Introduction
Glioblastoma (GBM; WHO grade IV) is the most common
and lethal primary malignant intracranial tumor (1). The invasion and migration of GBM
cells remain a significant cause of palindromia and mortality and
pose challenges for locally directed therapies such as surgery and
radiation therapy (2). Inhibiting
the invasion and migration of GBM cells has become an attractive
treatment strategy. Recent studies have demonstrated that
individual cells of certain cancers, such as GBM, have two
different interconvertible patterns of movement: A mesenchymal
pattern driven by Rac GTPase activation and an amoeboid pattern
driven by Rho GTPase activation (3,4).
Because the ability of GBM cells to switch between different
patterns of movement might limit the effectiveness of single
therapeutic agents targeting the migration and invasion of cancer
cells, simultaneously inhibiting Rac signaling and Rho signaling
may be an effective strategy for GBM therapy (5).
Every cell in the body, including cancer cells, is
surrounded by an intricate microenvironment. The in vivo
microenvironment provides physical support and strongly influences
the morphology, migration, proliferation and function of the cells
therein (4), but most publications
on cancer cells is based on the traditional two-dimensional (2D)
culture. This discrepancy may be an important reason for the
failure of the translation of numerous seemingly successful
preclinical studies into clinical applications. In recent years, an
increasing number of cell biology studies have been dedicated to
three-dimensional (3D) cultures in which the cells are fully
surrounded by matrix material and nutrition, in contrast to 2D
culture, where the cells grow on top of a stiff surface. The
biodegradability and porosity of 3D hydrogel may enable cancer
cells to move in both mesenchymal and amoeboid patterns (6).
In this study, we established the 3D hydrogel model
in vitro to investigate GBM cell motility inside the tumor
architecture. 3D hydrogel showed more advantages on studying
mesenchymal-amoeboid transition (MAT) and amoeboid-mesenchymal
transition (AMT) than the 2D monolayer culture. Simultaneous
treatment of NSC23766 [specific Ras-related C3 botulinum toxin
substrate 1 (Rac1) inhibitor] and Y27632 (selective ROCK1
inhibitor) abrogated U87 GBM cells migration through inhibiting
both MAT and AMT. Additionally, Y27632 induced integrin expression
which gave rise to the focal adhesion to facilitate the mesenchymal
invasion.
Materials and methods
Ethical approval
All procedures performed in studies involving
human participants were in accordance with the ethical standards of
the Institutional and/or National Research Committee and with the
1964 Helsinki Declaration and its later amendments or comparable
ethical standards. Informed consent was obtained from all
individual participants included in the study.
Cells, antibodies and reagents
Glioma cell line U87 was purchased from the Chinese
Academy of Sciences Cell Bank (Shanghai, China). Dulbecco's
Modified Eagle's Medium (DMEM) and fetal bovine serum (FBS) were
purchased from Gibco (Grand Island, NY, USA). Lipofectamine-3000™
and Alexa Fluor 488-labeled Goat Anti-Mouse IgG were purchased from
Invitrogen (Carlsbad, CA, USA). The antibiotic G418 was purchased
from Merck KGaA (Darmstadt, Germany). Plasmid pCMV-LifeAct-TagGFP2
and µ-Slide Chemotaxis3D were purchased from ibidi GmbH
(Martinsried, Germany). The specific inhibitors of NSC23766 and
Y27632 were purchased from Selleck Chemicals (Houston TX, USA).
Recombinant SDF-1was purchased from R&D Systems (Minneapolis,
MN, USA). 3D life dextran-CD hydrogel was purchased from CELLENDS
(Germany). Mouse anti-integrin αVβ3 monoclonal antibody was
purchased from Abcam (Cambridge, MA, USA).
Cell culture and plasmid
transfection
U87 cells were cultured in DMEM medium containing
10% FBS at 37°C under a humidified atmosphere with 5%
CO2. Lipofectin-3000™ was used for the transfection of
the Plasmid pCMV-LifeAct-TagGFP2 for 6 h. The entire process was
performed according to the manufacturer's instructions. The cells
were cultured with 800 µg/ml G418, and the result of transfection
was examined using an Olympus IX71 fluorescence microscope (Tianjin
Neurological Institute, Tianjin, China).
Targeted treatment and group
assignments
U87 cells were separated into 4 groups: The NSC23766
(Rac1 targeted inhibitor, 100 µM)-treated group; the Y-27632 (ROCK
targeted inhibitor, 10 µM)-treated group; the combined treatment
group with these two inhibitors; and the control group.
Wound healing cell migration
assays
U87 cells were seeded in 6-well dishes
(2×105 cells/well). On the next day, an artificial wound
was made with a sterile pipette tip. Targeted inhibitors were then
added as assigned to each group. The cells were then incubated at
37°C with 5% CO2. Images of the wound area 24 h after
injury were captured using an inverted microscope.
3D cultures in hydrogel and the
chemotaxis experiment
To prepare the hydrogel, the polymers, pH buffer,
water, peptides, crosslinker and cell suspension with final a
concentration of 106/ml were mixed as described in the
manufacturer's instructions. The final concentration of
themaleimide and thiol groups reached 2.5 mmol/l, thus making the
gel strength similar to brain tissue.
To observe the transformation of cell morphology in
the 3D hydrogel, the cells were treated with targeted inhibitors
according to their group assignments, and images were collected at
40-sec intervals at 37°C with 5% CO2 using laser
confocal microscopy (PerkinElmer, Inc., Waltham, MA, USA).
Cells in the hydrogel system prepared as described
above were pretreated with the targeted inhibitors for 3 h.
Chemotaxis experiments have been done once as previously described
(7,8). The hydrogel system was injected into
the observation channel of µ-Slide Chemotaxis3D, the
complete medium containing SDF-1 (100 nmol/ml) was injected into
the left reservoir, and the complete medium without SDF-1 was
injected into the right reservoir. For time-lapse microscopic
analyses, cells were cultured at 37°C with 5% CO2 for 12
h. Images were collected at 2-min intervals with a living cells
workstation (Nikon Ti-E; Nikon Corp., Tokyo, Japan).
Immunofluorescence staining
The integrin expression in the cells that had been
observed for morphology translation was evaluated by
immunofluorescence staining. Cells were fixed with 4%
paraformaldehyde for 15 min, permeabilized with 0.5% Triton X-100
for 20 min, and blocked with 0.5% BSA in PBS for 1 h.
Immunofluorescence staining was performed by incubating the
hydrogel system with a mouse anti-integrin αVβ3 monoclonal antibody
(1:50) overnight at 4°C and with Alexa Fluor 488-labeled goat
anti-mouse secondary antibody (1:1,000) for 2 h on the next day.
Immunofluorescence was visualized, and the images were captured
with an Olympus IX71 fluorescence microscope (Tianjin Neurological
Institute).
Patient data analysis
Patient data and gene expression datasets were
obtained using the R2 microarray analysis and visualization
platform (http://hgserver1.amc.nl/cgi-bin/r2/main.cgi). All
prognosis analyses were conducted online, and all data and P values
(log-rank test) were downloaded. Kaplan-Meier analysis and the
resulting survival curves were performed using GraphPad Prism
(version 6.0; GraphPad Software, Inc., La Jolla, CA, USA). All
cutoff values for separating high and low expression groups were
determined by the online R2 database algorithm. Medians were used
for separating high and low expression groups online.
Statistical analysis
All quantified data represent an average of at least
triplicate experiments unless otherwise indicated, and standard
deviations were calculated. All statistical analyses were performed
using GraphPad Prism 6.0. Comparisons among groups were determined
by multiple comparisons of ordinary one-way ANOVA, where P<0.05
was significant.
Results
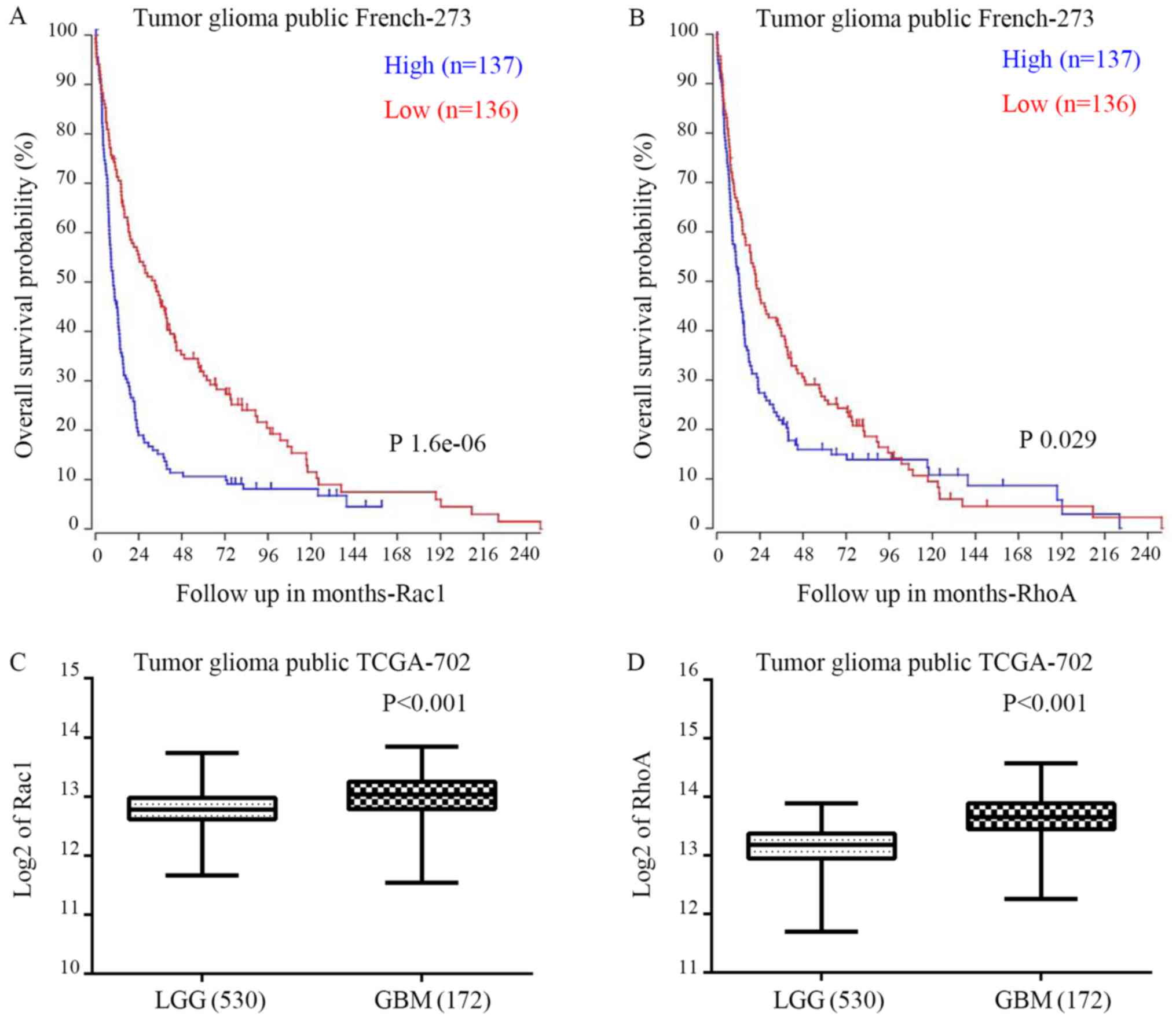
Rac1 and Ras homolog family member A
(RhoA) were closely related to the prognosis of patients
We evaluated the correlation between Rac1/RhoA and
overall survival (OS) through R2 genomics analysis and the
visualization platform database. Medians were used for separating
high and low expression groups by the online R2 database algorithm.
Rac1 was highly expressed in 137 of the 273 cases of glioma. Rac1
expression was negatively correlated with the patient OS in
French's data (P=1.6e-0.6; Fig.
1A). Similarly, RhoA was upregulated in 137 cases among the 273
patients. RhoA also showed negative correlation with the patient OS
(P=0.029; Fig. 1B). Accordingly,
in contrast to lower grade gliomas (LGG), both Rac1 and RhoA were
significantly upregulated in GBM patients according to TCGA data
(Fig. 1C and D).
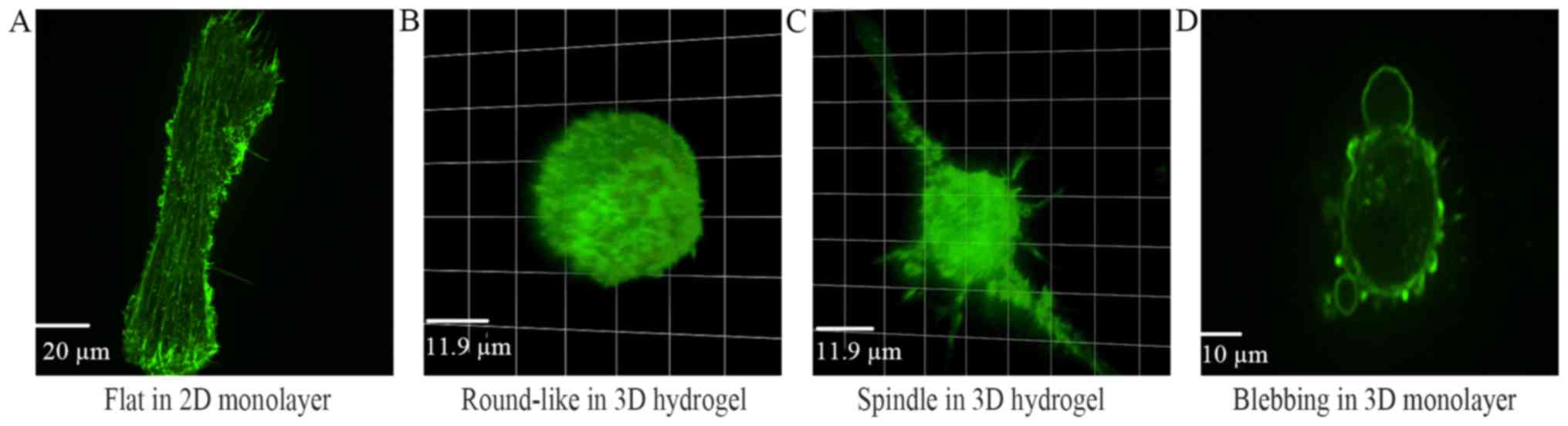
The morphology of cells in 2D
monolayer and 3D hydrogel
To investigate cell morphology in detail, U87 GBM
cells were transfected with plasmid pCMV-LifeAct-TagGFP2 and the
morphology was observed with a confocal microscopy. In the 2D
monolayer culture, U87 cells presented as flat and protruded
several broad lamellipodium without polarity (Fig. 2A). In the 3D hydrogel culture, the
cells exhibited as spindle or round-like (Fig. 2B-D). Most spindle cells contained
one or two spindly protrusion and harbored obvious polarity. Blebs
around the round-like cells were observed (Fig. 2D).
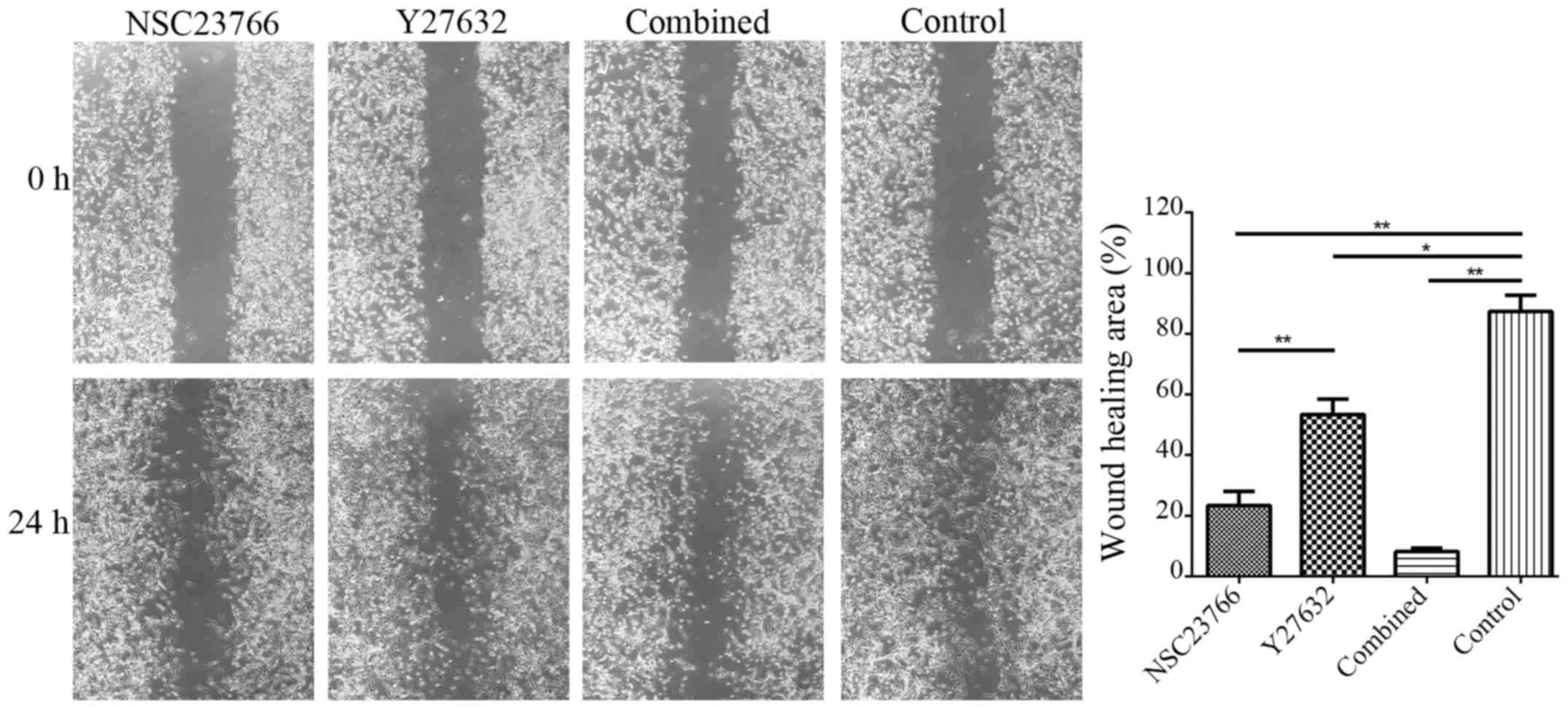
Migration-targeted inhibitions in the
2D monolayer
Considering the mesenchymal pattern driven by Rac
GTPase activation and the amoeboid pattern driven by Rho GTPase
activation, NSC23766 (specific Rac1 inhibitor) and Y27632
(selective ROCK1 inhibitor) were introduced to treat U87 glioma
cells in the traditional 2D monolayer culture. The migration of U87
cells to the wound area was analyzed at 24 h after injury and
treatment. As a result, the wound healed 23.35±8.10% in the
NSC23766-treated group, 53.38±8.80% in the Y27632-treated group,
8.18±1.93% in the combined treatment group, and 87.38±9.30% in the
control group. Cells in the combined treatment group migrated
significantly more slowly than did the control group (P<0.01;
Fig. 3). This finding revealed
that inhibiting both Rac1 and RhoA abolished the wound healing of
U87 GBM cells.
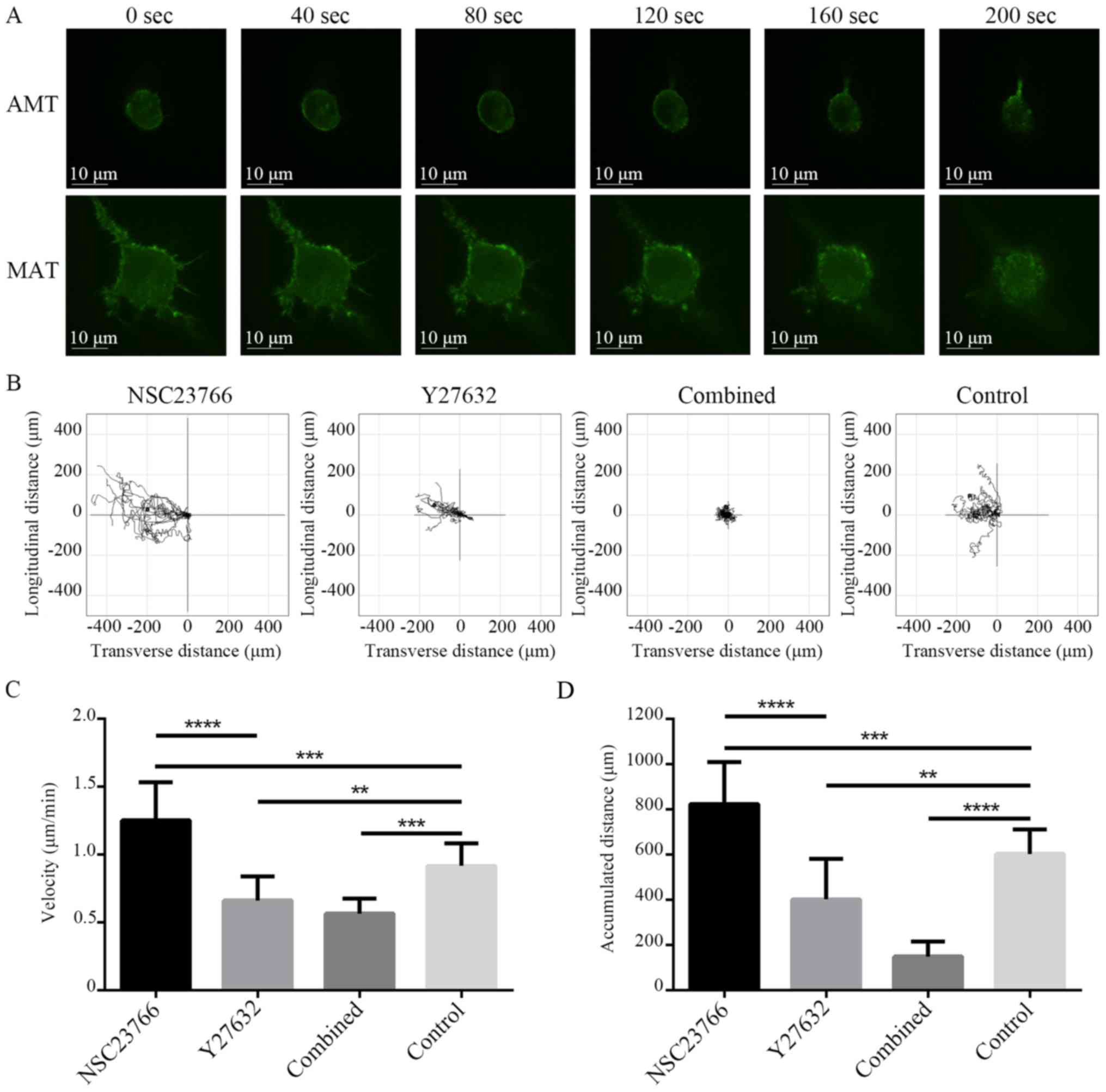
Targeted inhibited invasion and
migration in the 3D hydrogel
Because the tumor architecture strongly influences
the morphology, invasion, and migration of cells (9), the efficiency of targeted inhibition
was investigated in 3D hydrogel. U87 cells were treated with
targeted inhibitors according to their assigned groups, and the
morphology of these cells was observed with a confocal microscope
at 40-sec intervals (Fig. 4). The
morphology of the cells underwent MAT transformed gradually from
spindle- to round-like, and the protrusion disappeared (Fig. 4B). In contrast, the morphology of
the cells underwent AMT transformed gradually from round-like to
spindle-shaped, and the protrusion extended gradually (Fig. 4A).
Single cell migration towards SDF-1 was observed by
living cells workstation. U87 cells were pretreated with targeted
inhibitors for 3 h. Under the chemotaxis of SDF-1 for 12 h, the
invasion and migration of U87 cells were assessed by live cell
tracking in µ-slide chemotaxis chambers as described in the
materials and methods section. Images were collected at 2-min
intervals with time-lapse microscopy, and all cell trajectories
were recorded using the ImageJ software (Fig. 4A). In addition, movement velocity
and distance were calculated. The movement velocity and 12-h
movement distance of the different treatment groups were
1.253±0.282 µm/min, 0.663±0.178, 0.567±0.110 and 0.917±0.166 µm/min
(Fig. 4C) and 824.645±184.608,
402.703±179.196, 149.149±63.765 and 603.085±109.441 µm (Fig. 4D), respectively. The differences
were significant between the experimental groups and the control
group (P<0.05 for each). Interestingly, NSC23766 treated cells
moved much faster than were cells in other groups in the 3D
hydrogel (P<0.05; Fig. 4C and
D). However, NSC23766 treated cells moved slower than other
group cells in the 2D monolayer (P<0.05; Fig. 3).
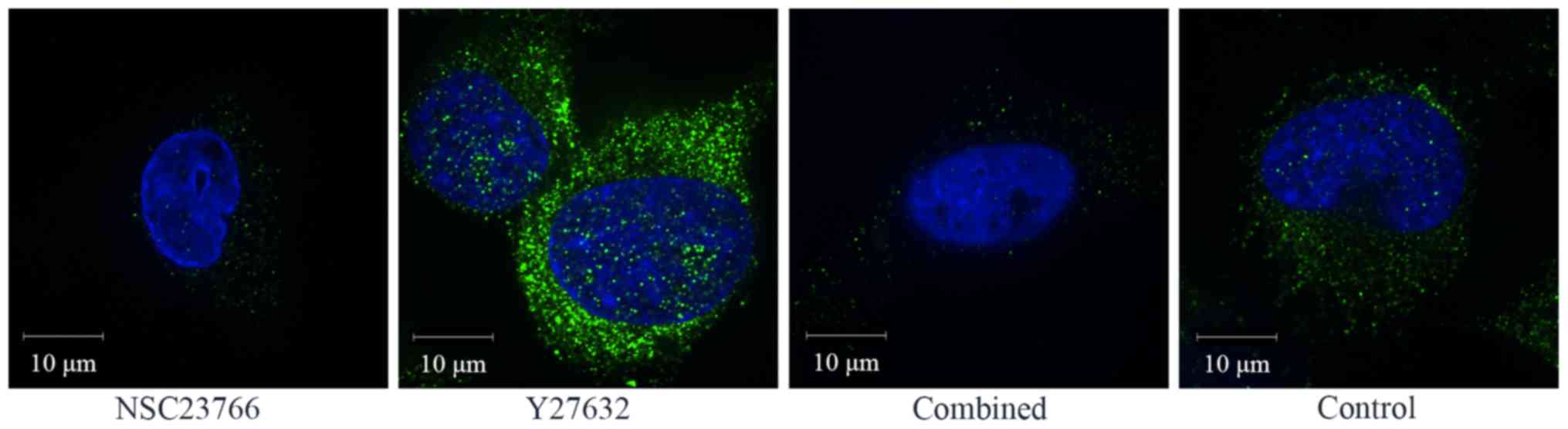
Integrin expression in targeted
inhibited cells
The formation of focal adhesion mediated by integrin
is an important process during the invasion of glioma cells.
Integrin αVβ3 expression was detected by immunofluorescence
staining to assess the adhesion of U87 cells. The integrin αVβ3
expression of the Y27632-treated group was much higher than in the
other groups. The integrin αVβ3 expression of the Y27632-treated
group and the combined treatment group was much lower than in the
control group (Fig. 5).
Discussion
GBM is the most common intracranial malignant tumor,
and it has a poor prognosis anda powerful capacity for invasion and
migration (2,10). GBM cells have two different
interconvertible patterns of movement: The mesenchymal pattern
driven by Rac GTPase activation and the amoeboid pattern driven by
Rho GTPase activation (3,4,11).
Mesenchymal pattern is the main motility model in the brain white
matter with a slower speed (0.1–1 µm/min) (12). The tumor cells with mesenchymal
motility pattern presented the spindle morphology and the invasion
dependent on the enzymatic hydrolysis of the extracellular matrix
(ECM) (13). Rac1 promotes the
polymerization from G-actins to F-actins and serves as the key
molecule of mesenchymal movement (14,15).
Amoeboid pattern is the main motility model in subpial,
subependymal and extravascular space with a faster speed (a maximum
of 4.0 µm/min can be achieved) (12). The cells with amoeboid motility
pattern exhibited a round-like morphology and the migration
dependent on the actomyosin contraction driven by RhoA rather than
the polarization driven by Rac1 (12,16).
The induction of actomyosin contractility via RhoA-ROCK signaling
leads to blebbing, thus causing amoeboid migration, but it also
regulates tail retraction in mesenchymal migration (2,16,17).
In this study, single-locus analysis revealed that patients with a
relatively higher Rac1/RhoA level harbored shorter OS (Fig. 1A). Additionally, the Rac1/RhoA
level was increased in GBM compared with the lower-grade glioma
(Fig. 1B).
Consistent with Stephen Paget's 128-year-old
‘seed-and-soil’ hypothesis, the microenvironment in vivo
provides a physical support and strongly influences the morphology,
migration, proliferation and function of cells therein (9), whereas most studies of cancer cells
are currently based on the traditional 2D culture. Cell properties
including extending vesicle, invasion, mesenchymal and amoeboid
movement are unable to be thoroughly studied in the 2D monolayer.
Despite the advantages of animal models on studying tumorigenesis
and progression, it is difficult to detect aimed molecules and
trace the cell motion in vivo (18). Even the organotypic brain slice
cannot be used for the morphological and tracing research of a
single cell (19). Cells in 3D
cultures mimicking the microenvironment in vivo are fully
surrounded by matrix material, in contrast to 2D culture, where
cells are grown on top of a stiff surface. The biodegradability and
porosity of the 3D hydrogel enable cancer cells to move in both
mesenchymal and amoeboid patterns (20,21).
In this study, 3D life dextran-CD hydrogelwas utilized for U87 GBM
cell culture. Because cell morphology and motor pattern can be
affected by the stiffness of the environment (22), the final concentrations of
maleimide and thiol groups were made up to 2.5 mmol/l; thus, the
gel strength is similar to brain tissue. The results suggested that
U87 cells had two diverse patterns, i.e., spindle- and round-like
(Fig. 2B-D), in 3D hydrogel,
whereas they presented a flat pattern in the 2D monolayer (Fig. 2A).
Because the conversion between different patterns of
movement might limit the efficiency of single therapeutic agents,
combined therapy targeting Rac1 and RhoA would be a promising
strategy to restrain the invasion and migration of GBM cells
(23,24). Rac1-WAVE signaling has been shown
to promote cytoskeletal reorganization and invadopodium formation
(17). ROCK is a downstream
molecular target of RhoA, which induces actomyosin contraction and
causes amoeboid migration. Thus, Rho activation promotes cell
contraction and hinders mesenchymal movement by modulating Rac-GAP
ARHGAP22 signaling (12,16). NSC23766 is a specific inhibitor of
the binding and activation of Rac1 GTPase and does not inhibit the
closely related targets, Cdc42 or RhoA. In this study, NSC23766 was
used to inhibit Rac1 signaling. Because RhoA contributes to both
amoeboid and mesenchymal modes of migration (25), Y27632 (selective ROCK1 inhibitor)
was selected to inhibit RhoA-ROCK signaling.
In the 2D monolayer, the cells in the
NSC23766-treated group migrated more slowly than did the cells in
both the control group and the Y27632-treated group in the wound
healing assay (P<0.05 for each, Fig. 3). Interestingly, the cells of the
NSC23766 treated group invaded and migrated faster than the cells
of the Y27632 treated group and the control group in 3D hydrogel
(P<0.05, P<0.05; Fig. 4D).
This result is consistent with the phenomenon that some tumor cells
migrate in an amoeboid pattern with a higher speed in vivo
(12). Otherwise, it probably
revealed the reason that numerous seemingly successful preclinical
studies based on the 2D model failed in translating into clinical
applications. At the same time, MAT was observed in the NSC23766
treated group and AMT was observed in the Y27632 treated group in
3D hydrogel (Fig. 4A), both of
which cannot be seen in the 2D monolayer. Additionally, the results
of combined inhibition targeting Rac1 and RhoA indicated that both
the movement velocity and 12-h movement distance were significantly
decreased (Fig. 4C and D).
In fact, the average movement velocity of U87 cells
treated by NSC23766 was not the same as the tumor cells moving in
an amoeboid pattern with the maximum of 4.0 µm/min. Several facts
might contribute to this difference. Firstly, 1.253±0.282 µm/min
was the general speed of 15 cells, and not all of the cells were
moving in an amoeboid pattern. Secondly, the stiffness of the ECM
influences the invasion and migration (26). The soft hydrogel lacks stiffness
and allows the glioma cells to move quickly in an amoeboid pattern.
Last but not least, the porosity of the hydrogel is an important
environmental factor for cell migration. Cell migration is fastest
at pore diameters that match or are slightly smaller than the
cells; migration speed decreases in large pore size matrices due to
the loss of cell-matrix interactions, but pore sizes much smaller
than the cell diameter trap cells in a physical cage and reduce
cell migration (27). In this
hydrogel, the average diameter was slightly smaller than the cell
size with a final concentration of 2.5 mmol/l of maleimide and
thiol groups, and it is difficult to avoid the fluctuation of
diameter within a certain range.
Integrin αVβ3 was identified as a driver of an
aggressive and metastatic tumor phenotype (28,29).
Here, the integrin αVβ3 expression of targeted inhibited U87 cells
was detected by immunofluorescence staining to assess the adhesion.
The results showed that the integrin αVβ3 expression of the
Y27632-treated group was much higher than those of the other
groups. The integrin αVβ3 expressions of the NSC23766-treated group
and combined treatment group were much lower than that of control
group (Fig. 5). This result
verified the fact that Rac signaling activates inversely with Rho
signaling.
In conclusion, we mainly investigated the invasion
and migration of U87 glioma cells and targeted therapy in 3D
hydrogel. The results showed that the U87 cells treated with
NSC23766 moved much faster than did the control cells in the 3D
chemotactic assay, whereas they moved much more slowly than did the
control cells in the 2D wound healing assay. This result probably
revealed the reason why numerous seemingly successful preclinical
studies based on 2D model have failed to be translated into
clinical applications. The effective inhibition of invasion and
migration by the combined targeting Rac1 and RhoA suggested that
combined targeted therapy may be a promising strategy for
preventing invasion and migration of GBM cells.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (nos. 81272782 and 81472352); and the
Tianjin Research Program of Application Foundation and Advanced
Technology (no. 15JCZDJC36200).
References
|
1
|
Stupp R, Hegi ME, Mason WP, van den Bent
MJ, Taphoorn MJ, Janzer RC, Ludwin SK, Allgeier A, Fisher B,
Belanger K, et al: Effects of radiotherapy with concomitant and
adjuvant temozolomide versus radiotherapy alone on survival in
glioblastoma in a randomised phase III study: 5-year analysis of
the EORTC-NCIC trial. Lancet Oncol. 10:459–466. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Friedl P and Wolf K: Tumour-cell invasion
and migration: Diversity and escape mechanisms. Nat Rev Cancer.
3:362–374. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sanz-Moreno V, Gadea G, Ahn J, Paterson H,
Marra P, Pinner S, Sahai E and Marshall CJ: Rac activation and
inactivation control plasticity of tumor cell movement. Cell.
135:510–523. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rao SS, Bentil S, DeJesus J, Larison J,
Hissong A, Dupaix R, Sarkar A and Winter JO: Inherent interfacial
mechanical gradients in 3D hydrogels influence tumor cell
behaviors. PloS One. 7:e358522012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Parri M and Chiarugi P: Rac and Rho
GTPases in cancer cell motility control. Cell Commun Signal.
8:232010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jiglaire Jiguet C, Baeza-Kallee N,
Denicolaï E, Barets D, Metellus P, Padovani L, Chinot O,
Figarella-Branger D and Fernandez C: Ex vivo cultures of
glioblastoma in three-dimensional hydrogel maintain the original
tumor growth behavior and are suitable for preclinical drug and
radiation sensitivity screening. Exp Cell Res. 321:99–108. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zengel P, Nguyen-Hoang A, Schildhammer C,
Zantl R, Kahl V and Horn E: µ-Slide chemotaxis: A new chamber for
long-term chemotaxis studies. BMC Cell Biol. 12:212011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gouwy M, De Buck M, Pörtner N, Opdenakker
G, Proost P, Struyf S and Van Damme J: Serum amyloid A
chemoattracts immature dendritic cells and indirectly provokes
monocyte chemotaxis by induction of cooperating CC and CXC
chemokines. Eur J Immunol. 45:101–112. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Thiele J, Ma Y, Bruekers SM, Ma S and Huck
WT: 25th anniversary article: Designer hydrogels for cell cultures:
A materials selection guide. Adv Mater. 26:125–147. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lu KV, Chang JP, Parachoniak CA, Pandika
MM, Aghi MK, Meyronet D, Isachenko N, Fouse SD, Phillips JJ,
Cheresh DA, et al: VEGF inhibits tumor cell invasion and
mesenchymal transition through a MET/VEGFR2 complex. Cancer Cell.
22:21–35. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mikheeva SA, Mikheev AM, Petit A, Beyer R,
Oxford RG, Khorasani L, Maxwell JP, Glackin CA, Wakimoto H,
González-Herrero I, et al: TWIST1 promotes invasion through
mesenchymal change in human glioblastoma. Mol Cancer. 9:1942010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Symons M and Segall JE: Rac and Rho
driving tumor invasion: Who's at the wheel? Genome Biol.
10:2132009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yamazaki D, Kurisu S and Takenawa T:
Involvement of Rac and Rho signaling in cancer cell motility in 3D
substrates. Oncogene. 28:1570–1583. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhong J, Paul A, Kellie SJ and O'Neill GM:
Mesenchymal migration as a therapeutic target in glioblastoma. J
Oncol. 2010:4301422010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wuichet K and Søgaard-Andersen L:
Evolution and diversity of the Ras superfamily of small GTPases in
prokaryotes. Genome Biol Evol. 7:57–70. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Haga RB and Ridley AJ: Rho GTPases:
Regulation and roles in cancer cell biology. Small GTPases.
7:207–221. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Murali A and Rajalingam K: Small Rho
GTPases in the control of cell shape and mobility. Cell Mol Life
Sci. 71:1703–1721. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rape AD and Kumar S: A composite hydrogel
platform for the dissection of tumor cell migration at tissue
interfaces. Biomaterials. 35:8846–8853. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ren B, Yu S, Chen C, Wang L, Liu Z, Wu Q,
Wang L, Zhao K and Yang X: Invasion and anti-invasion research of
glioma cells in an improved model of organotypic brain slice
culture. Tumori. 101:390–397. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Flemming A: Cancer: Multifunctional
nanodevice reverses drug resistance. Nat Rev Drug Discov.
14:3092015. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shea LD, Woodruff TK and Shikanov A:
Bioengineering the ovarian follicle microenvironment. Annu Rev
Biomed Eng. 16:29–52. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Souza GR, Molina JR, Raphael RM, Ozawa MG,
Stark DJ, Levin CS, Bronk LF, Ananta JS, Mandelin J, Georgescu MM,
et al: Three-dimensional tissue culture based on magnetic cell
levitation. Nat Nanotechnol. 5:291–296. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Reymond N, Im JH, Garg R, Vega FM, d'Agua
Borda B, Riou P, Cox S, Valderrama F, Muschel RJ and Ridley AJ:
Cdc42 promotes transendothelial migration of cancer cells through
β1 integrin. J Cell Biol. 199:653–668. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Navarro-Lérida I, Pellinen T, Sanchez SA,
Guadamillas MC, Wang Y, Mirtti T, Calvo E and Del Pozo MA: Rac1
nucleocytoplasmic shuttling drives nuclear shape changes and tumor
invasion. Dev Cell. 32:318–334. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sadok A, McCarthy A, Caldwell J, Collins
I, Garrett MD, Yeo M, Hooper S, Sahai E, Kuemper S, Mardakheh FK
and Marshall CJ: Rho kinase inhibitors block melanoma cell
migration and inhibit metastasis. Cancer Res. 75:2272–2284. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mishima T, Naotsuka M, Horita Y, Sato M,
Ohashi K and Mizuno K: LIM-kinase is critical for the
mesenchymal-to-amoeboid cell morphological transition in 3D
matrices: Biochem Biophys Res Commun. 392:577–581. 2010.
|
|
27
|
Panková K, Rösel D, Novotný M and Brábek
J: The molecular mechanisms of transition between mesenchymal and
amoeboid invasiveness in tumor cells. Cell Mol Life Sci. 67:63–71.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Desgrosellier JS, Barnes LA, Shields DJ,
Huang M, Lau SK, Prévost N, Tarin D, Shattil SJ and Cheresh DA: An
integrin alpha(v)beta(3)-c-Src oncogenic unit promotes
anchorage-independence and tumor progression. Nat Med.
15:1163–1169. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Seguin L, Kato S, Franovic A, Camargo MF,
Lesperance J, Elliott KC, Yebra M, Mielgo A, Lowy AM, Husain H, et
al: An integrin β3-KRAS-RalB complex drives tumour
stemness and resistance to EGFR inhibition. Nat Cell Biol.
16:457–468. 2014. View Article : Google Scholar : PubMed/NCBI
|