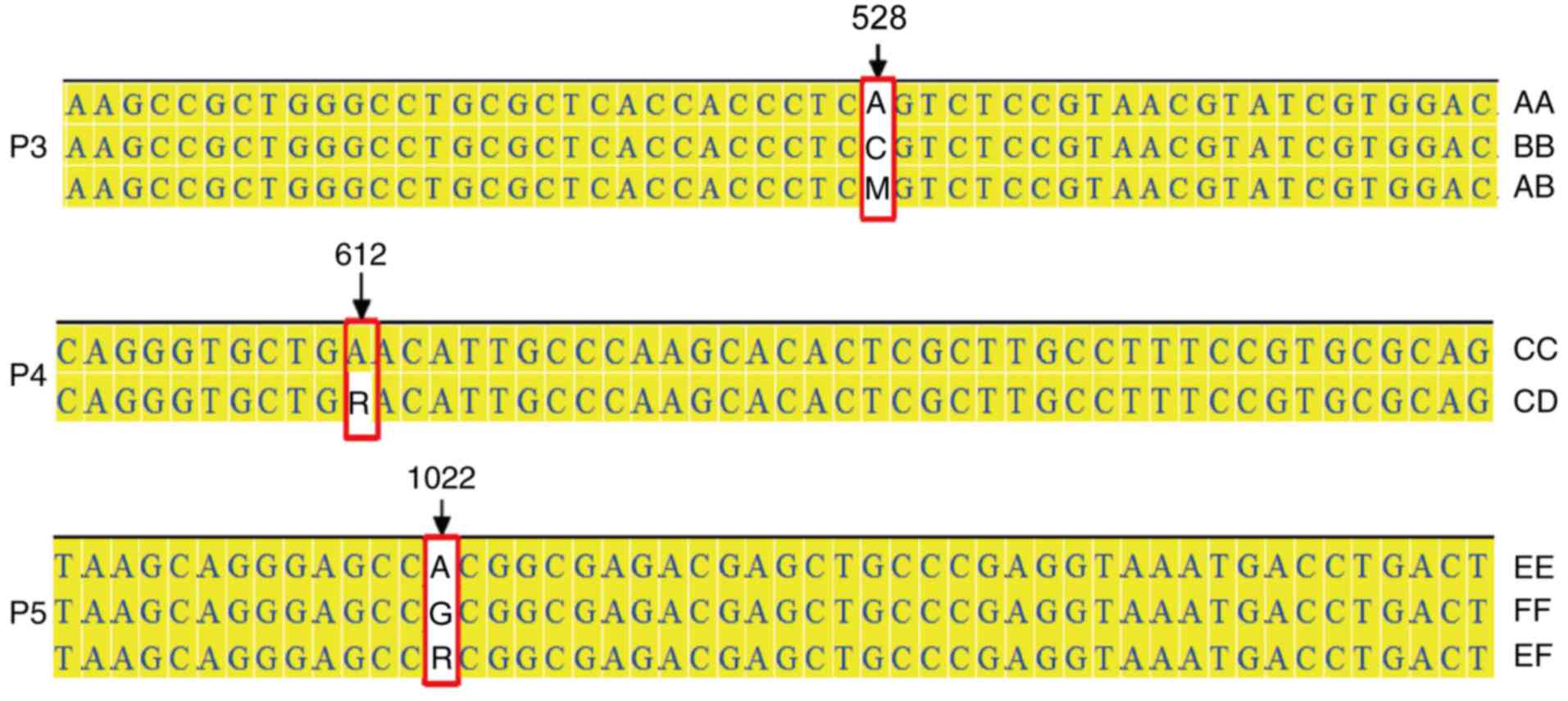
are demonstrated in Fig. 2 and
SNPs in each genotype of exon 2 were compared and illustrated in
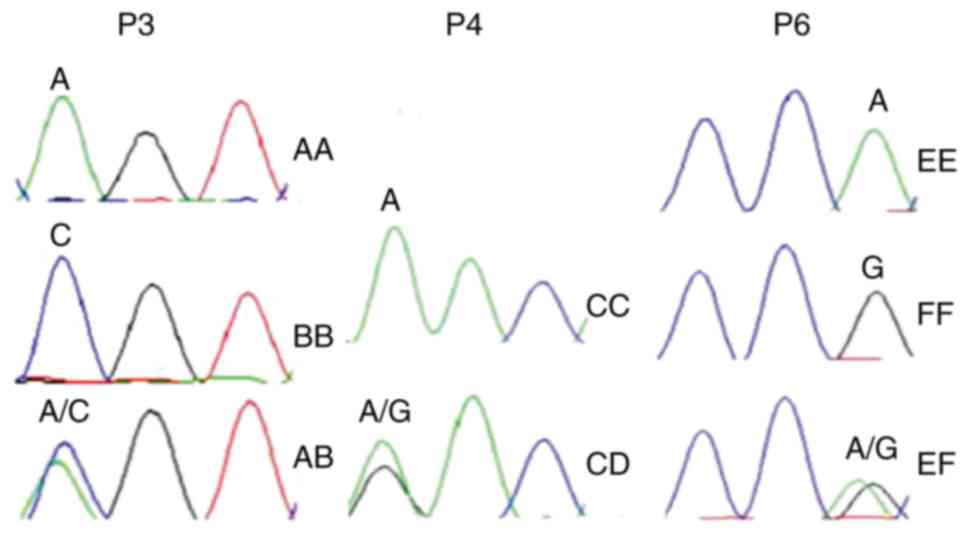
Fig. 3. DNA sequence oscillograms
of different genotypes are illustrated in Fig. 4 and the 3 different SNPs are
presented in Table II. Except for
the SNP in allele F, these SNPs cause the following amino acid
changes: g.528 A→C (147 Ser→Arg) in allele B; g.612 A→G (175
Asn→Asp) in allele D (Fig. 5).
| Table II.Three SNPs identified in CD14
gene. |
Table II.
Three SNPs identified in CD14
gene.
| Primer pair | Allele | SNP |
|---|
| 3 | B | g.528 A→C |
| 4 | D | g.612 A→G |
| 6 | F | g.1022 A→G |
Prediction of CD14 molecular structure
and motif analysis of exon 2
Via the haplotype analysis, only three protein
variants encoded by CD14 were predicted in animals. Protein
information, types, combinations and their associations with the
morbidity of mastitis are listed in Table III, and the different sequences
of these three proteins are presented in Fig. 6.
| Table III.Protein types encoded by CD14,
protein combinations and the association with bovine mastitis. |
Table III.
Protein types encoded by CD14,
protein combinations and the association with bovine mastitis.
| Protein type and
protein combination | Alleles | Frequency of each
protein type or protein group (%) | Number of clinical
individuals | Number of
non.clinical individuals | Morbidity (%) |
|---|
| 1 | AA, CC | 49.2 | 20 | 46 | 30.3a |
| 2 | BB, CC | 19.4 | 12 | 14 | 46.2b |
| 3 | AA, DD | 0 | 0 | 0 | 0 |
| 1 and 2 | A/B, CC | 19.4 | 21 | 5 | 80.8b |
| 1 and 3 | AA, C/D | 12 | 13 | 3 | 81.3b |
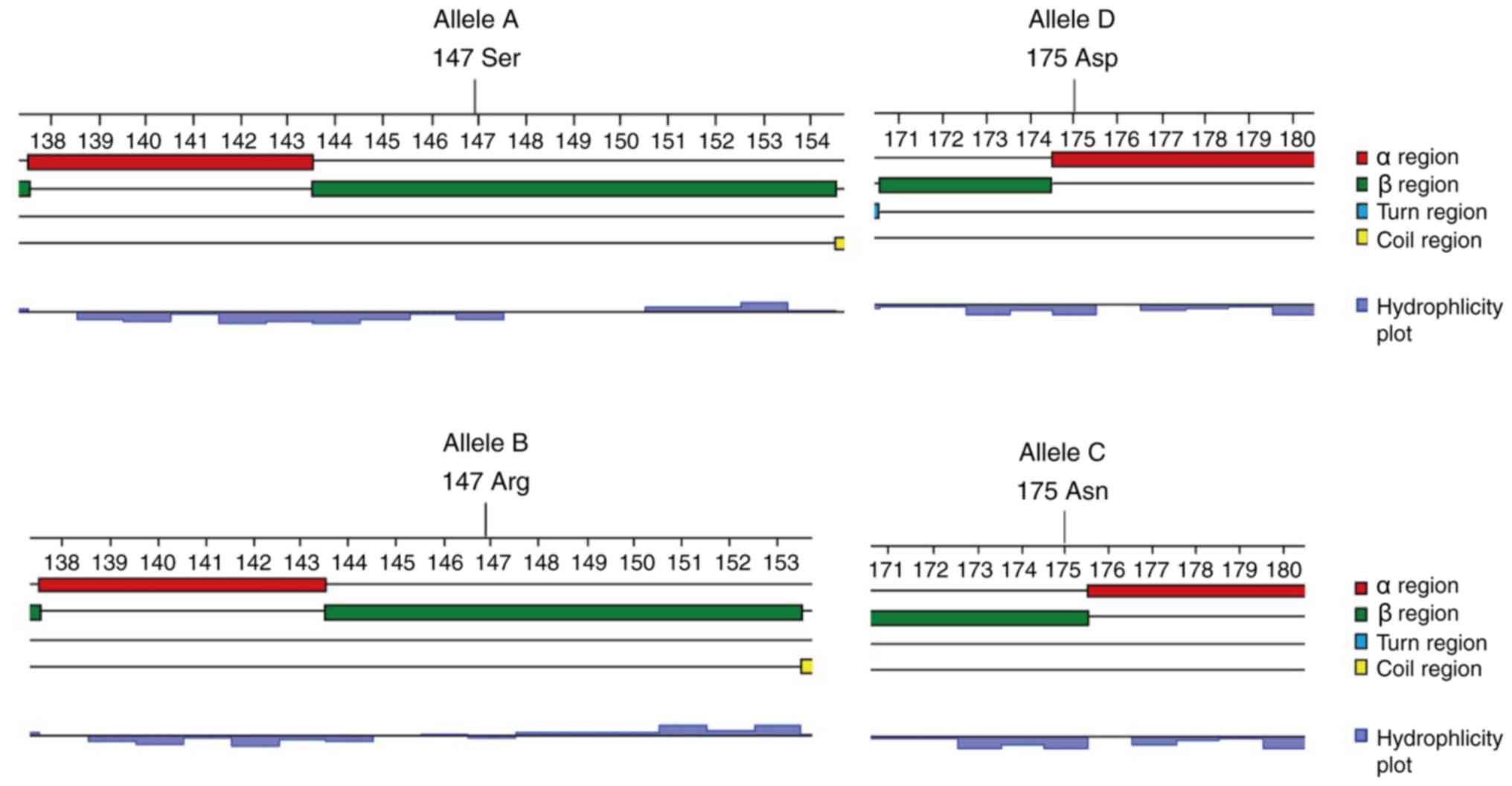
Secondary structure analysis confirmed that bovine
CD14 was a mixed-type protein with extensive hydrophilic and
hydrophobic regions consisting of 36.4% α-helix, 38.7% β-pleated
sheet, 0.7% turns and 4.2% coiled coils domains. The two amino acid
mutations lead to changes of secondary structure: 147 Ser→Arg in
allele B made a β-pleated sheet a little shorter; 175 Asn→Asp in
allele D made β-pleated sheet fall into α-helix (Fig. 7). 147 Ser→Arg in allele B made the
isoelectric point of protein 2 higher than protein 1; 175 Asn→Asp
in allele D caused the isoelectric point of protein 3 lower than
protein 1 (Tables IV–VI).
| Table IV.Isoelectric point of each
protein. |
Table IV.
Isoelectric point of each
protein.
| Predicted
protein | Isoelectric
point |
|---|
| 1 | 5.32 |
| 2 | 5.54 |
| 3 | 5.17 |
| Table VI.Key SNPs in exon 2 of CD14
gene, and their influence on the protein structure and motif
profiles. |
Table VI.
Key SNPs in exon 2 of CD14
gene, and their influence on the protein structure and motif
profiles.
| Allele | SNP | Primary
structure | Secondary
structure | Motif profiles | Location in the 3D
structure |
|---|
| B | g.528 A→C | 147 Ser→Arg | Shorter β-pleated
sheets | Protein kinase C
phosphorylation site moved forward | Concave surface of
CD14 protein |
| D | g.612 A→G | 175 Asn→Asp | β-pleated sheets
into α-helices | No motif
profile | Concave surface of
CD14 protein |
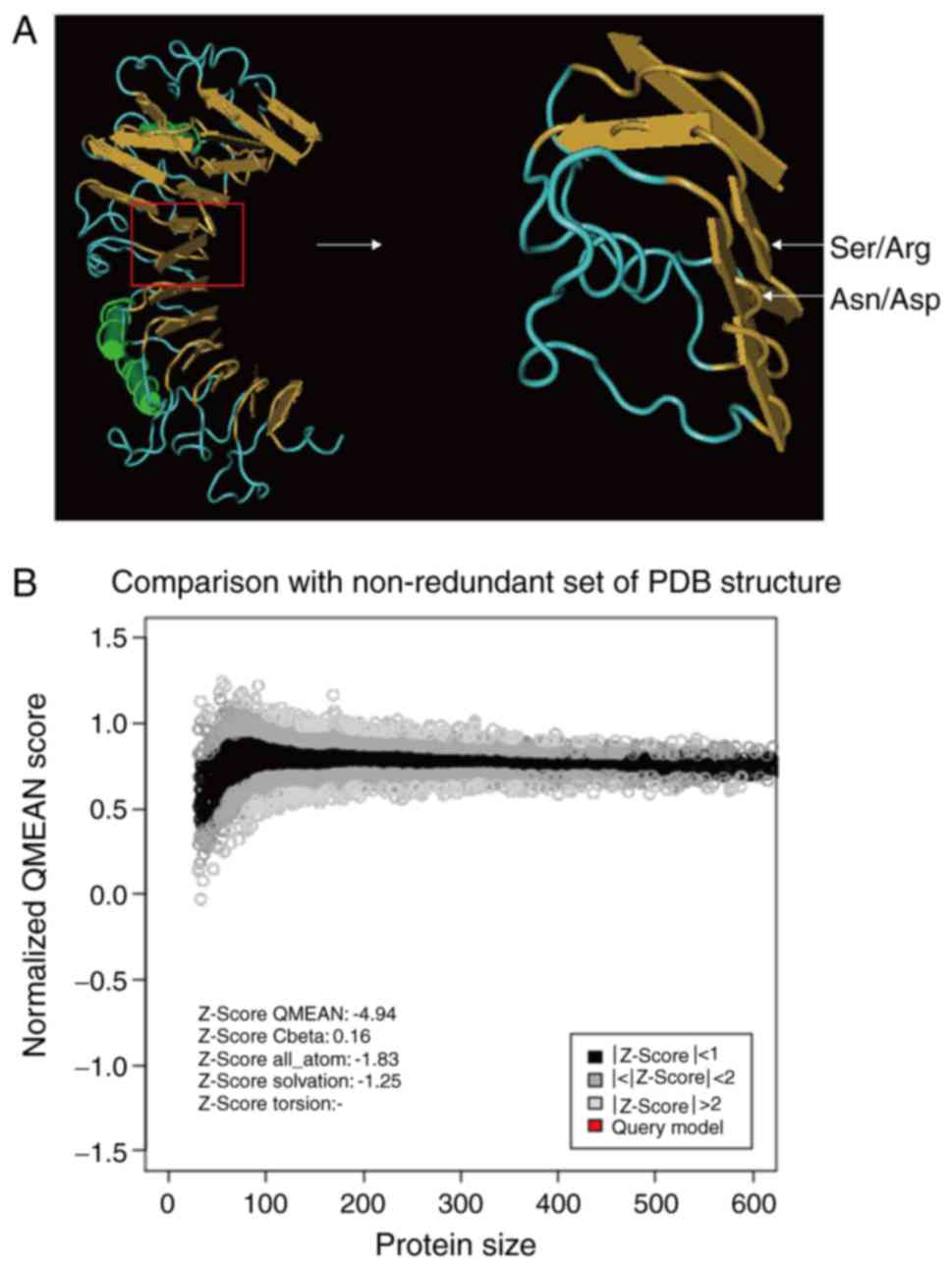
3D structures of these three proteins were also
predicted (Fig. 8A). The QMEAN
score was 0.457 (mean Z score: −4.94; Fig. 8B), which demonstrates the
reliability of the model (29). It
was demonstrated that 147 Ser→Arg induced a protein kinase C (PKC)
phosphorylation site moving forward in protein 3 by using the
PROSITE database Motif assay (Fig.
6).
Genetic analysis of the genotypes and
SNPs
The genetic information of the CD14 gene is listed
in Table VII. In the fragment
amplified by P3, three genotypes were found (AA, AB and BB), and
allele A accounts for 70.90% and in a moderate polymorphism
(0.2<PIC<0.5); Two genotypes were found (CC and CD) amplified
by P4, allele C is prevalent accounting for 94.03% and in low
polymorphism (PIC<0.25); A total of three genotypes were found
(EE, EF and FF) amplified by P6, allele E accounts for 70.15% and
in a moderate polymorphism (0.2<PIC<0.5). Genetic linkage
analysis indicated that alleles B and F were in high linkage
disequilibrium (P<0.01; Table
V). χ2 test revealed that genotyping by P4 and P6
was in Hardy-Weinberg equilibrium (P>0.05) but P3 was not
(P<0.01; Table VII).
| Table VII.Distribution of genotypes and alleles
of CD14 gene. |
Table VII.
Distribution of genotypes and alleles
of CD14 gene.
| Genotype | Number of
cattle | Genotype frequency
(%) | Allele | Allele frequency
(%) | χ2 test
for Hardy Weinberg equilibrium | Polymorphism
information content |
| Primer pair 3 |
|
|
|
| P<0.01 | 0.31 |
| AA | 82 | 61.20 | A | 70.9 |
|
|
| BB | 26 | 19.40 | B | 29.1 |
|
|
| AB | 26 | 19.40 |
|
|
|
| Primer pair 4 |
|
|
|
| P>0.05 | 0.12 |
| CC | 118 | 88.06 | C | 94.03 |
|
|
| CD | 16 | 11.94 | D | 5.97 |
|
|
| Primer pair 6 |
|
|
|
| P>0.05 | 0.33 |
| EE | 64 | 47.76 | E | 70.15 |
|
|
| FF | 10 |
7.46 | F | 29.85 |
|
|
| EF | 60 | 44.78 |
|
|
|
|
| Table V.Linkage disequilibrium between A, B,
E and F alleles. |
Table V.
Linkage disequilibrium between A, B,
E and F alleles.
| P3 allele | P6 allele | Burrows | Correlation | χ2 | P-value |
|---|
| A | E | 0.0810 | 0.03258 | 14.12 | 0.0002 |
| B | F | 0.0810 | 0.03258 | 14.12 | 0.0002 |
Association studies
The association between SNPs and mastitis morbidity
are presented in Table VIII,
and AB has a higher morbidity rate than AA (χ2=12.97,
P<0.01) and BB (χ2=6.72, P<0.01) in the fragments
amplified by P3, while susceptibility to mastitis of CD individuals
is higher than for CC (χ2=7.44, P<0.01) in the
fragments amplified by P4 (P<0.01; Table VIII). However, SNPs in the
fragments amplified by P6 had no difference among different
genotypes (P>0.05). Protein types (including combinations of
them) and their risk of disease are illustrated in Table III, and prevalence rates of
protein 1/protein 2 and protein 1/protein 3 combinations were
higher than that of protein 1 (P<0.01) and protein 2
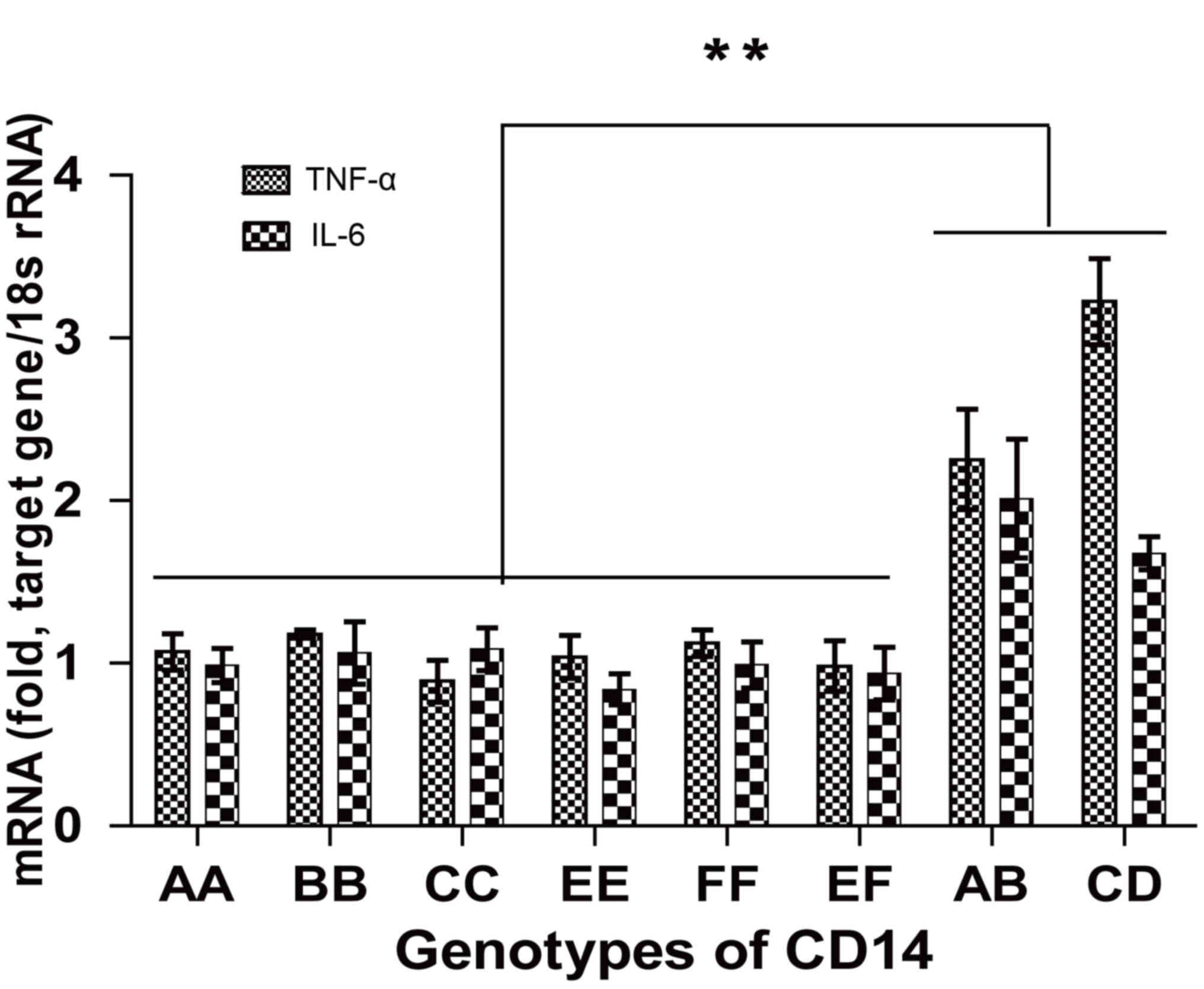
(P<0.05). QRT-qPCR analysis of the clinical samples, revealed
that mRNA expression levels of TNF-α and IL-6 of AB and CD
genotypes were higher than that of the other genotypes (P<0.01;
Fig. 9).
| Table VIII.Statistical analysis of association
between genotypes and susceptibility of clinical mastitis in
Chinese Holstein. |
Table VIII.
Statistical analysis of association
between genotypes and susceptibility of clinical mastitis in
Chinese Holstein.
| Genotype | Number of clinical
individuals | Number of
non-clinical individuals | Morbidity (%) |
|---|
| Primer pair 3 |
|
|
|
| AA | 33 | 49 | 40.2 |
| BB | 12 | 14 | 46.2 |
| AB | 21 | 5 | 80.8a |
| Primer pair 4 |
|
|
|
| CC | 53 | 65 | 44.9 |
| CD | 13 | 3 | 81.3a |
| Primer pair 6 |
|
|
|
| EE | 30 | 34 | 46.9 |
| FF | 4 | 6 | 40 |
| EF | 28 | 32 | 46.7 |
Discussion
The DD genotype may be susceptible to pathogens, and
eventually lead to morbidity in embryonic or adult animals, but
real evidences of lethality remains to be identified. Previous
studies have suggested that SNPs in the immune response-associated
genes contribute to susceptibility to diseases among individuals
(30–32). LPS is the first molecule that GN
bacteria recognize and induce infection (5,9).
Generally, sCD14 in the milk or blood blocks mCD14 function of
macrophages or PMN (8). When GN
bacteria invade the host body, mCD14 on monocytes, macrophages and
PMNs will form a complex with LPS; Notably, sCD14 also binds to
LPS, mediating activation of cells without mCD14 under low
concentration of LPS (33,34). Subsequently, the sCD14-LPS complex
induces the release and translocation of nuclear factor-κB (NF-κB),
which upregulates the expression of pro-inflammatory cytokines
(33). It has been demonstrated
that recombinant sCD14 can relieve inflammation activation in the
mammary gland induced by E. coli (17).
Two SNPs in the coding sequence (CDS) of
CD14 result in primary and secondary structure changes to the
protein, shifting of a PKC phosphorylation site and changes in the
mRNA expression level of TNF-α and IL-6
CD14 serves an essential role in host immunity,
which is the cellular receptor for LPS (8,35).
In the present study, three SNPs located in the coding sequence,
two of which lead to amino acids changes, in which g.528 A→C (147
Ser→Arg) occurs at a functional site, causing a PKC phosphorylation
site to move forward. PKC functions in the LPS-induced immune
responses through activation of mitogen-activated protein
kinase/NF-κB and subsequent transcriptional activation of the
target genes. In addition, previous studies revealed that TNF-α and
IL-6 can be translationally regulated by PKC activation in LPS
response (35,36). Phosphorylation, a
post-translational modification (37) exerts significant effects on many
physiological processes, such as signaling transduction, gene
regulation and cell differentiation. The 528 site SNP found in the
present study may affect PKC phosphorylation. In addition, g.528
A→C (147 Ser→Arg) and g.612 A→G (175Asn→Asp) can change the
isoelectric points of protein 2 and protein 3. As a result, it may
change the secondary structure leading to a functional change of
CD14. AA genotype crossed with BB and the offspring of cross, AB,
perhaps induced mutations of its linked genes, and eventually
elevated TNF-α and IL-6.
SNPs in CD14 protein may change morbidity by
altering CD14 expression on the surface of neutrophils
Researchers indicated that the CDS mutations
occurring in CD14 may change its expression on neutrophils
(38). Neutrophils can protect
animals from GN bacterial infection via building a defense line,
and CD14 confers neutrophils protection against LPS sensitivity
(39), which is indispensable for
innate immunity. LBP strengthens the formation of toll-like
receptor 4-CD14-myeloid differentiation protein 2 complexes,
participate in inflammation signaling pathway transduction,
promoting cytokine release (40–42).
These cytokines mediate the antibiosis and change the abundance of
molecules on surface of neutrophils (33). The present study will shed light on
controlling the inflammatory response and will contribute to the
development of therapeutics for bovine mastitis.
In conclusion, a total of three SNPs were identified
in CDS region of bovine CD14 gene in Chinese Holstein. SNPs
g.528A→C (147 Ser→Arg) and g.612 A→G (175 Asn→Asp) that may be
associated with the expression of CD14 on the surface of
neutrophils. The morbidity ratio of AB and CD was higher than that
of other genotypes, suggesting CD14 genotyping could serve as a
molecular marker for breeding anti-mastitis dairy cows.
Acknowledgements
The present study was supported by the Chinese
National Science Foundation (grant nos. 31372207 and 31572265), a
Start-up grant from Nanjing Agricultural University (grant no.
804090), the ‘Sanxin’ Research Program of Jiangsu Province [grant
no. SXGC(2016)312] and Agricultural Innovation fund of Jiangsu
Province [grant no. CX(17)1005].
References
|
1
|
Baravalle C, Dallard BE, Cadoche MC,
Pereyra EA, Neder VE, Ortega HH and Calvinho LF: Proinflammatory
cytokines and CD14 expression in mammary tissue of cows following
intramammary inoculation of Panax ginseng at drying off. Vet
Immunol Immunopathol. 144:52–60. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Erskine RJ, Tyler JW, Riddell MG Jr and
Wilson RC: Theory, use, and realities of efficacy and food safety
of antimicrobial treatment of acute coliform mastitis. J Am Vet Med
Assoc. 198:980–984. 1991.PubMed/NCBI
|
|
3
|
Ziv G: Treatment of peracute and acute
mastitis. Vet Clin North Am Food Anim Pract. 8:1–15. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ribeiro MG, Motta RG, Paes AC, Allendorf
SD, Salerno T, Siqueira AK, Fernandes MC and Lara GHB: Peracute
bovine mastitis caused by Klebsiella pneumoniae. Arq Bras Med Vet
Zootec. 60:485–488. 2008. View Article : Google Scholar
|
|
5
|
Cheng J, Li J, Zhang W, Cai Y and Wang G:
Mutations in lipopolysaccharide-binding protein (LBP) gene change
the susceptibility to clinical mastitis in Chinese Holstein. Mol
Biol Rep. 39:9601–9612. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang Y, Zarlenga DS, Paape MJ and Dahl GE:
Recombinant bovine soluble CD14 sensitizes the mammary gland to
lipopolysaccharide. Vet Immunol Immunopathol. 86:115–124. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Banks KL and Michaels FH: Stimulation and
killing of bovine mononuclear leukocytes by bacterial
lipopolysaccharide (endotoxin). Am J Vet Res. 46:1568–1572.
1985.PubMed/NCBI
|
|
8
|
Chen YC, Wang SY and King CC: Bacterial
lipopolysaccharide inhibits dengue virus infection of primary human
monocytes/macrophages by blockade of virus entry via a
CD14-dependent mechanism. J Virol. 73:2650–2657. 1999.PubMed/NCBI
|
|
9
|
Landmann R, Ludwig C, Obrist R and Obrecht
JP: Effect of cytokines and lipopolysaccharide on CD14 antigen
expression in human monocytes and macrophages. J Cell Biochem.
47:317–329. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Erridge C, Guerrero E Bennet and Poxton
IR: Structure and function of lipopolysaccharides. Microbes Infect.
4:837–851. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guerra S, Lohman I Carla, LeVan TD, Wright
AL, Martinez FD and Halonen M: The differential effect of genetic
variation on soluble CD14 levels in human plasma and milk. Am J
Reprod Immunol. 52:204–211. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Van Miert A.S.J.P.A.M.: Acute phase
response and non cellulardefence mechanisms. Flemish Vet J.
62:69–72. 1991.
|
|
13
|
Beamer LJ, Carroll SF and Eisenberg D: The
three-dimensional structure of human
bactericidal/permeability-increasing protein: Implications for
understanding protein-lipopolysaccharide interactions. Biochem
Pharmacol. 57:225–229. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Loppnow H, Stelter F, Schonbeck U,
Schluter C, Ernst M, Schutt C and Flad HD: Endotoxin activates
human vascular smooth muscle cells despite lack of expression of
CD14 mRNA or endogenous membrane CD14. Infect Immun. 63:1020–1026.
1995.PubMed/NCBI
|
|
15
|
Pugin J, Ulevitch RJ and Tobias PS:
Activation of endothelial cells by endotoxin: Direct versus
indirect pathways and the role of CD14. Prog Clin Biol Res.
392:369–373. 1995.PubMed/NCBI
|
|
16
|
Qureshi ST, Lariviere L, Leveque G,
Clermont S, Moore KJ, Gros P and Malo D: Endotoxin-tolerant mice
have mutations in Toll-like receptor 4 (TLR-4). J Exp Med.
189:615–625. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee JW, Paape MJ, Elsasser TH and Zhao X:
Recombinant soluble CD14 reduces severity of intramammary infection
by Escherichia coli. Infect Immun. 71:4034–4039. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Burvenich C, Van Merris V, Mehrzad J,
Diez-Fraile A and Duchateau L: Severity of E. colimastitis is
mainly determined by cow factors. Vet Res. 34:521–564. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hartel CH, Finas D, Ahrens P, Kattner E,
Schaible T, Muller D, Segerer H, Albrecht K, Moller J, Diedrich K,
et al: Polymorphisms of genes involved in innate immunity:
Association with preterm delivery. Mol Hum Reprod. 10:911–915.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Donati M, Berglundh T, Hytonen AM,
Hahn-Zoric M, Hanson LA and Padyukov L: Association of the −159
CD14 gene polymorphism and lack of association of the −308 TNFA and
Q551R IL-4RA polymorphisms with severe chronic periodontitis in
Swedish Caucasians. J Clin Periodontol. 32:474–479. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nishimura S, Zaitsu M, Hara M, Yokota G,
Watanabe M, Ueda Y, Imayoshi M, Ishii E, Tasaki H and Hamasaki Y: A
polymorphism in the promoter of the CD14 gene (CD14/-159) is
associated with the development of coronary artery lesions in
patients with Kawasaki disease. J Pediatr. 143:357–362. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rosas-Taraco AG, Revol A, Salinas-Carmona
MC, Rendon A, Caballero-Olin G and Arce-Mendoz AY: CD14 C (−159) T
polymorphism is a risk factor for development of pulmonary
tuberculosis. J Infect Dis. 196:1698–1706. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pal A, Sharma A, Bhattacharya TK,
Chatterjee PN and Chakravarty AK: Molecular characterization and
SNP detection of CD14 gene of crossbred cattle. Mol Biol Int.
2011:5073462011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang P, Lu C, Li J, Xu J, Liu Z, Wang Q,
Wang Z, Huo J, Li H, Teng Y, et al: Mutations in HSP70-2 gene
change the susceptibility to clinical mastitis in Chinese Holstein.
Gene. 559:62–72. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hopp TP: Protein surface analysis. Methods
for identifying antigenic determinants and other interaction sites.
J Immunol Methods. 88:1–18. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hopp TP: Retrospective: 12 years of
antigenic determinant predictions and more. Pept Res. 6:183–190.
1993.PubMed/NCBI
|
|
27
|
Schwede T, Kopp J, Guex N and Peitsch MC:
SWISS-MODEL: An automated protein homology-modeling server. Nucleic
Acids Res. 31:3381–3385. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Guex N and Peitsch MC: SWISS-MODEL and the
Swiss-Pdb Viewer: An environment for comparative protein modelling.
Electrophoresis. 18:2714–2723. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Arnold K, Bordoli L, Kopp J and Schwede T:
The SWISS-MODEL workspace: A web-based environment for protein
structure homology modelling. Bioinformatics. 22:195–201. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Beutler B and Poltorak A: Positional
cloning of Lps, and the general role of toll-like receptors in the
innate immune response. Eur Cytokine Netw. 11:143–152.
2000.PubMed/NCBI
|
|
31
|
Smirnova I, Poltorak A, Chan EK, McBride C
and Beutler B: Phylogenetic variation and polymorphism at the
toll-like receptor 4 locus (TLR-4). Genome Biol. 1:RESEARCH0022000.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Michel O, LeVan TD, Stern D, Dentener M,
Thorn J, Gnat D, Beijer ML, Cochaux P, Holt PG, Martinez FD, et al:
Systemic responsiveness to lipopolysaccharide and polymorphisms in
the toll-like receptor 4 gene in human beings. J Allergy Clin
Immunol. 112:923–929. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Frey EA, Miller DS, Jahr TG, Sundan A,
Bazil V, Espevik T, Finlay BB and Wright SD: Soluble CD14
participates in the response of cells to lipopolysaccharide. J Exp
Med. 176:1665–1671. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pugin J, Schurer-Maly CC, Leturcq D,
Moriarty A, Ulevitch RJ and Tobias PS: Lipopolysaccharide
activation of human endothelial and epithelial cells is mediated by
lipopolysaccharide-binding protein and soluble CD14. Proc Natl Acad
Sci USA. 90:pp. 2744–2748. 1993; View Article : Google Scholar : PubMed/NCBI
|
|
35
|
McKenna TM, Li S and Tao S: PKC mediates
LPS- and phorbolinduced cardiac cell nitric oxide synthase activity
and hypocontractility. Am J Physiol. 269:H1891–H1898.
1995.PubMed/NCBI
|
|
36
|
Kim JK, Lee SM, Suk K and Lee WH: A novel
pathway responsible for lipopolysaccharide-induced translational
regulation of TNF-α and IL-6 expression involves protein kinase C
and fascin. J Immunol. 187:6327–6334. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim JH, Lee J, Oh B, Kimm K and Koh I:
Prediction of phosphorylation sites using SVMs. Bioinformatics.
20:3179–3184. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ibeagha-Awemu EM, Lee JW, Ibeagha AE and
Zhao X: Bovine CD14 gene characterization and relationship between
polymorphisms and surface expression on monocytes and
polymorphonuclear neutrophils. BMC Genet. 9:502008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Paape MJ, Lilius EM, Wiitanen PA, Kontio
MP and Miller RH: Intrammary defense against infections induced by
Escherichia coli cows. Am J Vet Res. 57:477–482. 1996.PubMed/NCBI
|
|
40
|
Cleveland MG, Gorham JD, Murphy TL,
Toumanen E and Murphy KM: Lipoteichoic acid preparations of
gram-positive bacteria induce interleukin-12 through a
CD14-dependent pathway. Infect Immun. 64:1906–1912. 1996.PubMed/NCBI
|
|
41
|
De Schepper S, De Ketelaere A, Bannerman
DD, Paape MJ, Peelman L and Burvenich C: The toll-like receptor-4
(TLR-4) pathway and its possible role in the pathogenesis of
Escherichia coli mastitis in dairy cattle. Vet Res. 39:52008.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Shin HJ, Lee H, Park JD, Hyun HC, Sohn HO,
Lee DW and Kim YS: Kinetics of binding of LPS to recombinant CD14,
TLR4 and MD-2 proteins. Mol Cells. 24:119–124. 2007.PubMed/NCBI
|