Introduction
Cardiovascular disease is the most common cause of
mortality worldwide, with >730,000,000 cases of coronary heart
disease-associated mortality reported by the World Health
Organization in 2016 (1). Clinical
studies have shown that morbidity and mortality rates of
cardiovascular disease in developing countries are increasing and
are estimated to reach 2,330,000,000 by 2030 (2). Atherosclerosis is one of the most
common causes of cardiovascular disease, which is characterized by
subsequent myocardial ischemia, insufficient blood supply to the
coronary arteries and hypoxia (3,4).
Atherosclerosis characterized by lipid deposition is the most
common and important type of arteriosclerosis in the intima of the
involved artery, proliferation of fibrous tissue, calcium
deposition, lesions in the middle layer of the artery and
accumulation of complex carbohydrates (5). Clinically, coronary heart disease is
a fundamental pathological phenotype of atherosclerosis (6,7).
Despite an increasing number of drugs devoted to the treatment of
atherosclerosis, the rates of atherosclerosis-associated morbidity
and mortality remain at high levels (8). Therefore, the prevention and
treatment of cardiovascular disease is a problem requiring urgent
attention in order to improve the quality of life of patients with
cardiovascular disease (9,10).
Fibroblast growth factor-21 (FGF-21) is an atypical
member of the FGF family, and is a multifunctional protein
predominantly secreted by adipose tissue, the pancreas and the
liver (11,12). FGF-21 is predominantly identified
as a momentous controller and regulator of glucose and lipid
metabolism, and of long-term energy balance (13,14).
FGF-21 has been found to be associated with various human diseases
and metabolic syndromes, including ageing, obesity, type 2 diabetes
mellitus and congenital hypothyroidism (15–17).
A previous study showed that FGF-21 resulted in insulin resistance
by inhibiting the activation of nuclear factor (NF)-κB (18). FGF-21 also functions as an
endocrine hormone by inhibiting somatic growth leading to growth
hormone resistance (19). FGF-21
has been reported to be associated with lipid metabolism and the
incidence of cardiovascular disease (20). However, the mechanisms underlying
the association of FGF-21 and cardiovascular disease remain to be
fully elucidated.
In previous years, FGF-21 has been developed as a
promising metabolic regulator and a potential drug for various
human diseases, including ageing, type 2 diabetes and
cardiovascular disease (21,22).
Although the role of FGF-21 in metabolic regulation has been
partially elucidated, the specific signaling pathways have not been
reported or discussed in cardiovascular disease. A previous study
showed that FGF-21 is beneficial for the regulation of inflammatory
factors. Coincidentally, inflammatory aggravation is found in
vessels in the progress of atherosclerosis, which is associated
with blood lipid disorder. FGF-21 can regulate insulin phosphate
metabolism and repair endothelial cells, which may contribute to
recovery in cardiovascular disease. Endothelial dysfunction
accounts for the most characteristic marker in patients with
atherosclerosis, which is caused by oxidative stress. Therefore,
FGF-21 may be associated with atherosclerosis.
The present study investigated the therapeutic
effects and primary mechanism of FGF-21 on a rat model of
atherosclerosis induced by vitamin D3 and a high-fat
diet. The function of FGF-21 on the formation of neointimal cells
and endothelial-dependent relaxation was also investigated, and the
protein expression levels of Forkhead box O (FOXO) in animals in
the atherosclerosis model treated with different doses of FGF-21
were examined. The resulting data indicated that FGF-21
significantly downregulated levels of blood lipids, Rho kinase and
NF-κB, which contributed to atherosclerosis therapy and may be the
mechanisms underlying anti-atherosclerotic effects in the model
rats.
Materials and methods
Ethical approval
The present study was performed in strict accordance
with the recommendations in the Guide for the Care and Use of
Laboratory Animals of Affiliated Hospital of Nanjing University
Medical School (Nanjing, China). All experimental protocols and
animals were performed in accordance with National Institutes of
Health and approved by the Committee on the Ethics of Animal
Experiments Defence Research of Affiliated Hospital of Nanjing
University Medical School (Nanjing, China) (23). All surgery and sacrifice were
performed in a manner to minimize animal suffering.
Experimental animals
A total of 120 female Wistar rats (6–8 weeks old;
190±20 g) were purchased from Slack Experimental Animals Co., Ltd.
(Shanghai, China). All animals were housed under pathogen-free
conditions (license no. SCXK-2008-014). All rats were raised on a
12-h light/dark cycle. The animals had free access to water and the
temperature was 20±2°C with a humidity of 50±5%. All rats were
provided with a high-fat diet and intraperitoneal injections of
vitamin D3 (160,000 U/kg) once a month for a total of five months,
following a 7-day acclimatization period. The rats only received
the high-fat diet in the next 2 months. The rats were randomly
divided into three groups, in which animals received intravenous
treatment with PBS, FGF-21 (6 mg/kg/d) or atorvastatin (4.8
mg/kg/d), respectively, once every 3 days. The rats were starved
and sacrificed on day 30.
Tissue preparation
On day 30, the rats were sacrificed by
intraperitoneal injection of 5% urethane (1,000 mg/kg). The
abdominal aorta, full-length aorta and peripheral blood were
obtained in the experimental rats. The tissues and serum samples
were collected and stored to further analysis. Serum samples were
obtained by centrifugation at 1,500 × g and 4°C for 15 min. The
serum levels of low-density lipoprotein cholesterol, high-density
lipoprotein cholesterol, triglyceride and total cholesterol were
measured according to the methods described in a previous study
(24). The tissues were
homogenized using electrically-driven tissue homogenizer (ABI,
Grand Island, NY, USA) and prepared for reverse
transcription-quantitative polymerase chain reaction (RT-qPCR),
immunofluorescence and western blot analyses.
RT-qPCR analysis
Total RNA was obtained from on endothelial cells of
the experimental rats using an RNAeasy Mini kit (Qiagen, Inc.,
Gaithersburg, MD, USA). The expression levels of FGF-21, chemokine
(C-C motif) ligand (Ccl)2, Ccl5, intercellular adhesion molecule 1
(Icam1) and tumor necrosis factor (TNF)α in the cells were measured
using β-actin as an endogenous control (25) (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). The forward and reverse
primers were synthesized by Invitrogen; Thermo Fisher Scientific,
Inc. PCR amplification was preliminary denaturation at 94°C for 2
min, followed by 45 cycles of 95°C for 30 sec, annealing
temperature reduced to 58°C for 30 sec, and 72°C for 10 min using a
volume of 20 µl containing 50 ng of genomic DNA, 200 µM dNTP, 2.5
units of Taq DNA polymerase and 200 µM primers (Table I). The relative changes in mRNA
expression were calculated using the 2−ΔΔCq method
(25). The results are expressed
as the n-fold change compared with the control.
 | Table I.Sequences of primers used. |
Table I.
Sequences of primers used.
|
| Sequence |
|---|
|
|
|
|---|
| Gene name | Reverse | Forward |
|---|
| Ccl2 |
5′-GATCTCAGTGCAGAGGCTCG-3′ |
5′-TGCTTGTCCAGGTGGTCCAT-3′ |
| Ccl5 |
5′-GTGAGGAACAAGCCAGAG-3′ |
5′-TGACCAGAAGAAGGAATGC-3′ |
| Icam1 |
5′-GGAACCCATTGCCCGAGC-3′ |
5′-GGTGAGGATTGCATTAGGTC-3′ |
| TNFα |
5′-TCCAGACTTCCTTGAGACA-3′ |
5′-GGCGATTACAGACACAACT-3′ |
| β-actin |
5′-GTTGGTCTACCGGGACTCAA-3′ |
5′-CTGAACCCTAAGGCCAACCG-3′ |
Western blot analysis
The endothelial cells in the experimental rats were
homogenized in lysate buffer containing protease-inhibitor and were
centrifuged at 6,000 × g at 4°C for 10 min. The supernatant of the
mixture was used for analysis of the proteins of interest. For the
detection of purpose protein, transmembrane proteins were extracted
using a Transmembrane Protein Extraction kit (Qiagen, Inc.)
according to the manufacturer's protocol. SDS assays were performed
as previously described (26). For
western blot analysis, primary antibodies: p65 (ab16502; 1:1,000;
Abcam, Cambridge, UK), NF-κB (ab97726; 1:1,000; Abcam) were added
following blocking in 5% skimmed milk for 1 h at 37°C. This was
followed by incubation with horseradish peroxidase-conjugated
anti-rabbit IgG (1721019; 1:5,000; Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) for 24 h at 4°C. The results were visualized by
using a chemiluminescence detection system (1705060; Bio-Rad
Laboratories, Inc.).
Histological examination
Tissues were fixed for 48 h, dehydrated through a
graded series of ethanol, embedded in paraffin wax, and cut into 4
µm tissue sections. The sections were stained with hematoxylin and
eosin (H&E) and observed under a light microscope (Olympus
BX51; Olympus Corporation, Tokyo, Japan).
Immunofluorescence
The endothelial cells in the experimental rats were
incubated with primary antibodies, followed by appropriate
secondary antibodies: pro-inflammatory M1 (ab110273; 1:2,000;
Abcam) and anti-inflammatory M2 (ab110274; 1:2,000; Abcam) were
added as targets for primary antibodies for the immobilized cells
for 12 h at 4°C. The horseradish peroxidase-conjugated anti-rabbit
IgG (1721019; 1:5,000; Bio-Rad Laboratories, Inc.) was incubated
with the endothelial cells following washing with PBS for 2 h at
37°C. In addition, the localization and levels of NF-κB were
analyzed using fluorescein isothiocyanate secondary Ab and nuclear
counterstaining (DAPI) using a fluorescence microscope (Leica DMi8;
Leica Microsystems GmbH, Wetzlar, Germany) (27).
Oxidative stress
Intracellular superoxide in arterial tissues was
analyzed using fluorescence microscopy combined with expressed as
number of DHE-positive cells (DAPI staining) and
superoxide-sensitive fluorescent dye dihydroethidium (2 mmol/l,
DHE; Invitrogen; Thermo Fisher Scientific, Inc.). The activity of
NADPH-dependent oxidase in the endothelial cell homogenates was
measured using chemiluminescence by NADPH (100 mmol/l) and
lucigenin (5 mmol/l).
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical tests for data analysis included Fisher's exact test,
log-rank test, χ2 test, and Student's two-tailed t-test.
Multivariate statistical analysis was performed using a Cox
regression model. Statistical analyses were performed using SPSS
version 19.0 (IBM SPSS, Armonk, NY, USA). P<0.05 was considered
to indicate a statistically significant difference.
Results
Differences in the expression of
FGF-21, blood glucose, blood lipids and body weight in rats with
atherosclerosis
In order to examine the role of FGF-21 on
atherosclerosis, the present study first investigated the
expression level of FGF-21 in the peripheral blood of
atherosclerosis rats, with healthy rats as a control. The results,
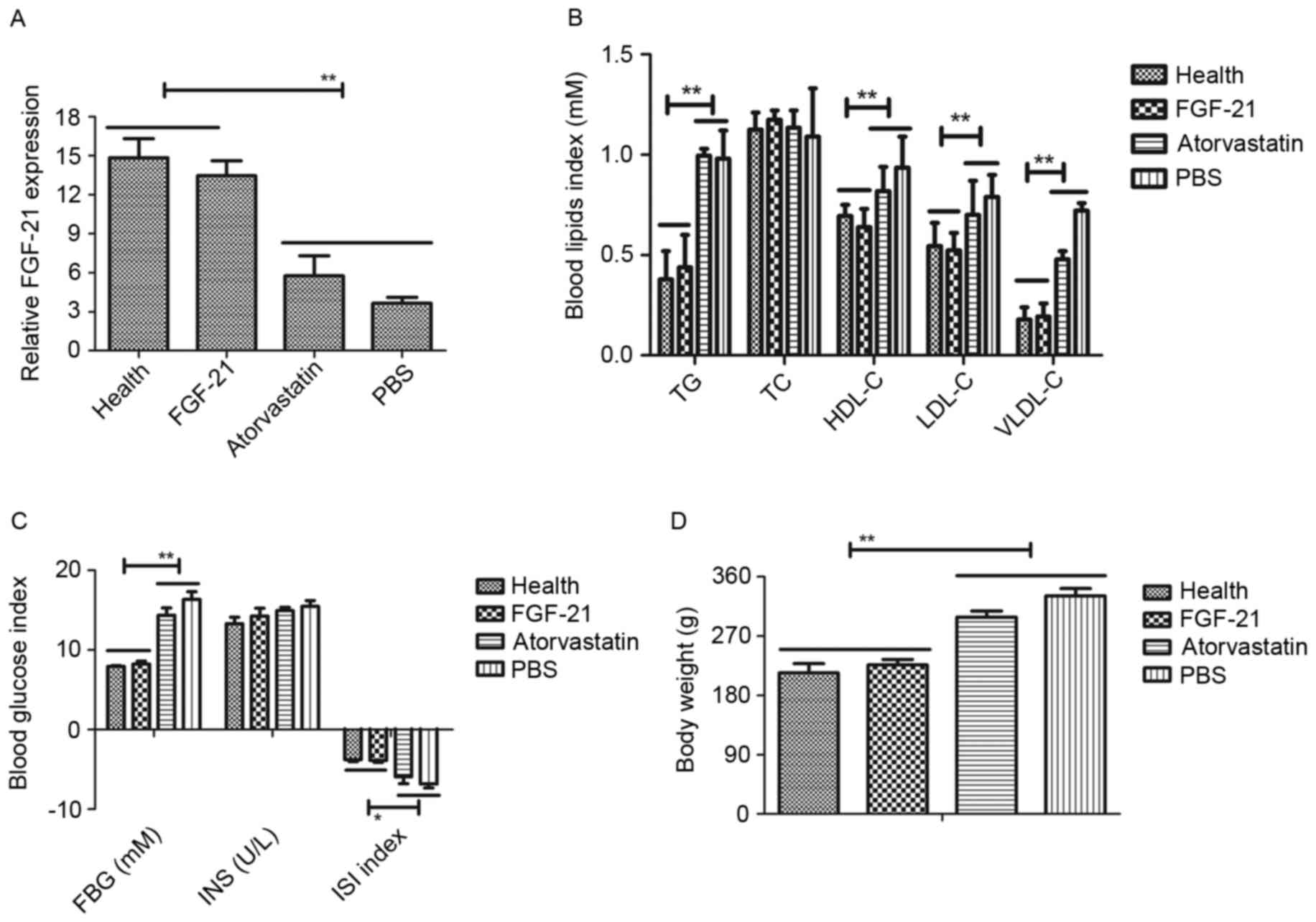
as presented in Fig. 1A, showed
that the expression levels of FGF-21 were downregulated in the rats
with atherosclerosis, compared with the healthy rats. However,
FGF-21 therapy improved recovery to normal levels of FGF-21. The
present study also examined the blood glucose and blood lipids
levels to determine the therapeutic effects of FGF-21 on
atherosclerosis. As presented in Fig.
1B and C, blood glucose and blood lipids were increased in rats
with atherosclerosis, compared with the rats administered with PBS
or atorvastatin. The therapeutic effects of FGF-21 on body weight
on the rats with atherosclerosis were also examined. The data
indicated that FGF-21 inhibited the increase in body weight,
compared with PBS or atorvastatin (Fig. 1D). Collectively, these data
suggested that FGF-21 was downregulated in atherosclerosis, and
that the restoration of FGF-21 had beneficial effects on blood
glucose, blood lipids and body weight in the rats with
atherosclerosis.
Effects of FGF-21 treatment on
endothelial cell ultrastructure, endothelial-dependent relaxation
and balloon-induced neointimal formation
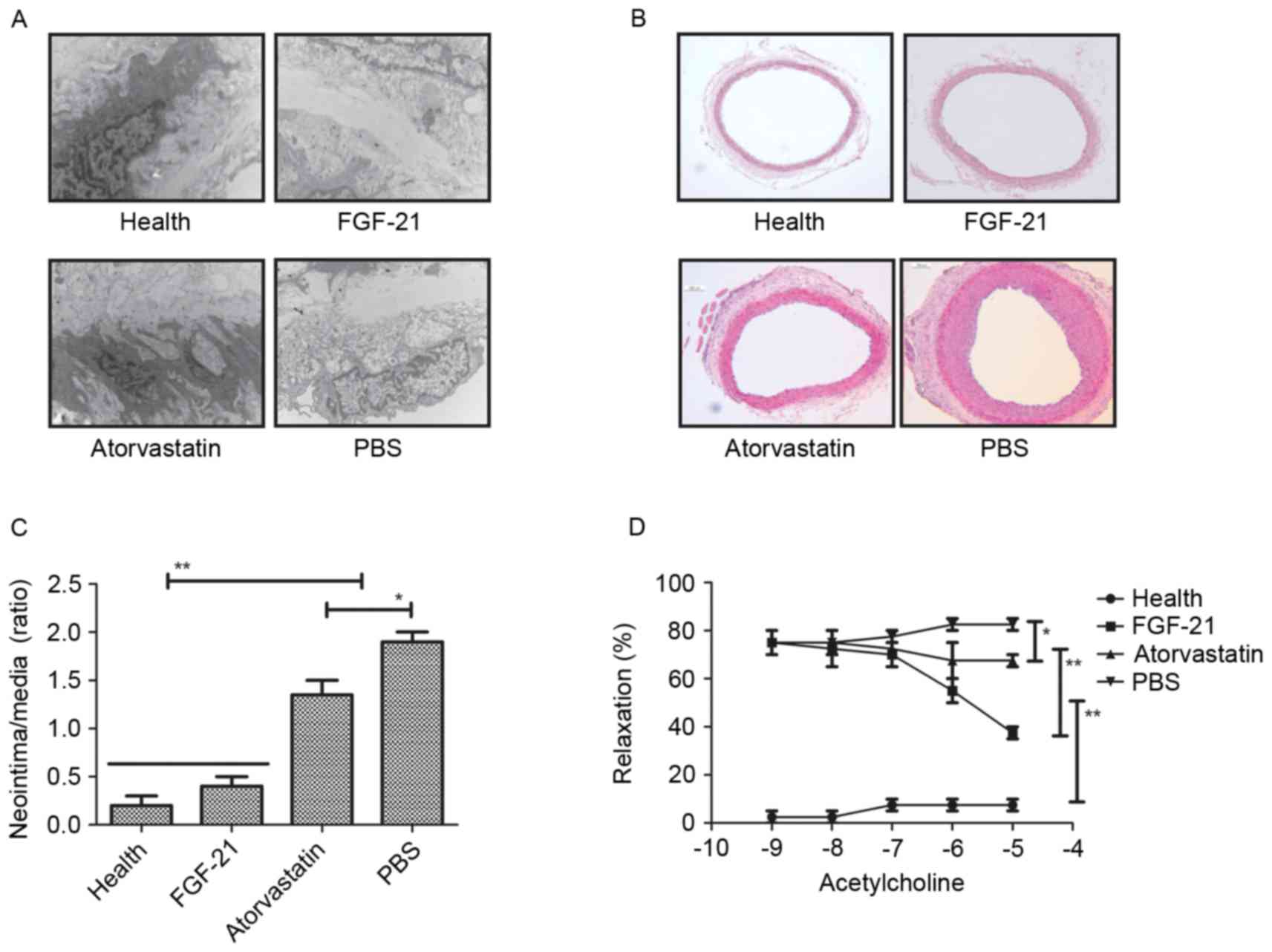
To analyze the effect of FGF-21 on the endothelial
cells in the experimental rat, H&E staining was used to observe
alteration in the endothelial cells. It was found that FGF-21
treatment markedly altered the ultrastructure of the endothelial
cells in the aorta in the rats with atherosclerosis, compared with
the rats administered with PBS or atorvastatin (Fig. 2A). In addition, the formation of
neointima was analyzed in the carotid arteries of rats with
atherosclerosis. The results revealed that FGF-21 significantly
improved neointimal formation, and that the neointimal formation
was higher, compared with that in either the PBS or
atorvastatin-treated groups. (Fig. 2B
and C). The present study also examined the effects of FGF-21
on endothelial-dependent relaxation in the experimental rats to
confirm the improvement of vessel function. As shown in Fig. 2D, the results revealed that
endothelial-dependent relaxation was increased in the injured
carotid arteries of the rats with atherosclerosis. Taken together,
the therapeutic effects of FGF-21 provided an advantage in the
treatment of rats with atherosclerosis.
Effects of FGF-21 treatment on
histopathological changes in atherosclerosis in rats
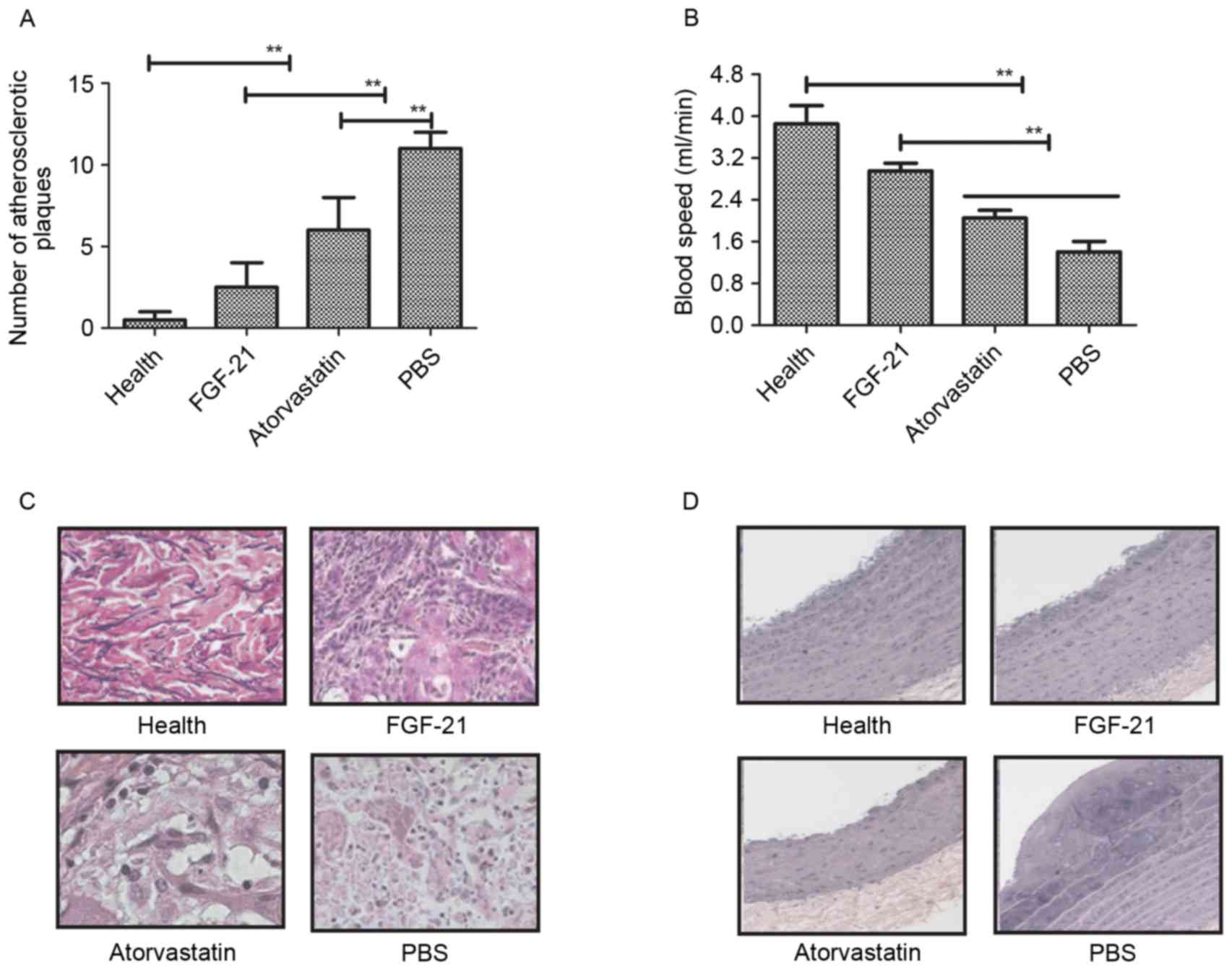
In order to identify the beneficial effects of
FGF-21 on blood vessels, histopathologal changes were detected
using H&E and immunohistochemical staining in the rat model of
atherosclerosis. The data showed that the unit area of
atherosclerotic plaques was significantly lower, compared with the
model and atorvastatin groups (Fig.
3A). The blood velocity increased following FGF-21 treatment
(Fig. 3B) and, as shown in
Fig. 3C, adipocytes were decreased
following FGF-21 treatment. There were marked pathological changes
to the normal functioning in the aortas of the rats with
atherosclerosis following FGF-21 treatment, compared with PBS and
atorvastatin (Fig. 3D). Taken
together, these findings suggested that FGF-21 treatment led to a
visible decrease in pathological changes, including the vessel
walls and atherosclerotic plaques, in rats with
atherosclerosis.
Effects of FGF-21 treatment on
atherosclerosis via the NF-κB signaling pathway
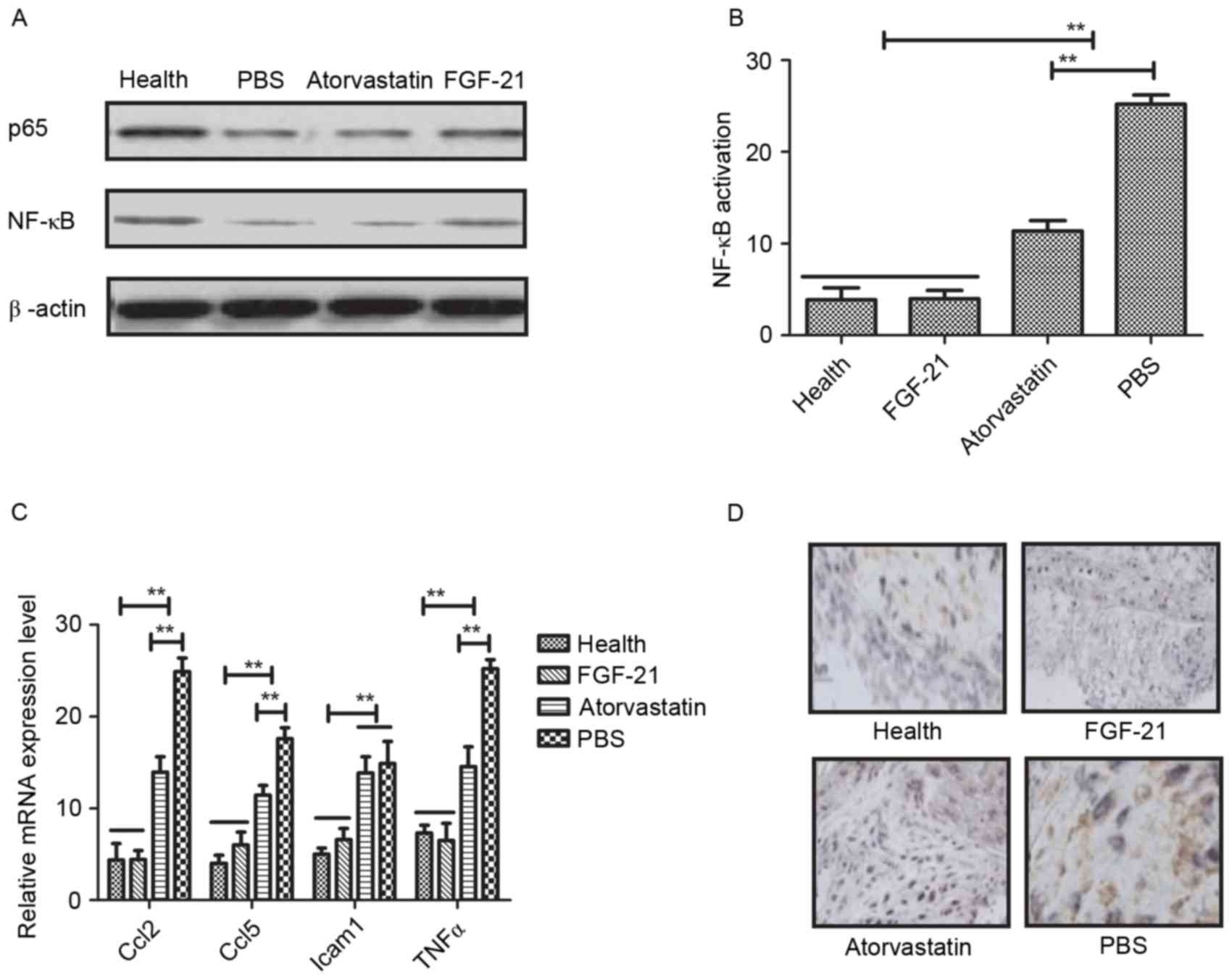
NF-κB is a crucial regulator in inflammatory
responses, which controls the expression of several genes involved
in atherogenesis (28). In the
present study, the associations between FGF-21 and the NF-κB
signaling pathway were analyzed in rats with atherosclerosis. It
was observed that FGF-21 inhibited the nuclear import of activated
NF-κB (p65) in the vascular endothelial cells of the experimental
rats (Fig. 4A). The data indicated
that FGF-21 inhibited the activation of NF-κB in vascular cells
isolated from rats with atherosclerosis (Fig. 4B). In addition, it was found that
FGF-21 treatment decreased the expression levels of
anti-inflammatory genes involved in the NF-κB signaling pathway,
including Ccl2, Ccl5, Icam1 and TNFα, determined using RT-qPCR
analysis (Fig. 4C). The potential
benefits of FGF-21 were also examined, which revealed marked NF-κB
inactivation in the FGF-21-treated rats (Fig. 4D). Taken together, it was concluded
that FGF-21 improved atherosclerosis via the NF-κB signaling
pathway.
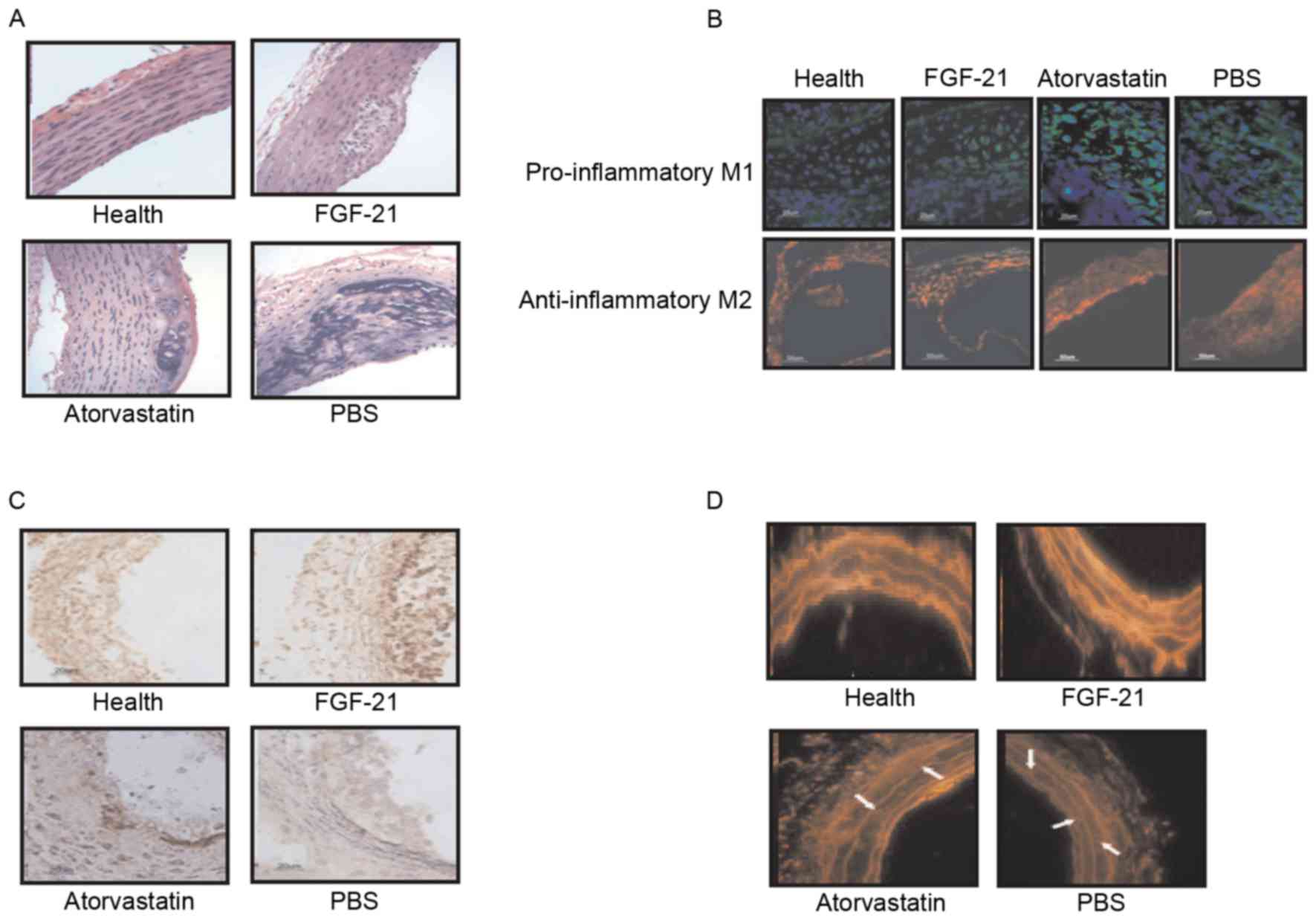
FGF-21 treatment decreases oxidative
stress and atherosclerotic plaques in rats with
atherosclerosis
Further confirming the effects of FGF-21 treatment
on rats with atherosclerosis, the data obtained showed that lesion
size was reduced by FGF-21 treatment, in addition to a significant
decrease in the number of macrophages within the atherosclerotic
plaques (Fig. 5A). Regression
analysis revealed a correlation of macrophage content with lesion
area (r=0.724; P=0.0042) and NF-κB staining (r=0.746; P=0.0031) in
the experimental model, indicating a higher inflammatory component
in larger lesions. It was also observed that pro-inflammatory M1
and anti-inflammatory M2 were increased in the plaques of the
FGF-21-treated rats (Fig. 5B). The
results of immunofluorescence demonstrated a higher relative
collagen and vascular smooth muscle cell (VSMC) content in the
aortic lesions of the FGF-21-treated rats, compared with the PBS
and atorvastatin groups (Fig. 5C).
Furthermore, superoxide generation in the aortic root region of the
experimental animals was analyzed using DHE staining. As
demonstrated in Fig. 5D, FGF-21
treatment reduced the number of DHE-labeled nuclei in the plaques,
suggesting that FGF-21 treatment inhibited oxidative stress through
the NF-κB signaling pathway. Taken together, these data suggested
that FGF-21 mediated inflammation and oxidative stress through the
NF-κB signaling pathway in rats with atherosclerosis, which may be
a potential drug candidate for the treatment of
atherosclerosis.
Discussion
Atherosclerosis is one of the leading causes of
cardiovascular disease-associated mortality worldwide, and is
characterized as a chronic and multifactorial disease occurring in
the arterial wall (29,30). A previous study indicated that
atherosclerotic plaques are the most important clinical features,
characterized by leukocyte infiltration and lipid accumulation
(6). The continuous inflammation
in the artery leads to atherosclerotic plaques and thrombus
formation, which further exacerbates the disease and the difficulty
of treatment (31). Leukocyte
infiltration and lipid accumulation have been reported to
contribute to different process in plaque formation in the
development of atherosclerosis through a serious inflammatory
responses and metabolic disorders (32). Therefore, controlling leukocyte
infiltration and lipid accumulation may be beneficial for recovery
in patients with atherosclerosis. In the present study, data
indicated that lower levels of leukocyte infiltration and lipid
accumulation were observed in FGF-21-treated rats with
atherosclerosis, resulting in improvement through a decrease in the
number of atherosclerotic plaques.
FGF-21 has been demonstrated to be associated with
glucose metabolism, lipid metabolism, hyperglycemia and
thermogenesis, which are performed in a hormone-like manner
(33). The upstream and downstream
signaling pathways of FGF-21 remain to be fully elucidated and are
the focus of numerous investigations. In the present study, the
therapeutic effects of FGF-21 on atherosclerosis were investigated
in a rat model of atherosclerosis. Previously, the association
between FGF-21 concentration and restoration has been reported to
be associated with insulin sensitivity and metabolic syndrome, and
that FGF-21 injection presents a potential multifunctional
regulator in improving insulin resistance and obesity-associated
metabolic disorders (34). A
number of reports on FGF-21 have focused on its efficacy in
diagnosing and treating type 2 diabetes (34,35).
Fewer reports have described the association between FGF-21 and
atherosclerosis in preclinical and clinical studies. It is known
that the expression of FGF-21 is significantly upregulated when
cells are in a state of fasting/starvation, and regulates glucose
and lipid metabolism (36). The
hormone-like FGF-21 is involved in the regulation of diverse
metabolic pathways, including the NF-κB signaling pathway. In the
present study, FGF-21 was shown to regulate leukocyte infiltration
and lipid accumulation via the NF-κB signaling pathway in rats with
atherosclerosis.
The interaction between NF-κB and atherosclerosis in
vascular inflammation and atherosclerosis has been investigated and
discussed in previous studies (37,38).
The activation of NF-κB has been detected in human atherosclerotic
plaques, and the modulation of NF-κB inflammatory activity has been
shown to limit disease progression in mice (39). NF-κB is a major regulator of
inflammation, which controls the expression of several functional
genes involved in atherogenesis (40). In addition, several cellular
assembly and molecular inflammatory responses are involved in the
progress of atherosclerosis, including the release of
pro-inflammatory and inflammatory cytokines, dysfunction of VSMCs,
and activation of leukocyte infiltration and migration (41). The NF-κB signaling pathway is also
a crucial factor regulating the expression of genes in different
processes of atherosclerotic formation, including lipid
modification, adhesion of leukocytes, monocyte differentiation and
VSMC proliferation (42). NF-κB is
also vital in the initiation and development of the atherosclerotic
process through mediating pro-inflammatory transcription factors in
endothelial cells in the artery (43). The aberrant activation of NF-κB is
observed in the majority of human cardiovascular diseases (44,45).
Evidence also suggests that poor survival rates and outcomes of the
atherosclerotic process are associated with aberrant activation of
the NF-κB signaling pathway (46,47).
In conclusion, the findings of the present study
suggested that FGF-21 improved and maintained the morphology of the
vascular endothelium in rats with atherosclerosis by activating the
NF-κB signaling pathway. Data showed that FGF-21 treatment
downregulated the levels of inflammatory factors and inhibited
oxidative stress. It was also demonstrated that FGF-21 inhibited
the nuclear importing of activated NF-κB (p65) in vascular
endothelial cells and downregulated the levels of physiological
NF-κB to protect the vascular endothelium in the experimental rats,
which may offer potential as an alternative for the treatment of
atherosclerosis.
References
|
1
|
Tarugi P, Averna M, Di Leo E, Cefalù AB,
Noto D, Magnolo L, Cattin L, Bertolini S and Calandra S:
Corrigendum to ‘Molecular diagnosis of hypobetalipoproteinemia: An
ENID review’ [Atherosclerosis 195 (2) (2007) 19–27].
Atherosclerosis. 253:e12016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shah P, Bajaj S, Virk H, Bikkina M and
Shamoon F: Rapid progression of coronary atherosclerosis: A review.
Thrombosis. 2015:6349832015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Boesen ME, Singh D, Menon BK and Frayne R:
A systematic literature review of the effect of carotid
atherosclerosis on local vessel stiffness and elasticity.
Atherosclerosis. 243:211–222. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kousios A, Kouis P and Panayiotou AG:
Matrix metalloproteinases and subclinical atherosclerosis in
chronic kidney disease: A systematic review. Int J Nephrol.
2016:94980132016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ayyappan Plakkal J, Paul A and Goo YH:
Lipid droplet-associated proteins in atherosclerosis (Review). Mol
Med Rep. 13:4527–4534. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gholami S, Salavati A, Houshmand S, Werner
TJ and Alavi A: Assessment of atherosclerosis in large vessel
walls: A comprehensive review of FDG-PET/CT image acquisition
protocols and methods for uptake quantification. J Nucl Cardiol.
22:468–479. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Crickx E, Saussine A, Vignon-Pennamen MD,
Cordoliani F, Mouly F, Bagot M and Rybojad M: Diffuse dermal
angiomatosis associated with severe atherosclerosis: Two cases and
review of the literature. Clin Exp Dermatol. 40:521–524. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Merashli M, Ster IC and Ames PR:
Subclinical atherosclerosis in Behcet's disease: A systematic
review and meta-analysis. Semin Arthritis Rheum. 45:502–510. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shah NR and Mahmoudi M: The role of DNA
damage and repair in atherosclerosis: A review. J Mol Cell Cardiol.
86:147–157. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Olubamwo OO, Onyeka IN, Miettola J,
Kauhanen J and Tuomainen TP: Hepatitis C as a risk factor for
carotid atherosclerosis-a systematic review. Clin Physiol Funct
Imaging. 36:249–260. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Eto K: FGF-21, a newcomer in the field of
hypertension research. J Hum Hypertens. 27:343–344. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Reinehr T, Woelfle J, Wunsch R and Roth
CL: Fibroblast growth factor 21 (FGF-21) and its relation to
obesity, metabolic syndrome, and nonalcoholic fatty liver in
children: A longitudinal analysis. J Clin Endocrinol Metab.
97:2143–2150. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Suomalainen A, Elo JM, Pietiläinen KH,
Hakonen AH, Sevastianova K, Korpela M, Isohanni P, Marjavaara SK,
Tyni T, Kiuru-Enari S, et al: FGF-21 as a biomarker for
muscle-manifesting mitochondrial respiratory chain deficiencies: A
diagnostic study. Lancet Neurol. 10:806–818. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lin Z, Wu Z, Yin X, Liu Y, Yan X, Lin S,
Xiao J, Wang X, Feng W and Li X: Serum levels of FGF-21 are
increased in coronary heart disease patients and are independently
associated with adverse lipid profile. PLoS One. 5:e155342010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ren G, Yin J, Wang W, Li L and Li D:
Fibroblast growth factor (FGF)-21 signals through both FGF
receptor-1 and 2. Sci China Life Sci. 53:1000–1008. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kharitonenkov A, Dunbar JD, Bina HA,
Bright S, Moyers JS, Zhang C, Ding L, Micanovic R, Mehrbod SF,
Knierman MD, et al: FGF-21/FGF-21 receptor interaction and
activation is determined by betaKlotho. J Cell Physiol. 215:1–7.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kharitonenkov A, Shiyanova TL, Koester A,
Ford AM, Micanovic R, Galbreath EJ, Sandusky GE, Hammond LJ, Moyers
JS, Owens RA, et al: FGF-21 as a novel metabolic regulator. J Clin
Invest. 115:1627–1635. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Salehi MH, Kamalidehghan B, Houshmand M,
Aryani O, Sadeghizadeh M and Mossalaeie MM: Association of
fibroblast growth factor (FGF-21) as a biomarker with primary
mitochondrial disorders, but not with secondary mitochondrial
disorders (Friedreich Ataxia). Mol Biol Rep. 40:6495–6499. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gahete MD, Córdoba-Chacón J, Luque RM and
Kineman RD: The rise in growth hormone during starvation does not
serve to maintain glucose levels or lean mass but is required for
appropriate adipose tissue response in female mice. Endocrinology.
154:263–269. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu D, Sun CY, Sun GP, Ren GP, Ye XL, Zhu
SL, Wang WF, Xu PF, Li SJ, Wu Q, et al: The synergistic effect of
FGF-21 and insulin on regulating glucose metabolism and its
mechanism. Yao Xue Xue Bao. 49:977–984. 2014.(In Chinese).
PubMed/NCBI
|
|
21
|
Matuszek B, Lenart-Lipińska M, Duma D,
Solski J and Nowakowski A: Evaluation of concentrations of FGF-21-a
new adipocytokine in type 2 diabetes. Endokrynol Pol. 61:50–54.
2010.PubMed/NCBI
|
|
22
|
Yu Y, Bai F, Wang W, Liu Y, Yuan Q, Qu S,
Zhang T, Tian G, Li S, Li D and Ren G: Fibroblast growth factor 21
protects mouse brain against D-galactose induced aging via
suppression of oxidative stress response and advanced glycation end
products formation. Pharmacol Biochem Behav. 133:122–131. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
In: Guidance for the Description of Animal
Research in Scientific Publications. Washington, DC: 2011
|
|
24
|
Sartang Mohammadi M, Mazloomi SM, Tanideh
N and Zadeh Rezaian A: The effects of probiotic soymilk fortified
with omega-3 on blood glucose, lipid profile, haematological and
oxidative stress and inflammatory parameters in streptozotocin
nicotinamide-induced diabetic rats. J Diabetes Res.
2015:6963722015.PubMed/NCBI
|
|
25
|
Xiao S, Wang J and Xiao N: MicroRNAs as
noninvasive biomarkers in bladder cancer detection: A diagnostic
meta-analysis based on qRT-PCR data. Int J Biol Markers.
31:e276–e285. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wai-Hoe L, Wing-Seng L, Ismail Z and
Lay-Harn G: SDS-PAGE-based quantitative assay for screening of
kidney stone disease. Biol Proced Online. 11:145–160. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yahav G, Hirshberg A, Salomon O, Amariglio
N, Trakhtenbrot L and Fixler D: Fluorescence lifetime imaging of
DAPI-stained nuclei as a novel diagnostic tool for the detection
and classification of B-cell chronic lymphocytic leukemia.
Cytometry A. 89:644–652. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mallavia B, Recio C, Oguiza A, Ortiz-Muñoz
G, Lazaro I, Lopez-Parra V, Lopez-Franco O, Schindler S, Depping R,
Egido J and Gomez-Guerrero C: Peptide inhibitor of NF-κB
translocation ameliorates experimental atherosclerosis. Am J
Pathol. 182:1910–1921. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wong MC, Zhang DX and Wang HH: Rapid
emergence of atherosclerosis in Asia: A systematic review of
coronary atherosclerotic heart disease epidemiology and
implications for prevention and control strategies. Curr Opin
Lipidol. 26:257–269. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Singh TP, Vangaveti VN and Malabu UH:
Dipeptidyl peptidase-4 inhibitors and their potential role in the
management of atherosclerosis-A review. Diabetes Metab Syndr.
9:223–229. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu GC, Leng RX, Lu Q, Fan YG, Wang DG and
Ye DQ: Subclinical atherosclerosis in patients with inflammatory
bowel diseases: A systematic review and meta-analysis. Angiology.
68:447–461. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cuspidi C, Sala C, Tadic M, Gherbesi E,
Grassi G and Mancia G: Nondipping pattern and carotid
atherosclerosis: A systematic review and meta-analysis. J
Hypertens. 34:385–392. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chartoumpekis DV, Habeos IG, Ziros PG,
Psyrogiannis AI, Kyriazopoulou VE and Papavassiliou AG: Brown
adipose tissue responds to cold and adrenergic stimulation by
induction of FGF21. Mol Med. 17:736–740. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Charoenphandhu N, Suntornsaratoon P,
Krishnamra N, Sa-Nguanmoo P, Tanajak P, Wang X, Liang G, Li X,
Jiang C, Chattipakorn N and Chattipakorn S: Fibroblast growth
factor-21 restores insulin sensitivity but induces aberrant bone
microstructure in obese insulin-resistant rats. J Bone Miner Metab.
35:142–149. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fan H, Sun X, Zhang H, Liu J, Zhang P, Xu
Y, Pan Q and Wang G: Effect of metformin on fibroblast growth
factor-21 levels in patients with newly diagnosed type 2 diabetes.
Diabetes Technol Ther. 18:120–126. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang G, Liu J, Yang N, Hu Y, Zhang H, Miao
L, Yao Z and Xu Y: Levothyroxine treatment restored the decreased
circulating fibroblast growth factor 21 levels in patients with
hypothyroidism. Eur J Intern Med. 31:94–98. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen Y, Zhao H and Ren X: Estrogen and
progestogen inhibit NF-κB in atherosclerotic tissues of
ovariectomized ApoE (−/-) mice. Climacteric. 19:357–363. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hsueh TP, Sheen JM, Pang JH, Bi KW, Huang
CC, Wu HT and Huang ST: The anti-atherosclerotic effect of naringin
is associated with reduced expressions of cell adhesion molecules
and chemokines through NF-κB Pathway. Molecules. 21:pii: E195.
2016. View Article : Google Scholar
|
|
39
|
Qiu L, Xu R, Wang S, Li S, Sheng H, Wu J
and Qu Y: Honokiol ameliorates endothelial dysfunction through
suppression of PTX3 expression, a key mediator of
IKK/IkappaB/NF-κB, in atherosclerotic cell model. Exp Mol Med.
47:e1712015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kim KM, Choi JY, Yoo SE, Park MY, Lee BS,
Ko YH, Sung SH, Shin HM and Park JE: HMCO5, herbal extract,
inhibits NF-kappaB expression in lipopolysaccharide treated
macrophages and reduces atherosclerotic lesions in cholesterol fed
mice. J Ethnopharmacol. 114:316–324. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Asare Y, Shagdarsuren E, Schmid JA,
Tilstam PV, Grommes J, El Bounkari O, Schütz AK, Weber C, de
Winther MP, Noels H and Bernhagen J: Endothelial CSN5 impairs NF-κB
activation and monocyte adhesion to endothelial cells and is highly
expressed in human atherosclerotic lesions. Thromb Haemost.
110:141–152. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sun R, Xiao L and Duan S: High expression
of ubiquitin conjugates and NF-κB in unstable human intracranial
atherosclerotic plaques. J Cell Physiol. 227:784–788. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sigala F, Savvari P, Liontos M, Sigalas P,
Pateras IS, Papalampros A, Basdra EK, Kolettas E, Papavassiliou AG
and Gorgoulis VG: Increased expression of bFGF is associated with
carotid atherosclerotic plaques instability engaging the NF-κB
pathway. J Cell Mol Med. 14:2273–2280. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Pan S, Lei L, Chen S, Li H and Yan F:
Rosiglitazone impedes Porphyromonas gingivalis-accelerated
atherosclerosis by downregulating the TLR/NF-κB signaling pathway
in atherosclerotic mice. Int Immunopharmacol. 23:701–708. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ye X, Jiang X, Guo W, Clark K and Gao Z:
Overexpression of NF-κB p65 in macrophages ameliorates
atherosclerosis in apoE-knockout mice. Am J Physiol Endocrinol
Metab. 305:E1375–E1383. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lazaro I, Oguiza A, Recio C, Mallavia B,
Madrigal-Matute J, Blanco J, Egido J, Martin-Ventura JL and
Gomez-Guerrero C: Targeting HSP90 ameliorates nephropathy and
atherosclerosis through suppression of NF-κB and STAT signaling
pathways in diabetic mice. Diabetes. 64:3600–3613. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bhat OM, Kumar PU, Giridharan NV, Kaul D,
Kumar MJ and Dhawan V: Interleukin-18-induced atherosclerosis
involves CD36 and NF-κB crosstalk in Apo E−/− mice. J Cardiol.
66:28–35. 2015. View Article : Google Scholar : PubMed/NCBI
|